
Impact of Melatonin on RAW264.7 Macrophages during Mechanical Strain

 ,
,  and
and
Abstract
:1. Introduction
2. Results
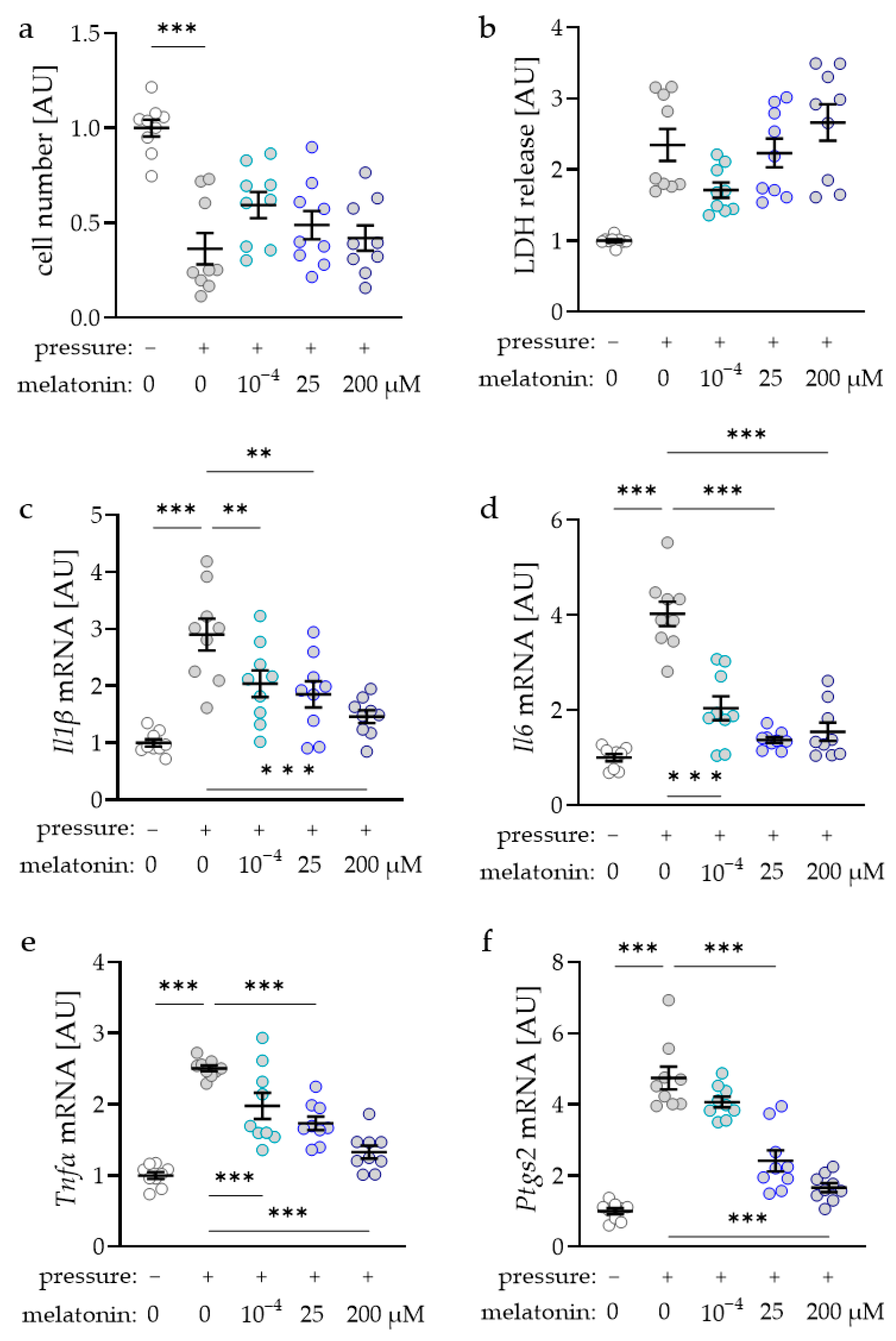
2.1. Determination of the Concentration Dependence of Melatonin on Macrophages during Mechanical Strain
2.2. Impact of Melatonin on Expression of Inflammatory Genes during Tensile Strain
2.3. Impact of Melatonin on Expression of Inflammatory Genes and Proteins during Compressive Strain
3. Discussion
4. Materials and Methods
4.1. Cell Culture Experiments
4.2. Determination of Cell Number
4.3. Lactate Dehydrogenase (LDH) Assay
4.4. RNA Isolation
4.5. cDNA Synthesis
4.6. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.7. Western Blot Analysis
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Statistical Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lombardo, G.; Vena, F.; Negri, P.; Pagano, S.; Barilotti, C.; Paglia, L.; Colombo, S.; Orso, M.; Cianetti, S. Worldwide prevalence of malocclusion in the different stages of dentition: A systematic review and meta-analysis. Eur. J. Paediatr. Dent. 2020, 21, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, S.S.; Gheware, A.; Kamatagi, L.; Pasumarthy, S.; Pawar, V.; Fatangare, M. Dental caries and its relationship to malocclusion in permanent dentition among 12–15 year old school going children. J. Int. Oral Health 2014, 6, 27–30. [Google Scholar] [PubMed]
- Feldens, C.A.; Dos Santos Dullius, A.I.; Kramer, P.F.; Scapini, A.; Busato, A.L.S.; Vargas-Ferreira, F. Impact of malocclusion and dentofacial anomalies on the prevalence and severity of dental caries among adolescents. Angle Orthod. 2015, 85, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Staufer, K.; Landmesser, H. Effects of crowding in the lower anterior segment--a risk evaluation depending upon the degree of crowding. J. Orofac. Orthop. 2004, 65, 13–25. [Google Scholar] [CrossRef]
- Meikle, M.C. The tissue, cellular, and molecular regulation of orthodontic tooth movement: 100 years after Carl Sandstedt. Eur. J. Orthod. 2006, 28, 221–240. [Google Scholar] [CrossRef] [Green Version]
- Schröder, A.; Bauer, K.; Spanier, G.; Proff, P.; Wolf, M.; Kirschneck, C. Expression kinetics of human periodontal ligament fibroblasts in the early phases of orthodontic tooth movement. J. Orofac. Orthop. 2018, 79, 337–351. [Google Scholar] [CrossRef]
- Kanzaki, H.; Chiba, M.; Shimizu, Y.; Mitani, H. Periodontal ligament cells under mechanical stress induce osteoclastogenesis by receptor activator of nuclear factor kappaB ligand up-regulation via prostaglandin E2 synthesis. J. Bone Miner. Res. 2002, 17, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Schröder, A.; Käppler, P.; Nazet, U.; Jantsch, J.; Proff, P.; Cieplik, F.; Deschner, J.; Kirschneck, C. Effects of Compressive and Tensile Strain on Macrophages during Simulated Orthodontic Tooth Movement. Mediat. Inflamm. 2020, 2020, 2814015. [Google Scholar] [CrossRef]
