
Effects of Pollen Sources on Fruit Set and Fruit Characteristics of ‘Fengtangli’ Plum (Prunus salicina Lindl.) Based on Microscopic and Transcriptomic Analysis
 , , and
, , and
Abstract
:1. Introduction
2. Results
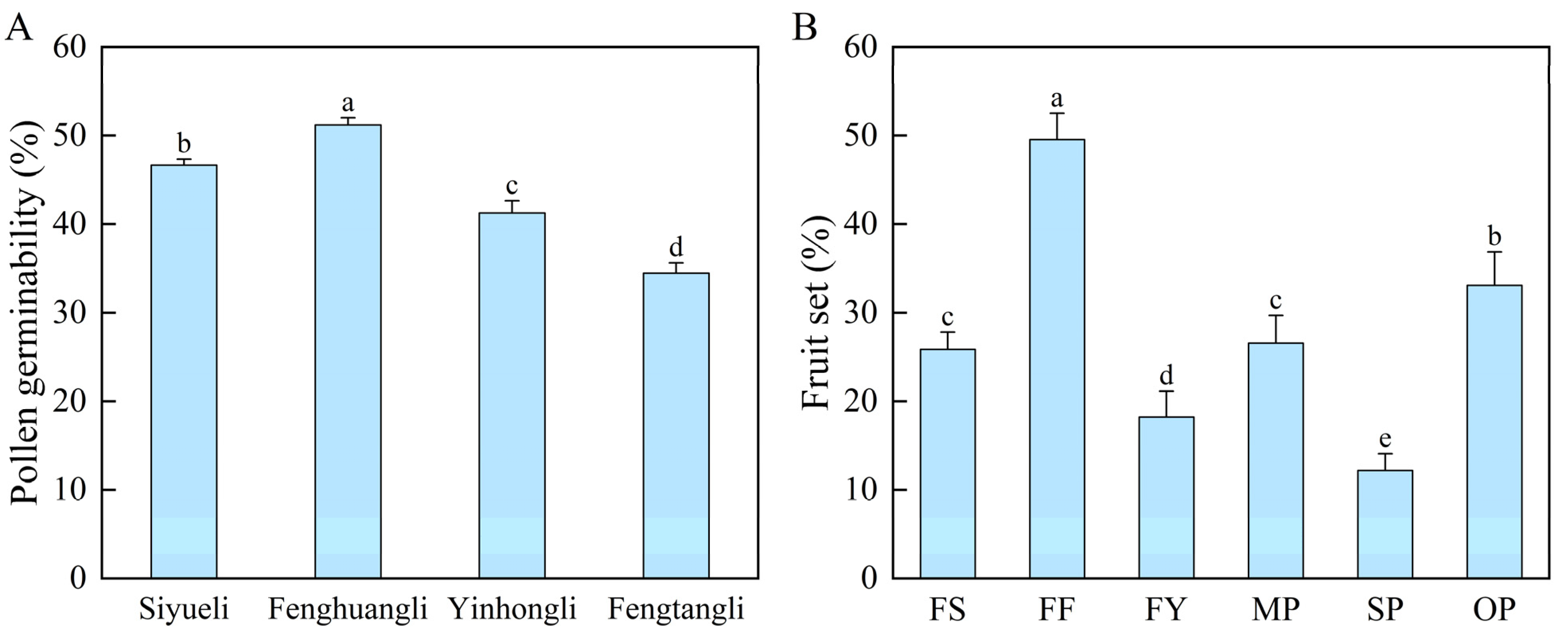
2.1. Pollen Germinability
2.2. Fruit Set
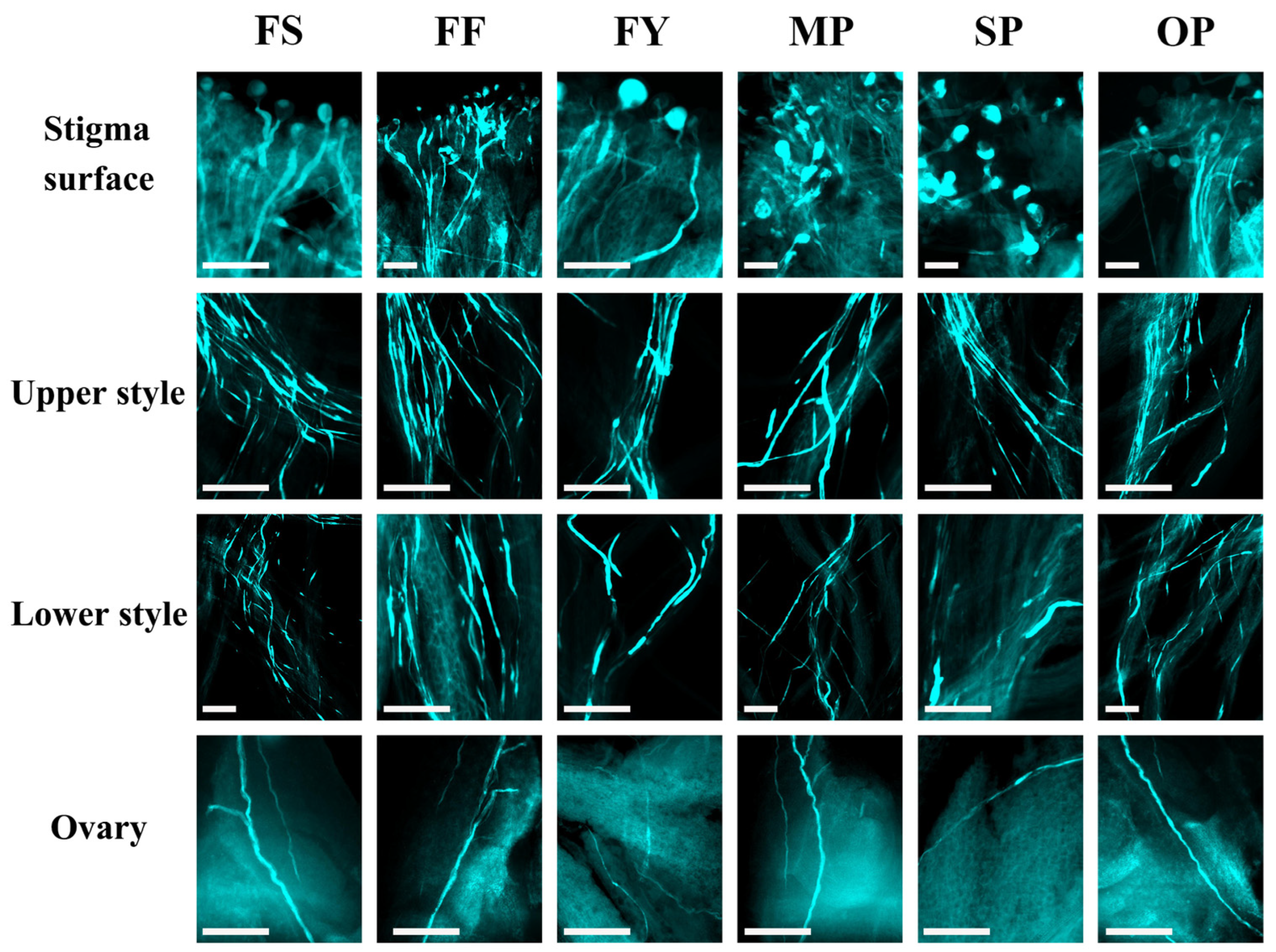
2.3. Pollen Tube Growth In Vivo
