
Identification and Expression Analysis of Dsx and Its Positive Transcriptional Regulation of IAG in Black Tiger Shrimp (Penaeus monodon)

 ,
,
Abstract
:1. Introduction
2. Results
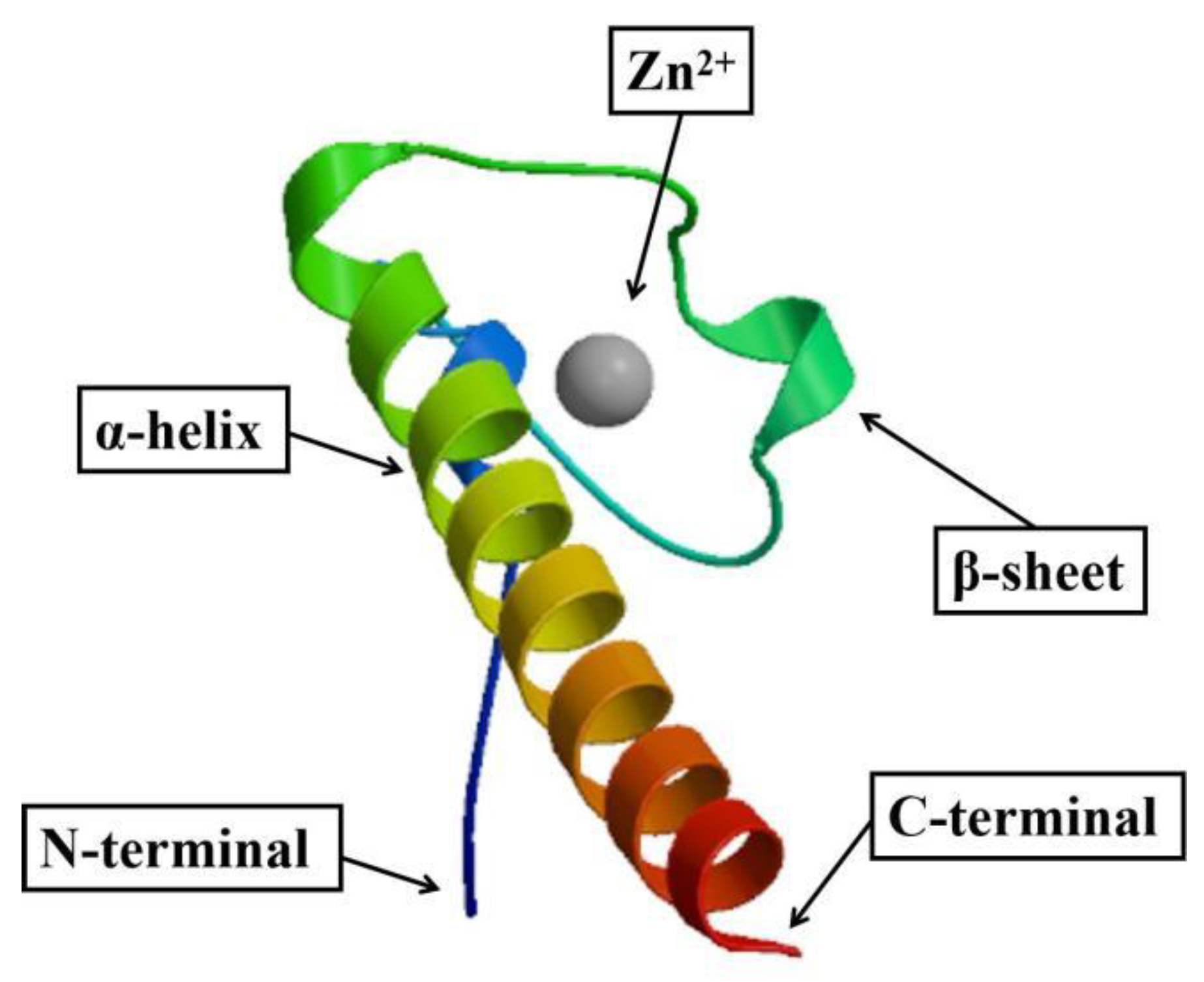
2.1. cDNA Cloning and Sequence Analysis of PmDsx
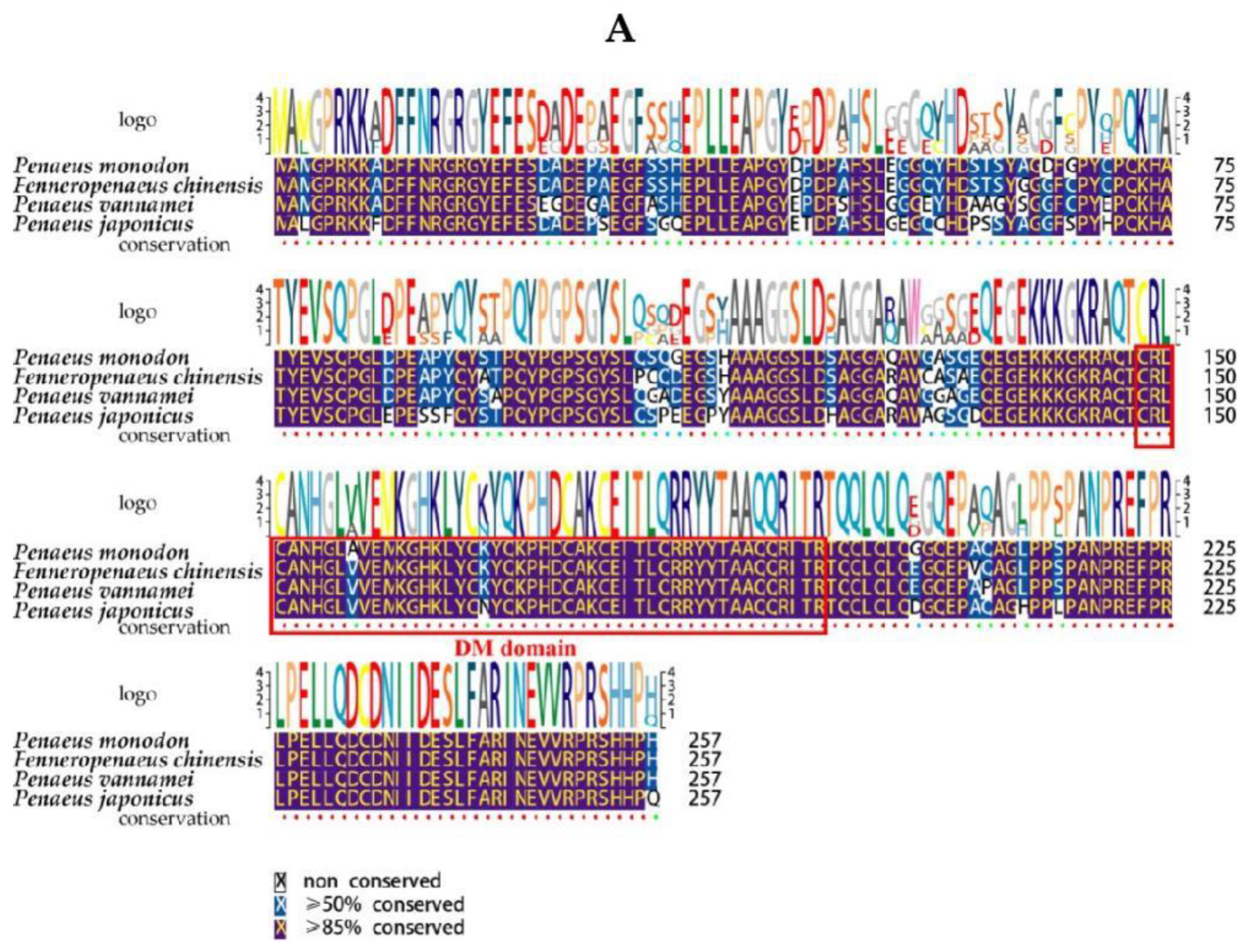
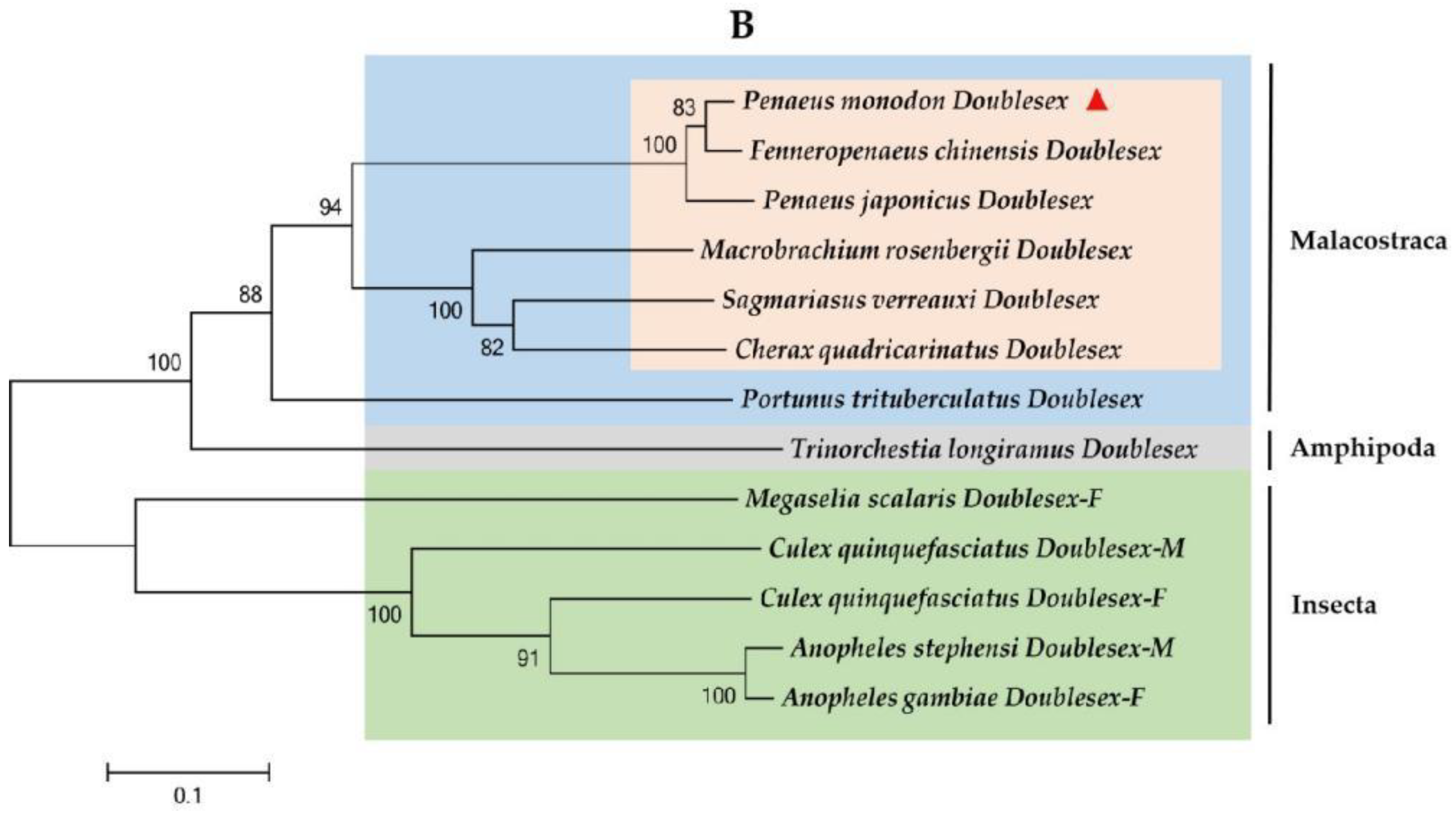
2.2. Multiple Sequence Alignment and Phylogenetic Analysis of PmDsx
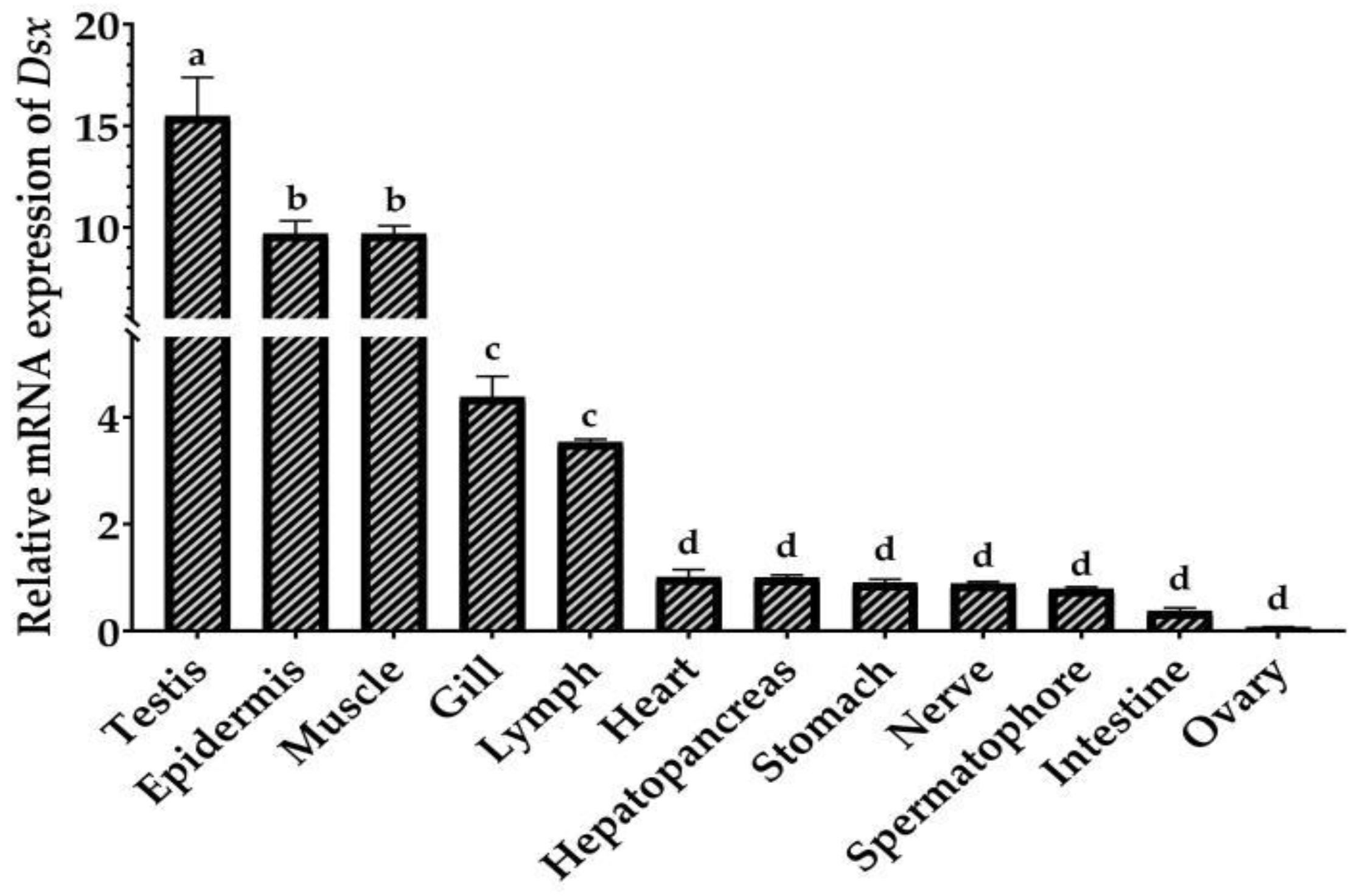
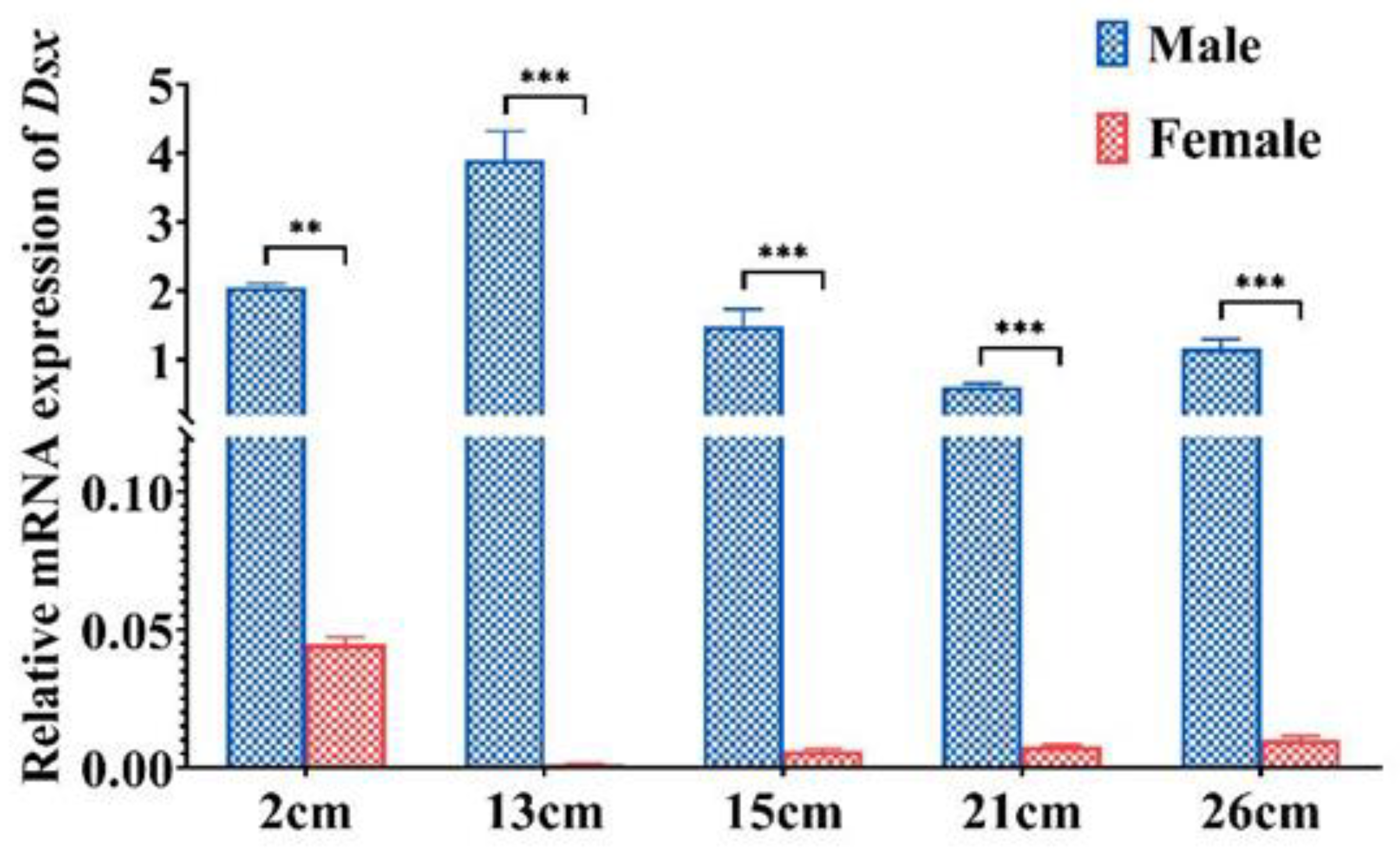
2.3. Tissue Distribution and Sexually Dimorphic Expression of PmDsx
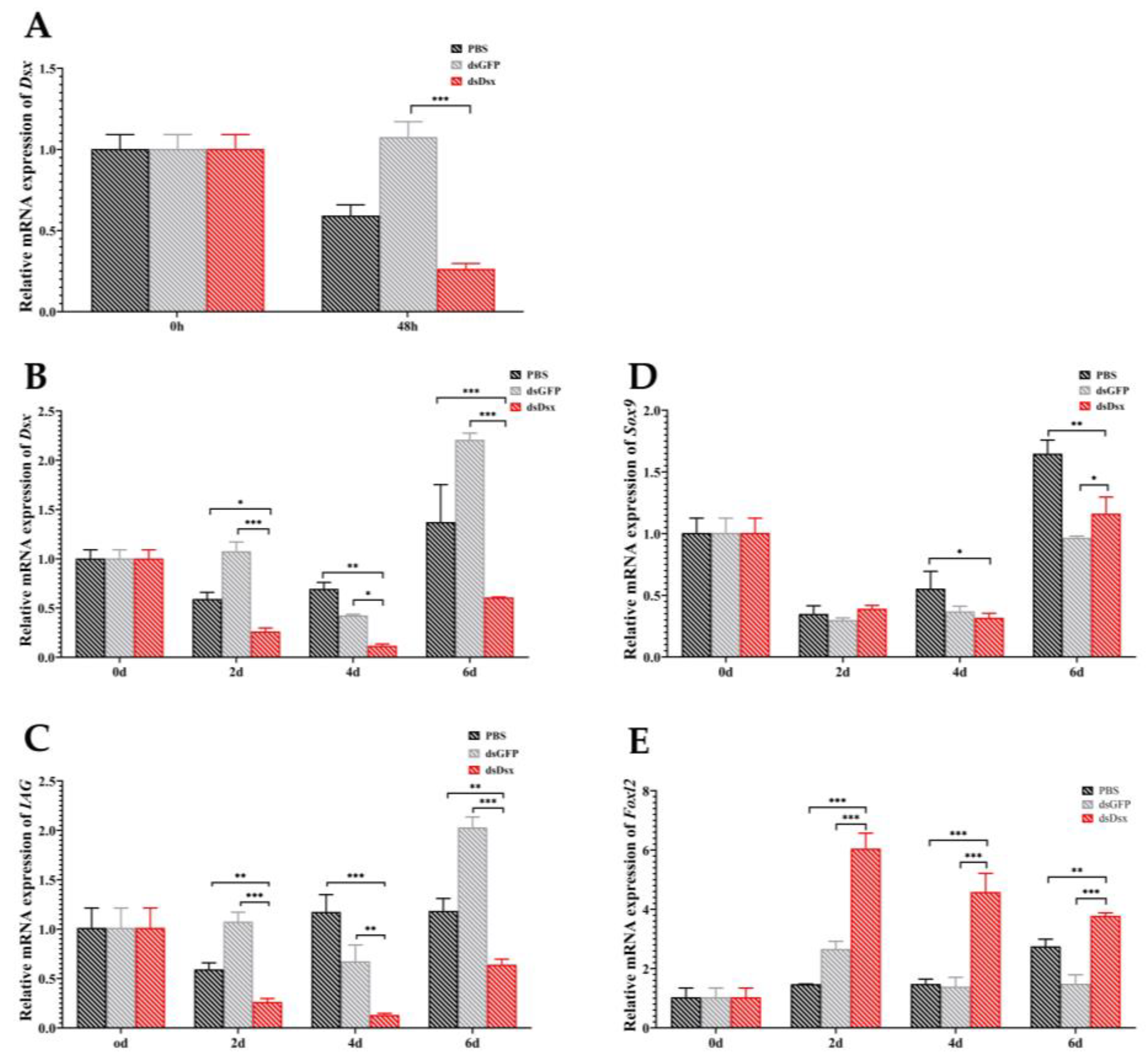
2.4. RNA Interference of PmDsx Reduces the Transcript Level of Sex-Related Genes
2.5. Prediction of Transcription Factor Binding Sites in the Promoter Region of PmIAG
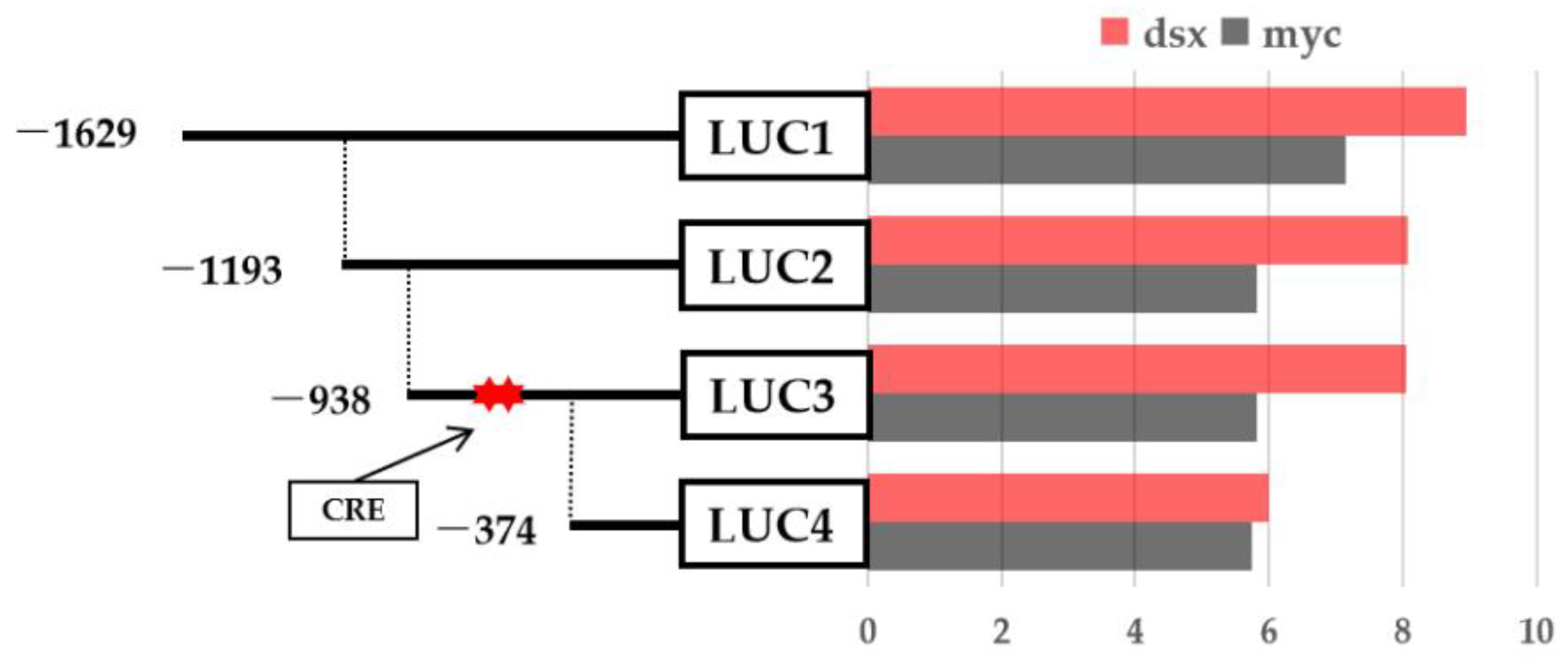
2.6. PmDsx Regulates the Transcriptional Activity of PmIAG
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Cell Culture
4.2. RNA Isolation and Molecular Cloning
4.3. Bioinformatics Analysis of PmDsx
4.4. Real-Time Quantitative PCR Analysis
4.5. RNA Interference
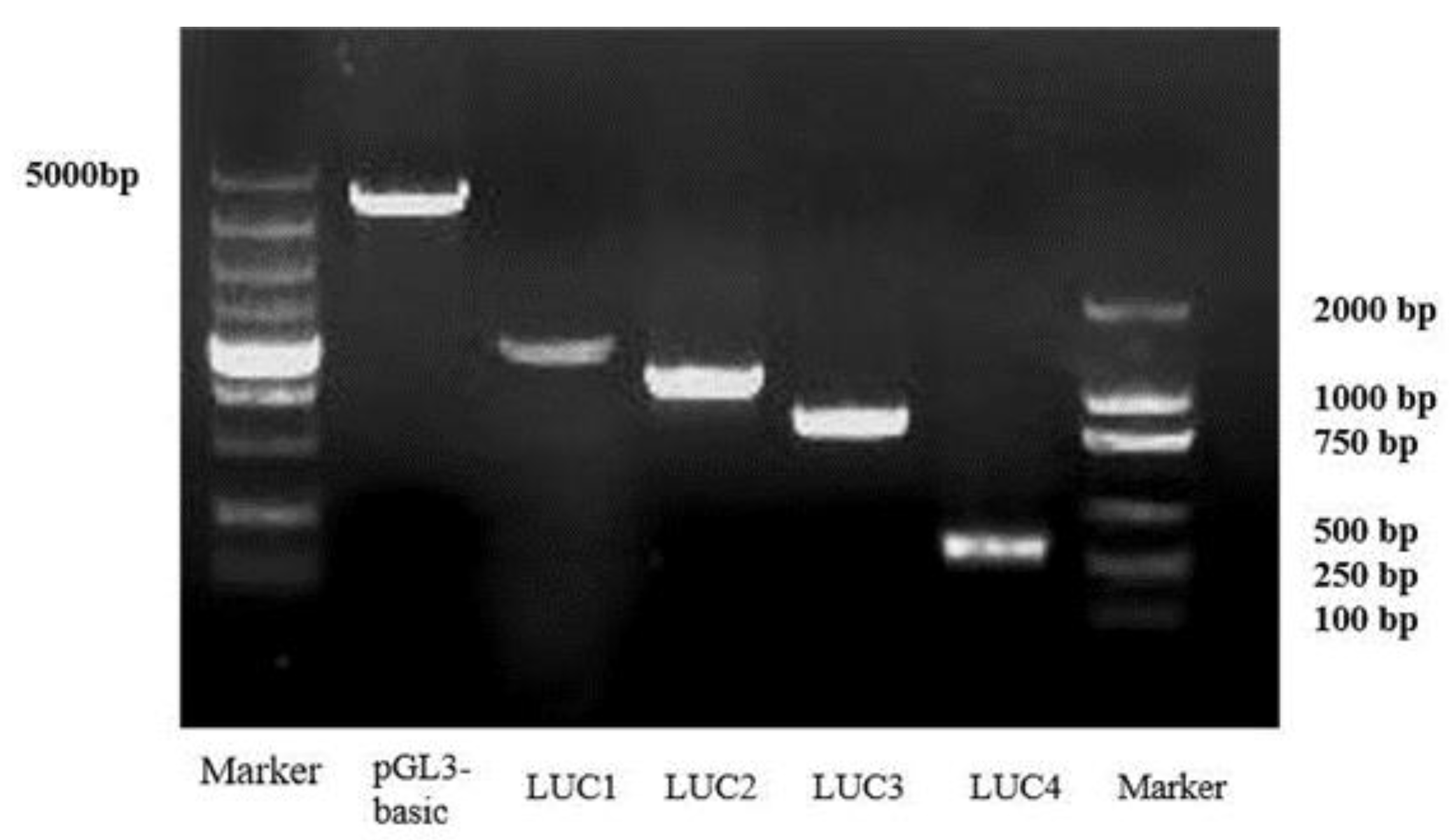
4.6. Promoter Cloning and Plasmid Construction
4.7. Dual-Luciferase Reporter Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cronin, L.E. Anatomy and histology of the male reproductive system of Callinectes sapidus Rathbun. J. Morphol. 1947, 81, 209–239. [Google Scholar] [CrossRef]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Raviv, S.; Glazer, L.; Sagi, A. Temporal silencing of an androgenic gland-specific insulin-like gene affecting phenotypical gender differences and spermatogenesis. Endocrinology 2009, 150, 1278–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, T.; Rosen, O.; Sagi, A. From the discovery of the crustacean androgenic gland to the insulin-like hormone in six decades. Gen. Comp. Endocrinol. 2011, 173, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.Y.; Lin, J.Y.; Guo, S.Z.; Chen, Y.; Li, J.