
Functional Characterization of Heat Shock Factor (CrHsf) Families Provide Comprehensive Insight into the Adaptive Mechanisms of Canavalia rosea (Sw.) DC. to Tropical Coral Islands


Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of the C. rosea Hsf Family
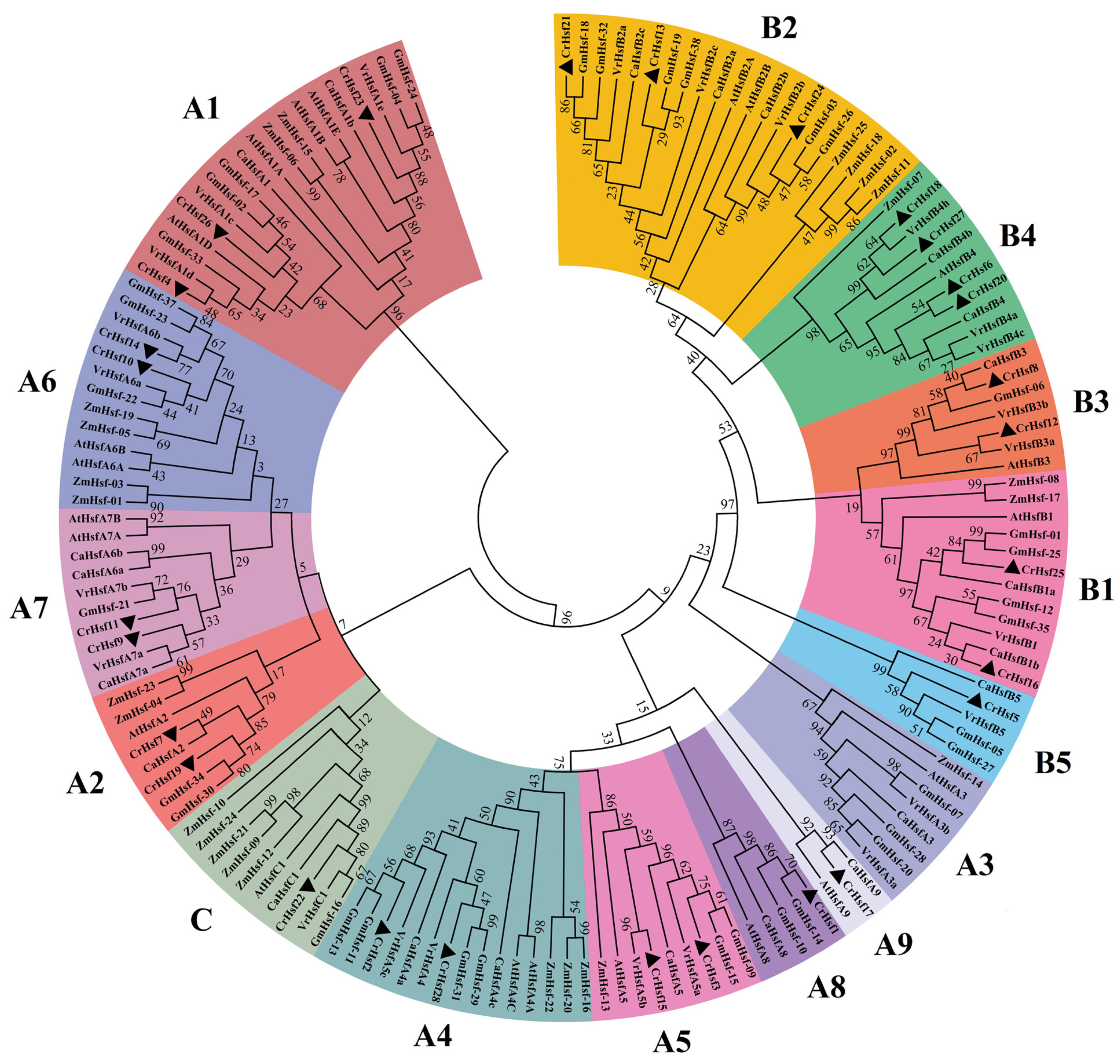
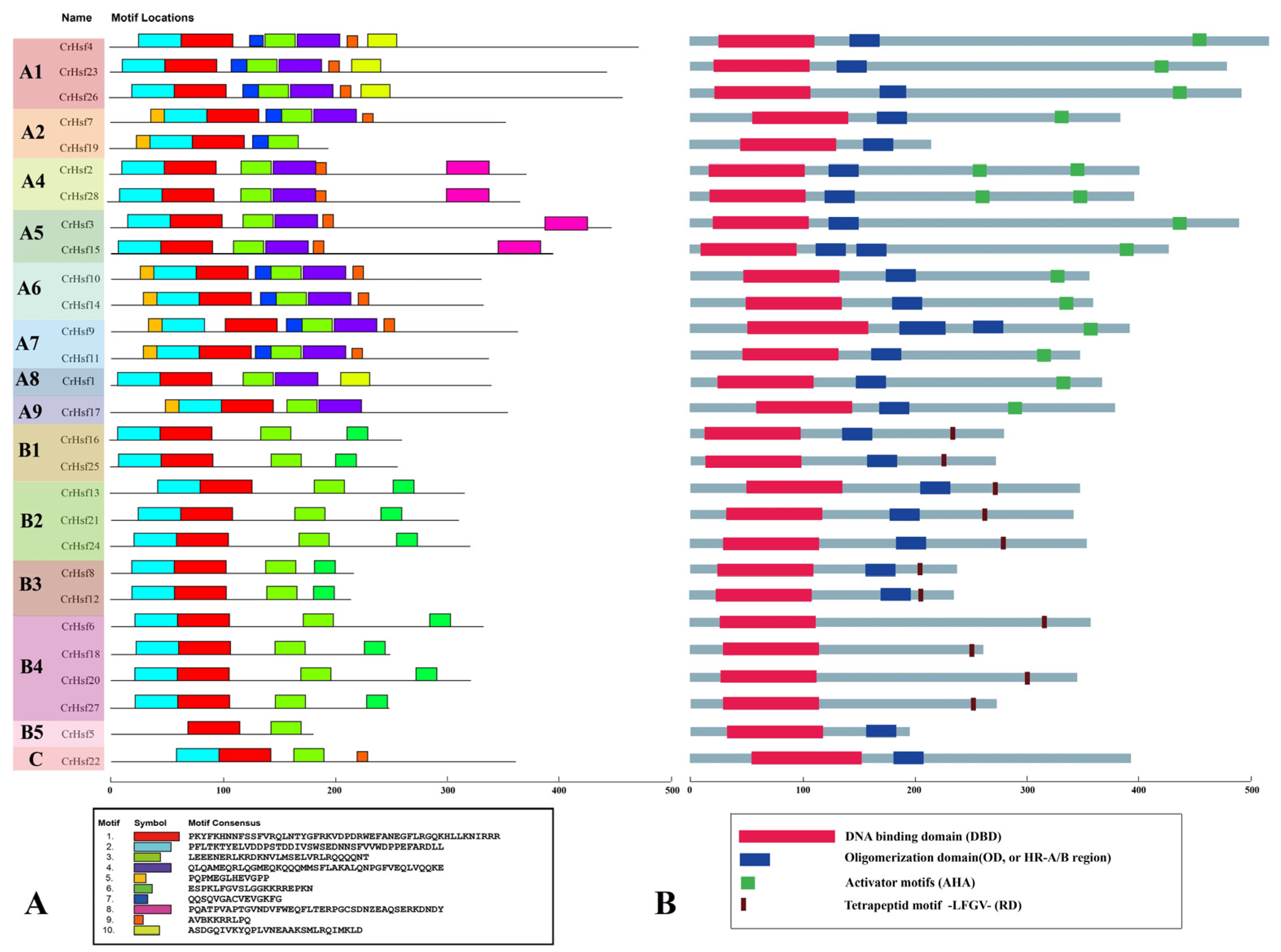
2.2. Phylogenetic Analysis, Classification, and Conserved Motif Analyses of CrHsf Proteins
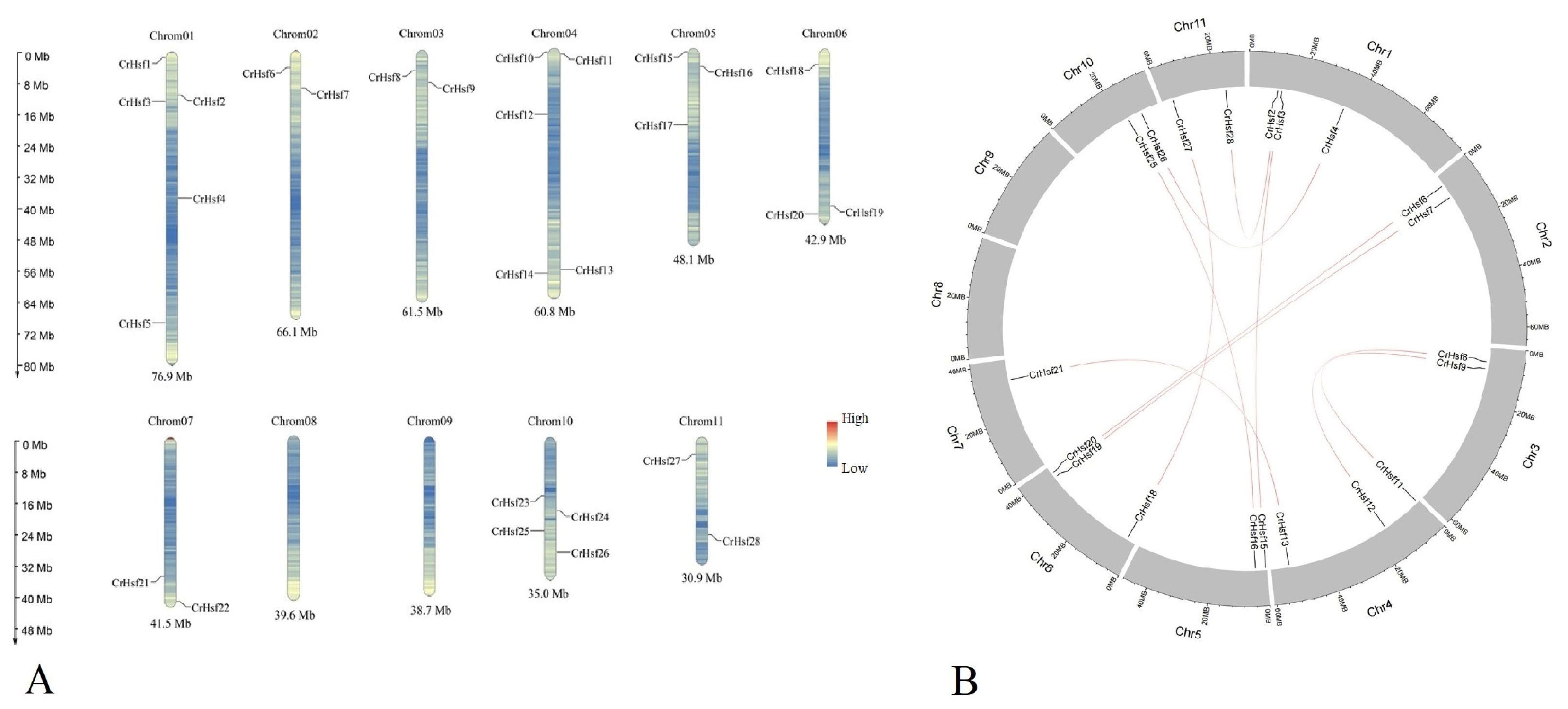
2.3. Chromosomal Location and Duplication of CrHsf Genes
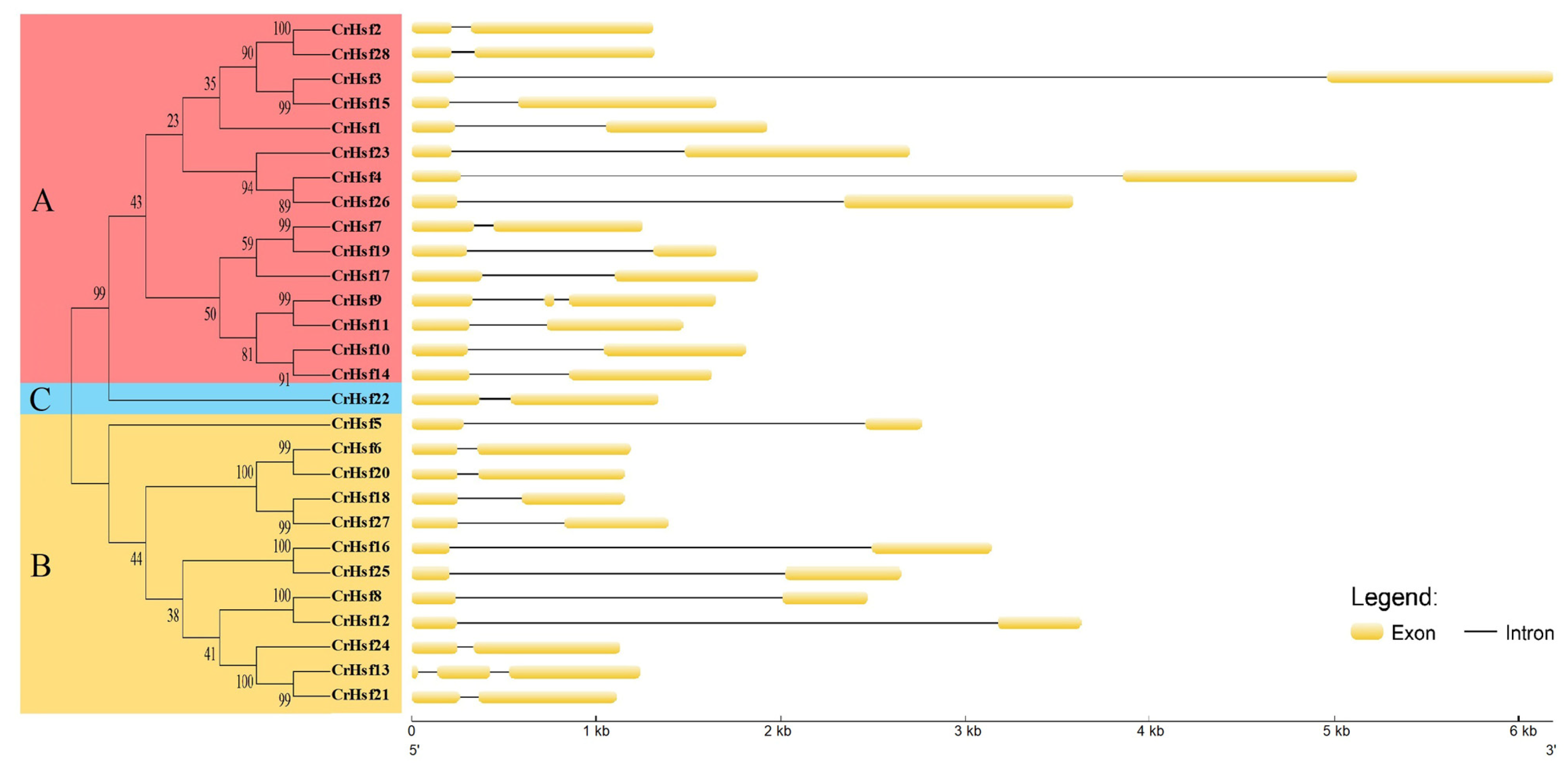
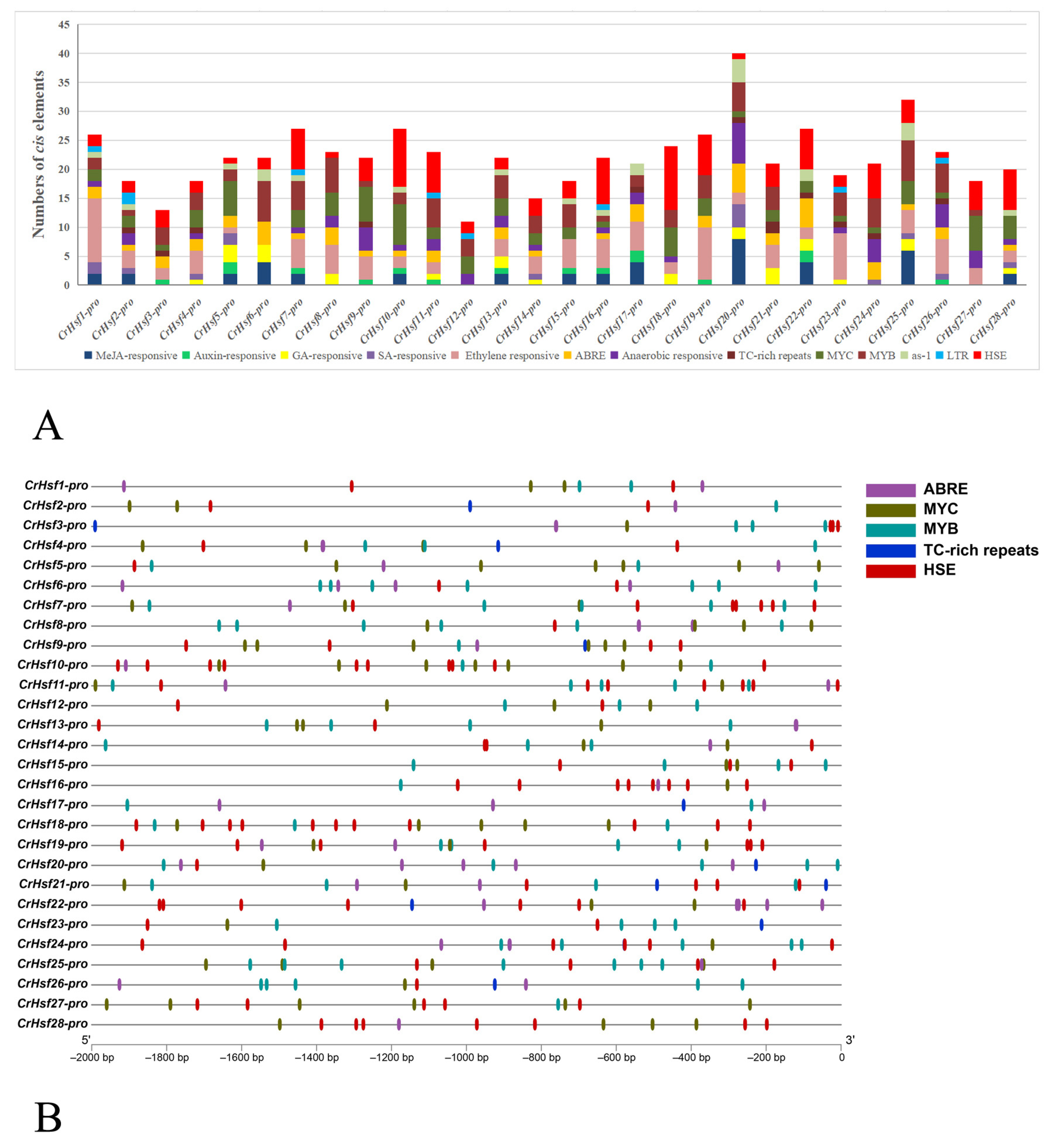
2.4. Gene Structures and Cis-Acting Element Analyses in CrHsf Promoter Regions
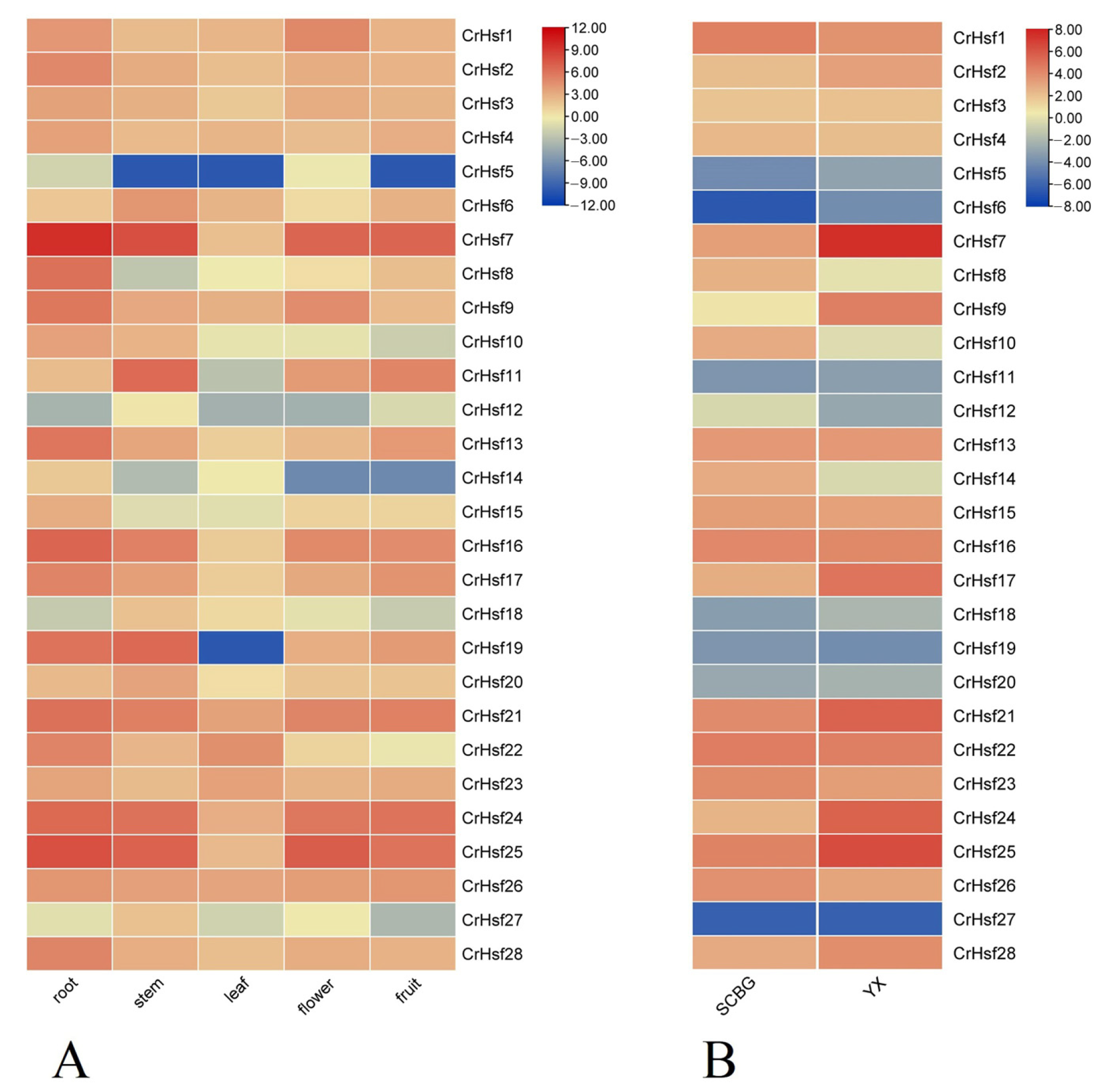
2.5. Expression Profiles of CrHsf Genes in Different Tissues or in Response to Different Habitats
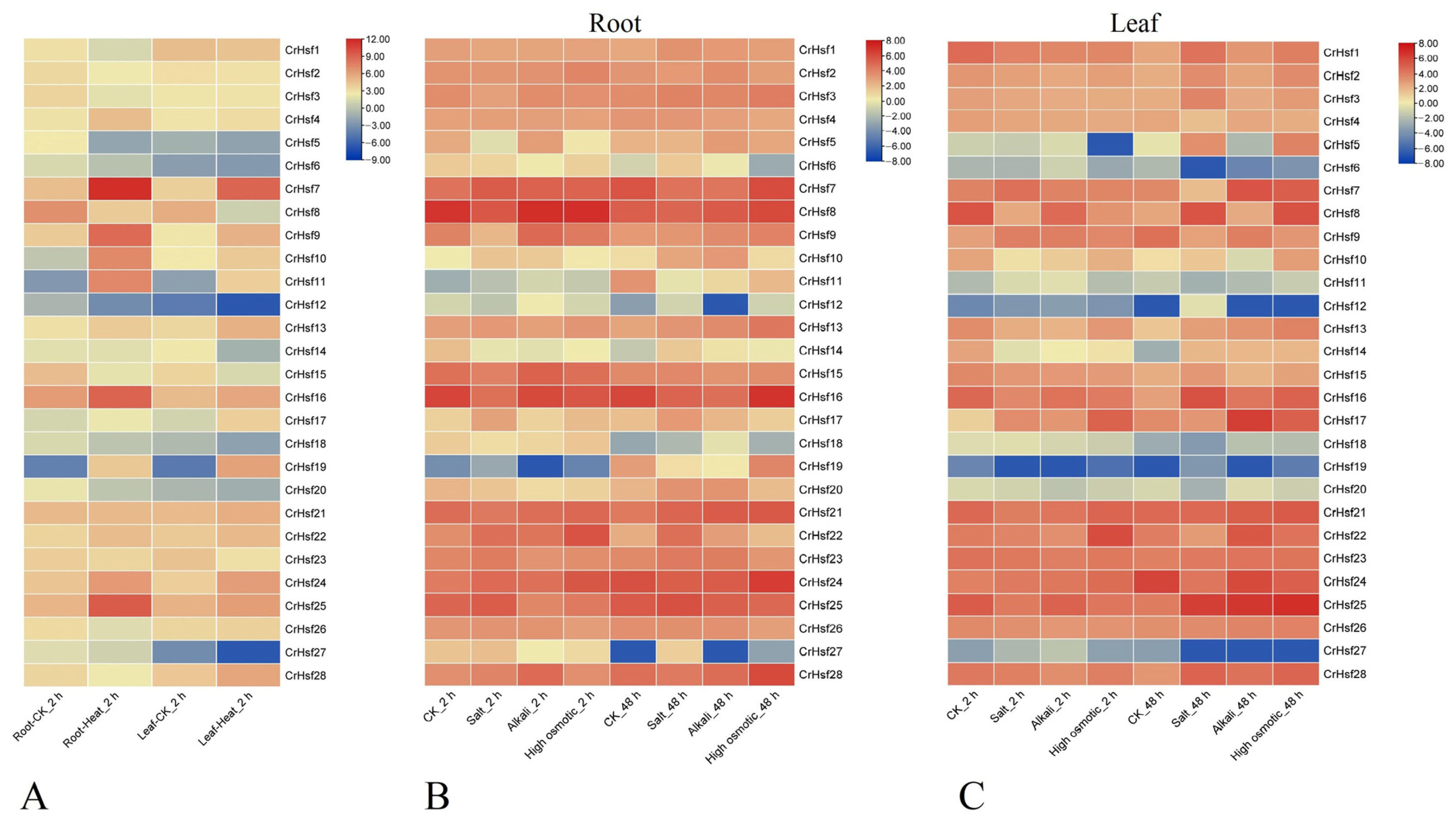
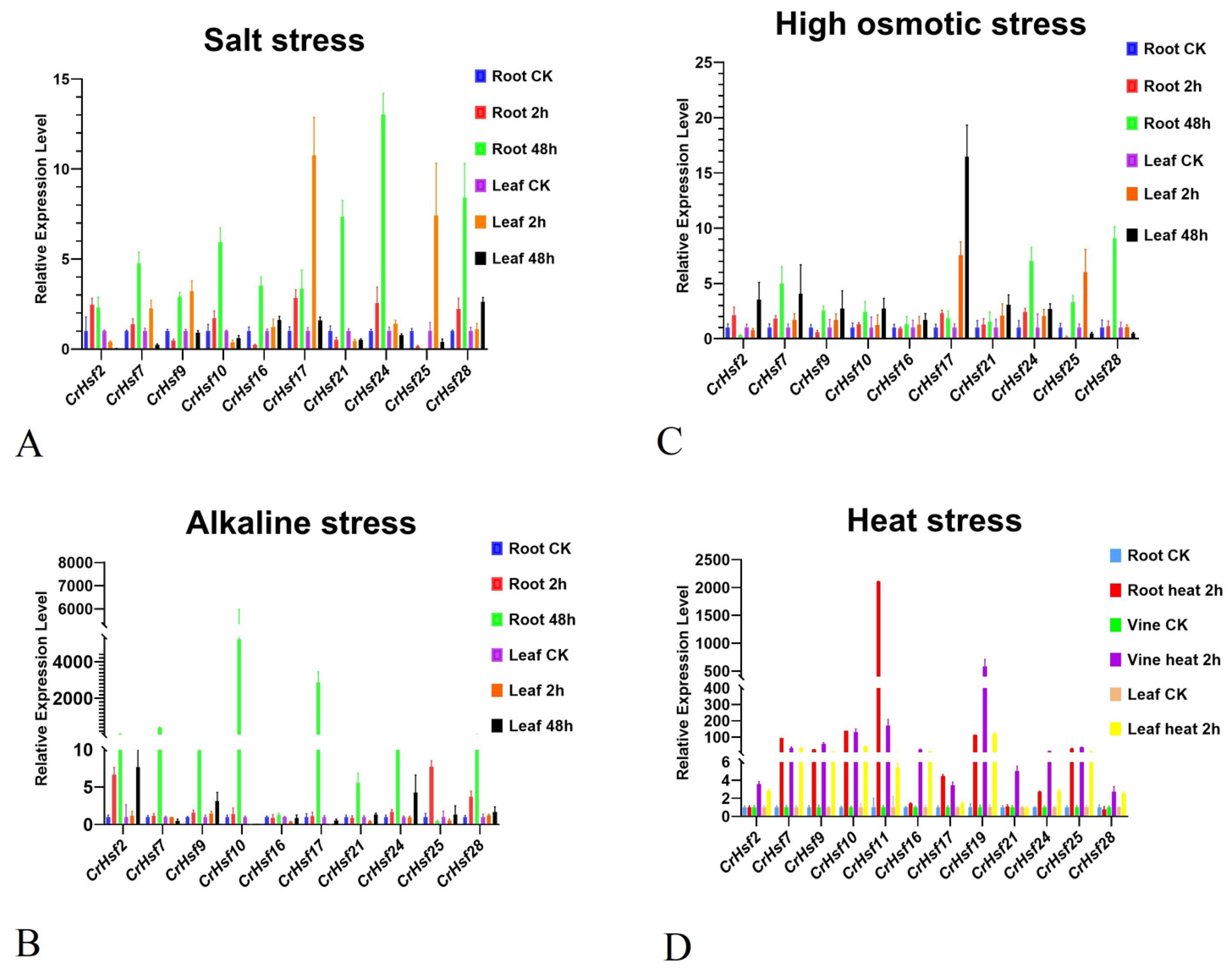
2.6. Expression Profiles of CrHsf Genes in Response to Different Abiotic Stresses
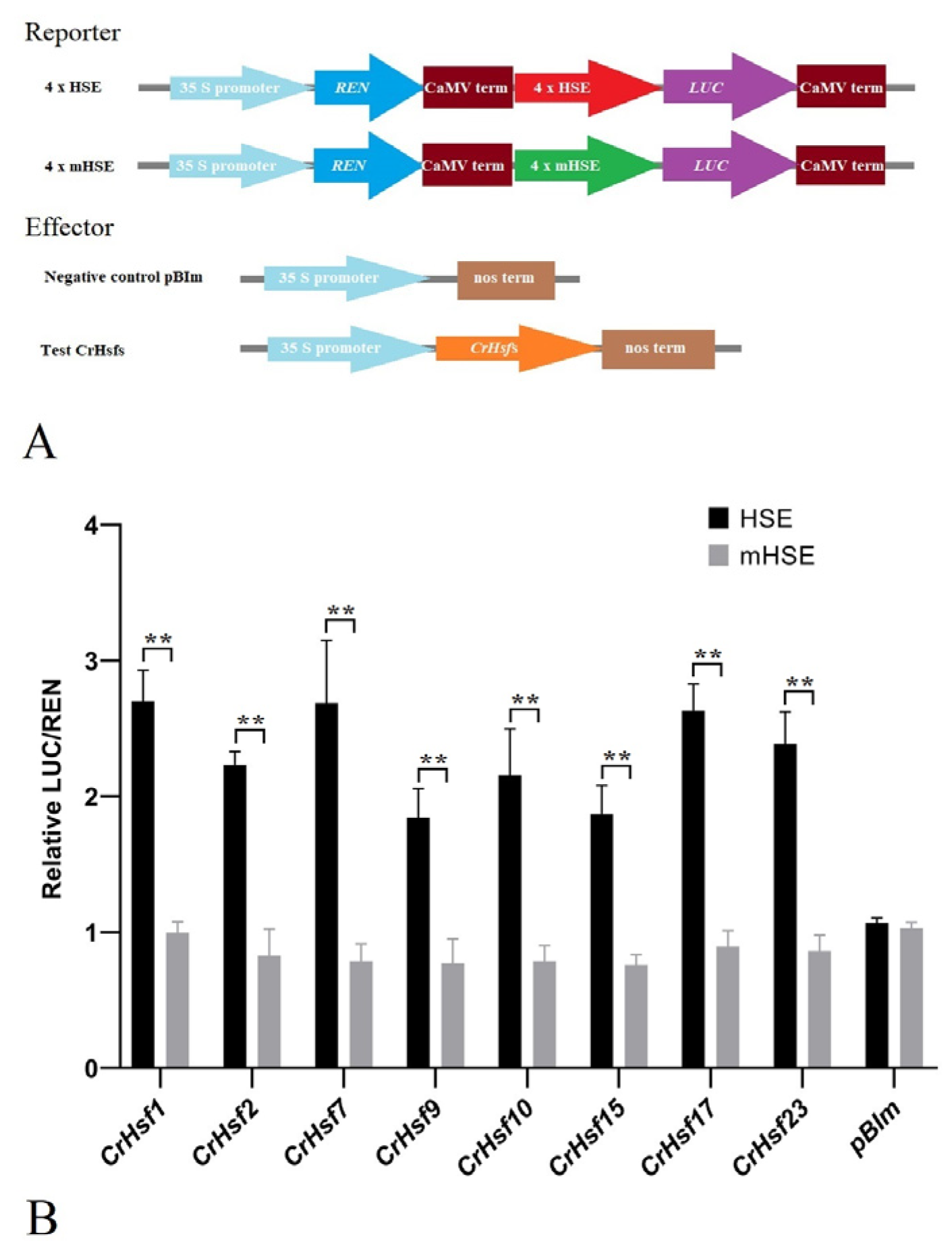
2.7. Transactivation Activity Analysis of the CrHsf Proteins
2.8. Heterologous Expression of CrHsfs Confers Abiotic Tolerance in Transgenic Yeast
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Stress Treatments
4.2. Identification and Bioinformatics Analysis of the Hsf Family in C. rosea
4.3. Phylogenetic and Sequence Conservation Analysis of C. rosea
4.4. Gene Duplication and Gene Collinearity Analysis
4.5. Analysis of Cis-Acting Elements of CrHsf Promoters
4.6. Expression Profile Analysis of CrHsfs and Other Abiotic-Related Genes in Various Tissues/Organs or under Multiple Stresses
4.7. Cloning of CrHsf cDNAs and Transcriptional Activity Analysis of CrHsfs
4.8. Functional Identification of CrHsfs in Yeast
4.9. Statistical Analysis
5. Conclusions and Prospects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siqueira-Silva, A.I.; da Silva, L.C.; Azevedo, A.A.; Oliva, M.A. Iron plaque formation and morphoanatomy of roots from species of restinga subjected to excess iron. Ecotoxicol. Environ. Saf. 2012, 78, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Supriya, P.; Sridhar, K.R. Impact of electron beam irradiation on the bioactive principles of seeds of coastal sand dune wild legumes (Canavalia spp.). Recent Pat. Food Nutr. Agric. 2019, 10, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liu, N.; Ren, H.; Jian, S. Physiology and biochemical characteristics of Canavalia maritime under stress. J. Trop. Subtrop. Bot. 2019, 27, 157–163. [Google Scholar]
- Andrási, N.; Pettkó-Szandtner, A.; Szabados, L. Diversity of plant heat shock factors: Regulation, interactions, and functions. J. Exp. Bot. 2021, 72, 1558–1575. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Shi, Y.; Yang, S. Molecular regulation of plant responses to environmental temperatures. Mol. Plant 2020, 13, 544–564. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Euring, D.; Cha, J.Y.; Lin, Z.; Lu, M.; Huang, L.J.; Kim, W.Y. Plant hormone-mediated regulation of heat tolerance in response to global climate change. Front. Plant Sci. 2021, 11, 627969. [Google Scholar] [CrossRef] [PubMed]
- Medina, E.; Kim, S.H.; Yun, M.; Choi, W.G. Recapitulation of the function and role of ROS generated in response to heat stress in plants. Plants 2021, 10, 371. [Google Scholar] [CrossRef] [PubMed]
- Hayes, S.; Schachtschabel, J.; Mishkind, M.; Munnik, T.; Arisz, S.A. Hot topic: Thermosensing in plants. Plant Cell Environ. 2021, 44, 2018–2033. [Google Scholar] [CrossRef]
- Scharf, K.D.; Berberich, T.; Ebersberger, I.; Nover, L. The plant heat stress transcription factor (Hsf) family: Structure, function and evolution. Biochim. Biophys. Acta 2012, 1819, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Fragkostefanakis, S.; Roeth, S.; Schleiff, E.; Scharf, K.D. Prospects of engineering thermotolerance in crops through modulation of heat stress transcription factor and heat shock protein networks. Plant Cell Environ. 2015, 38, 1881–1895. [Google Scholar] [CrossRef]
- Nover, L.; Bharti, K.; Döring, P.; Mishra, S.K.; Ganguli, A.; Scharf, K.D. Arabidopsis and the heat stress transcription factor world: How many heat stress transcription factors do we need? Cell Stress Chaperones 2001, 6, 177–189. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Ma, X.; Luo, D.X.; Gong, Z.H.; Lu, M.H. The plant heat stress transcription factors (HSFs): Structure, regulation, and function in response to abiotic stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Huang, W.; Liu, J.; Yang, Z.; Huang, B. Molecular regulation and physiological functions of a novel FaHsfA2c cloned from tall fescue conferring plant tolerance to heat stress. Plant Biotechnol. J. 2017, 15, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Liang, J.; Wang, C.; Zhao, X.; Zhong, X.; Cao, X.; Li, G.; He, J.; Yi, M. Overexpression of lily HsfA3s in Arabidopsis confers increased thermotolerance and salt sensitivity via alterations in proline catabolism. J. Exp. Bot. 2018, 69, 2005–2021. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.J.; Chen, D.; Lynne Mclntyre, C.; Fernanda Dreccer, M.; Zhang, Z.B.; Drenth, J.; Kalaipandian, S.; Chang, H.; Xue, G.P. Heat shock factor C2a serves as a proactive mechanism for heat protection in developing grains in wheat via an ABA-mediated regulatory pathway. Plant Cell Environ. 2018, 41, 79–98. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, L.; Cao, W.; Wang, J.; Yu, J.; Yang, Z.; Huang, B. Characterization and functional analysis of FaHsfC1b from Festuca arundinacea conferring heat tolerance in Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, K.; Kajita, T.; Murata, J.I.N.; Tateishi, Y. Phylogeography and genetic structure of Hibiscus tiliaceus─Speciation of a pantropical plant with sea-drifted seeds. Mol. Ecol. 2006, 15, 2871–2881. [Google Scholar] [CrossRef] [PubMed]