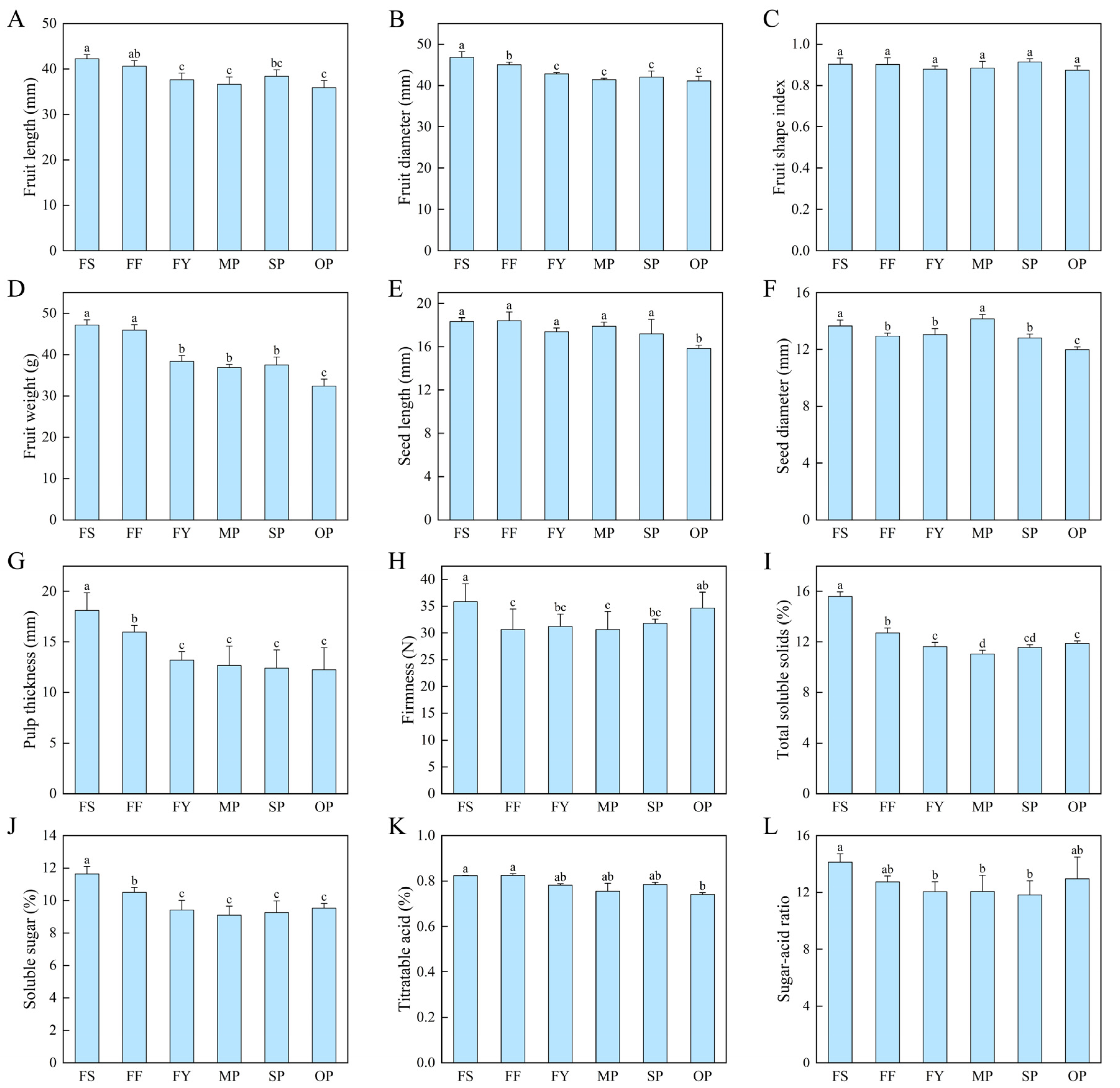
2.4. Metaxenia Effect on Fruit Characteristics
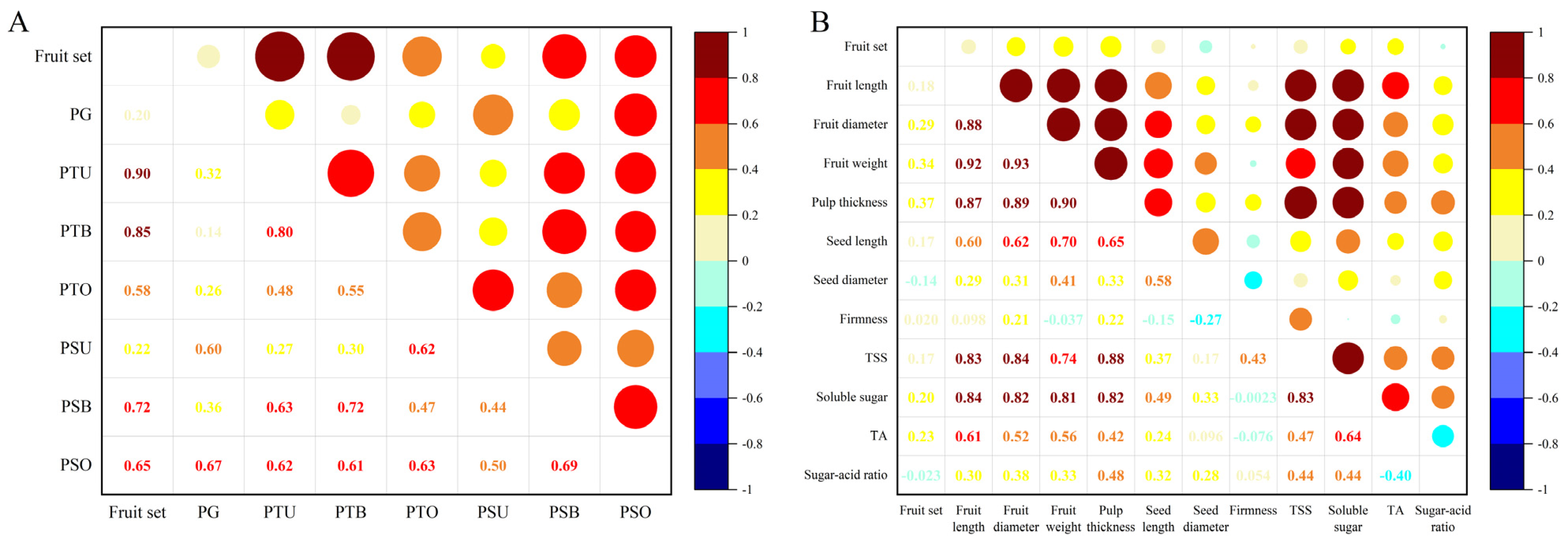
2.5. Correlation Analysis
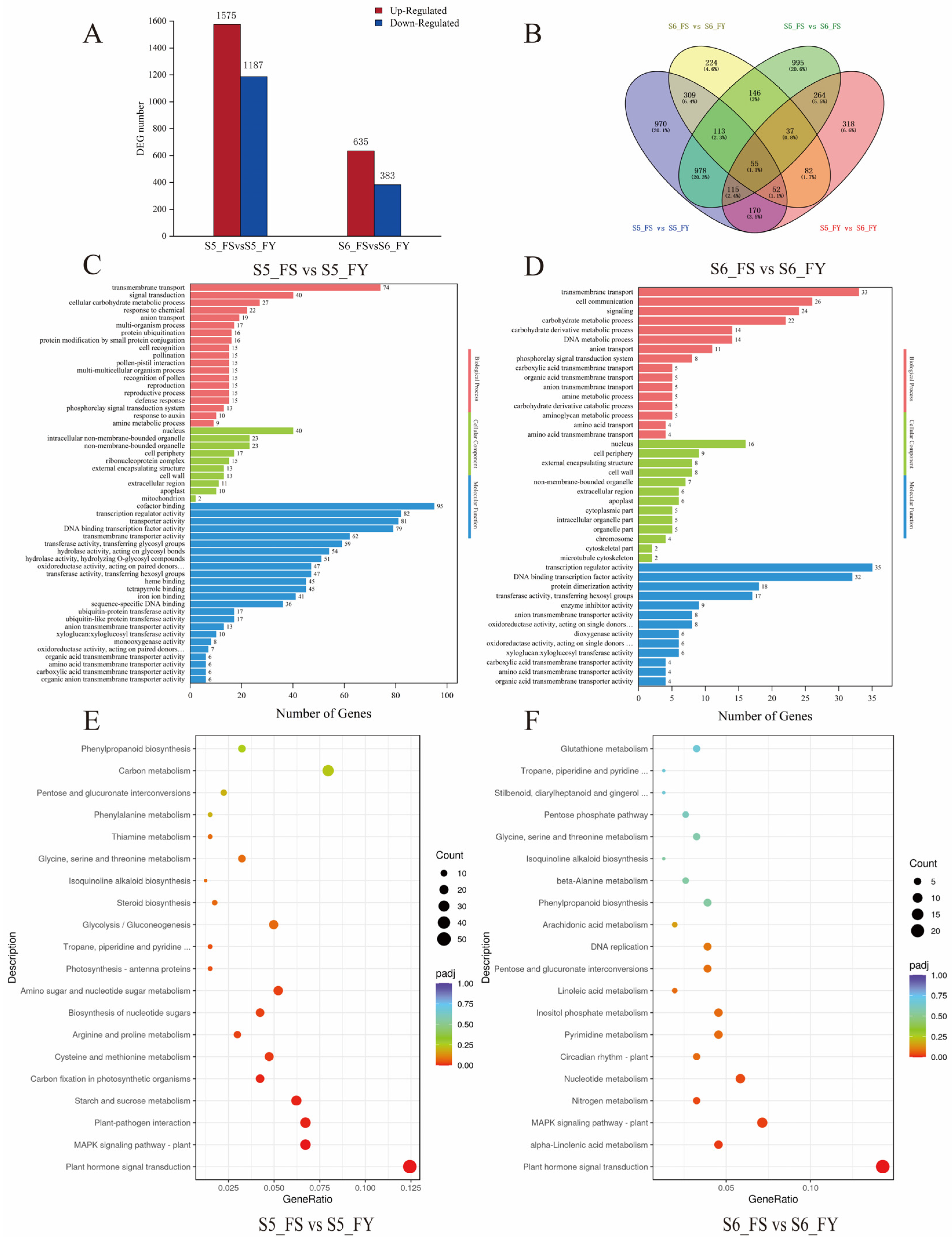
2.6. Effects of Metaxenia on Transcriptomic Alterations
2.7. Effects of Metaxenia on Functional Enrichments of DEGs
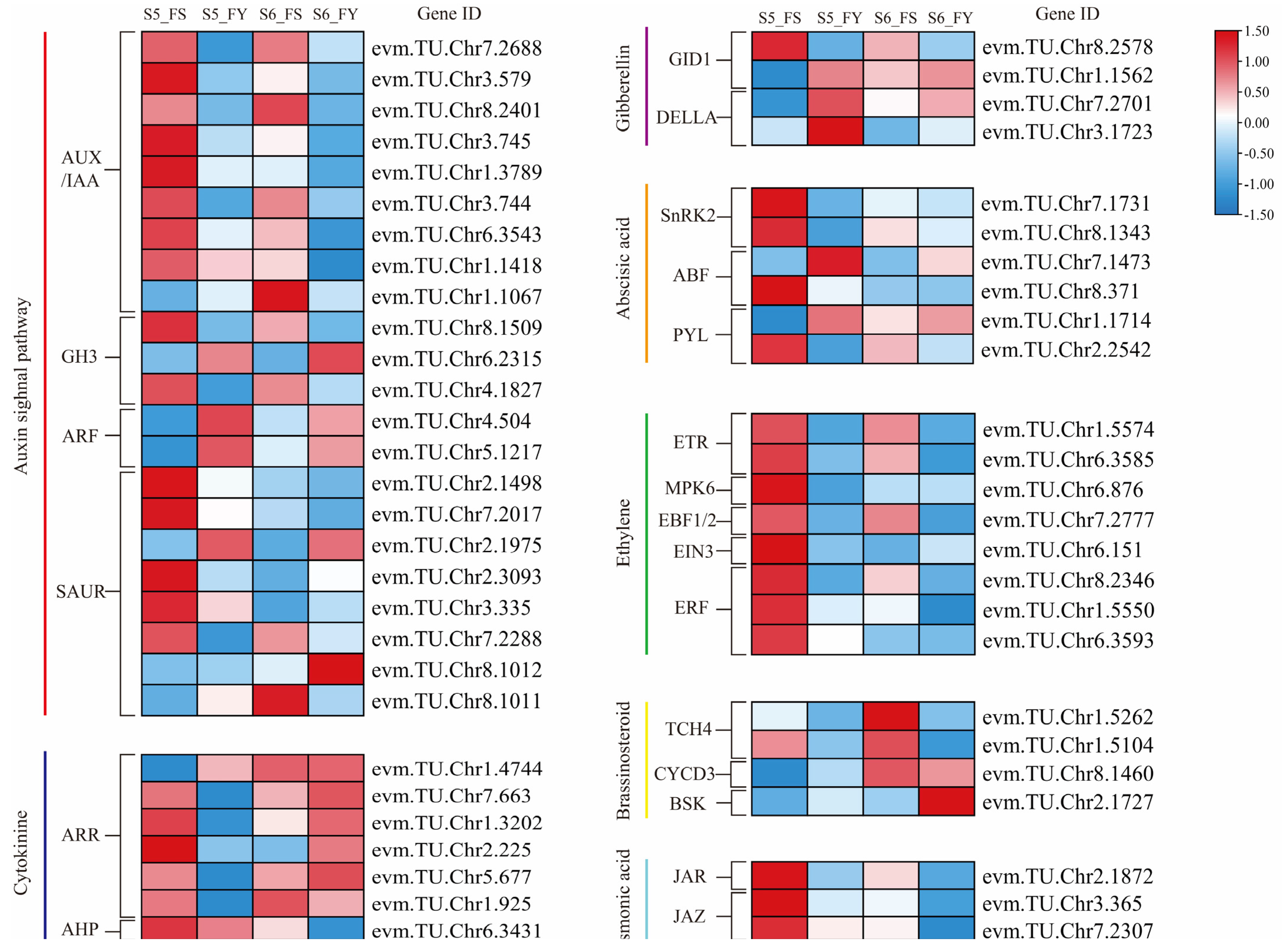
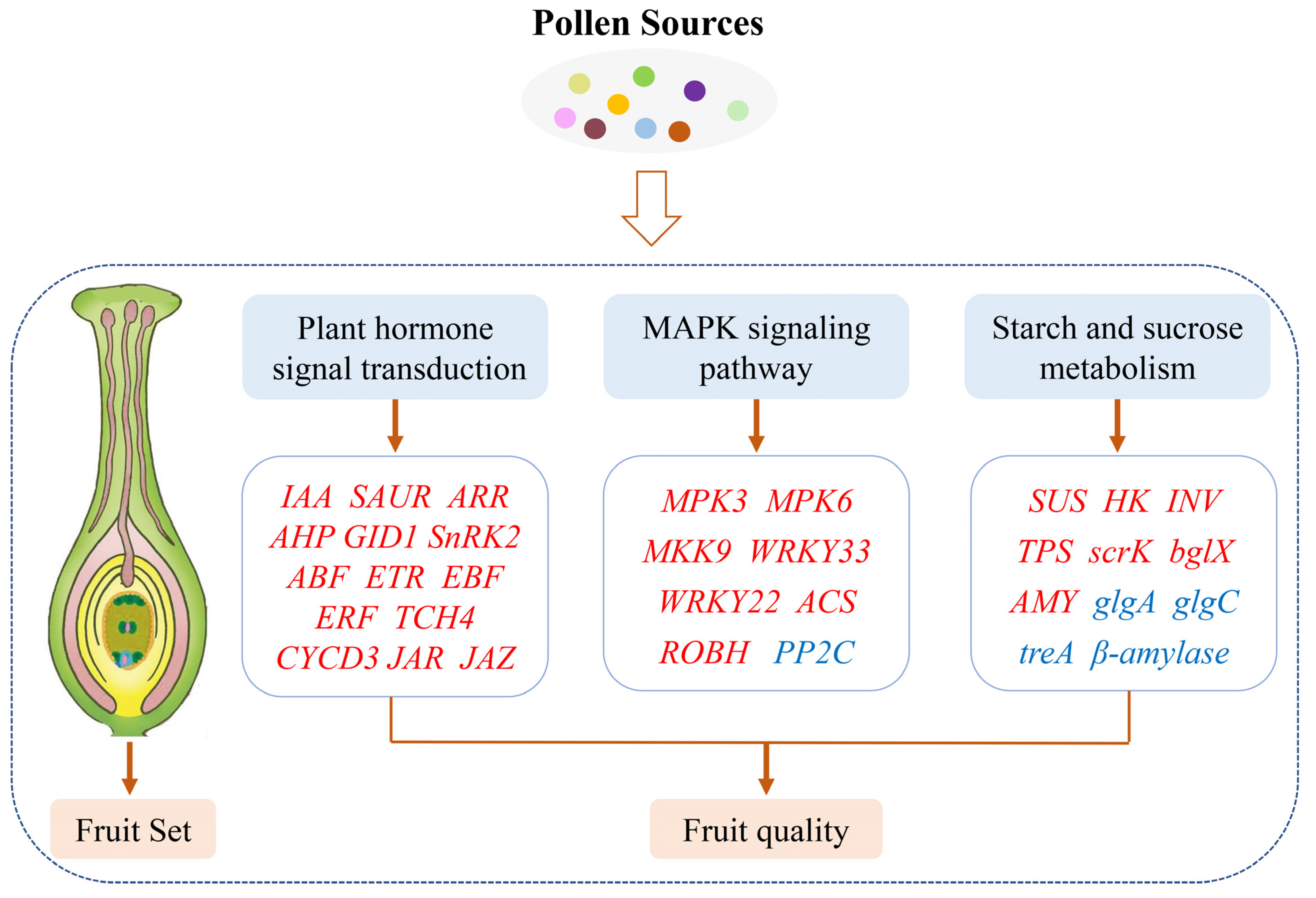