-L.; Qiu, G.F. Molecular characterization and expression analysis of an insulin-like gene from the androgenic gland of the oriental river prawn, Macrobrachium nipponense. Gen. Comp. Endocrinol. 2013, 185, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Bai, H.; Xiong, Y.; Fu, H.; Jiang, S.; Jiang, F.; Jin, S.; Sun, S.; Qiao, H.; Zhang, W. Molecular characterization of insulin-like androgenic gland hormone-binding protein gene from the oriental river prawn Macrobrachium nipponense and investigation of its transcriptional relationship with the insulin-like androgenic gland hormone gene. Gen. Comp. Endocrinol. 2015, 216, 152–160. [Google Scholar] [CrossRef]
- Mareddy, V.R.; Rosen, O.; Thaggard, H.B.; Manor, R.; Kuballa, A.V.; Aflalo, E.D.; Sagi, A.; Paterson, B.; Elizur, A. Isolation and characterization of the complete cDNA sequence encoding a putative insulin-like peptide from the androgenic gland of Penaeus monodon. Aquaculture 2011, 318, 364–370. [Google Scholar] [CrossRef]
- Li, S.; Li, F.; Sun, Z.; Xiang, J. Two spliced variants of insulin-like androgenic gland hormone gene in the Chinese shrimp, Fenneropenaeus chinensis. Gen. Comp. Endocrinol. 2012, 177, 246–255. [Google Scholar] [CrossRef]
- Sroyraya, M.; Chotwiwatthanakun, C.; Stewart, M.J.; Soonklang, N.; Kornthong, N.; Phoungpetchara, I.; Hanna, P.J.; Sobhon, P. Bilateral eyestalk ablation of the blue swimmer crab, Portunus pelagicus, produces hypertrophy of the androgenic gland and an increase of cells producing insulin-like androgenic gland hormone. Tissue Cell 2010, 42, 293–300. [Google Scholar] [CrossRef]
- Jin, S.; Fu, Y.; Hu, Y.; Fu, H.; Jiang, S.; Xiong, Y.; Qiao, H.; Zhang, W.; Gong, Y.; Wu, Y. Identification of candidate genes from androgenic gland in Macrobrachium nipponense regulated by eyestalk ablation. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Liu, F.; Shi, W.; Ye, H.; Zeng, C.; Zhu, Z. Insulin-like androgenic gland hormone 1 (IAG1) regulates sexual differentiation in a hermaphrodite shrimp through feedback to neuroendocrine factors. Gen. Comp. Endocrinol. 2021, 303, 113706. [Google Scholar] [CrossRef]
