
Promoter of Vegetable Soybean GmTIP1;6 Responds to Diverse Abiotic Stresses and Hormone Signals in Transgenic Arabidopsis
Abstract
:1. Introduction
2. Results
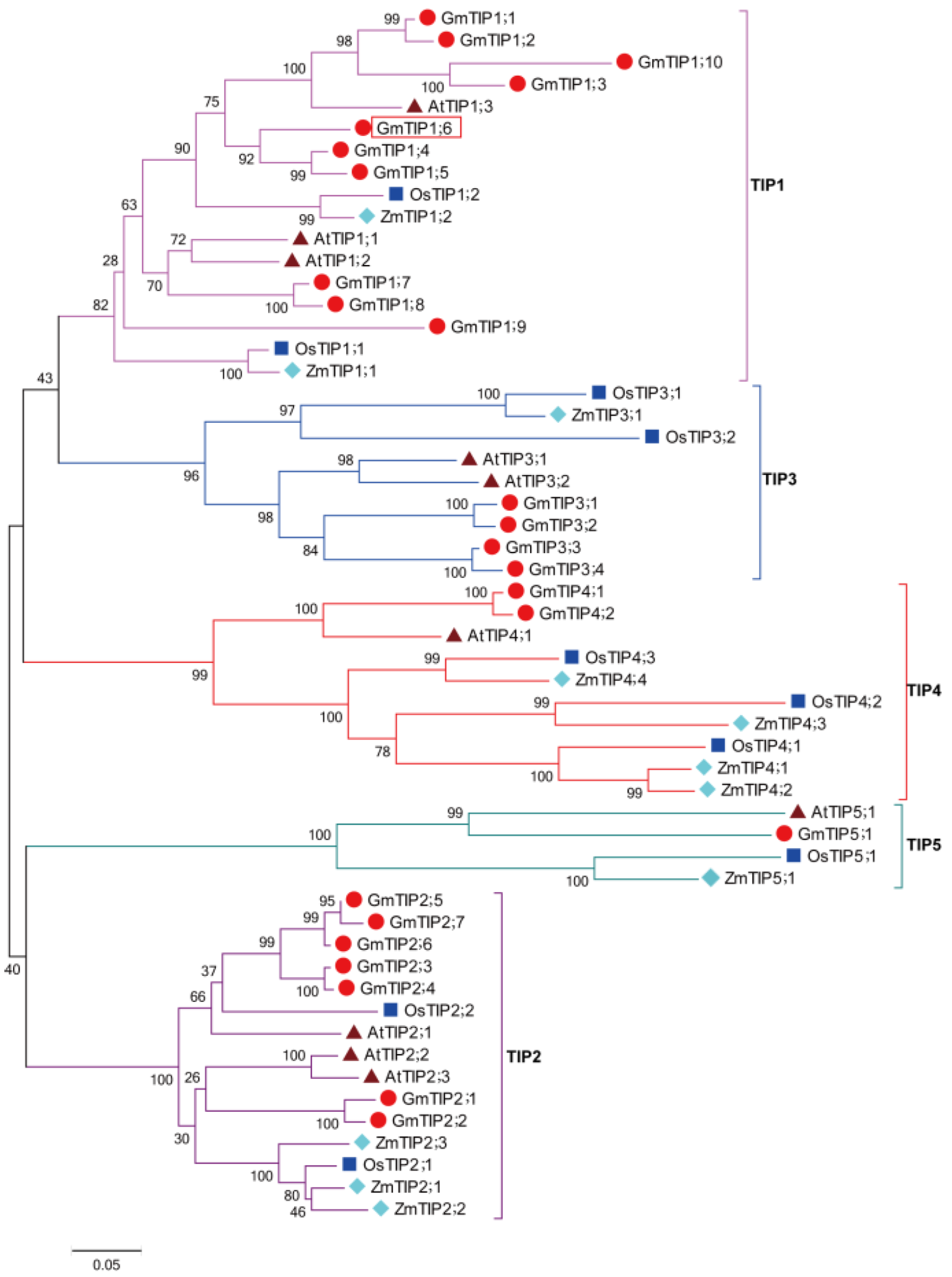
2.1. Phylogenetic Analysis of GmTIP1;6 Gene
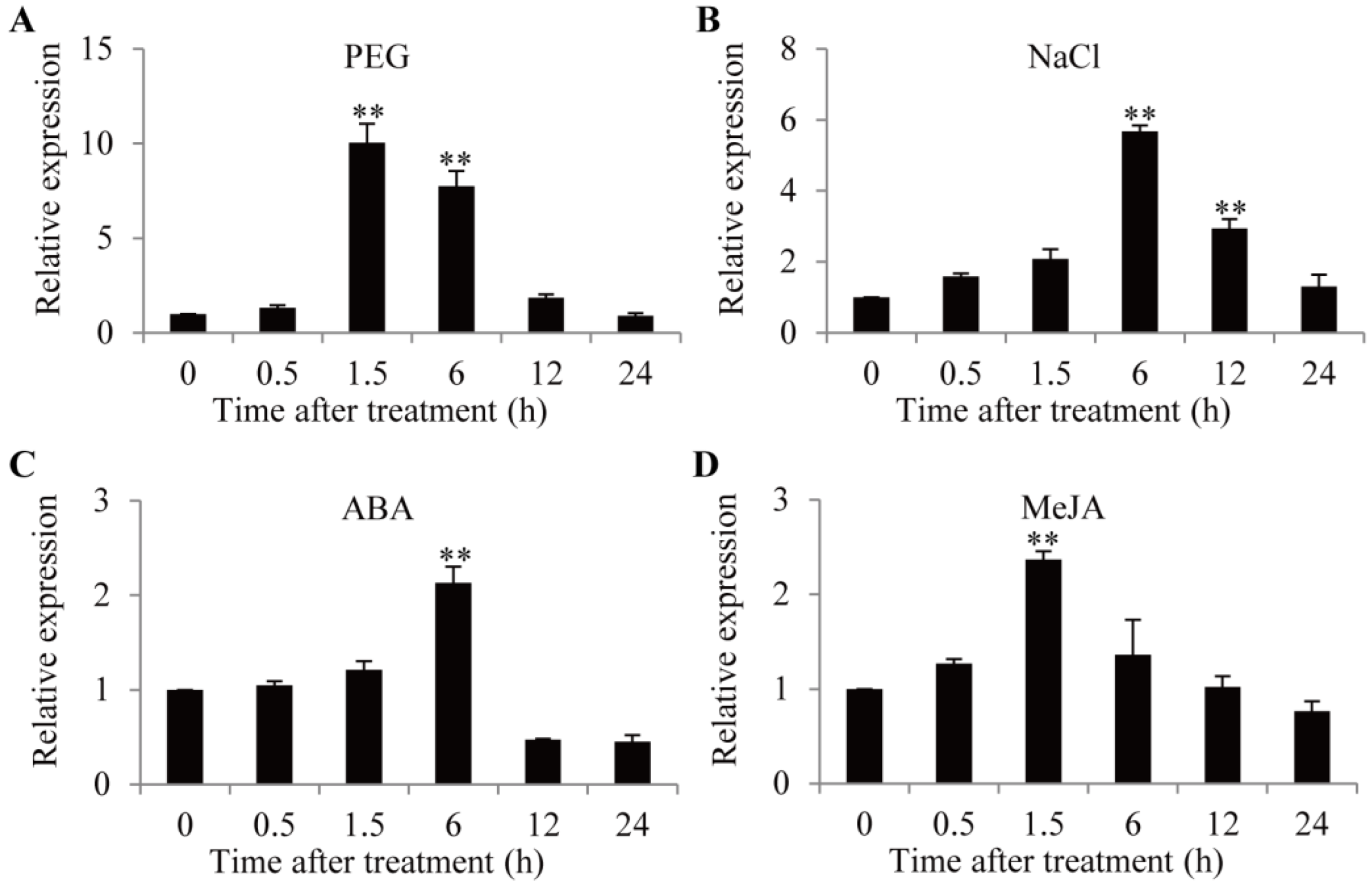
2.2. Expression Patterns of GmTIP1;6 Gene
2.3. Isolation and Cis-Acting Element Distribution of GmTIP1;6 Promoter
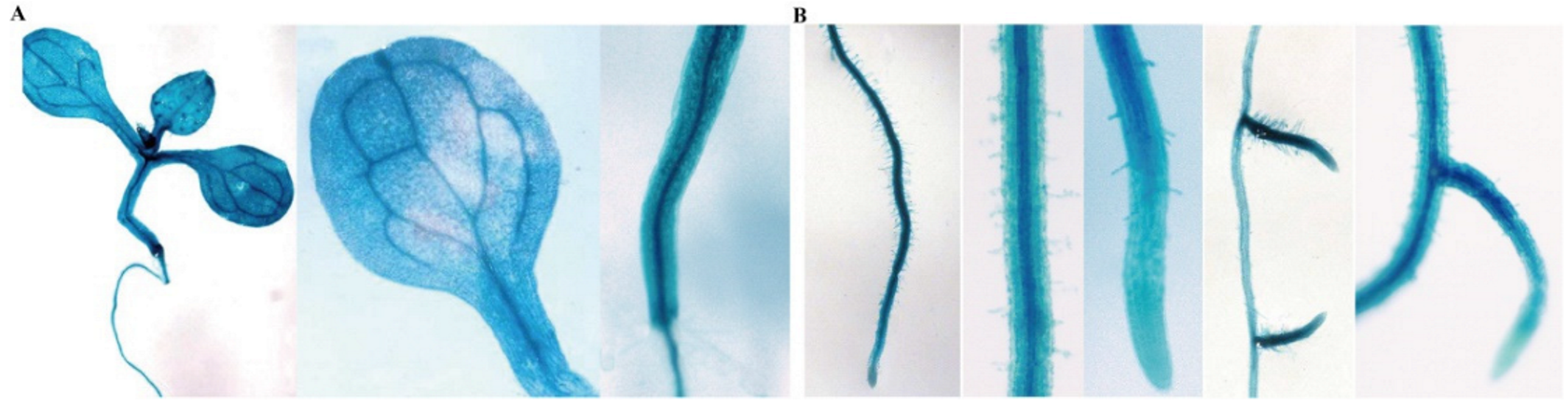
2.4. Histochemical Localization of GmTIP1;6 Promoter
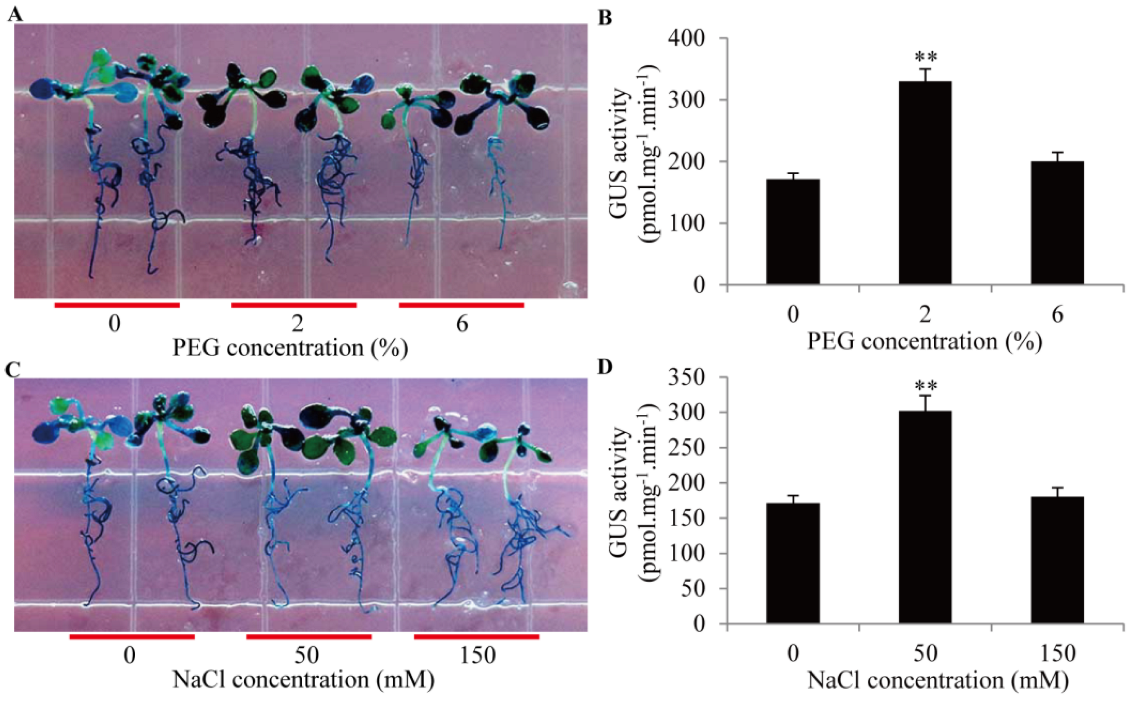
2.5. Activities of GmTIP1;6 Promoter in Response to Drought and Salt Stresses
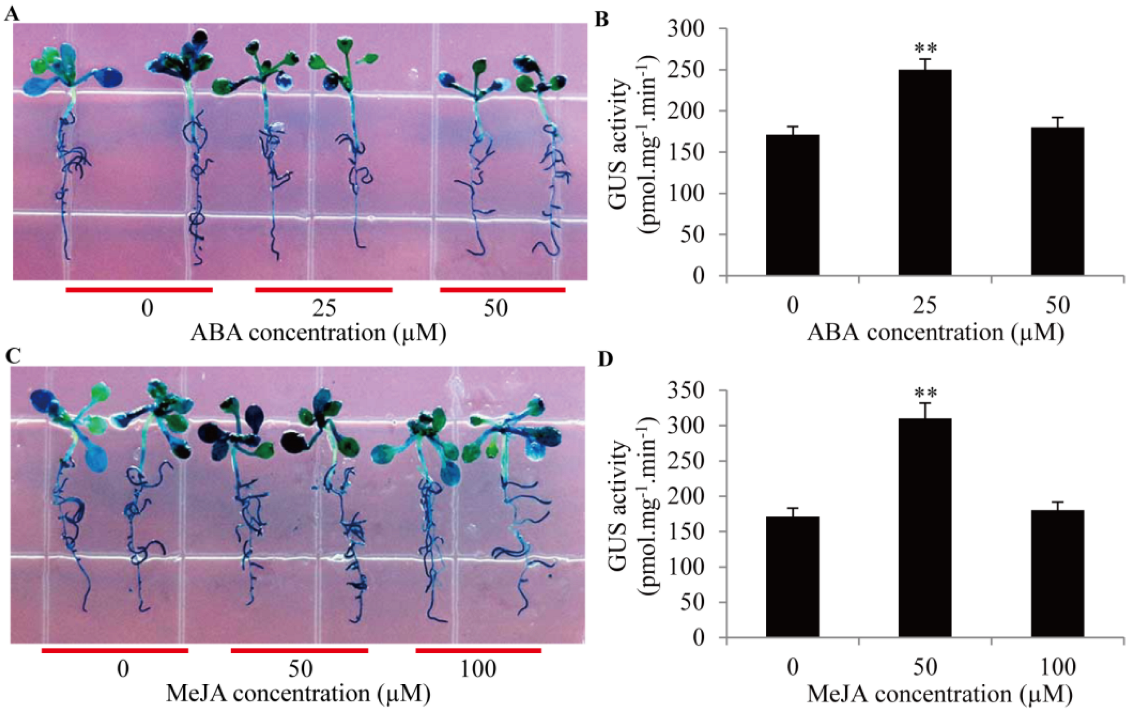
2.6. Activities of GmTIP1;6 Promoter in Response to ABA and MeJA Signals
3. Discussion
4. Materials and Methods
4.1. Stress and Hormonal Treatments for Vegetable Soybean
4.2. Phylogenetic Analysis for GmTIP1;6
4.3. RNA Extraction and qRT-PCR Analysis for GmTIP1;6
4.4. DNA Extraction and Promoter Cloning of GmTIP1;6
4.5. Cis-Acting Element Analysis of GmTIP1;6 Promoter
4.6. Vector Construction of GmTIP1;6 Promoter and Transgenic Arabidopsis Generation
4.7. Stress and Hormone Treatments for Transgenic Arabidopsis
4.8. GUS Staining and Activity Detection for Transgenic Arabidopsis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Liu, N.; Niu, Y.; Zhang, G.; Feng, Z.; Bo, Y.; Lian, J.; Wang, B.; Gong, Y. Genome sequencing and population resequencing provide insights into the genetic basis of domestication and diversity of vegetable soybean. Hortic. Res. 2022, 9, uhab052. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.; He, S.; Wang, C.; Lai, Z.; Lin, D.; Chen, S. A modified roger’s distance algorithm for mixed quantitative-qualitative phenotypes to establish a core collection for Taiwanese vegetable soybeans. Front. Plant Sci. 2021, 11, 612106. [Google Scholar] [CrossRef]
- Zhang, B.; Lord, N.; Kuhar, T.; Duncan, S.; Huang, H.; Ross, J.; Rideout, S.; Arancibia, R.; Reiter, M.; Li, S.; et al. ‘VT Sweet’: A vegetable soybean cultivar for commercial edamame production in the mid-Atlantic USA. J. Plant Regist. 2022, 16, 29–33. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, W.; Zhou, Y.; Ji, P.; Wan, Y.; Shi, S.; Yang, Z.; Gong, Y.; Mu, F.; Chen, S. Integrative analysis of metabolome and transcriptome reveals the improvements of seed quality in vegetable soybean (Glycine max (L.) Merr.). Phytochemistry 2022, 200, 113216. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Liu, H.; Li, S.; Zhang, W.; Wang, Q.; Zhang, H.; Liu, X.; Cui, X.; Chen, X.; Tang, W.; et al. GWAS and identification of candidate genes associated with seed soluble sugar content in vegetable soybean. Agron. J. 2022, 12, 1470. [Google Scholar] [CrossRef]
- Zhang, G.; Hu, Q.; Xu, S.; Gong, Y. Polyamines play a positive role in salt tolerant mechanisms by activating antioxidant enzymes in roots of vegetable soybean. Legume Res. 2013, 36, 234–240. [Google Scholar]
- Xu, S.; Liu, N.; Mao, W.; Hu, Q.; Wang, G.; Gong, Y. Identification of chilling-responsive microRNAs and their targets in vegetable soybean (Glycine max L.). Sci. Rep. 2016, 6, 26619. [Google Scholar] [CrossRef] [Green Version]
- Moloi, M.J.; van der Merwe, R. Drought tolerance responses in vegetable-type soybean involve a network of biochemical mechanisms at flowering and pod-filling stages. Plants 2021, 10, 1502. [Google Scholar] [CrossRef]
- Chen, G.; Yan, W.; Yang, L.; Gai, J.; Zhu, Y. Overexpression of StNHX1, a novel vacuolar Na+/H+ antiporter gene from Solanum torvum, enhances salt tolerance in transgenic vegetable soybean. Hortic. Environ. Biote. 2014, 55, 213–221. [Google Scholar] [CrossRef]
- Yan, W.; Chen, G.; Yang, L.; Gai, J.; Zhu, Y. Overexpression of the rice phosphate transporter gene OsPT6 enhances tolerance to low phosphorus stress in vegetable soybean. Sci. Hortic. 2014, 177, 71–76. [Google Scholar]
- Zhang, G.; Zhu, W.; Gai, J.; Zhu, Y.; Yang, L. Enhanced salt tolerance of transgenic vegetable soybeans resulting from overexpression of a novel Δ1-pyrroline-5-carboxylate synthetase gene from Solanum torvum Swartz. Hortic. Environ. Biotechnol. 2015, 56, 94–104. [Google Scholar] [CrossRef]
- Wang, X.; Gao, F.; Yang, S.; Gai, J.; Zhu, Y. Overexpression of the StP5CS gene promotes nodulation and nitrogen fixation in vegetable soybean under drought stress. Legume Res. 2019, 42, 603–608. [Google Scholar] [CrossRef]
- Gautam, A.; Pandey, A.K. Aquaporins responses under challenging environmental conditions and abiotic stress tolerance in plants. Bot. Rev. 2021, 87, 467–495. [Google Scholar] [CrossRef]
- Maurel, C.; Reizer, J.; Schroeder, J.I.; Chrispeels, M.J. The vacuolar membrane protein gamma-TIP creates water specific channels in Xenopus oocytes. EMBO J. 1993, 12, 2241–2247. [Google Scholar] [CrossRef]
- Singh, R.K.; Deshmukh, R.; Muthamilarasan, M.; Rani, R.; Prasad, M. Versatile roles of aquaporin in physiological processes and stress tolerance in plants. Plant Physiol. Bioch. 2020, 149, 178–189. [Google Scholar] [CrossRef]
- Pawłowicz, I.; Masajada, K. Aquaporins as a link between water relations and photosynthetic pathway in abiotic stress tolerance in plants. Gene 2019, 687, 166–172. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Penna, S.; Nguyen, D.V.; Tran, L.S.P. Multifaceted roles of aquaporins as molecular conduits in plant responses to abiotic stresses. Crit. Rev. Biotechnol. 2016, 36, 389–398. [Google Scholar] [CrossRef]
- Jauh, G.Y.; Phillips, T.E.; Rogers, J.C. Tonoplast intrinsic protein isoforms as markers for vacuolar functions. Plant Cell 1999, 11, 1867–1882. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Liu, N.; Zhang, G.; Niu, F.; Xu, S.; Gong, Y. Investigation of the AQP family in soybean and the promoter activity of TIP2;6 in heat stress and hormone responses. Int. J. Mol. Sci. 2019, 20, 262. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.; Kim, D.G.; Kim, Y.O.; Kim, J.S.; Kang, H. An expression analysis of a gene family encoding plasma membrane aquaporins in response to abiotic stresses in Arabidopsis thaliana. Plant Mol. Biol. 2004, 54, 713–725. [Google Scholar] [CrossRef]
- Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M.; Maeshima, M. Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell Physiol. 2005, 46, 1568–1577. [Google Scholar] [CrossRef] [Green Version]
- Chaumont, F.; Barrieu, F.; Wojcik, E.; Chrispeels, M.J.; Jung, R. Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol. 2001, 125, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Sudhakaran, S.; Thakral, V.; Padalkar, G.; Rajora, N.; Dhiman, P.; Raturi, G.; Sonah, H. Significance of solute specificity, expression, and gating mechanism of tonoplast intrinsic protein during development and stress response in plants. Physiol. Plant. 2021, 172, 258–274. [Google Scholar] [CrossRef] [PubMed]
