
A Temporal Comparative RNA Transcriptome Profile of the Annexin Gene Family in the Salivary versus Lacrimal Glands of the Sjögren’s Syndrome-Susceptible C57BL/6.NOD-Aec1Aec2 Mouse

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
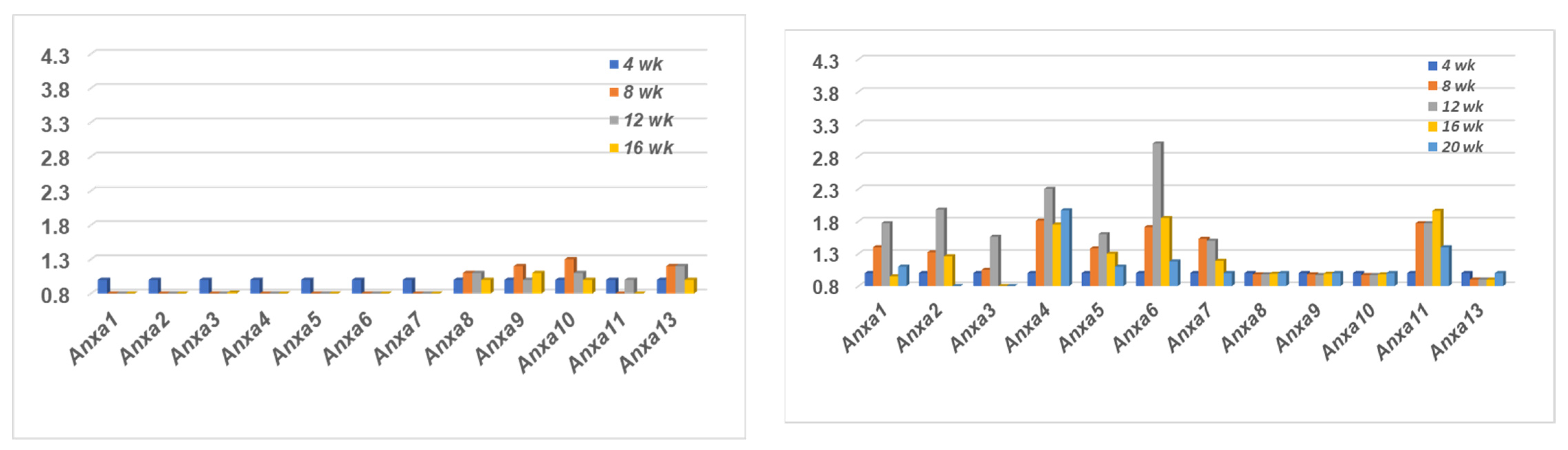
2.1. Comparison of the Annexin Transcriptome Profiles Expressed in the Salivary Glands of SSNS C57BL/6J and SSS C57BL/6.NOD-Aec1Aec2 Mice
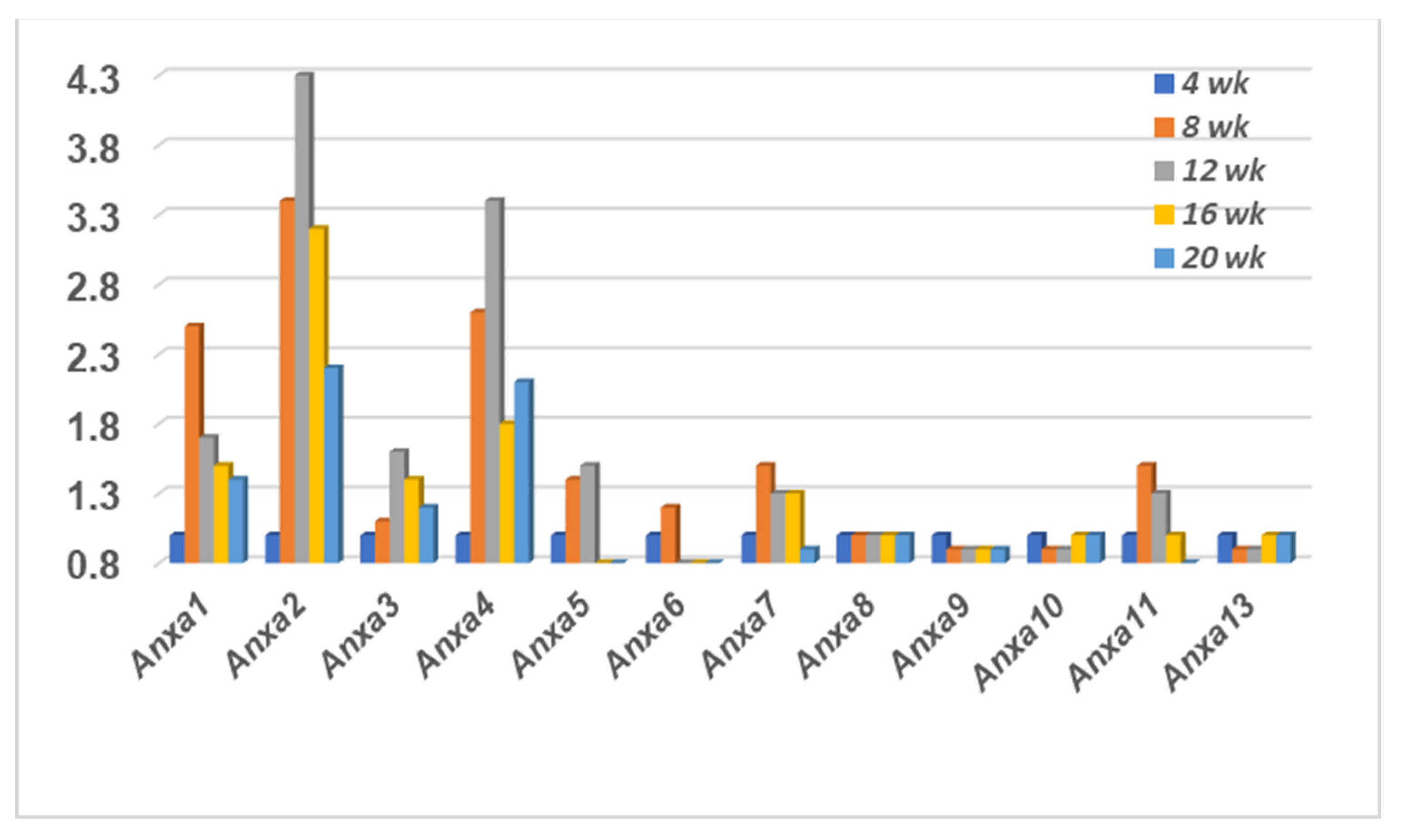
2.2. The Annexin Transcriptome Profile Expressed in the Lacrimal Glands of SSS C57BL/6.NOD-Aec1Aec2 Mice
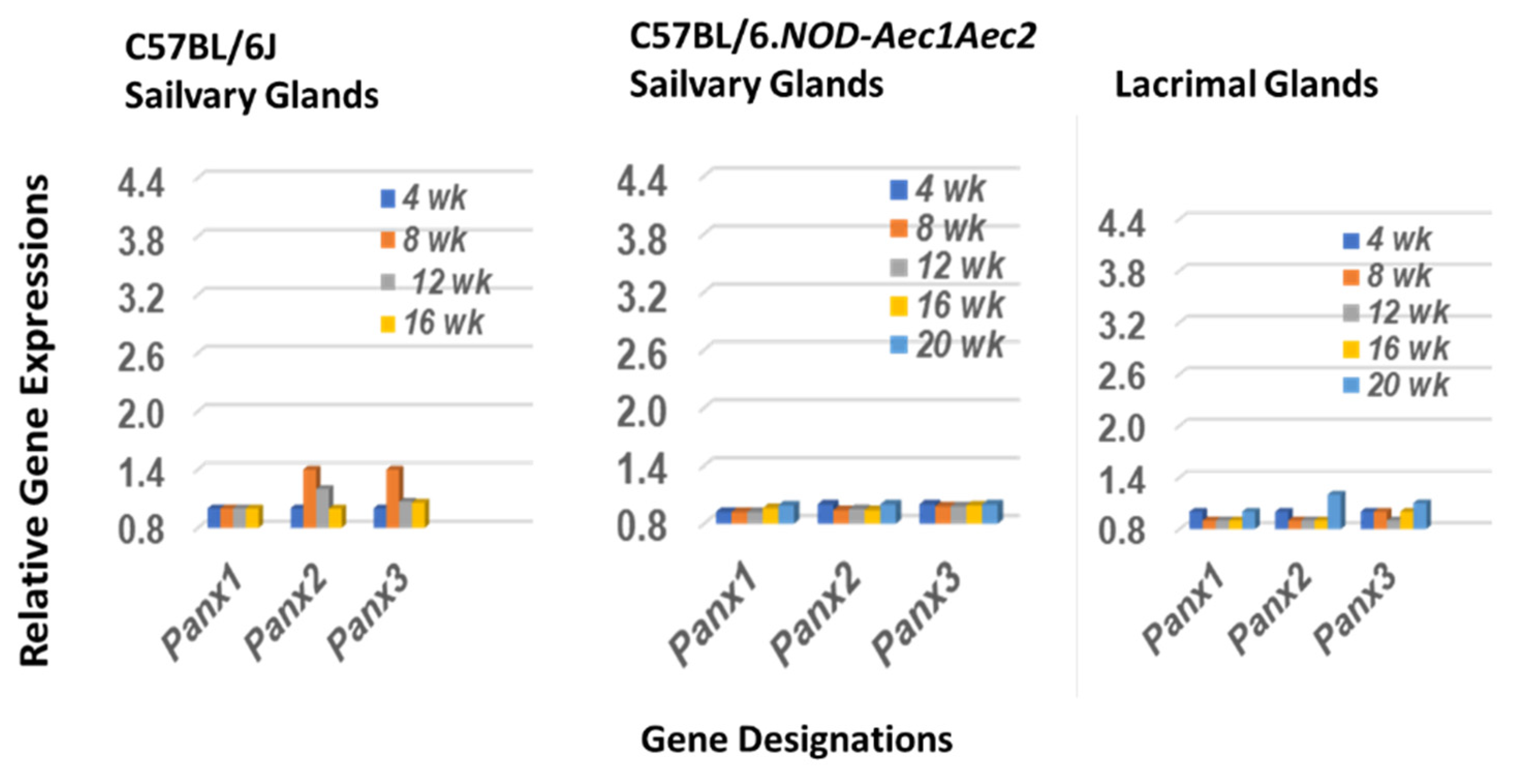
2.3. The Pannexin Transcription Profile in Salivary and Lacrimal Glands of SSNS C57BL/6J and SSS C57BL/6.NOD-Aec1Aec2 Mice during the Early Covert Phase of SS
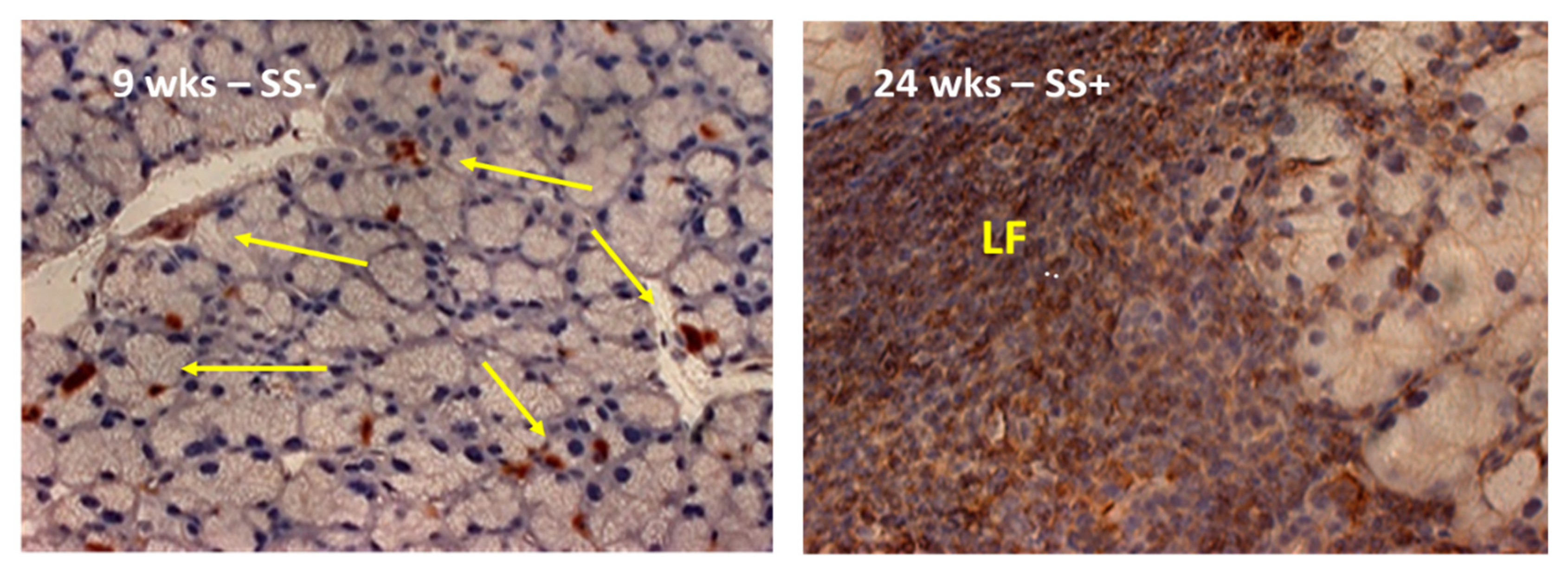
2.4. A Visual Comparison between the Anax Gene Expression Profiling and the Histological Landscape Showing the Distinct Increased Levels of Leukocytic Infiltrations of the Lacrimal Glands between Early-Stage versus Late-Stage Development of the SS-like Disease in the C57BL/6.NOD-Aec1Aec2 Mice
3. Discussion
4. Materials and Methods
4.1. Animal Model
4.2. RNA Preparations
4.3. Microarray Data Analyses
4.4. Histology
5. Conclusions, Summary and Contribution to the Field
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Anxa | Annexin gene |
| Ca++ | Calcium ion |
| DAMPs | Danger-associated molecular patterns |
| dsRNA | double-stranded RNA |
| FRP | Formyl peptide receptor |
| GCMRA | Robust multiarray average |
| GTPase | GTP catalyzing enzymes |
| KO | Knock-out (gene) |
| LF | Lymphocytic focus |
| LIMMA | Linear model for microarray analysis |
| NOD | Non-obese diabetic (mouse) |
| Panx | Pannexin gene |
| PAMPS | Pathogen-assocated molecular patterns |
| PRR | Pathogen recognition receptor |
| pSS-MALT | Primary Sjögren’s syndrome—mucosal-associated lymphoid tissue lymphoma |
| RA | Rheumatoid arthritis |
| RNA | Ribonuclear acid |
| SLE | Systemic l erythmetosus |
| SS | Sjögren’s syndrome |
| SSS | Sjögren’s syndrome—susceptible |
| SSNS | Sjögren’s syndrome—non-susceptible |
References
- Jonsson, R.; Dowman, S.; Gordon, T. Sjogren’s Syndrome. In Arthritis and Allied Conditions—A Textbook in Rheumatology; Koopman, W., Moreland, L., Eds.; Lippincott WIlliams & Wilkins: Philadelphia, PA, USA, 2004; pp. 1681–1705. [Google Scholar]
- Fox, R.I. Sjogren’s syndrome. Lancet 2005, 366, 321–331. [Google Scholar] [CrossRef]
- Routsias, J.G.; Tzioufas, A.G. Sjogren’s syndrome—Study of autoantigens and autoantibodies. Clin. Rev. Allergy Immunol. 2007, 32, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Nordmark, G.; Eloranta, M.L.; Ronnblom, L. Primary Sjogren’s syndrome and the type I interferon system. Curr. Pharm. Biotechnol. 2012, 13, 2054–2062. [Google Scholar] [CrossRef]
- Shiboski, S.C.; Shiboski, C.H.; Criswell, L.A.; Baer, A.N.; Challacombe, S.; Lanfranchi, H.; Schiodt, M.; Umehara, H.; Vivino, F.; Zhao, Y.; et al. American College of Rheumatology classification criteria for Sjogren’s syndrome: A data-driven, expert consensus approach in the Sjogren’s International Collaborative Clinical Alliance Cohort. Arthritis Care Res. 2012, 64, 475–487. [Google Scholar] [CrossRef]
- Shen, L.; Suresh, L.; Lindemann, M.; Xuan, J.; Kowal, P.; Malyavantham, K.; Ambrus, J.L., Jr. Novel autoantibodies in Sjogren’s syndrome. Clin. Immunol. 2012, 145, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Maier-Moore, J.S.; Koelsch, K.A.; Smith, K.; Lessard, C.J.; Radfar, L.; Lewis, D.; Kurien, B.T.; Wolska, N.; Deshmukh, U.; Rasmussen, A.; et al. Antibody-Secreting Cell Specificity in Labial Salivary Glands Reflects the Clinical Presentation and Serology in Patients with Sjogren’s Syndrome. Arthritis Rheumatol. 2014, 66, 3445–3456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goules, A.V.; Tzioufas, A.G.; Moutsopoulos, H.M. Classification criteria of Sjogren’s syndrome. J. Autoimmun. 2014, 48–49, 42–45. [Google Scholar] [CrossRef]
- Reksten, T.R.; Jonsson, M.V. Sjogren’s Syndrome An Update on Epidemiology and Current Insights on Pathophysiology. Oral Maxillofac. Surg. