
Combined Metabolome and Transcriptome Analyses Unveil the Molecular Mechanisms of Fruit Acidity Variation in Litchi (Litchi chinensis Sonn.)
 ,
,
Abstract
:1. Introduction
2. Results
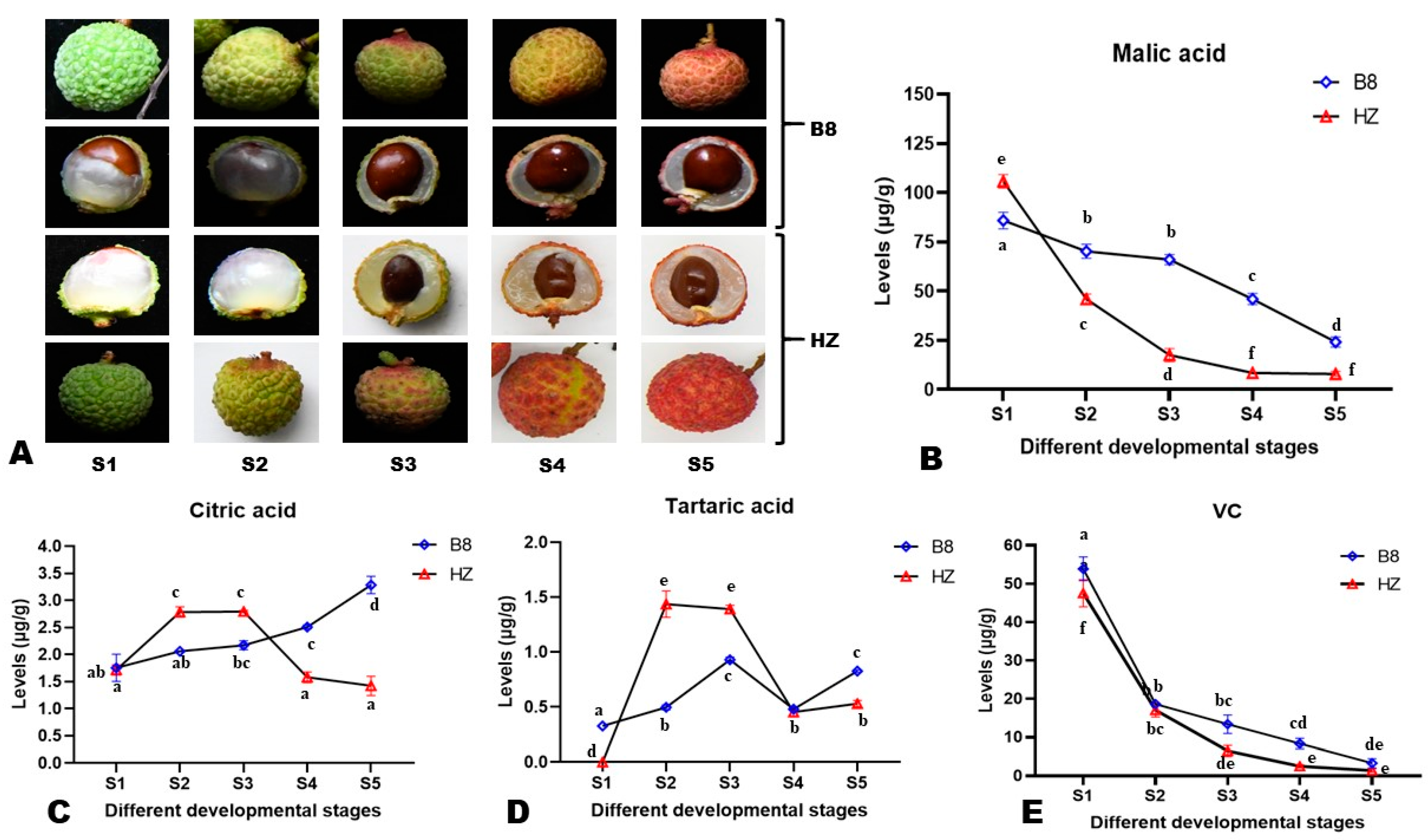
2.1. Dynamic Changes in the Content of Major Organic Acids in Litchi Pulp during Fruit Development
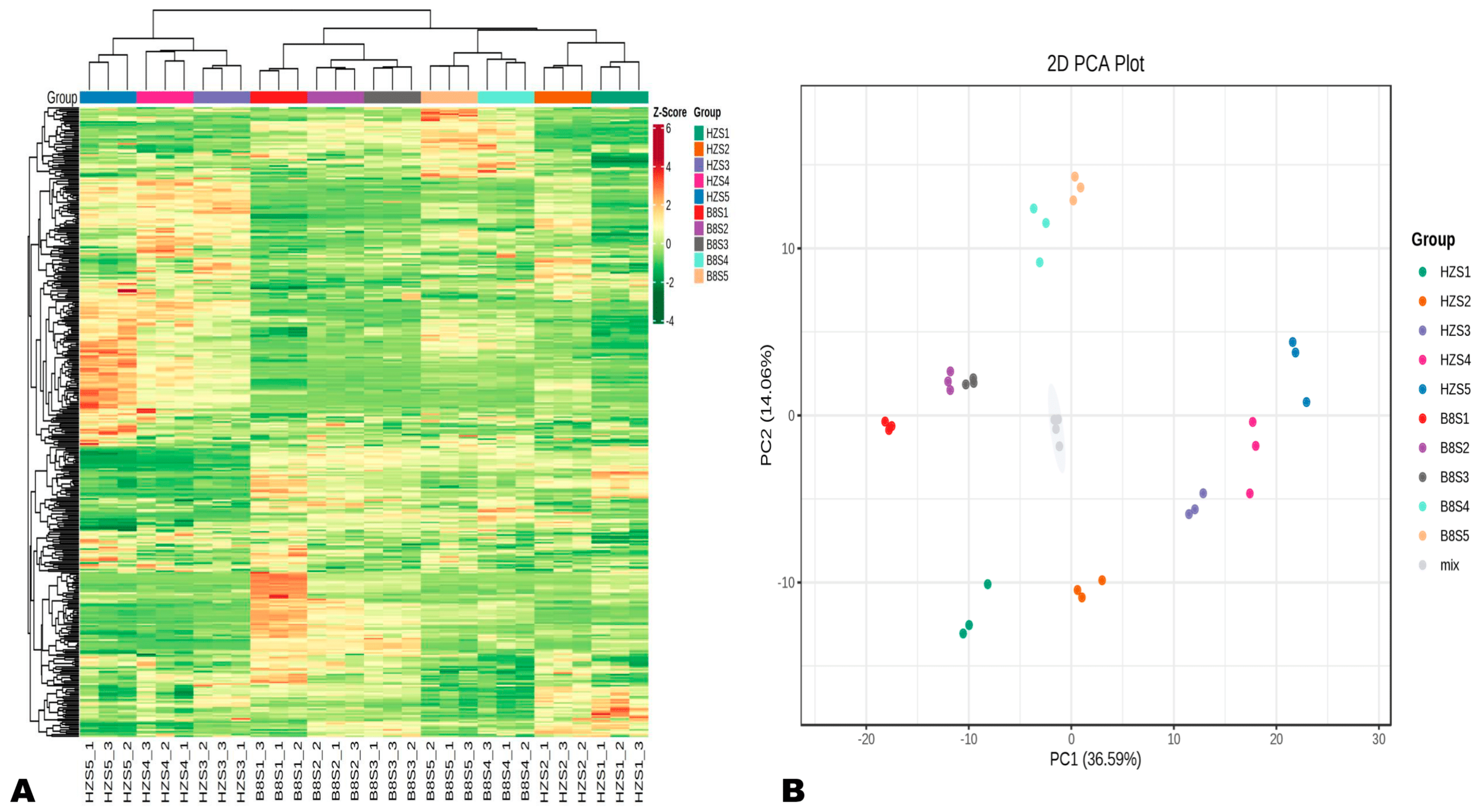
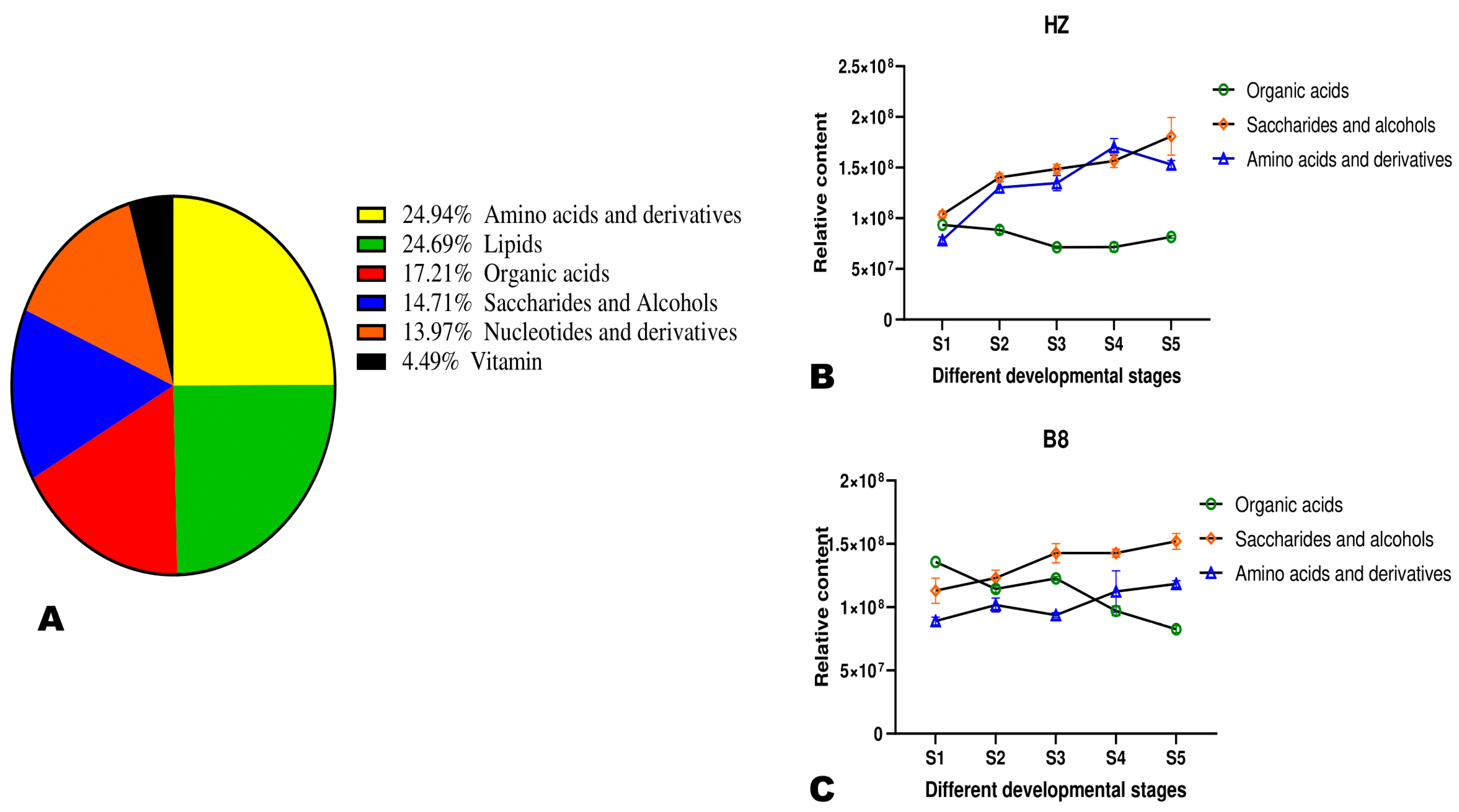
2.2. Diversity and Variation of Metabolites in Litchi Pulp during Fruit Development
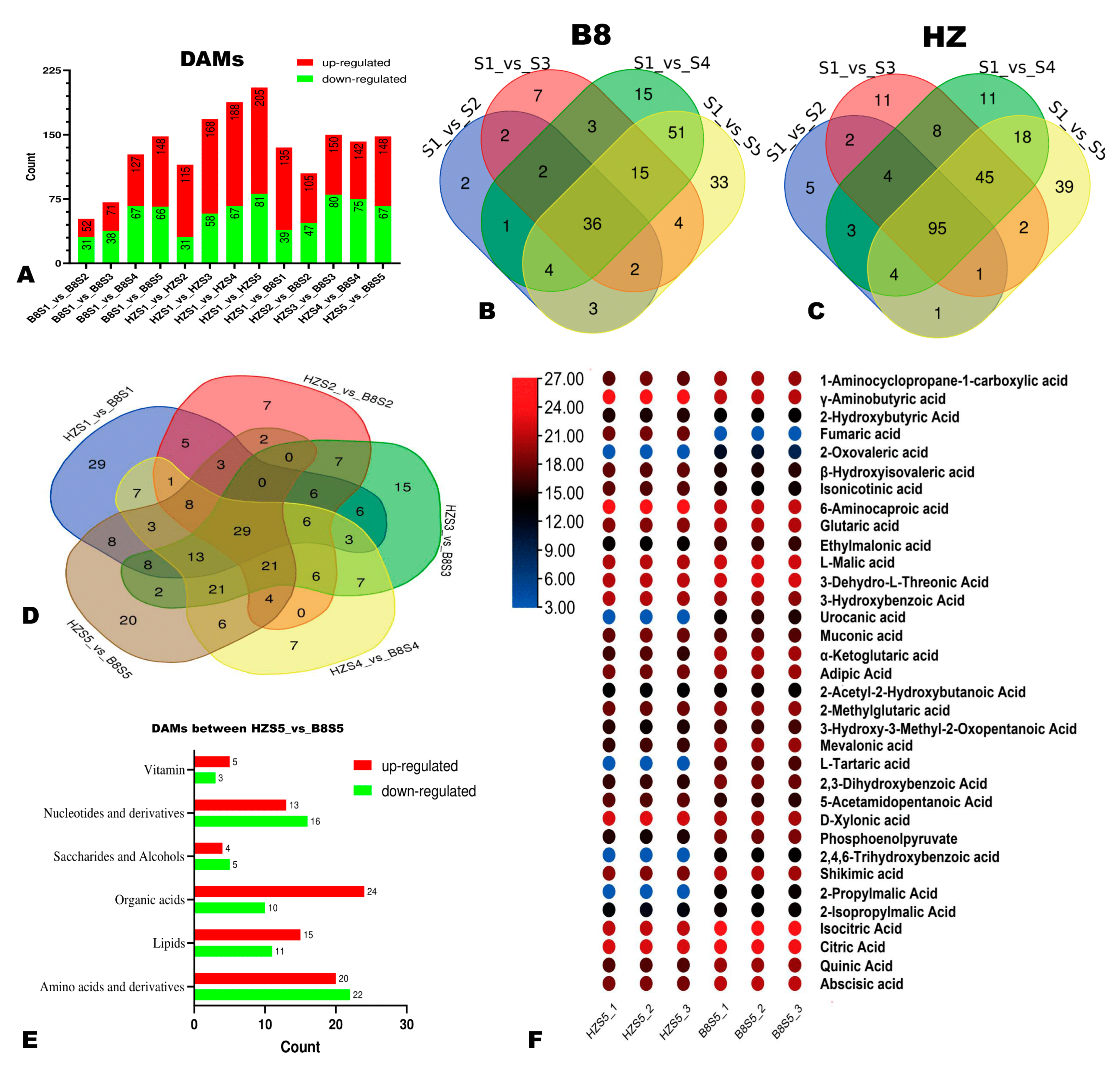
2.3. Differentially Accumulated Metabolites (DAMs) in HZ and B8 Pulps
2.4. Transcriptome Profiles of HZ and B8 Pulps during Fruit Development
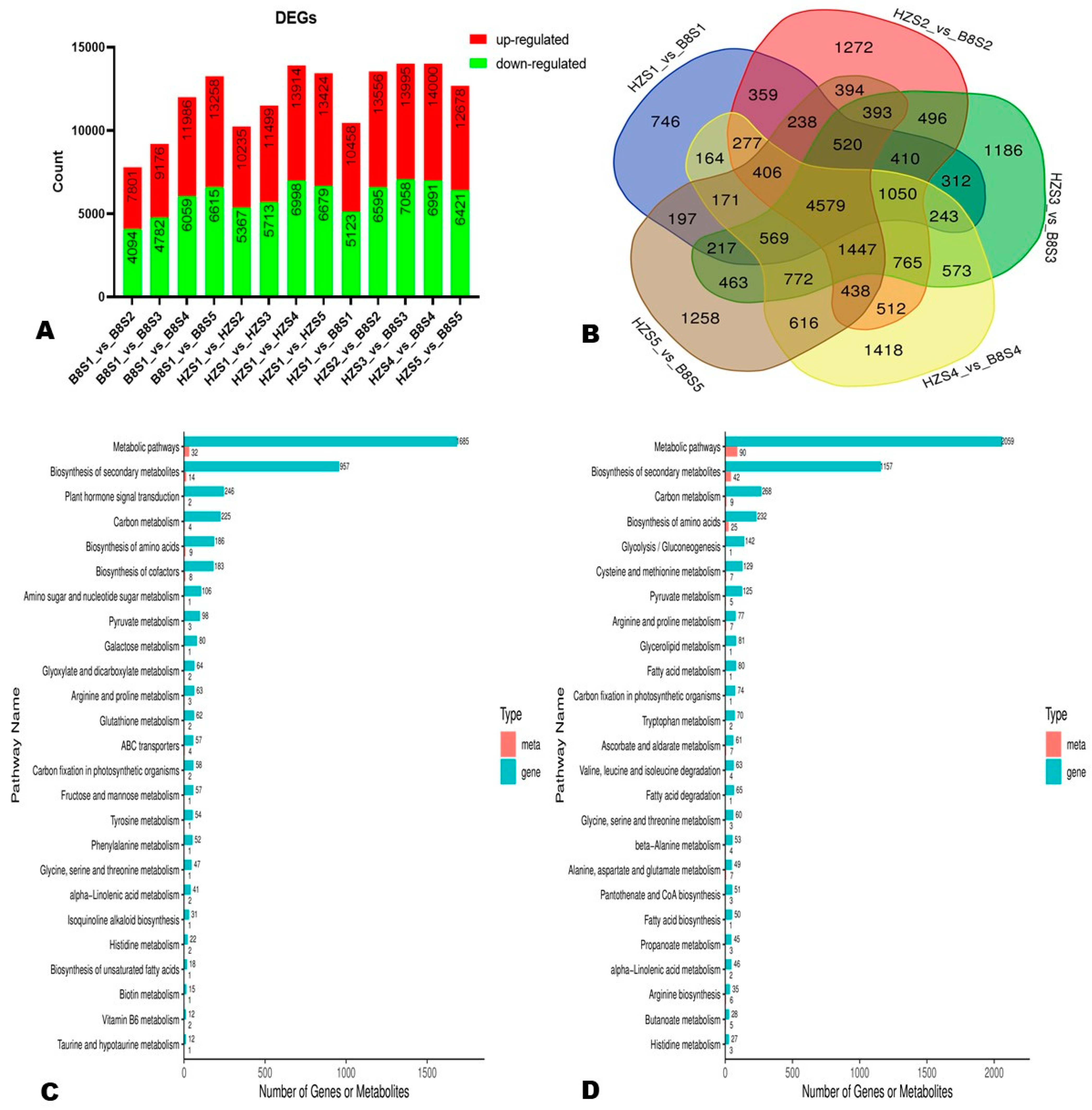
2.5. Differentially Expressed Genes (DEGs) between HZ and B8
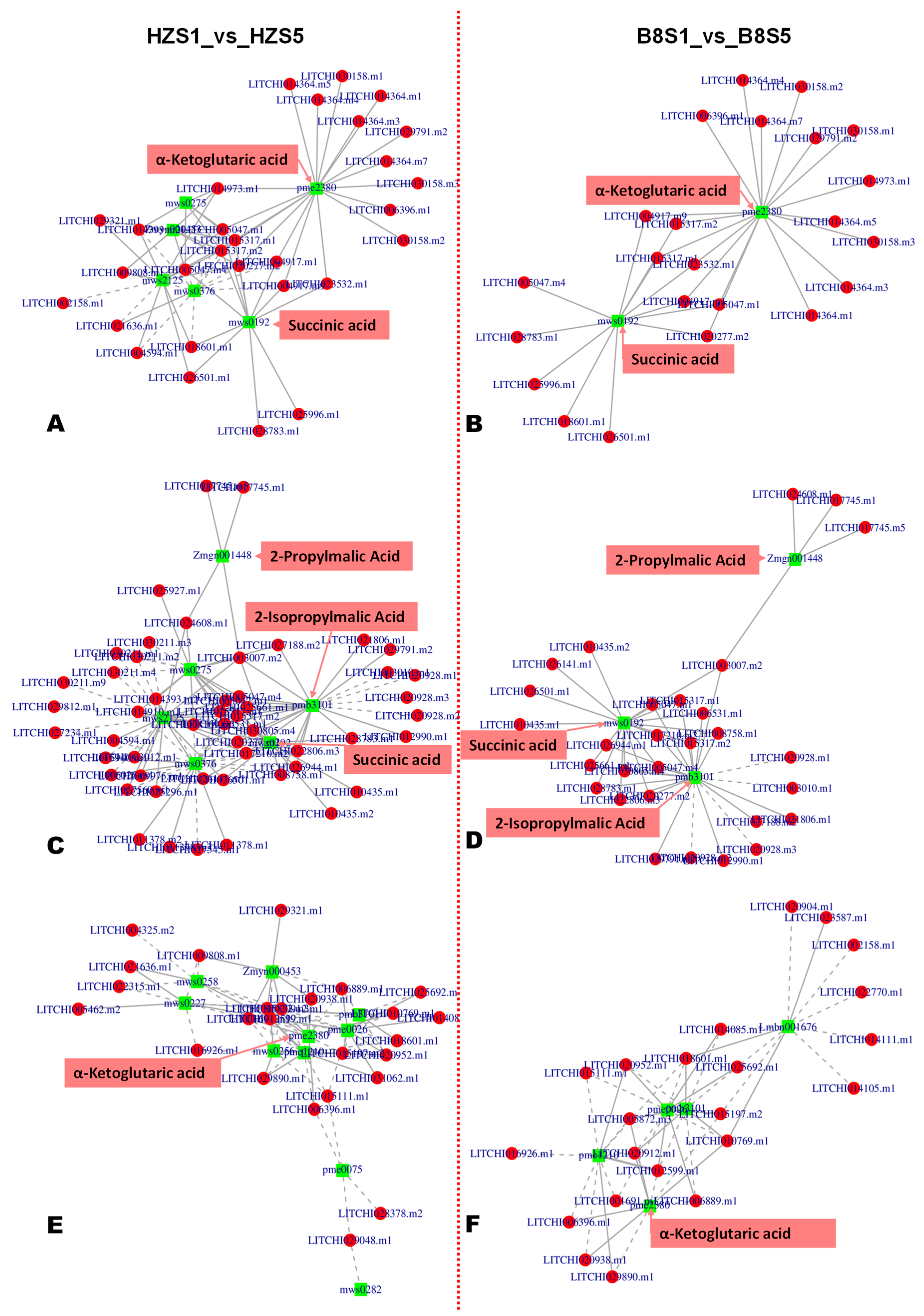
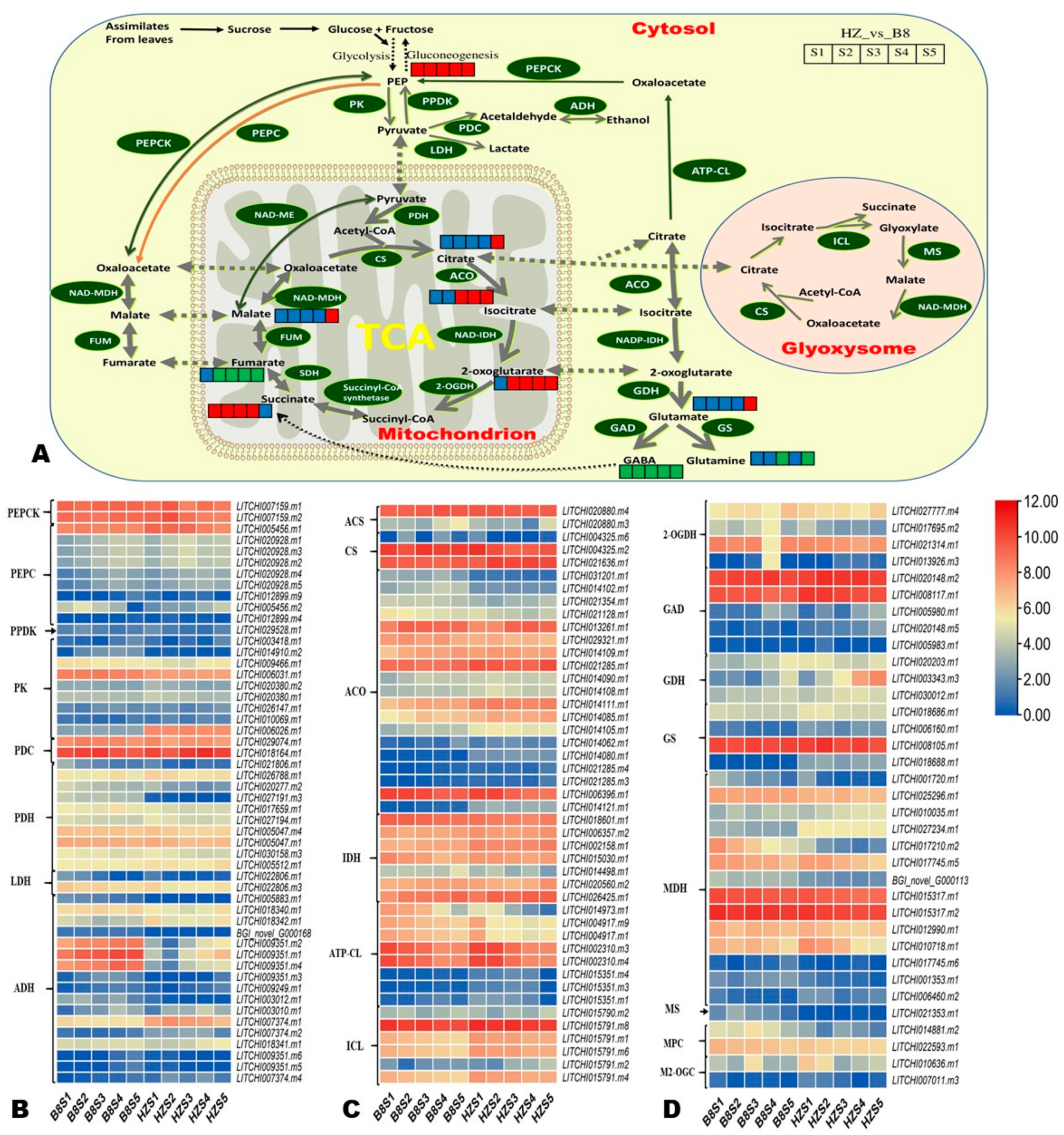
2.6. Candidate Genes for Controlling Litchi Fruit Acidity
2.7. Quantitative Real-Time-PCR (qRT-PCR) Validation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Sampling
4.2. HPLC Analysis of Organic Acids
4.3. Metabolomics Analysis
4.4. Transcriptomics Sequencing
4.5. Differentially Expressed Genes (DEGs) Analysis and Functional Annotations
4.6. qRT-PCR Analysis
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Veerappan, K.; Natarajan, S.; Chung, H.; Park, J. Molecular insights of fruit quality traits in peaches, prunus persica. Plants 2021, 10, 2191. [Google Scholar] [CrossRef]
- Bai, Y.; Dougherty, L.; Cheng, L.; Zhong, G.Y.; Xu, K. Uncovering co-expression gene network modules regulating fruit acidity in diverse apples. BMC Genom. 2015, 16, 612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Zhao, L.; Jiang, X.; Cherono, S.; Liu, J.J.; Ogutu, C.; Ntini, C.; Zhang, X.; Han, Y. Assessment of organic acid accumulation and its related genes in peach. Food Chem. 2021, 334, 127567. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wei, X.; Ali, M.M.; Rizwan, H.M.; Li, B.; Li, H.; Jia, K.; Yang, X.; Ma, S.; Li, S.; et al. Changes in the content of organic acids and expression analysis of citric acid accumulation-related genes during fruit development of yellow (Passiflora edulis f. flavicarpa) and purple (Passiflora edulis f. edulis) passion fruits. Int. J. Mol. Sci. 2021, 22, 5765. [Google Scholar] [CrossRef] [PubMed]
- Etienne, A.; Génard, M.; Lobit, P.; Mbeguié-A-Mbéguié, D.; Bugaud, C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, R.P.; Famiani, F. Organic acids in fruits: Metabolism, functions and contents. Hortic. Rev. 2018, 45, 371–430. [Google Scholar] [CrossRef]
- Saradhuldhat, P.; Paull, R.E. Pineapple organic acid metabolism and accumulation during fruit development. Sci. Hortic. 2007, 112, 297–303. [Google Scholar] [CrossRef]
- García-Gómez, B.E.; Salazar, J.A.; Nicolás-Almansa, M.; Razi, M.; Rubio, M.; Ruiz, D.; Martínez-Gómez, P. Molecular bases of fruit quality in prunus species: An integrated genomic, transcriptomic, and metabolic review with a breeding perspective. Int. J. Mol. Sci. 2021, 22, 333. [Google Scholar] [CrossRef]
- Etienne, A.; Génard, M.; Lobit, P.; Bugaud, C. Modeling the vacuolar storage of malate shed lights on pre- and post-harvest fruit acidity. BMC Plant Biol. 2014, 14, 310. [Google Scholar] [CrossRef]
- Famiani, F.; Farinelli, D.; Moscatello, S.; Battistelli, A.; Leegood, R.C.; Walker, R.P. The contribution of stored malate and citrate to the substrate requirements of metabolism of ripening peach (Prunus persica L. Batsch) flesh is negligible. Implications for the occurrence of phosphoenolpyruvate carboxykinase and gluconeogenesis. Plant Physiol. Biochem. 2016, 101, 33–42. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Tyerman, S.D.; Gilliham, M.; Xu, B. γ-Aminobutyric acid (GABA) signalling in plants. Cell. Mol. Life Sci. 2017, 74, 1577–1603. [Google Scholar] [CrossRef] [PubMed]