- Regon, P.; Panda, P.; Kshetrimayum, E.; Panda, S.K. Genome-wide comparative analysis of tonoplast intrinsic protein (TIP) genes in plants. Funct. Integr. Genom. 2014, 14, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, K.; Fukaya, T. Molecular architecture of enhancer-promoter interaction. Curr. Opin. Cell. Biol. 2022, 74, 62–70. [Google Scholar] [CrossRef]
- Zhou, S.; Du, G.; Kang, Z.; Li, J.; Chen, J.; Li, H.; Zhou, J. The application of powerful promoters to enhance gene expression in industrial microorganisms. World J. Microb. Biot. 2017, 33, 23. [Google Scholar] [CrossRef]
- Nevoigt, E.; Fischer, C.; Mucha, O.; Matthäus, F.; Stahl, U.; Stephanopoulos, G. Engineering promoter regulation. Biotechnol. Bioeng. 2007, 96, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Juven-Gershon, T.; Cheng, S.; Kadonaga, J.T. Rational design of a super core promoter that enhances gene expression. Nat. Methods 2006, 3, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lee, J.H.; Poindexter, M.R.; Shao, Y.; Liu, W.; Lenaghan, S.C.; Ahkami, A.H.; Blumwald, B.; Stewart, C.N., Jr. Rational design and testing of abiotic stress-inducible synthetic promoters from poplar cis-regulatory elements. Plant Biotechnol. J. 2021, 19, 1354–1369. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Xu, Y.; Huang, D.; Miao, H.; Liu, J.; Jia, C.; Hu, W.; Valarezo, A.V.; Xu, B.; Jin, Z. Identification of a novel promoter from banana aquaporin family gene (MaTIP1;2) which responses to drought and salt-stress in transgenic Arabidopsis thaliana. Plant Physiol. Biochem. 2018, 128, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tong, J.; He, X.; Xu, Z.; Xu, L.; Wei, P.; Huang, Y.; Brestic, M.; Ma, H.; Shao, H. A novel soybean intrinsic protein gene, GmTIP2;3, involved in responding to osmotic Stress. Front. Plant Sci. 2016, 6, 1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, M.M. Phenomorphological characterization of vegetable soybean germplasm lines for commercial production. Crop Sci. 2015, 55, 1274–1279. [Google Scholar] [CrossRef]