2.8. DEGs Related to Plant Hormone Signal Transduction
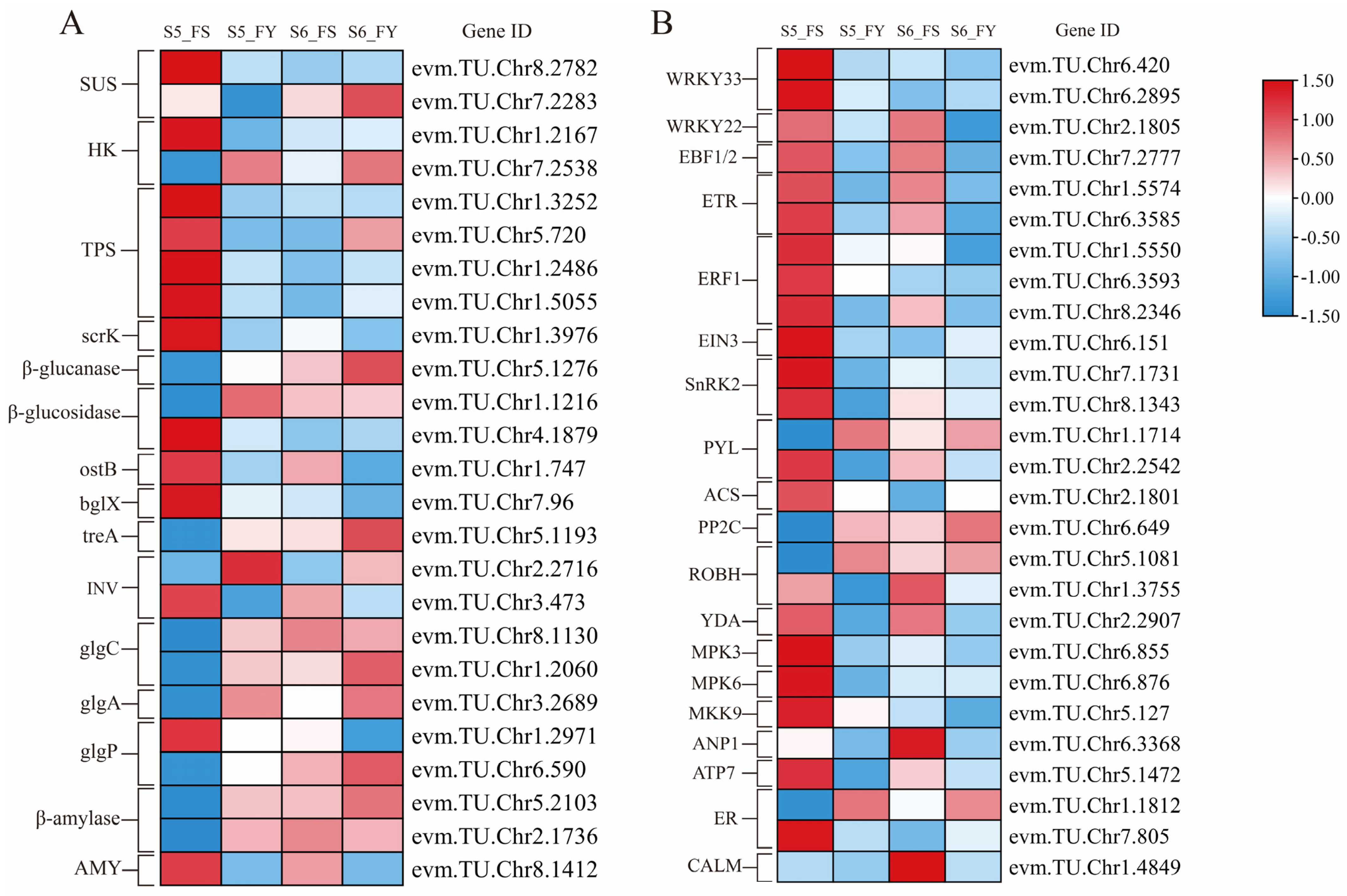
2.9. DEGs Related to Starch and Sucrose Metabolism
2.10. DEGs Related to MAPK Signaling Pathway
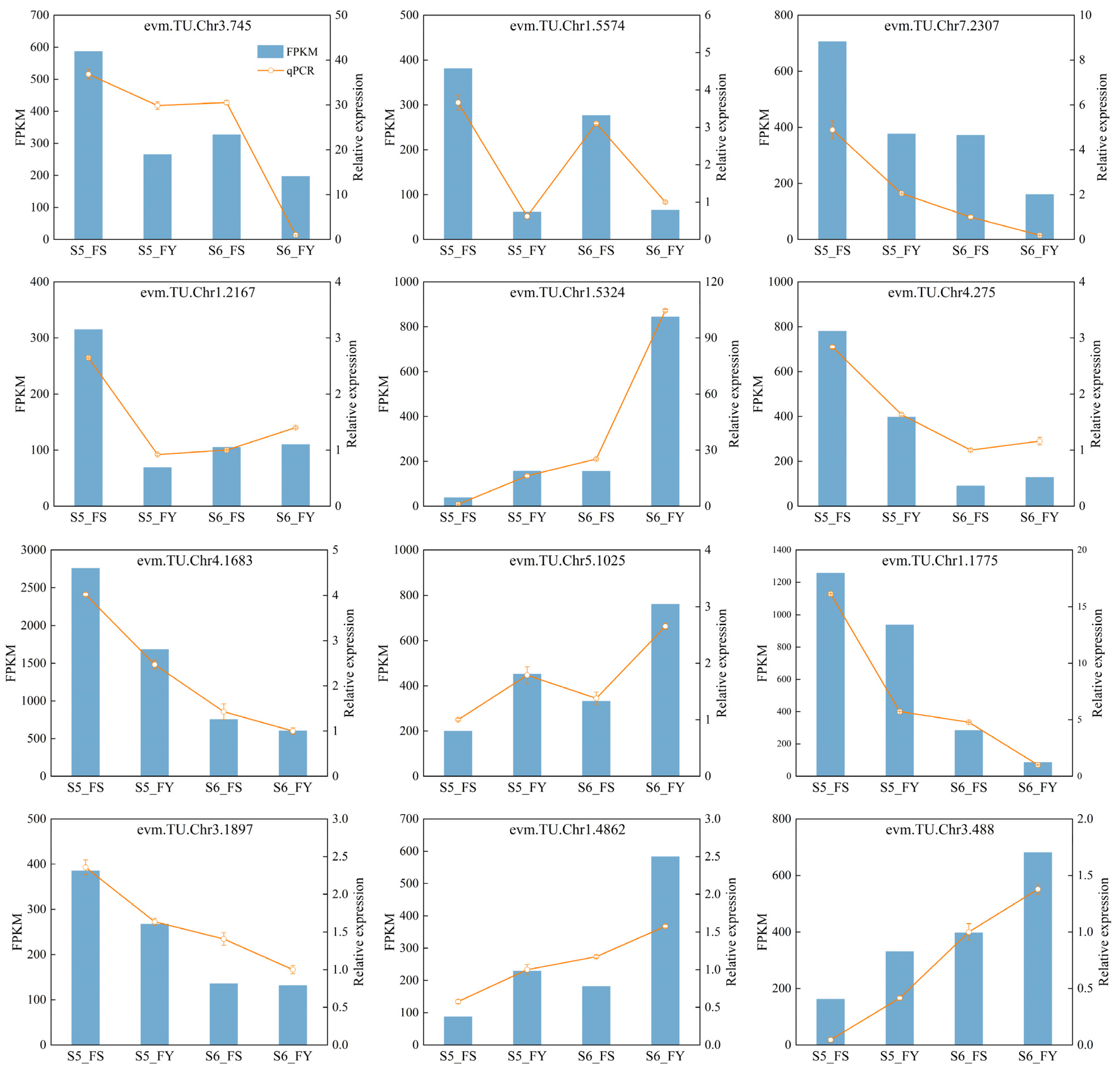
2.11. qRT-PCR Verification for RNA-seq Data
3. Discussion
3.1. Pollen Germinability, Pollen Tube Growth and Fruit Set
3.2. Metaxenia Effect on ‘Fengtangli’ Plum Fruit Characteristics
4. Materials and Methods
4.1. Plant Materials
4.2. Pollen Germinability
4.3. Pollination Experiments
4.4. Evaluation of Fruit Set
4.5. Pollen Tube Growth In Vivo
4.6. Determination of Fruit Characteristics
4.7. RNA Extraction, Library Construction and Sequencing
4.8. Analysis of the RNA-seq Data
4.9. Functional Classification and Pathway Enrichment
4.10. Quantitative Real-Time PCR (qRT-PCR) Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glowacka, A.; Sitarek, M.; Rozpara, E.; Podwyszynska, M. Pomological characteristics and ploidy levels of Japanese plum (Prunus salicina Lindl.) cultivars preserved in Poland. Plants 2021, 10, 884. [Google Scholar] [CrossRef]
- FAOSTAT. 2020. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 13 June 2022).