- Tan, K.; Zhou, M.; Jiang, H.; Jiang, D.; Li, Y.; Wang, W. siRNA-Mediated MrIAG Silencing Induces Sex Reversal in Macrobrachium rosenbergii. Mar. Biotechnol. 2020, 22, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Rosen, O.; Sagi, A. Timing sexual differentiation: Full functional sex reversal achieved through silencing of a single insulin-like gene in the prawn, Macrobrachium rosenbergii. Biol. Reprod. 2012, 86, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Li, F.; Wang, L.; Wu, F.; Wang, J.; Fan, X.; Liu, T. Molecular characteristics and abundance of insulin-like androgenic gland hormone and effects of RNA interference in Eriocheir sinensis. Anim. Reprod. Sci. 2020, 215, 106332. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Lu, B.; Wang, G.; Ye, H. Transcriptional inhibition of sp-iag by crustacean female sex hormone in the mud crab, scylla paramamosain. Int. J. Mol. Sci. 2020, 21, 5300. [Google Scholar] [CrossRef]
- Burtis, K.C.; Baker, B.S. Drosophila doublesex gene controls somatic sexual differentiation by producing alternatively spliced mRNAs encoding related sex-specific polypeptides. Cell 1989, 56, 997–1010. [Google Scholar] [CrossRef]
- Ruiz, M.F.; Alvarez, M.; Eirín-López, J.M.; Sarno, F.; Kremer, L.; Barbero, J.L.; Sánchez, L. An unusual role for doublesex in sex determination in the dipteran Sciara. Genetics 2015, 200, 1181–1199. [Google Scholar] [CrossRef] [Green Version]
- Shirangi, T.R.; Wong, A.M.; Truman, J.W.; Stern, D.L. Doublesex Regulates the Connectivity of a Neural Circuit Controlling Drosophila Male Courtship Song. Dev. Cell 2016, 37, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhan, S.; Chen, S.; Zeng, B.; Li, Z.; James, A.A.; Tan, A.; Huang, Y. Sexually dimorphic traits in the