- Li, P.S.; Yu, T.F.; He, G.H.; Chen, M.; Zhou, Y.B.; Chai, S.C.; Xu, Z.S.; Ma, Y.Z. Genome-wide analysis of the Hsf family in soybean and functional identification of GmHsf-34 involvement in drought and heat stresses. BMC Genom. 2014, 15, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Chidambaranathan, P.; Jagannadham, P.T.K.; Satheesh, V.; Kohli, D.; Basavarajappa, S.H.; Chellapilla, B.; Kumar, J.; Jain, P.K.; Srinivasan, R. Genome-wide analysis identifies chickpea (Cicer arietinum) heat stress transcription factors (Hsfs) responsive to heat stress at the pod development stage. J. Plant Res. 2018, 131, 525–542. [Google Scholar] [CrossRef]
- Yabuta, Y. Functions of heat shock transcription factors involved in response to photooxidative stresses in Arabidopsis. Biosci. Biotechnol. Biochem. 2016, 80, 1254–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Hu, W.; Qian, Y.; Ren, Q.; Zhang, J. Genome-wide identification, classification and expression analysis of the Hsf and Hsp70 gene families in maize. Gene 2021, 770, 145348. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.L.; Yuan, S.N.; Zhang, H.N.; Zhang, Y.Y.; Zhang, Y.J.; Wang, G.Y.; Li, Y.Q.; Li, G.L. Heat-response patterns of the heat shock transcription factor family in advanced development stages of wheat (Triticum aestivum L.) and thermotolerance-regulation by TaHsfA2-10. BMC Plant Biol. 2020, 20, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Huang, Z.; Ma, R.; Chen, J.; Zhang, Z.; Yrjälä, K. Genome-wide identification and analysis of the heat shock transcription factor family in moso bamboo (Phyllostachys edulis). Sci. Rep. 2021, 11, 1–19. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Chai, M.; Chen, H.; Aslam, M.; Niu, X.; Qin, Y.; Cai, H. Genome-wide identification, classification, and expression analysis of the HSF gene family in pineapple (Ananas comosus). PeerJ 2021, 9, e11329. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Zou, T.; Mei, Q.; Wang, Z.; Zhang, M.; Jian, S. Genome-wide analysis of the late embryogenesis abundant (LEA) and abscisic acid-, stress-, and ripening-induced (ASR) gene superfamily from Canavalia rosea and their roles in salinity/alkaline and drought tolerance. Int. J. Mol. Sci. 2021, 22, 4554. [Google Scholar] [CrossRef]