- Huan, C.; Xu, Q.; Shuling, S.; Dong, J.; Zheng, X. Effect of benzothiadiazole treatment on quality and anthocyanin biosynthesis in plum fruit during storage at ambient temperature. J. Sci. Food Agric. 2021, 101, 3176–3185. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Lu, Y.; Li, Q.; Lin, Z.; Qiu, J.; Peng, C.; Brennan, C.S.; Guo, X. The combination of hot air and chitosan treatments on phytochemical changes during postharvest storage of ‘Sanhua’ plum fruits. Foods 2019, 8, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liaudanskas, M.; Okuleviciute, R.; Lanauskas, J.; Kviklys, D.; Zymone, K.; Rendyuk, T.; Zvikas, V.; Uselis, N.; Janulis, V. Variability in the content of phenolic compounds in plum fruit. Plants 2020, 9, 1611. [Google Scholar] [CrossRef]
- Wang, L.; Sang, W.; Xu, R.; Cao, J. Alteration of flesh color and enhancement of bioactive substances via the stimulation of anthocyanin biosynthesis in ‘Friar’ plum fruit by low temperature and the removal. Food Chem. 2020, 310, 125862. [Google Scholar] [CrossRef]
- Xu, R.; Wang, L.; Li, K.; Cao, J.; Zhao, Z. Integrative transcriptomic and metabolomic alterations unravel the effect of melatonin on mitigating postharvest chilling injury upon plum (cv. Friar) fruit. Postharvest Biol. Technol. 2022, 186, 111819. [Google Scholar] [CrossRef]
- Yu, J.; Li, W.; You, B.; Yang, S.; Xian, W.; Deng, Y.; Huang, W.; Yang, R. Phenolic profiles, bioaccessibility and antioxidant activity of plum (Prunus salicina Lindl). Food Res. Int. 2021, 143, 110300. [Google Scholar] [CrossRef] [PubMed]
- Kubo, K.; Entani, T.; Takara, A.; Wang, N.; Fields, A.M.; Hua, Z.; Toyoda, M.; Kawashima, S.; Ando, T.; Isogai, A.; et al. Collaborative non-self recognition system in S-RNase-based self-incompatibility. Science 2010, 330, 796–799. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Iezzoni, A.F. The S-RNase-based gametophytic self-incompatibility system in Prunus exhibits distinct genetic and molecular features. Sci. Hortic. 2010, 124, 423–433. [Google Scholar] [CrossRef]
- Beppu, K.; Kumai, M.; Yamane, H.; Tao, R.; Kataoka, I. Molecular and genetic analyses of the S-haplotype of the self-compatible Japanese plum (Prunus salicina Lindl.) ‘Methley’. J. Horticult. Sci. Biotechnol. 2012, 87, 493–498. [Google Scholar] [CrossRef]
- Cachi, A.M.; Wunsch, A. Characterization and mapping of non-S gametophytic self-compatibility in sweet cherry (Prunus avium L.). J. Exp. Bot. 2011, 62, 1847–1856. [Google Scholar] [CrossRef] [Green Version]
- Halasz, J.; Pedryc, A.; Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytol. 2007, 176, 792–803. [Google Scholar] [CrossRef]
- Kodad, O.; Socias i Company, R.; Alonso, J.M. Unilateral recognition of the Sf allele in almond. Sci. Hortic. 2015, 185, 29–33. [Google Scholar] [CrossRef]
- Sapir, G.; Stern, R.A.; Shafir, S.; Goldway, M. Full compatibility is superior to semi-compatibility for fruit set in Japanese plum (Prunus salicina Lindl.) cultivars. Sci. Hortic. 2008, 116, 394–398. [Google Scholar] [CrossRef]
- Kämper, W.; Thorp, G.; Wirthensohn, M.; Brooks, P.; Trueman, S.J. Pollen paternity can affect kernel size and nutritional composition of self-incompatible and new self-compatible almond cultivars. Agronomy 2021, 11, 326. [Google Scholar] [CrossRef]
- Guerra, M.E.; Guerrero, B.I.; Casadomet, C.; Rodrigo, J. Self-(in)compatibility, S-RNase allele identification, and selection of pollinizers in new Japanese plum-type cultivars. Sci. Hortic. 2020, 261, 109022. [Google Scholar] [CrossRef]
- Herrera, S.; Lora, J.; Hormaza, J.I.; Herrero, M.; Rodrigo, J. Optimizing production in the new generation of apricot cultivars: Self-incompatibility, S-RNase allele identification, and incompatibility group assignment. Front. Plant Sci. 2018, 9, 527. [Google Scholar] [CrossRef] [Green Version]
- Alinezhad Jahromi, H.; Zarei, A.; Mohammadkhani, A. Analysis the effects of pollen grain sources on the fruits set and their characteristics of ‘Clementine’ mandarin using microscopic and molecular approaches. Sci. Hortic. 2019, 249, 347–354. [Google Scholar] [CrossRef]
- Guerra, M.E.; Wunsch, A.; Lopez-Corrales, M.; Rodrigo, J. Flower emasculation as the cause for lack of fruit set in Japanese plum crosses. J. Am. Soc. Hortic. Sci. 2010, 135, 556–562. [Google Scholar] [CrossRef] [Green Version]
- Guerra, M.E.; Rodrigo, J. Japanese plum pollination: A review. Sci. Hortic. 2015, 197, 674–686. [Google Scholar] [CrossRef]
- Milatović, D.; Nikolić, D.; Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Hortic. Sci. 2013, 40, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.J.; He, F.J.; Xiong, C.Z.; Zhu, F.R.; Okamoto, G. Influences of cross pollination on pollen tube growth and fruit set in Zuili plums (Prunus salicina). J. Integr. Plant Biol. 2008, 50, 203–209. [Google Scholar] [CrossRef]
- Radunić, M.; Jazbec, A.; Ercisli, S.; Čmelik, Z.; Ban, S.G. Pollen-pistil interaction influence on the fruit set of sweet cherry. Sci. Hortic. 2017, 224, 358–366. [Google Scholar] [CrossRef]
- Denney, J.O. Xenia includes metaxenia. HortScience 1992, 27, 722–728. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Shi, Z.; Zhu, Q.; Tao, L.; Liang, W.; Zhao, Z. Transcriptome sequencing and differential expression analysis of seed starch accumulation in Chinese chestnut metaxenia. BMC Genom. 2021, 22, 617. [Google Scholar] [CrossRef] [PubMed]
- Herbert, S.W.; Walton, D.A.; Wallace, H.M. Pollen-parent affects fruit, nut and kernel development of Macadamia. Sci. Hortic. 2019, 244, 406–412. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, C.; Yao, J.L.; Deng, C.H.; Chen, S.; Chen, J.; Wang, Z.; Yu, Q.; Cheng, Y.; Xu, J. Citrus mangshanensis pollen confers a xenia effect on linalool oxide accumulation in pummelo fruit by enhancing the expression of a cytochrome P450 78A7 gene CitLO1. J. Agric. Food Chem. 2019, 67, 9468–9476. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, Z.; Mao, Y.; Hu, Y.; Yang, L.; Wang, Y.; Zhang, L.; Shen, X. Effects of quince pollen pollination on fruit qualities and phenolic substance contents of apples. Sci. Hortic. 2019, 256, 108628. [Google Scholar] [CrossRef]
- Sabir, A. Xenia and metaxenia in grapes: Differences in berry and seed characteristics of maternal grape cv. ‘Narince’ (Vitis vinifera L.) as influenced by different pollen sources. Plant Biol. 2015, 17, 567–573. [Google Scholar] [CrossRef]
- Cheng, X.; Zhang, J.; Wang, H.; Chen, T.; Li, G.; Yan, C.; Jin, Q.; Lin, Y.; Cai, Y. Effects of metaxenia on stone cell formation in pear (Pyrus bretschneideri) based on transcriptomic analysis and functional characterization of the lignin-related gene PbC4H2. Forests 2020, 11, 53. [Google Scholar] [CrossRef]
- Doi, K.; Inoue, R.; Iwasaki, N. Seed weight mediates effects of pollen on berry weight, ripening, and anthocyanin content in highbush blueberry. Sci. Hortic. 2021, 288, 110313. [Google Scholar] [CrossRef]
- Kodad, O.; Estopanan, G.; Juan, T.; Company, R.S.I. Xenia effects on oil content and fatty acid and tocopherol concentrations in autogamous almond cultivars. J. Agric. Food Chem. 2009, 57, 10809–10813. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.S. A novel mechanism for xenia? Hortscience 2008, 43, 706. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Canio, W.; Kessler, S.; Sinha, N. Developmental changes due to long-distance movement of a homeobox fusion transcript in tomato. Science 2001, 293, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Fattorini, R.; Glover, B.J. Molecular mechanisms of pollination biology. Annu. Rev. Plant Biol. 2020, 71, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Sharafi, Y. Pollen viability and longevity in some selected genotypes of peach, plum, prune and sour cherry. J. Med. Plants Res. 2011, 5, 275–279. [Google Scholar]
- Sakata, T.; Takahashi, H.; Nishiyama, I.; Higashitani, A. Effects of high temperature on the development of pollen mother cells and microspores in barley Hordeum vulgare L. J. Plant Res. 2000, 113, 395–402. [Google Scholar] [CrossRef]
- Glišić, I.; Milatović, D.; Cerović, R.; Radičević, S.; Đorđević, M.; Milošević, N. Examination of self-compatibility in promising plum (Prunus domestica L.) genotypes developed at the Fruit Research Institute, Čačak. Sci. Hortic. 2017, 224, 156–162. [Google Scholar] [CrossRef]
- Beltran, R.; Valls, A.; Cebrian, N.; Zornoza, C.; Garcia Breijo, F.; Reig Arminana, J.; Garmendia, A.; Merle, H. Effect of temperature on pollen germination for several Rosaceae species: Influence of freezing conservation time on germination patterns. PeerJ 2019, 7, e8195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.J.; Zhao, T.Y.; Ouyang, X.K.; Zhao, X.Y.; Dai, X.; Gao, X.Q. Pollen-specific gene SKU5-SIMILAR 13 enhances growth of pollen tubes in the transmitting tract in Arabidopsis. J. Exp. Bot. 2021, 73, 696–710. [Google Scholar] [CrossRef]
- Zhang, D.; Wengier, D.; Shuai, B.; Gui, C.P.; Muschietti, J.; McCormick, S.; Tang, W.H. The pollen receptor kinase LePRK2 mediates growth-promoting signals and positively regulates pollen germination and tube growth. Plant Physiol. 2008, 148, 1368–1379. [Google Scholar] [CrossRef] [Green Version]
- Sankaranarayanan, S.; Jamshed, M.; Samuel, M.A. Proteomics approaches advance our understanding of plant self-incompatibility response. J. Proteome Res. 2013, 12, 4717–4726. [Google Scholar] [CrossRef]
- Pereira, A.M.; Moreira, D.; Coimbra, S.; Masiero, S. Paving the way for fertilization: The role of the transmitting tract. Int. J. Mol. Sci. 2021, 22, 2603. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.Y.; Wang, H.; Wu, H. A floral transmitting tissue-specific glycoprotein attracts pollen tubes and stimulates their growth. Cell 1995, 82, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Muhlemann, J.K.; Younts, T.L.B.; Muday, G.K. Flavonols control pollen tube growth and integrity by regulating ROS homeostasis during high-temperature stress. Proc. Natl. Acad. Sci. USA 2018, 115, E11188–E11197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolić, D.; Milatović, D. Examining self-compatibility in plum (Prunus domestica L.) by fluorescence microscopy. Genetika 2010, 42, 387–396. [Google Scholar] [CrossRef]
- Duarte, M.O.; Oliveira, D.M.T.; Borba, E.L. Two self-incompatibility sites occur simultaneously in the same Acianthera species (Orchidaceae, Pleurothallidinae). Plants 2020, 9, 1758. [Google Scholar] [CrossRef]
- Kwon, J.H.; Jun, J.H.; Nam, E.Y.; Chung, K.H.; Yoon, I.K.; Yun, S.K.; Kim, S.J. Selection of a suitable pollinizer for ‘Summer Fantasia’ plum. Hortscience 2017, 52, 1182–1187. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Gao, Z.; Zhang, Z. Identification of S-genotypes and novel S-RNase alleles in Japanese apricot cultivars native to China. Sci. Hortic. 2010, 123, 459–463. [Google Scholar] [CrossRef]
- Nebel, B.; Trump, I.J. Xenia and metaxenia in apples: II. Proc. Natl. Acad. Sci. USA 1932, 18, 356–359. [Google Scholar] [CrossRef]
- Fattahi, R.; Mohammadzedeh, M.; Khadivi-Khub, A. Influence of different pollen sources on nut and kernel characteristics of hazelnut. Sci. Hortic. 2014, 173, 15–19. [Google Scholar] [CrossRef]
- Żurawicz, E.; Studnicki, M.; Kubik, J.; Pruski, K. A careful choice of compatible pollinizers significantly improves the size of fruits in red raspberry (Rubus idaeus L.). Sci. Hortic. 2018, 235, 253–257. [Google Scholar] [CrossRef]
- Zhang, J.; Meng, Z.; Gong, W. Effect of pollen xenia on fruit quality of Frinar plum cultivar. J. Fruit Sci. 2009, 26, 836–839. [Google Scholar]
- Godoy, F.; Kuhn, N.; Munoz, M.; Marchandon, G.; Gouthu, S.; Deluc, L.; Delrot, S.; Lauvergeat, V.; Arce-Johnson, P. The role of auxin during early berry development in grapevine as revealed by transcript profiling from pollination to fruit set. Hortic. Res. 2021, 8, 140. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Shao, Y.; Ge, S.; Zhang, M.; Zhang, T.; Hu, X.; Liu, Y.; Walker, J.; Zhang, S.; Xu, J. A MAPK cascade downstream of IDA-HAE/HSL2 ligand-receptor pair in lateral root emergence. Nat. Plants 2019, 5, 414–423. [Google Scholar] [CrossRef]
- Zhou, J.G.; Mu, Q.; Wang, X.Y.; Zhang, J.; Yu, H.Z.; Huang, T.Z.; He, Y.X.; Dai, S.J.; Meng, X.Z. Multilayered synergistic regulation of phytoalexin biosynthesis by ethylene, jasmonate, and MAPK signaling pathways in Arabidopsis. Plant Cell 2022, 34, 3066–3087. [Google Scholar] [CrossRef] [PubMed]
- Desnoues, E.; Baldazzi, V.; Genard, M.; Mauroux, J.B.; Lambert, P.; Confolent, C.; Quilot-Turion, B. Dynamic QTLs for sugars and enzyme activities provide an overview of genetic control of sugar metabolism during peach fruit development. J. Exp. Bot. 2016, 67, 3419–3431. [Google Scholar] [CrossRef] [Green Version]
- Tao, H.; Sun, H.; Wang, Y.; Song, X.; Guo, Y. New insights on ‘GALA’ apple fruit development: Sugar and acid accumulation: A transcriptomic approach. J. Plant Growth Regul. 2020, 39, 680–702. [Google Scholar] [CrossRef]
- Zhang, Q.; Shan, C.; Song, W.; Cai, W.; Zhou, F.; Ning, M.; Tang, F. Transcriptome analysis of starch and sucrose metabolism change in Gold Queen Hami melons under different storage temperatures. Postharvest Biol. Technol. 2021, 174, 111445. [Google Scholar] [CrossRef]
- Cerović, R.; Fotirić Akšić, M.; Meland, M. Success rate of individual pollinizers for the pear cultivars “Ingeborg” and “Celina” in a Nordic climate. Agronomy 2020, 10, 970. [Google Scholar] [CrossRef]
- Chen, J.C.; Fang, S.C. The long pollen tube journey and in vitro pollen germination of Phalaenopsis orchids. Plant Reprod. 2016, 29, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, D.; Baraket, G.; Perez, V.; Ben Mustapha, S.; Salhi-Hannachi, A.; Hormaza, J.I. Analysis of self-incompatibility and genetic diversity in diploid and hexaploid plum genotypes. Front. Plant Sci. 2019, 10, 896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Feng, C.; Peng, W.; Hao, J.; Wang, J.; Pan, J.; He, Y. Chromosome-level draft genome of a diploid plum (Prunus salicina). GigaScience 2020, 9, giaa130. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Pollen Grains Germination on Stigma (%) | Pistils (%) with Pollen Tubes | Number of Pollen Tubes | ||||
|---|---|---|---|---|---|---|---|
| At the Upper Third of Style | At the Base of Style | Penetrated to Ovary | At the Upper Third of Style | At the Base of Style | Penetrated to Ovary | ||
| FS | 61.64 ± 3.54 a 1 | 95.45 ± 4.55 a | 54.55 ± 4.54 c | 36.36 ± 3.76 ab | 33.17 ± 5.49 b | 11.33 ± 2.42 c | 1.71 ± 0.49 b |
| FF | 56.91 ± 5.66 ab | 91.30 ± 4.35 a | 78.26 ± 4.30 a | 43.48 ± 2.22 a | 47.33 ± 5.08 a | 23.17 ± 4.31 a | 2.57 ± 0.53 a |
| FY | 53.20 ± 4.10 bc | 92.31 ± 3.85 a | 65.38 ± 4.01 b | 26.92 ± 2.62 cd | 26.50 ± 3.15 c | 9.67 ± 5.09 cd | 0.86 ± 0.38 cd |
| MP | 63.72 ± 2.87 a | 92.00 ± 4.00 a | 76.00 ± 3.80 a | 44.00 ± 2.40 a | 31.83 ± 4.26 b | 16.00 ± 4.15 b | 1.53 ± 0.52 b |
| SP | 48.14 ± 4.53 c | 79.17 ± 4.17 b | 33.33 ± 4.06 d | 20.83 ± 3.39 d | 21.00 ± 3.22 d | 6.17 ± 1.47 d | 0.43 ± 0.44 d |
| OP | 45.92 ± 2.32 c | 83.33 ± 4.16 b | 67.67 ± 2.51 b | 29.17 ± 4.17 bc | 35.67 ± 4.32 b | 19.00 ± 2.53 ab | 1.29 ± 0.48 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, L.; Wang, T.; Hu, J.; Yang, X.; Yao, Y.; Jin, Z.; Huang, Z.; Sun, G.; Xiong, B.; Liao, L.; et al. Effects of Pollen Sources on Fruit Set and Fruit Characteristics of ‘Fengtangli’ Plum (Prunus salicina Lindl.) Based on Microscopic and Transcriptomic Analysis. Int. J. Mol. Sci. 2022, 23, 12959. https://doi.org/10.3390/ijms232112959
Deng L, Wang T, Hu J, Yang X, Yao Y, Jin Z, Huang Z, Sun G, Xiong B, Liao L, et al. Effects of Pollen Sources on Fruit Set and Fruit Characteristics of ‘Fengtangli’ Plum (Prunus salicina Lindl.) Based on Microscopic and Transcriptomic Analysis. International Journal of Molecular Sciences. 2022; 23(21):12959. https://doi.org/10.3390/ijms232112959
Chicago/Turabian StyleDeng, Lijun, Tie Wang, Juan Hu, Xinxia Yang, Yuan Yao, Zhenghua Jin, Zehao Huang, Guochao Sun, Bo Xiong, Ling Liao, and et al. 2022. "Effects of Pollen Sources on Fruit Set and Fruit Characteristics of ‘Fengtangli’ Plum (Prunus salicina Lindl.) Based on Microscopic and Transcriptomic Analysis" International Journal of Molecular Sciences 23, no. 21: 12959. https://doi.org/10.3390/ijms232112959