- Hlahla, J.M.; Mafa, M.S.; Van der Merwe, R.; Alexander, O.; Duvenhage, M.M.; Kemp, G.; Moloi, M.J. The photosynthetic efficiency and carbohydrates responses of six edamame (Glycine max. L. Merrill) cultivars under drought stress. Plants 2022, 11, 394. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, V.; Pandey, N.; Ramteke, R.; Verghese, P.; Reddy, R.; Onkarappa, T.; Mehtre, S.P.; Gupta, S.; Satpute, G.K.; Mohan, Y.; et al. GGE biplot analysis of vegetable type soybean genotypes under multi-environmental conditions in India. J. Environ. Biol. 2021, 42, 247–253. [Google Scholar] [CrossRef]
- Comlekcioglu, N.; Simsek, M. Effects of deficit irrigation on yield and yield components of vegetable soybean [Glycine max L. (Merr.)] in semi-arid conditions. Afr. J. Biotechnol. 2011, 10, 6227–6234. [Google Scholar]
- Keatinge, J.D.H.; Easdown, W.J.; Yang, R.Y.; Chadha, M.L.; Shanmugasundaram, S. Overcoming chronic malnutrition in a future warming world: The key importance of mungbean and vegetable soybean. Euphytica 2011, 180, 129–141. [Google Scholar] [CrossRef]
- Rojas-Pierce, M. Targeting of tonoplast proteins to the vacuole. Plant Sci. 2013, 211, 132–136. [Google Scholar] [CrossRef]
- Gattolin, S.; Sorieul, M.; Frigerio, L. Tonoplast intrinsic proteins and vacuolar identity. Biochem. Soc. Trans. 2010, 38, 769–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauh, G.Y.; Fischer, A.M.; Grimes, H.D.; Ryan, C.A., Jr.; Rogers, J.C. δ-tonoplast intrinsic protein defines unique plant vacuole functions. Proc. Natl. Acad. Sci. USA 1998, 95, 12995–12999. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Xu, S.; Liu, N.; Hang, G.; Hu, Q.; Xu, Z.; Gong, Y. Identification of the AQP members involved in abiotic stress responses from Arabidopsis. Gene 2018, 646, 64–73. [Google Scholar] [CrossRef]
- Footitt, S.; Clewes, R.; Feeney, M.; Finch-Savage, W.E.; Frigerio, L. Aquaporins influence seed dormancy and germination in response to stress. Plant Cell Environ. 2019, 2, 2325–2339. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Sun, W. Arabidopsis seed-specific vacuolar aquaporins are involved in maintaining seed longevity under the control of ABSCISIC ACID INSENSITIVE 3. J. Exp. Bot. 2015, 66, 4781–4794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, F.; Bousser, A.; Sissoëff, I.; Hoarau, J.; Mahé, A. Characterization in maize of ZmTIP2-3, a root-specific tonoplast intrinsic protein exhibiting aquaporin activity. J. Exp. Bot. 2004, 55, 539–541. [Google Scholar] [CrossRef] [PubMed]
- Hofte, H.; Hubbard, L.; Reizer, J.; Ludevid, D.; Herman, E.M.; Chrispeels, M.J. Vegetative and seed-specific forms of tonoplast intrinsic protein in the vacuolar membrane of Arabidopsis thaliana. Plant Physiol. 1992, 99, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Xin, S.; Yu, G.; Sun, L.; Qiang, X.; Xu, N.; Cheng, X. Expression of tomato SlTIP2;2 enhances the tolerance to salt stress in the transgenic Arabidopsis and interacts with target proteins. J. Plant Res. 2014, 127, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Wang, M.; Zhang, L.; Wei, H.; Wang, H.; Lu, J.; Yu, S. Overexpression of a cotton aquaporin gene GhTIP1;1-like confers cold tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2022, 23, 1361. [Google Scholar] [CrossRef]
- Guo, A.; Hao, J.; Su, Y.; Li, B.; Zhao, N.; Zhu, M.; Huang, Y.; Tian, B.; Shi, G.; Hua, J. Two aquaporin genes, GhPIP2;7 and GhTIP2;1, positively regulate the tolerance of upland cotton to salt and osmotic stresses. Front. Plant Sci. 2021, 12, 780486. [Google Scholar] [CrossRef]
- Wang, L.L.; Chen, A.P.; Zhong, N.Q.; Liu, N.; Wu, X.M.; Wang, F.; Yang, C.L.; Romero, M.F.; Xia, G.X. The Thellungiella salsuginea tonoplast aquaporin TsTIP1;2 functions in protection against multiple abiotic stresses. Plant Cell Physiol. 2014, 55, 148–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.; Agarwal, P.; Shanware, A.; Sane, V.A. Heterologous expression of two Jatropha aquaporins imparts drought and salt tolerance and improves seed viability in transgenic Arabidopsis thaliana. PLoS ONE 2015, 10, e0128866. [Google Scholar] [CrossRef]