- Soares-Cavalcanti, N.M.; Belarmino, L.C.; Kido, E.A.; Pandolfi, V.; Marcelino-Guimarães, F.C.; Rodrigues, F.A.; Pereira, G.A.; Benko-Iseppon, A.M. Overall picture of expressed heat shock factors in Glycine max, Lotus japonicus and Medicago truncatula. Genet. Mol. Biol. 2012, 35, 247–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wang, R.; Jin, H.; Ding, Y.; Cai, C. Molecular characterization and expression profile analysis of heat shock transcription factors in mungbean. Front. Genet. 2019, 9, 736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, E.; Kim, K.M.; Lee, J.H. Genome-wide analysis and molecular characterization of heat shock transcription factor family in Glycine max. J. Genet. Genom. 2013, 40, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Guo, Q.; Pang, X.; Zhang, P.; Kong, D.; Liu, J. New insights into evolution of plant heat shock factors (Hsfs) and expression analysis of tea genes in response to abiotic stresses. Plants. 2020, 9, 311. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.; Samraj, S.; Jiménez, J.; Gómez, C.; Liu, T.; Begcy, K. Genome-wide identification of heat shock factors and heat shock proteins in response to UV and high intensity light stress in lettuce. BMC Plant Biol. 2021, 21, 1–20. [Google Scholar] [CrossRef]
- Zhou, L.; Yu, X.; Wang, D.; Li, L.; Zhou, W.; Zhang, Q.; Wang, X.; Ye, S.; Wang, Z. Genome-wide identification, classification and expression profile analysis of the HSF gene family in Hypericum perforatum. PeerJ 2021, 9, e11345. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Yan, L.; Li, H.; Lian, X.; Cheng, J.; Wang, W.; Zheng, X.; Wang, X.; Li, J.; Ye, X.; et al. Genome-wide identification of HSF family in peach and functional analysis of PpHSF5 involvement in root and aerial organ development. PeerJ 2021, 9, e10961. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Atif, R.M.; Azhar, M.T.; Peng, Z.; Li, H.; Qin, G.; Jia, Y.; Pan, Z.; He, S.; Qayyum, A.; et al. Genome wide identification, classification and functional characterization of heat shock transcription factors in cultivated and ancestral cottons (Gossypium spp.). Int. J. Biol. Macromol. 2021, 182, 1507–1527. [Google Scholar] [CrossRef] [PubMed]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, S.; Raza, A.; Iqbal, J.; Shaukat, M.; Mahmood, T. Analyzing the regulatory role of heat shock transcription factors in plant heat stress tolerance: A brief appraisal. Mol. Biol. Rep. 2022, 49, 5771–5785. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wu, J.; Ji, Q.; Wang, C.; Luo, L.; Yuan, Y.; Wang, Y.; Wang, J. Genome-wide analysis of heat shock transcription factor families in rice and Arabidopsis. J. Genet. Genom. 2008, 35, 105–118. [Google Scholar] [CrossRef]