- McArthur, A.J.; Hunt, A.E.; Gillette, M.U. Melatonin action and signal transduction in the rat suprachiasmatic circadian clock: Activation of protein kinase C at dusk and dawn. Endocrinology 1997, 138, 627–634. [Google Scholar] [CrossRef]
- Redman, J.; Armstrong, S.; Ng, K.T. Free-running activity rhythms in the rat: Entrainment by melatonin. Science 1983, 219, 1089–1091. [Google Scholar] [CrossRef]
- Lekic, P.; McCulloch, C.A.G. Periodontal ligament cell populations: The central role of fibroblasts in creating a unique tissue. Anat. Rec. 1996, 245, 327–341. [Google Scholar] [CrossRef]
- Tan, D.-X.; Manchester, L.C.; Esteban-Zubero, E.; Zhou, Z.; Reiter, R.J. Melatonin as a Potent and Inducible Endogenous Antioxidant: Synthesis and Metabolism. Molecules 2015, 20, 18886–18906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Chen, S.; Zeng, S.; Zhao, Y.; Zhu, C.; Deng, B.; Zhu, G.; Yin, Y.; Wang, W.; Hardeland, R.; et al. Melatonin in macrophage biology: Current understanding and future perspectives. J. Pineal Res. 2019, 66, e12547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowes, D.A.; Webster, N.R.; Murphy, M.P.; Galley, H.F. Antioxidants that protect mitochondria reduce interleukin-6 and oxidative stress, improve mitochondrial function, and reduce biochemical markers of organ dysfunction in a rat model of acute sepsis. Br. J. Anaesth. 2013, 110, 472–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell. Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Ekmekcioglu, C. Melatonin receptors in humans: Biological role and clinical relevance. Biomed. Pharmacother. 2006, 60, 97–108. [Google Scholar] [CrossRef]
- Jockers, R.; Delagrange, P.; Dubocovich, M.L.; Markus, R.P.; Renault, N.; Tosini, G.; Cecon, E.; Zlotos, D.P. Update on melatonin receptors: IUPHAR Review 20. Br. J. Pharmacol. 2016, 173, 2702–2725. [Google Scholar] [CrossRef] [Green Version]
- Lewy, A.J. The dim light melatonin onset, melatonin assays and biological rhythm research in humans. Biol. Signals Recept. 1999, 8, 79–83. [Google Scholar] [CrossRef]
- Voultsios, A.; Kennaway, D.J.; Dawson, D. Salivary melatonin as a circadian phase marker: Validation and comparison to plasma melatonin. J. Biol. Rhythms 1997, 12, 457–466. [Google Scholar] [CrossRef]
- Vakkuri, O. Diurnal rhythm of melatonin in human saliva. Acta Physiol. Scand. 1985, 124, 409–412. [Google Scholar] [CrossRef]
- Miles, A.; Philbrick, D.R.; Shaw, D.M.; Tidmarsh, S.F.; Pugh, A.J. Salivary melatonin estimation in clinical research. Clin. Chem. 1985, 31, 2041–2042. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Hidalgo, M.; de La Lastra, C.A.; Carrascosa-Salmoral, M.P.; Naranjo, M.C.; Gomez-Corvera, A.; Caballero, B.; Guerrero, J.M. Age-related changes in melatonin synthesis in rat extrapineal tissues. Exp. Gerontol. 2009, 44, 328–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvetnoy, I.M. Extrapineal melatonin: Location and role within diffuse neuroendocrine system. Histochem. J. 1999, 31, 1–12. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Semak, I.; Kim, T.-K.; Janjetovic, Z.; Slominski, R.M.; Zmijewski, J.W. Melatonin, mitochondria, and the skin. Cell. Mol. Life Sci. 2017, 74, 3913–3925. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Hardeland, R.; Zmijewski, M.A.; Slominski, R.M.; Reiter, R.J.; Paus, R. Melatonin: A Cutaneous Perspective on its Production, Metabolism, and Functions. J. Investig. Dermatol. 2018, 138, 490–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, J.A.; Kim, B.G.; Lin, W.L.; Cho, M.I. Melatonin promotes osteoblast differentiation and bone formation. J. Biol. Chem. 1999, 274, 22041–22047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakade, O.; Koyama, H.; Ariji, H.; Yajima, A.; Kaku, T. Melatonin stimulates proliferation and type I collagen synthesis in human bone cells in vitro. J. Pineal Res. 1999, 27, 106–110. [Google Scholar] [CrossRef]
- Radio, N.M.; Doctor, J.S.; Witt-Enderby, P.A. Melatonin enhances alkaline phosphatase activity in differentiating human adult mesenchymal stem cells grown in osteogenic medium via MT2 melatonin receptors and the MEK/ERK (1/2) signaling cascade. J. Pineal Res. 2006, 40, 332–342. [Google Scholar] [CrossRef]
- Satomura, K.; Tobiume, S.; Tokuyama, R.; Yamasaki, Y.; Kudoh, K.; Maeda, E.; Nagayama, M. Melatonin at pharmacological doses enhances human osteoblastic differentiation in vitro and promotes mouse cortical bone formation in vivo. J. Pineal Res. 2007, 42, 231–239. [Google Scholar] [CrossRef]
- Sethi, S.; Radio, N.M.; Kotlarczyk, M.P.; Chen, C.-T.; Wei, Y.-H.; Jockers, R.; Witt-Enderby, P.A. Determination of the minimal melatonin exposure required to induce osteoblast differentiation from human mesenchymal stem cells and these effects on downstream signaling pathways. J. Pineal Res. 2010, 49, 222–238. [Google Scholar] [CrossRef]