- Song, S.; Zhang, D.; Ma, F.; Xing, W.; Huang, D.; Wu, B.; Chen, J.; Chen, D.; Xu, B.; Xu, Y. Genome-wide Identification and expression analyses of the aquaporin gene family in passion fruit (Passiflora edulis), revealing PeTIP3-2 to be involved in drought stress. Int. J. Mol. Sci. 2022, 23, 5720. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; An, Y.; Wu, J.; He, S.; Sun, L.; Hao, F. Wheat TaTIP4;1 confers enhanced tolerance to drought, salt and osmotic stress in Arabidopsis and rice. Int. J. Mol. Sci. 2022, 23, 2085. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Ji, W.; Bai, X.; Cai, H.; Zhu, D.; Sun, X.L.; Chen, L.J.; Zhu, Y.M. A novel Glycine soja tonoplast intrinsic protein gene responds to abiotic stress and depresses salt and dehydration tolerance in transgenic Arabidopsis thaliana. J. Plant Physiol. 2011, 168, 1241–1248. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, M.; Zhou, L.; Quan, T.; Xia, G. Heterologous expression of the wheat aquaporin gene TaTIP2;2 compromises the abiotic stress tolerance of Arabidopsis thaliana. PLoS ONE 2013, 8, e79618. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Nguyen, N.A.; Deshmukh, R.K.; Patil, G.B.; Prince, S.J.; Valliyodan, B.; Mutava, R.; Pike, S.M.; Gassmann, W.; Nguyen, H.T. Soybean TIP gene family analysis and characterization of GmTIP1;5 and GmTIP2;5 water transport activity. Front. Plant Sci. 2016, 7, 1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 4. [Google Scholar] [CrossRef]
- Fox, A.R.; Maistriaux, L.C.; Chaumont, F. Toward understanding of the high number of plant aquaporin isoforms and multiple regulation mechanisms. Plant Sci. 2017, 264, 179–187. [Google Scholar] [CrossRef]
- Kreida, S.; Törnroth-Horsefield, S. Structural insights into aquaporin selectivity and regulation. Curr. Opin. Struct. Biol. 2015, 33, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Han, C.S.; Kim, S.; Lee, S.E.; Choi, S.; Kim, S.H.; Sun Yoon, I.; Hwang, Y.S. Cross-talk between ABA and sugar signaling is mediated by the ACGT core and CE1 element reciprocally in OsTIP3;1 promoter. J. Plant Physiol. 2018, 224, 103–111. [Google Scholar] [CrossRef]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Harshitha, R.; Arunraj, D.R. Real-time quantitative PCR: A tool for absolute and relative quantification. Biochem. Mol. Biol. Int. 2021, 49, 800–812. [Google Scholar] [CrossRef]
- Yang, Z.; Du, H.; Xing, X.; Li, W.; Kong, Y.; Li, X.; Zhang, C. A small heat shock protein, GmHSP17.9, from nodule confers symbiotic nitrogen fixation and seed yield in soybean. Plant Biotechnol. J. 2022, 20, 103. [Google Scholar] [CrossRef]
- Duan, Z.; Zhang, M.; Zhang, Z.; Liang, S.; Fan, L.; Yang, X.; Yuan, Y.; Pan, Y.; Zhou, G.; Liu, S.; et al. Natural allelic variation of GmST05 controlling seed size and quality in soybean. Plant Biotechnol. J. 2022, 20, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Gao, L.; Ke, M.; Gao, Z.; Tu, T.; Huang, L.; Chen, J.; Guan, Y.; Huang, X.; Chen, X. GmPIN1-mediated auxin asymmetry regulates leaf petiole angle and plant architecture in soybean. J. Integr. Plant Biol. 2022, 64, 1325–1338. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element Name | Core Sequence | Number | Location (bp) | Function | |
|---|---|---|---|---|---|
| (+) Sense Strand | (−) Antisense Strand | ||||
| MBS | CAACTG | 1 | +266 | Drought responsive | |
| ABRE-element | ACGTG | 5 | +1233 | −585 −1235 | ABA responsive |
| +1236 | |||||
| +1291 | |||||
| CGTCA-motif | CGTCA | 2 | +447 | −444 | MeJA responsive |
| TGACG-motif | TGACG | 2 | +444 | −447 | MeJA responsive |
| TCA-element | TCAGAAGAGG | 1 | +853 | SA responsive | |
| P-box | CCTTTTG | 1 | −1153 | GA responsive | |
| TATC-box | TATCCCA | 1 | +614 | GA responsive | |
| TGA-element | AACGA | 1 | +1413 | Auxin responsive | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Z.; Liu, N.; Zhang, G.; Bu, Y.; Wang, B.; Gong, Y. Promoter of Vegetable Soybean GmTIP1;6 Responds to Diverse Abiotic Stresses and Hormone Signals in Transgenic Arabidopsis. Int. J. Mol. Sci. 2022, 23, 12684. https://doi.org/10.3390/ijms232012684
Feng Z, Liu N, Zhang G, Bu Y, Wang B, Gong Y. Promoter of Vegetable Soybean GmTIP1;6 Responds to Diverse Abiotic Stresses and Hormone Signals in Transgenic Arabidopsis. International Journal of Molecular Sciences. 2022; 23(20):12684. https://doi.org/10.3390/ijms232012684
Chicago/Turabian StyleFeng, Zhijuan, Na Liu, Guwen Zhang, Yuanpeng Bu, Bin Wang, and Yaming Gong. 2022. "Promoter of Vegetable Soybean GmTIP1;6 Responds to Diverse Abiotic Stresses and Hormone Signals in Transgenic Arabidopsis" International Journal of Molecular Sciences 23, no. 20: 12684. https://doi.org/10.3390/ijms232012684