- Zhang, Q.; Geng, J.; Du, Y.; Zhao, Q.; Zhang, W.; Fang, Q.; Yin, Z.; Li, J.; Yuan, X.; Fan, Y.; et al. Heat shock transcription factor (Hsf) gene family in common bean (Phaseolus vulgaris): Genome-wide identification, phylogeny, evolutionary expansion and expression analyses at the sprout stage under abiotic stress. BMC Plant Biol. 2022, 22, 1–15. [Google Scholar] [CrossRef]
- Mallick, B.; Kumari, M.; Pradhan, S.K.; Acharya, G.C.; Naresh, P.; Das, B.; Shashankar, P. Genome-wide analysis and characterization of heat shock transcription factors (Hsfs) in common bean (Phaseolus vulgaris L.). Funct. Integr. Genomics. 2022, 22, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Huang, S.; Qian, J.; Qing, H.; Wan, Z.; Cheng, H.; Zhang, C. Genome-wide identification of petunia HSF genes and potential function of PhHSF19 in benzenoid/phenylpropanoid biosynthesis. Int. J. Mol. Sci. 2022, 23, 2974. [Google Scholar] [CrossRef]
- Bian, X.H.; Li, W.; Niu, C.F.; Wei, W.; Hu, Y.; Han, J.Q.; Lu, X.; Tao, J.J.; Jin, M.; Qin, H.; et al. A class B heat shock factor selected for during soybean domestication contributes to salt tolerance by promoting flavonoid biosynthesis. New Phytol. 2020, 225, 268–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Qi, X.; Zhang, Y.; Zhang, R.; Wang, C.; Sun, T.; Zheng, J.; Lu, Y. Genome-wide analysis of the heat shock transcription factor gene family in Sorbus pohuashanensis (Hance) Hedl identifies potential candidates for resistance to abiotic stresses. Plant Physiol. Biochem. 2022, 175, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Miao, J.; He, J.; Tian, X.; Gao, K.; Ma, C.; Tian, X.; Men, W.; Li, H.; Bi, H.; et al. Wheat heat shock factor TaHsfA2d contributes to plant responses to phosphate deficiency. Plant Physiol. Biochem. 2022, 185, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liu, W.; Yu, L.; Guo, Z.; Chen, Z.; Jiang, S.; Xu, H.; Fang, H.; Wang, Y.; Zhang, Z.; et al. HEAT SHOCK FACTOR A8a modulates flavonoid synthesis and drought tolerance. Plant Physiol. 2020, 184, 1273–1290. [Google Scholar] [CrossRef] [PubMed]
- Shyamli, P.S.; Pradhan, S.; Panda, M.; Parida, A. De novo whole-genome assembly of Moringa oleifera helps identify genes regulating drought stress tolerance. Front. Plant Sci. 2021, 12, 766999. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, W.; Ni, D.; Wang, M.; Guo, G. Genome-wide characterization of tea plant (Camellia sinensis) Hsf transcription factor family and role of CsHsfA2 in heat tolerance. BMC Plant Biol. 2020, 20, 1–17. [Google Scholar] [CrossRef]
- Lavania, D.; Dhingra, A.; Grover, A. Analysis of transactivation potential of rice (Oryza sativa L.) heat shock factors. Planta 2018, 247, 1267–1276. [Google Scholar] [CrossRef]
- Li, S.J.; Liu, S.C.; Lin, X.H.; Grierson, D.; Yin, X.R.; Chen, K.S. Citrus heat shock transcription factor CitHsfA7-mediated citric acid degradation in response to heat stress. Plant Cell Environ. 2022, 45, 95–104. [Google Scholar] [CrossRef]