- Liu, J.; Huang, F.; He, H.-W. Melatonin effects on hard tissues: Bone and tooth. Int. J. Mol. Sci. 2013, 14, 10063–10074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, J.-H.; Cho, Y.-C.; Sung, I.-Y.; Kim, I.-R.; Park, B.-S.; Kim, Y.-D. Melatonin promotes osteoblast differentiation and mineralization of MC3T3-E1 cells under hypoxic conditions through activation of PKD/p38 pathways. J. Pineal Res. 2014, 57, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Koyama, H.; Nakade, O.; Takada, Y.; Kaku, T.; Lau, K.H.W. Melatonin at pharmacologic doses increases bone mass by suppressing resorption through down-regulation of the RANKL-mediated osteoclast formation and activation. J. Bone Miner. Res. 2002, 17, 1219–1229. [Google Scholar] [CrossRef]
- Kitaura, H.; Yoshimatsu, M.; Fujimura, Y.; Eguchi, T.; Kohara, H.; Yamaguchi, A.; Yoshida, N. An anti-c-Fms antibody inhibits orthodontic tooth movement. J. Dent. Res. 2008, 87, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Davidovitch, Z. (Eds.) Biological Mechanisms of Tooth Movement; 2., Auflage; John Wiley & Sons: New York, NY, USA, 2015; ISBN 1118688872. [Google Scholar]
- Deng, W.-G.; Tang, S.-T.; Tseng, H.-P.; Wu, K.K. Melatonin suppresses macrophage cyclooxygenase-2 and inducible nitric oxide synthase expression by inhibiting p52 acetylation and binding. Blood 2006, 108, 518–524. [Google Scholar] [CrossRef]
- Mayo, J.C.; Sainz, R.M.; Tan, D.-X.; Hardeland, R.; Leon, J.; Rodriguez, C.; Reiter, R.J. Anti-inflammatory actions of melatonin and its metabolites, N1-acetyl-N2-formyl-5-methoxykynuramine (AFMK) and N1-acetyl-5-methoxykynuramine (AMK), in macrophages. J. Neuroimmunol. 2005, 165, 139–149. [Google Scholar] [CrossRef]
- Gilad, E.; Wong, H.R.; Zingarelli, B.; Virág, L.; O’Connor, M.; Salzman, A.L.; Szabó, C. Melatonin inhibits expression of the inducible isoform of nitric oxide synthase in murine macrophages: Role of inhibition of NFkappaB activation. FASEB J. 1998, 12, 685–693. [Google Scholar] [CrossRef]
- Xia, M.-Z.; Liang, Y.-L.; Wang, H.; Chen, X.; Huang, Y.-Y.; Zhang, Z.-H.; Chen, Y.-H.; Zhang, C.; Zhao, M.; Xu, D.-X.; et al. Melatonin modulates TLR4-mediated inflammatory genes through MyD88- and TRIF-dependent signaling pathways in lipopolysaccharide-stimulated RAW264.7 cells. J. Pineal Res. 2012, 53, 325–334. [Google Scholar] [CrossRef]
- Lissoni, P.; Barni, S.; Tancini, G.; Brivio, F.; Tisi, E.; Zubelewicz, B.; Braczkowski, R. Role of the pineal gland in the control of macrophage functions and its possible implication in cancer: A study of interactions between tumor necrosis factor-alpha and the pineal hormone melatonin. J. Biol. Regul. Homeost. Agents 1994, 8, 126–129. [Google Scholar]
- Murakami, Y.; Yuhara, K.; Takada, N.; Arai, T.; Tsuda, S.; Takamatsu, S.; Machino, M.; Fujisawa, S. Effect of melatonin on cyclooxygenase-2 expression and nuclear factor-kappa B activation in RAW264.7 macrophage-like cells stimulated with fimbriae of Porphyromonas gingivalis. In Vivo 2011, 25, 641–647. [Google Scholar]