silkworm, Bombyx mori, are regulated by doublesex. Insect Biochem. Mol. Biol. 2017, 80, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Rideout, E.J.; Dornan, A.J.; Neville, M.C.; Eadie, S.; Goodwin, S.F. Control of sexual differentiation and behavior by the doublesex gene in Drosophila melanogaster. Nat. Neurosci. 2010, 13, 458–466. [Google Scholar] [CrossRef] [Green Version]
- Coschigano, K.T.; Wensink, P.C. Sex-specific transcriptional regulation by the male and female doublesex proteins of Drosophila. Genes Dev. 1993, 7, 42–54. [Google Scholar] [CrossRef]
- Xu, J.; Wang, Y.; Li, Z.; Ling, L.; Zeng, B.; James, A.A.; Tan, A.; Huang, Y. Transcription activator-like effector nuclease (TALEN)-mediated female-specific sterility in the silkworm, Bombyx mori. Insect Mol. Biol. 2014, 23, 800–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Kobayashi, K.; Watanabe, H.; Iguchi, T. Environmental sex determination in the branchiopod crustacean Daphnia magna: Deep conservation of a Doublesex gene in the sex-determining pathway. PLoS Genet. 2011, 7, e1001345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, P.; Zhou, T.; Zhang, Y.; Chen, Y.; Yi, J.; Lin, W.; Guo, Z.; Xu, A.; Yang, S.; Chan, S. Potential involvement of a DMRT family member (Mr-Dsx) in the regulation of sexual differentiation and moulting in the giant river prawn Macrobrachium rosenbergii. Aquac. Res. 2019, 50, 3037–3049. [Google Scholar] [CrossRef]
- Yu, Y.Q.; Ma, W.M.; Zeng, Q.G.; Qian, Y.Q.; Yang, J.S.; Yang, W.J. Molecular Cloning and Sexually Dimorphic Expression of Two Dmrt Genes in the Giant Freshwater Prawn, Macrobrachium rosenbergii. Agric. Res. 2014, 3, 181–191. [Google Scholar] [CrossRef]
- Li, S.; Li, F.; Yu, K.; Xiang, J. Identification and characterization of a doublesex gene which regulates the expression of insulin-like androgenic gland hormone in Fenneropenaeus chinensis. Gene 2018, 649, 1–7. [Google Scholar] [CrossRef]