- Liu, Z.; Li, G.; Zhang, H.; Zhang, Y.; Zhang, Y.; Duan, S.; Sheteiwy, M.S.A.; Zhang, H.; Shao, H.; Guo, X. TaHsfA2-1, a new gene for thermotolerance in wheat seedlings: Characterization and functional roles. J. Plant Physiol. 2020, 246–247, 153135. [Google Scholar] [CrossRef]
- Chen, S.; Yu, M.; Li, H.; Wang, Y.; Lu, Z.; Zhang, Y.; Liu, M.; Qiao, G.; Wu, L.; Han, X.; et al. SaHsfA4c from Sedum alfredii Hance enhances cadmium tolerance by regulating ROS-scavenger activities and heat shock proteins expression. Front. Plant Sci. 2020, 11, 142. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Gojobori, T. Simple methods for estimating the numbers of synonymous and nonsynonymous nucleotide substitutions. Mol. Biol. Evol. 1986, 3, 418–426. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Liu, G.; Chang, Y.; He, C.; Shi, H. Heat shock transcription factor 3 regulates plant immune response through modulation of salicylic acid accumulation and signaling in cassava. Mol. Plant Pathol. 2018, 19, 2209–2220. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene Locus | Class | CDS (bp) | Mw (kDa) | PI | II | GRAVY | Subcellular Localization (WoLF_PSORT) |
|---|---|---|---|---|---|---|---|---|
| CrHsf1 | 01T000116 | A8 | 1104 | 41.85 | 4.9 | 51.75 | −0.678 | nucl: 13.5, cyto_nucl: 7.5 |
| CrHsf2 | 01T000944 | A4 | 1209 | 45.78 | 4.9 | 55.10 | −0.752 | nucl: 14 |
| CrHsf3 | 01T001060 | A5 | 1452 | 54.53 | 5.64 | 61.31 | −0.777 | nucl: 14 |
| CrHsf4 | 01T002065 | A1 | 1533 | 56.16 | 4.89 | 58.37 | −0.637 | nucl: 14 |
| CrHsf5 | 01T002937 | B5 | 588 | 22.39 | 8.23 | 56.00 | −0.598 | nucl: 13, pero: 1 |
| CrHsf6 | 02T004235 | B4 | 1080 | 40.28 | 8.62 | 47.44 | −0.599 | nucl: 14 |
| CrHsf7 | 02T004720 | A2 | 1146 | 42.50 | 5.03 | 66.71 | −0.541 | nucl: 13.5, cyto_nucl: 7.5 |
| CrHsf8 | 03T007873 | B3 | 705 | 26.76 | 8.85 | 58.96 | −0.828 | nucl: 13, pero: 1 |
| CrHsf9 | 03T008076 | A7 | 1179 | 44.72 | 5.23 | 61.06 | −0.669 | cyto: 4.5, E.R.: 3, cyto_pero: 3, nucl: 2, vacu: 2, mito: 1, plas: 1 |
| CrHsf10 | 04T011078 | A6 | 1074 | 41.35 | 4.76 | 62.10 | −0.791 | nucl: 14 |
| CrHsf11 | 04T011116 | A7 | 1047 | 40.66 | 5.58 | 56.11 | −0.798 | nucl: 14 |
| CrHsf12 | 04T011879 | B3 | 699 | 26.88 | 8.82 | 52.24 | −0.711 | nucl: 14 |
| CrHsf13 | 04T013612 | B2 | 1029 | 38.46 | 8.98 | 52.93 | −0.720 | nucl: 14 |
| CrHsf14 | 04T013679 | A6 | 1080 | 41.63 | 5.08 | 49.36 | −0.710 | nucl: 14 |
| CrHsf15 | 05T014277 | A5 | 1281 | 48.70 | 5.34 | 63.89 | −0.854 | nucl: 14 |
| CrHsf16 | 05T014583 | B1 | 849 | 30.92 | 5.61 | 42.86 | −0.668 | nucl: 13, pero: 1 |
| CrHsf17 | 05T015720 | A9 | 1152 | 43.64 | 5.88 | 42.28 | −0.712 | nucl: 14 |