- Gilliham, M.; Tyerman, S.D. Linking Metabolism to Membrane Signaling: The GABA–Malate Connection. Trends Plant Sci. 2016, 21, 295–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubimendi, J.P.; Martinatto, A.; Valacco, M.P.; Moreno, S.; Andreo, C.S.; Drincovich, M.F.; Tronconi, M.A. The complex allosteric and redox regulation of the fumarate hydratase and malate dehydratase reactions of Arabidopsis thaliana Fumarase 1 and 2 gives clues for understanding the massive accumulation of fumarate. FEBS J. 2018, 285, 2205–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, C.L.; Andreo, C.S.; Drincovich, M.F.; Gerrard Wheeler, M.C. Fumarate and cytosolic pH as modulators of the synthesis or consumption of C4 organic acids through NADP-malic enzyme in Arabidopsis thaliana. Plant Mol. Biol. 2013, 81, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Castro, A.I.; Oidor-Chan, V.H.; Bustamante-Camilo, P.; Pelayo-Zaldívar, C.; Díaz De León-Sánchez, F.; Mendoza-Espinoza, J.A. Chemical Characterization and Evaluation of the Antihyperglycemic Effect of Lychee (Litchi chinensis Sonn.) cv. Brewster. J. Med. Food 2022, 25, 61–69. [Google Scholar] [CrossRef]
- Bhushan, B.; Pal, A.; Narwal, R.; Meena, V.S.; Sharma, P.C.; Singh, J. Combinatorial approaches for controlling pericarp browning in Litchi (Litchi chinensis) fruit. J. Food Sci. Technol. 2015, 52, 5418–5426. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Gao, H.; Chen, Z.; Li, T.; Zhang, Z.; Yun, Z.; Jiang, Y. Metabolic variations in the pulp of four litchi cultivars during pulp breakdown. Food Res. Int. 2021, 140, 110080. [Google Scholar] [CrossRef]
- Yao, P.; Gao, Y.; Simal-Gandara, J.; Farag, M.A.; Chen, W.; Yao, D.; Delmas, D.; Chen, Z.; Liu, K.; Hu, H.; et al. Litchi (Litchi chinensis Sonn.): A comprehensive review of phytochemistry, medicinal properties, and product development. Food Funct. 2021, 12, 9527–9548. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, K.; Wang, K.; Zhu, J.; Hu, Z. Nutrient components, health benefits, and safety of litchi (Litchi chinensis Sonn.): A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2139–2163. [Google Scholar] [CrossRef]