- Zhikang, H.; Shigui, J.; Falin, Z.; Jianhua, H.; Qibin, Y. Establishment of a sex identification method for black tiger shrimp (Penaeus monodon) in embryonic and early postembryonic development based on an InDel molecular marker. S. China Fish. Sci. 2020, 16. [Google Scholar] [CrossRef]
- Zhu, L.; Wilken, J.; Phillips, N.B.; Narendra, U.; Chan, G.; Stratton, S.M.; Kent, S.B.; Weiss, M.A. Sexual dimorphism in diverse metazoans is regulated by a novel class of intertwined zinc fingers. Genes Dev. 2000, 14, 1750–1764. [Google Scholar] [CrossRef]
- Bayrer, J.R.; Zhang, W.; Weiss, M.A. Dimerization of doublesex is mediated by a cryptic ubiquitin-associated domain fold: Implications for sex-specific gene regulation. J. Biol. Chem. 2005, 280, 32989–32996. [Google Scholar] [CrossRef] [Green Version]
- Boggs, R.T.; Gregor, P.; Idriss, S.; Belote, J.M.; McKeown, M. Regulation of sexual differentiation in D. melanogaster via alternative splicing of RNA from the transformer gene. Cell 1987, 50, 739–747. [Google Scholar] [CrossRef]
- Inoue, K.; Hoshijima, K.; Higuchi, I.; Sakamoto, H.; Shimura, Y. Binding of the Drosophila transformer and transformer-2 proteins to the regulatory elements of doublesex primary transcript for sex-specific RNA processing. Proc. Natl. Acad. Sci. USA 1992, 89, 8092–8096. [Google Scholar] [CrossRef]
- Zhang, Y.; Waiho, K.; Ikhwanuddin, M.; Ma, H. Identification of sex-related genes from the three-spot swimming crab portunus sanguinolentus and comparative analysis with the crucifix crab charybdis feriatus. Animals 2021, 11, 1946. [Google Scholar] [CrossRef]
- Zheng, J.; Cai, L.; Jia, Y.; Chi, M.; Cheng, S.; Liu, S.; Li, F.; Gu, Z. Identification and functional analysis of the doublesex gene in the redclaw crayfish, Cherax quadricarinatus. Gene Expr. Patterns 2020, 37, 119129. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Zhong, J.; Zhang, Z.; Sheng, Y.; Wang, Y. Identification and functional analysis of the doublesex gene in the mud crab Scylla paramamosain. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2022, 266, 111150. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.S.; Uppendahl, L.D.; Chowdhury, M.A.; Ip, P.L.; Siegal, M.L. The female-specific Doublesex isoform regulates pleiotropic transcription factors to pattern genital development in Drosophila. Development 2011, 138, 1099–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.-Y.; Huang, J.-H.; Yang, Q.-B.; Zhou, F.-L.; Jiang, S.; Li, Y.-D.; Jiang, S.-G.; Yang, L.-S. Molecular characterization and functional analysis of DMRT11E in black tiger shrimp (Penaeus monodon). Aquac. Reports J. 2022, 22, 100982. [Google Scholar] [CrossRef]
- Li, M.H.; Yang, H.H.; Li, M.R.; Sun, Y.L.; Jiang, X.L.; Xie, Q.P.; Wang, T.R.; Shi, H.J.; Sun, L.N.; Zhou, L.Y. Antagonistic roles of Dmrt1 and Foxl2 in sex differentiation via estrogen production in tilapia as demonstrated by TALENs. Endocrinology 2013, 154, 4814–4825. [Google Scholar] [CrossRef]
- Wei, L.; Li, X.; Li, M.; Tang, Y.; Wei, J.; Wang, D. Dmrt1 directly regulates the transcription of the testis-biased Sox9b gene in Nile tilapia (Oreochromis niloticus). Gene 2019, 687, 109–115. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, S.; Fu, H.; Qiao, H.; Sun, S.; Zhang, W.; Jiang, S.; Gong, Y.; Xiong, Y.; Wu, Y. Identification and characterization of the DMRT11E gene in the oriental river prawn macrobrachium nipponense. Int. J. Mol. Sci. 2019, 20, 1734. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Lu, Z.; Qin, Z.; Zhao, L.; Pan, G.; Shen, H.; Zhang, M.; Liang, R.; Lin, L.; Zhang, K. Insight into the regulatory relationships between the insulin-like androgenic gland hormone gene and the insulin-like androgenic gland hormone-binding protein gene in giant freshwater prawns (Macrobrachium rosenbergii). Int. J. Mol. Sci. 2020, 21, 4207. [Google Scholar] [CrossRef]
- Chen, K.; Li, S.; Xiang, J.; Sagi, A.; Li, F. Transcriptome analysis reveals the endocrine regulation on the expression of iag in litopenaeus vannamei. J. Mar. Sci. Eng. 2021, 9, 677. [Google Scholar] [CrossRef]
- Shi, L.; Han, S.; Fei, J.; Zhang, L.; Ray, J.W.; Wang, W.; Li, Y. Molecular Characterization and Functional Study of Insulin-Like Androgenic Gland Hormone Gene in the Red Swamp Crayfish, Procambarus clarkii. Genes 2019, 10, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyadarshi, H.; Das, R.; Pavan-Kumar, A.; Gireesh-Babu, P.; Javed, H.; Kumar, S.; Marappan, M.; Somdutt; Krishna, G.; Chaudhari, A. Silencing and augmentation of IAG hormone transcripts in adult Macrobrachium rosenbergii males affects morphotype transformation. J. Exp. Biol. 2017, 220, 4101–4108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.; Liu, Y.; Yuan, J.; Zhang, X.; Ventura, T.; Ma, K.Y.; Sun, S.; Song, C.; Zhan, D.; Yang, Y. The Chinese mitten crab genome provides insights into adaptive plasticity and developmental regulation. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yi, W.; Zarkower, D. Similarity of DNA binding and transcriptional regulation by Caenorhabditis elegans MAB-3 and Drosophila melanogaster DSX suggests conservation of sex determining mechanisms. Development 1999, 126, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence(5′–3′) | Purpose |
|---|---|---|
| PmDsx-F | GTGGAACCTGAGGCAAACG | ORF Validation |
| PmDsx-R | AGACGGCGGAGGAAAAGG | ORF Validation |
| PmDsx-qF | TCCAACGACGCTACTACACGG | qPCR |
| PmDsx-qR | TTGATGCGGGCAAACAGG | qPCR |
| dsPmDsx-T7-F | TAATACGACTCACTATAGGGAGAGTACCAGCCACAGAAGCACG | RNAi |
| dsPmDsx-T7-R | TAATACGACTCACTATAGGGAGAAGACGGCGGAGGAAAAGG | RNAi |
| dsPmDsx-F | GTACCAGCCACAGAAGCACG | RNAi |
| dsPmDsx-R | AGACGGCGGAGGAAAAGG | RNAi |
| dsGFP-T7-F | TAATACGACTCACTATAGGGAGACCGACAAGCAGAAGAACGGCATCA | RNAi |
| dsGFP-T7-R | TAATACGACTCACTATAGGGAGATCACGAACTCCAGCAGGACCATGTGA | RNAi |
| dsGFP-F | CCGACAAGCAGAAGAACGGCATCA | RNAi |