| CrHsf18 | 06T017427 | B4 | 810 | 31.30 | 6.63 | 55.13 | −0.639 | nucl: 14 |
| CrHsf19 | 06T018996 | A2 | 636 | 23.78 | 6.12 | 42.87 | −0.643 | nucl: 14 |
| CrHsf20 | 06T019137 | B4 | 1044 | 39.60 | 7.73 | 51.42 | −0.710 | nucl: 14 |
| CrHsf21 | 07T020710 | B2 | 1008 | 37.00 | 6.27 | 61.01 | −0.645 | nucl: 14 |
| CrHsf22 | 07T021167 | C | 1173 | 43.35 | 6.77 | 48.59 | −0.608 | nucl: 12.5, cyto_nucl: 7, chlo: 1 |
| CrHsf23 | 10T026034 | A1 | 1440 | 53.42 | 5.35 | 55.90 | −0.556 | nucl: 14 |
| CrHsf24 | 10T026239 | B2 | 1044 | 37.90 | 5.16 | 53.51 | −0.517 | nucl: 14 |
| CrHsf25 | 10T026586 | B1 | 834 | 30.97 | 8.98 | 39.60 | −0.928 | nucl: 11, cyto: 1, mito: 1, plas: 1 |
| CrHsf26 | 10T027023 | A1 | 1488 | 54.79 | 4.99 | 64.79 | −0.678 | nucl: 14 |
| CrHsf27 | 11T027981 | B4 | 810 | 31.43 | 7.10 | 59.72 | −0.622 | nucl: 14 |
| CrHsf28 | 11T029135 | A4 | 1197 | 45.65 | 5.06 | 52.51 | −0.764 | nucl: 14 |
| Protein-Class | Length | DBD | HR-A/B | NLS | NES | AHA | RD |
|---|---|---|---|---|---|---|---|
| CrHsf4-A1 | 510 aa | 30–119 | 148–175 | N.D. | N.D. | (447)DLFNNPLFWD | N.D. |
| CrHsf23-A1 | 479 aa | 23–103 | 130–157 | (215)ITGGNKKRRLHRQ | N.D. | (414)DEFWELFFMP | N.D. |
| CrHsf26-A1 | 495 aa | 24–113 | 174–190 | N.D. | N.D. | (431)DDFLSNPSIW | N.D. |
| CrHsf7-A2 | 381 aa | 54–143 | 170–190 | (141)LLKTIKRRRNVT, (250)VRRKRRLTAS | N.D. | (320)GSVWEDLLNQ (360)DDDWTEDLQS | N.D. |
| CrHsf19-A2 | 211 aa | 41–130 | 158–178 | N.D. | N.D. | N.D. | N.D. |
| CrHsf2-A4 | 402 aa | 14–103 | 125–152 | N.D. | (142)L | (254)VAFWEAIAQD (338)DVFWEQFLTE | N.D. |
| CrHsf28-A4 | 398 aa | 14–103 | 120–175 | (208)VDRKRRLPRS | (134)LEKLKHEKEQL | (256)MAFWENLARD (340)DVFWEQFLTE | N.D. |
| CrHsf3-A5 | 483 aa | 18–107 | 125–145 | (208)LSAYNKKRRLPQV | (203)L | (430)DVFWEQFLTE | N.D. |
| CrHsf15-A5 | 426 aa | 9–98 | 116–136, 151–171 | (199)LSAYNKKRRLPQV | (307)L | (386)DMFWEQFLTE | N.D. |
| CrHsf10-A6 | 357 aa | 43–132 | 159–200 | (240)LCKKRRRPID | (285)L, (342)L | (317)EVFWEDFLNE | N.D. |
| CrHsf14-A6 | 359 aa | 46–135 | 164–205 | (245)FSKKRRRPID | (284)LEFEVDL | (322)EVFWQNLLNE | N.D. |
| CrHsf9-A7 | 392 aa | 51–160 | 189–222, 251–271 | (270)MSKKRRRPIE | (197)LVL | (348)EGFWEELFSE | N.D. |
| CrHsf11-A7 | 348 aa | 46–135 | 159–179 | (239)LTKKRRRQIE | (279)L, (281)M | (306)EQFWEEVLFG | N.D. |
| CrHsf1-A8 | 367 aa | 19–108 | 143–170 | N.D. | (178)LQ | (317)SPFLGNVQDS | N.D. |
| CrHsf17-A9 | 383 aa | 68–157 | 168–195 | (231)RMARKPAFVEQLIQKIKRKRELDGNDMDKRPRL | (189)I, (192)L | (278)QGFQSELNGL | N.D. |
| CrHsf16-B1 | 282 aa | 10–99 | 137–171 | (260)NCRKRGRQDPIAAGAKQLKT | (281)I | N.D. | 234 |
| CrHsf25-B1 | 277 aa | 10–99 | 160–180 | N.D. | (185)IAFLKERL | N.D. | 222 |
| CrHsf13-B2 | 342 aa | 48–137 | 202–222 | N.D. | (212)L | N.D. | 278 |