- Dubocovich, M.L.; Masana, M.I.; Iacob, S.; Sauri, D.M. Melatonin receptor antagonists that differentiate between the human Mel1a and Mel1b recombinant subtypes are used to assess the pharmacological profile of the rabbit retina ML1 presynaptic heteroreceptor. Naunyn Schmiedebergs. Arch. Pharmacol. 1997, 355, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-K.; Kleszczynski, K.; Janjetovic, Z.; Sweatman, T.; Lin, Z.; Li, W.; Reiter, R.J.; Fischer, T.W.; Slominski, A.T. Metabolism of melatonin and biological activity of intermediates of melatoninergic pathway in human skin cells. FASEB J. 2013, 27, 2742–2755. [Google Scholar] [CrossRef] [PubMed]
- García-Pergañeda, A.; Guerrero, J.M.; Rafii-El-Idrissi, M.; Paz Romero, M.; Pozo, D.; Calvo, J.R. Characterization of membrane melatonin receptor in mouse peritoneal macrophages: Inhibition of adenylyl cyclase by a pertussis toxin-sensitive G protein. J. Neuroimmunol. 1999, 95, 85–94. [Google Scholar] [CrossRef]
- García-Mauriño, S.; Pozo, D.; Calvo, J.R.; Guerrero, J.M. Correlation between nuclear melatonin receptor expression and enhanced cytokine production in human lymphocytic and monocytic cell lines. J. Pineal Res. 2000, 29, 129–137. [Google Scholar] [CrossRef]
- Muxel, S.M.; Pires-Lapa, M.A.; Monteiro, A.W.A.; Cecon, E.; Tamura, E.K.; Floeter-Winter, L.M.; Markus, R.P. NF-κB drives the synthesis of melatonin in RAW 264.7 macrophages by inducing the transcription of the arylalkylamine-N-acetyltransferase (AA-NAT) gene. PLoS ONE 2012, 7, e52010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutin, J.A.; Audinot, V.; Ferry, G.; Delagrange, P. Molecular tools to study melatonin pathways and actions. Trends Pharmacol. Sci. 2005, 26, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin: Signaling mechanisms of a pleiotropic agent. Biofactors 2009, 35, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-H.; Kang, J.W.; Lee, E.-M.; Kim, J.S.; Rhee, Y.H.; Kim, M.; Jeong, S.J.; Park, Y.G.; Kim, S.H. Melatonin promotes osteoblastic differentiation through the BMP/ERK/Wnt signaling pathways. J. Pineal Res. 2011, 51, 187–194. [Google Scholar] [CrossRef]
- Luchetti, F.; Betti, M.; Canonico, B.; Arcangeletti, M.; Ferri, P.; Galli, F.; Papa, S. ERK MAPK activation mediates the antiapoptotic signaling of melatonin in UVB-stressed U937 cells. Free Radic. Biol. Med. 2009, 46, 339–351. [Google Scholar] [CrossRef]
- Pandi-Perumal, S.R.; Trakht, I.; Srinivasan, V.; Spence, D.W.; Maestroni, G.J.M.; Zisapel, N.; Cardinali, D.P. Physiological effects of melatonin: Role of melatonin receptors and signal transduction pathways. Prog. Neurobiol. 2008, 85, 335–353. [Google Scholar] [CrossRef]