| dsGFP-R | TCACGAACTCCAGCAGGACCATGTGA | RNAi |
| PmSox9-qF | GCGAGGGACTTGGTAAATGTG | qPCR |
| PmSox9-qR | TGGTGGCTAGGATTGGTCTGA | qPCR |
| PmIAG-qF | GCCTTGAATCCGATGCGATAT | qPCR |
| PmIAG-qR | GCAGCACTCATCCTGTACGTTGT | qPCR |
| PmFoxl2-qF | AGGGAAGGGGAATTTCTGG | qPCR |
| PmFoxl2-qR | AAGCGTCGGGGTAGGTGTA | qPCR |
| EF-1α-qF | AAGCCAGGTATGGTTGTCAACTTT | Reference gene |
| EF-1α-qR | CGTGGTGCATCTCCACAGACT | Reference gene |
| LUC1-F | TAAAGGTGCCACGGGCCACTCAGG | Sequence Validation |
| LUC1-R | CATTTTTTTCTTTGTGTCTATGAG | Sequence Validation |
| LUC2-F | CAAGTGAAATACATCGGCATGTCAC | Sequence Validation |
| LUC2-R | CATTTTTTTCTTTGTGTCTATGAG | Sequence Validation |
| LUC3-F | ATTGGTACGAAGCGTGTCTGTTAT | Sequence Validation |
| LUC3-R | CATTTTTTTCTTTGTGTCTATGAG | Sequence Validation |
| LUC4-F | GATGAATTGCTGTGAGACGAGAGT | Sequence Validation |
| LUC4-R | CATTTTTTTCTTTGTGTCTATGAG | Sequence Validation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, W.-Y.; Huang, J.-H.; Zhou, F.-L.; Yang, Q.-B.; Li, Y.-D.; Jiang, S.; Jiang, S.-G.; Yang, L.-S. Identification and Expression Analysis of Dsx and Its Positive Transcriptional Regulation of IAG in Black Tiger Shrimp (Penaeus monodon). Int. J. Mol. Sci. 2022, 23, 12701. https://doi.org/10.3390/ijms232012701
Wei W-Y, Huang J-H, Zhou F-L, Yang Q-B, Li Y-D, Jiang S, Jiang S-G, Yang L-S. Identification and Expression Analysis of Dsx and Its Positive Transcriptional Regulation of IAG in Black Tiger Shrimp (Penaeus monodon). International Journal of Molecular Sciences. 2022; 23(20):12701. https://doi.org/10.3390/ijms232012701
Chicago/Turabian StyleWei, Wen-Ya, Jian-Hua Huang, Fa-Lin Zhou, Qi-Bin Yang, Yun-Dong Li, Song Jiang, Shi-Gui Jiang, and Li-Shi Yang. 2022. "Identification and Expression Analysis of Dsx and Its Positive Transcriptional Regulation of IAG in Black Tiger Shrimp (Penaeus monodon)" International Journal of Molecular Sciences 23, no. 20: 12701. https://doi.org/10.3390/ijms232012701