- Fan, S.; Wang, D.; Xie, H.; Wang, H.; Qin, Y.; Hu, G.; Zhao, J. Sugar transport, metabolism and signaling in fruit development of Litchi chinensis sonn.: A review. Int. J. Mol. Sci. 2021, 22, 11231. [Google Scholar] [CrossRef]
- Wang, H.C.; Huang, H.B.; Huang, X.M.; Hu, Z.Q. Sugar and acid compositions in the arils of Litchi chinensis Sonn.: Cultivar differences and evidence for the absence of succinic acid. J. Hortic. Sci. Biotechnol. 2006, 81, 57–62. [Google Scholar] [CrossRef]
- Liao, H.-z.; Lin, X.-k.; Du, J.-j.; Peng, J.-j.; Zhou, K.-b. Transcriptomic analysis reveals key genes regulating organic acid synthesis and accumulation in the pulp of Litchi chinensis Sonn. cv. Feizixiao. Sci. Hortic. 2022, 303, 111220. [Google Scholar] [CrossRef]
- Sweetlove, L.J.; Beard, K.F.M.; Nunes-Nesi, A.; Fernie, A.R.; Ratcliffe, R.G. Not just a circle: Flux modes in the plant TCA cycle. Trends Plant Sci. 2010, 15, 462–470. [Google Scholar] [CrossRef]
- Sweetman, C.; Deluc, L.G.; Cramer, G.R.; Ford, C.M.; Soole, K.L. Regulation of malate metabolism in grape berry and other developing fruits. Phytochemistry 2009, 70, 1329–1344. [Google Scholar] [CrossRef] [PubMed]
- Byeon, S.E.; Lee, J. Fruit maturity differentially affects fruit quality and responses of targeted metabolites in cold-stored figs (Ficus carica L.). J. Sci. Food Agric. 2021, 101, 673–683. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Yang, W.; Li, J.; Tang, W.; Gong, R. Transcriptomic and Metabolomic Analysis of Quality Changes during Sweet Cherry Fruit Development and Mining of Related Genes. Int. J. Mol. Sci. 2022, 23, 7402. [Google Scholar] [CrossRef]
- Dong, Y.; Østergaard, L. Fruit development and diversification. Curr. Biol. 2019, 29, R781–R785. [Google Scholar] [CrossRef]