- Lai, A.; Cox, C.D.; Chandra Sekar, N.; Thurgood, P.; Jaworowski, A.; Peter, K.; Baratchi, S. Mechanosensing by Piezo1 and its implications for physiology and various pathologies. Biol. Rev. Camb. Philos. Soc. 2022, 97, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Schröder, A.; Alefeld, A.; Forneck, A.; Spanier, G.; Deschner, J.; Proff, P.; Kirschneck, C. Impact of melatonin on periodontal ligament fibroblasts during mechanical strain. Eur. J. Orthod. 2022. [Google Scholar] [CrossRef] [PubMed]
- Nazet, U.; Schröder, A.; Spanier, G.; Wolf, M.; Proff, P.; Kirschneck, C. Simplified method for applying static isotropic tensile strain in cell culture experiments with identification of valid RT-qPCR reference genes for PDL fibroblasts. Eur. J. Orthod. 2020, 42, 359–370. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Gene Name | Forward Primer | Reverse Primer |
|---|---|---|---|
| Eef1a1 | Eukaryotic translation elongation factor-1-alpha-1 | AAAACATGATTACAGGCACATCCC | GCCCGTTCTTGGAGATACCAG |
| Gapdh | Glyceraldehyde-3-phosphate dehydrogenase | GTCATCCCAGAGCTGAACGG | ATGCCTGCTTCACCACCTTC |
| Il1β | Interleukin-1-beta | GTGTAATGAAAGACGGCACACC | ACCAGTTGGGGAACTCTGC |
| Il6 | Interleukin 6 | AAAGCCAGAGTCCTTCAGAGAG | CCTTAGCCACTCCTTCTGTGAC |
| Ptgs2 | Prostaglandin-endoperoxide-Synthase 2 | TCCCTGAAGCCGTACACATC | TCCCCAAAGATAGCATCTGGAC |
| Sdha | Succinate dehydrogenase complex, subunit A | AACACTGGAGGAAGCACACC | AGTAGGAGCGGATAGCAGGAG |
| Tbp | TATA box binding protein | CTATCACTCCTGCCACACCAG | CACGAAGTGCAATGGTCTTTAGG |
| Tnfα | Tumor necrosis factor alpha | ACAAGCCTGTAGCCCACGTC | TTGTTGTCTTTGAGATCCATGCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paddenberg, E.; Forneck, A.; Widbiller, M.; Smeda, M.; Jantsch, J.; Proff, P.; Kirschneck, C.; Schröder, A. Impact of Melatonin on RAW264.7 Macrophages during Mechanical Strain. Int. J. Mol. Sci. 2022, 23, 13397. https://doi.org/10.3390/ijms232113397
Paddenberg E, Forneck A, Widbiller M, Smeda M, Jantsch J, Proff P, Kirschneck C, Schröder A. Impact of Melatonin on RAW264.7 Macrophages during Mechanical Strain. International Journal of Molecular Sciences. 2022; 23(21):13397. https://doi.org/10.3390/ijms232113397
Chicago/Turabian StylePaddenberg, Eva, Anne Forneck, Matthias Widbiller, Martyna Smeda, Jonathan Jantsch, Peter Proff, Christian Kirschneck, and Agnes Schröder. 2022. "Impact of Melatonin on RAW264.7 Macrophages during Mechanical Strain" International Journal of Molecular Sciences 23, no. 21: 13397. https://doi.org/10.3390/ijms232113397