| CrHsf21-B2 | 335 aa | 28–117 | 182–202 | N.D. | (192)L, (201)SL | N.D. | 265 |
| CrHsf24-B2 | 347 aa | 25–114 | 180–207 | N.D. | (190)L, (195)M, (197)L | N.D. | 281 |
| CrHsf8-B3 | 234 aa | 22–111 | 154–174 | (174)TNMKRKCREL, (215)GGRDMKRNRAE | N.D. | N.D. | 201 |
| CrHsf12-B3 | 232 aa | 23–112 | 177–197 | (176)TIMKRKCREL, (215)GEREMKKRRDEI | N.D. | N.D. | 201 |
| CrHsf6-B4 | 359 aa | 25–114 | N.D. | N.D. | N.D. | N.D. | 312 |
| CrHsf18-B4 | 269 aa | 26–115 | N.D. | N.D. | N.D. | N.D. | 249 |
| CrHsf20-B4 | 347 aa | 25–114 | N.D. | N.D. | (338)L | N.D. | 299 |
| CrHsf27-B4 | 269 aa | 26–115 | N.D. | N.D. | N.D. | N.D. | 252 |
| CrHsf5-B5 | 195 aa | 31–124 | 152–182 | N.D. | (167)LELQM | N.D. | N.D. |
| CrHsf22-C | 390 aa | 65–154 | 174–208 | N.D. | (184)LKEEQKALEEEL | N.D. | N.D. |
| Duplicated Pair | Subfamily | Duplicate Type | Ka | Ks | Ka/Ks | p-Value (Fisher) | Positive Selection |
|---|---|---|---|---|---|---|---|
| CrHsf4/CrHsf26 | A1 | Segmental | 0.130461 | 0.519849 | 0.251 | 6.11 × 10−24 | No |
| CrHsf7/CrHsf19 | A2 | Segmental | 0.183841 | 1.25915 | 0.146 | 8.24 × 10−24 | No |
| CrHsf2/CrHsf28 | A4 | Segmental | 0.149982 | 0.613725 | 0.2444 | 4.17 × 10−21 | No |
| CrHsf3/CrHsf15 | A5 | Segmental | 0.169137 | 0.527677 | 0.3205 | 2.42 × 10−15 | No |
| CrHsf9/CrHsf11 | A7 | Segmental | 0.181549 | 0.581786 | 0.3121 | 8.48 × 10−14 | No |
| CrHsf16/CrHsf25 | B1 | Segmental | 0.257367 | 1.21105 | 0.2125 | 8.40 × 10−21 | No |
| CrHsf13/CrHsf21 | B2 | Segmental | 0.159903 | 0.969807 | 0.1649 | 9.10 × 10−32 | No |
| CrHsf8/CrHsf12 | B3 | Segmental | 0.140623 | 0.825046 | 0.1704 | 2.16 × 10−21 | No |
| CrHsf6/CrHsf20 | B4 | Segmental | 0.179072 | 0.98243 | 0.1823 | 4.90 × 10−30 | No |
| CrHsf18/CrHsf27 | B4 | Segmental | 0.101033 | 0.532705 | 0.1897 | 5.81 × 10−17 | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Wang, Z.; Jian, S. Functional Characterization of Heat Shock Factor (CrHsf) Families Provide Comprehensive Insight into the Adaptive Mechanisms of Canavalia rosea (Sw.) DC. to Tropical Coral Islands. Int. J. Mol. Sci. 2022, 23, 12357. https://doi.org/10.3390/ijms232012357
Zhang M, Wang Z, Jian S. Functional Characterization of Heat Shock Factor (CrHsf) Families Provide Comprehensive Insight into the Adaptive Mechanisms of Canavalia rosea (Sw.) DC. to Tropical Coral Islands. International Journal of Molecular Sciences. 2022; 23(20):12357. https://doi.org/10.3390/ijms232012357
Chicago/Turabian StyleZhang, Mei, Zhengfeng Wang, and Shuguang Jian. 2022. "Functional Characterization of Heat Shock Factor (CrHsf) Families Provide Comprehensive Insight into the Adaptive Mechanisms of Canavalia rosea (Sw.) DC. to Tropical Coral Islands" International Journal of Molecular Sciences 23, no. 20: 12357. https://doi.org/10.3390/ijms232012357