- Lin, Q.; Wang, C.; Dong, W.; Jiang, Q.; Wang, D.; Li, S.; Chen, M.; Liu, C.; Sun, C.; Chen, K. Transcriptome and metabolome analyses of sugar and organic acid metabolism in Ponkan (Citrus reticulata) fruit during fruit maturation. Gene 2015, 554, 64–74. [Google Scholar] [CrossRef]
- Walker, R.P.; Chen, Z.H.; Famiani, F. Gluconeogenesis in Plants: A Key Interface between Organic Acid/Amino Acid/Lipid and Sugar Metabolism. Molecules 2021, 26, 5129. [Google Scholar] [CrossRef]
- Fromm, H. GABA signaling in plants: Targeting the missing pieces of the puzzle. J. Exp. Bot. 2021, 71, 638–6245. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Tyerman, S.D.; Xu, B.; Bose, J.; Kaur, S.; Conn, V.; Domingos, P.; Ullah, S.; Wege, S.; Shabala, S.; et al. GABA signalling modulates plant growth by directly regulating the activity of plant-specific anion transporters. Nat. Commun. 2015, 6, 7879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, F.; Tercé-Laforgue, T.; Gonzalez-Moro, M.B.; Estavillo, J.M.; Sangwan, R.; Gallais, A.; Hirel, B. Glutamate dehydrogenase in plants: Is there a new story for an old enzyme? Plant Physiol. Biochem. 2003, 41, 565–576. [Google Scholar] [CrossRef]
- Xu, B.; Sai, N.; Gilliham, M. The emerging role of GABA as a transport regulator and physiological signal. Plant Physiol. 2021, 187, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Niu, X.Q.; Zheng, X.L.; Chen, X.; Zheng, G.H.; Wu, J.C. Comparative transcriptome analysis reveals key genes potentially related to organic acid and sugar accumulation in loquat. PLoS ONE 2021, 16, e0238873. [Google Scholar] [CrossRef] [PubMed]
- Chia, D.W.; Yoder, T.J.; Reiter, W.D.; Gibson, S.I. Fumaric acid: An overlooked form of fixed carbon in Arabidopsis and other plant species. Planta 2000, 211, 743–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, W.L.; Nunes-Nesi, A.; Fernie, A.R. Fumarate: Multiple functions of a simple metabolite. Phytochemistry 2011, 72, 838–843. [Google Scholar] [CrossRef]
- Ma, B.; Chen, J.; Zheng, H.; Fang, T.; Ogutu, C.; Li, S.; Han, Y.; Wu, B. Comparative assessment of sugar and malic acid composition in cultivated and wild apples. Food Chem. 2015, 172, 86–91. [Google Scholar] [CrossRef]
- Dossou, S.S.K.; Xu, F.; You, J.; Zhou, R.; Li, D.; Wang, L. Widely targeted metabolome profiling of different colored sesame (Sesamum indicum L.) seeds provides new insight into their antioxidant activities. Food Res. Int. 2022, 151, 110850. [Google Scholar] [CrossRef]
- Yi, D.; Zhang, H.; Lai, B.; Liu, L.; Pan, X.; Ma, Z.; Wang, Y.; Xie, J.; Shi, S.; Wei, Y. Integrative Analysis of the Coloring Mechanism of Red Longan Pericarp through Metabolome and Transcriptome Analyses. J. Agric. Food Chem. 2021, 69, 1806–1815. [Google Scholar] [CrossRef]
- Wang, J.G.; Gao, X.M.; Ma, Z.L.; Chen, J.; Liu, Y.N.; Shi, W.Q. Metabolomic and transcriptomic profiling of three types of litchi pericarps reveals that changes in the hormone balance constitute the molecular basis of the fruit cracking susceptibility of Litchi chinensis cv. Baitangying. Mol. Biol. Rep. 2019, 46, 5295–5308. [Google Scholar] [CrossRef]
- Hu, G.; Feng, J.; Xiang, X.; Wang, J.; Salojärvi, J.; Liu, C.; Wu, Z.; Zhang, J.; Liang, X.; Jiang, Z.; et al. Two divergent haplotypes from a highly heterozygous lychee genome suggest independent domestication events for early and late-maturing cultivars. Nat. Genet. 2022, 54, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene_ID | Chr | Gene Length | Log2FC_B8-vs.-HZ | Annotation | ||||
|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S3 | S4 | S5 | ||||
| LITCHI014080.m1 | Chr2 | 8049 | 4.06062 | 4.07972 | 3.57043 | 3.50206 | 4.81101 | Aconitate hydratase |
| LITCHI014121.m1 | Chr2 | 2381 | 4.21622 | 5.38847 | 3.22051 | 3.18061 | 3.40981 | |
| LITCHI005883.m1 | Chr14 | 10,553 | −5.7443 | −5.6668 | −4.9752 | −3.4136 | −2.6497 | Alcohol dehydrogenase |
| LITCHI009351.m2 | Chr7 | 2030 | −4.3612 | −8.2589 | −4.911 | −3.743 | −3.1405 | |
| LITCHI009351.m1 | Chr7 | 2717 | −4.7887 | −8.2142 | −4.8992 | −3.9994 | −3.1553 | |
| LITCHI009351.m4 | Chr7 | 1613 | −5.0403 | −5.6369 | −4.4061 | −4.5642 | −3.8755 | |
| LITCHI014973.m1 | Chr1 | 7813 | −2.3199 | −1.8379 | −3.1671 | −2.699 | −2.8939 | ATP citrate (pro-S)-lyase |
| LITCHI015351.m4 | Chr1 | 2941 | 3.24125 | 2.79842 | 3.06652 | 2.04997 | −1.1406 | |
| LITCHI004325.m6 | Chr14 | 5042 | 2.57725 | −4.5858 | NA | −1.2316 | −0.1214 | Citrate synthase |
| LITCHI020148.m5 | Chr12 | 7582 | 0.84976 | 2.50526 | 1.33812 | 1.30628 | 3.19904 | Glutamate decarboxylase |
| LITCHI003343.m3 | Chr6 | 2915 | 0.28903 | 2.36364 | 4.60759 | 4.35457 | 3.36485 | Glutamate dehydrogenase (NAD(P)+) |
| LITCHI014417.m1 | Chr1 | 766 | −7.4384 | −7.6799 | −7.6302 | −5.9068 | −4.2463 | Glutamate receptor, ionotropic, plant |
| LITCHI018688.m1 | Chr15 | 10,459 | 7.74389 | 6.88227 | 3.17553 | 6.89497 | 6.98392 | Glutamine synthetase |
| LITCHI017210.m2 | Chr1 | 4944 | −3.8161 | −4.6483 | −4.8736 | −5.3081 | −3.0973 | Malate dehydrogenase (oxaloacetate-decarboxylating) (NADP+) |
| LITCHI017210.m4 | Chr1 | 3658 | 5.80302 | 3.57723 | 3.80498 | −1.4727 | −0.0223 | |
| LITCHI017210.m3 | Chr1 | 3658 | 5.80302 | 3.57723 | 3.80498 | −1.4727 | −0.0223 | |
| LITCHI006460.m2 | Chr11 | 3827 | 1.20331 | 2.45027 | 0.9708 | 2.73029 | 3.01809 | Malate dehydrogenase |
| LITCHI021353.m1 | Chr12 | 1611 | −5.6461 | −7.5779 | −6.5181 | −3.9782 | −2.512 | Malate synthase |
| LITCHI014881.m2 | Chr1 | 685 | −1.8423 | −3.6235 | −2.9322 | −3.2504 | −2.7075 | Mitochondrial pyruvate carrier 2 |
| LITCHI012899.m9 | Chr2 | 2105 | 2.28868 | NA | 4.47104 | 0.66693 | −5.3621 | Phosphoenolpyruvate carboxylase |
| LITCHI005456.m2 | Chr14 | 15,180 | −1.1193 | −1.4388 | −0.656 | 0.06627 | 7.75064 | |
| LITCHI027191.m3 | Chr3 | 7665 | −5.1948 | −6.1722 | −7.9115 | −8.4677 | −8.1616 | Pyruvate dehydrogenase |
| LITCHI027947.m4 | Chr3 | 3598 | −2.4055 | −1.1367 | −0.0642 | −3.5808 | −2.4764 | Pyruvate dehydrogenase phosphatase |
| LITCHI014910.m2 | Chr1 | 4544 | 0.58152 | −2.1858 | −4.5882 | −7.1461 | −4.1166 | Pyruvate kinase |
| LITCHI006026.m1 | Chr14 | 7987 | 5.73614 | 5.63149 | 5.63661 | 5.5364 | 5.40993 | |
| LITCHI010636.m1 | Chr8 | 975 | 2.35935 | 2.91427 | −2.8211 | −1.5014 | 1.12691 | Solute carrier family 25 (mitochondrial oxoglutarate transporter) |
| LITCHI007011.m3 | Chr11 | 2142 | 0.83409 | 3.09493 | 0.71148 | −1.2857 | 2.30272 | |
| LITCHI013926.m3 | Chr2 | 3341 | NA | NA | 0.4415 | 3.81511 | 2.8162 | 2-oxoglutarate dehydrogenase |
| BGI_novel_G000750 | Chr4 | 6972 | 4.11965 | 1.87247 | 1.78248 | 2.097 | 0.82182 | Succinyl-CoA synthetase |
| LITCHI026501.m2 | Chr3 | 5631 | 3.79998 | 7.99249 | 7.43426 | 7.79962 | 7.75682 | Fumarate hydratase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Qi, Y.; Chen, X.; Yan, Q.; Chen, J.; Liu, H.; Shi, F.; Wen, Y.; Cai, C.; Ou, L. Combined Metabolome and Transcriptome Analyses Unveil the Molecular Mechanisms of Fruit Acidity Variation in Litchi (Litchi chinensis Sonn.). Int. J. Mol. Sci. 2023, 24, 1871. https://doi.org/10.3390/ijms24031871
Jiang Y, Qi Y, Chen X, Yan Q, Chen J, Liu H, Shi F, Wen Y, Cai C, Ou L. Combined Metabolome and Transcriptome Analyses Unveil the Molecular Mechanisms of Fruit Acidity Variation in Litchi (Litchi chinensis Sonn.). International Journal of Molecular Sciences. 2023; 24(3):1871. https://doi.org/10.3390/ijms24031871
Chicago/Turabian StyleJiang, Yonghua, Yingwei Qi, Xilong Chen, Qian Yan, Jiezhen Chen, Hailun Liu, Fachao Shi, Yingjie Wen, Changhe Cai, and Liangxi Ou. 2023. "Combined Metabolome and Transcriptome Analyses Unveil the Molecular Mechanisms of Fruit Acidity Variation in Litchi (Litchi chinensis Sonn.)" International Journal of Molecular Sciences 24, no. 3: 1871. https://doi.org/10.3390/ijms24031871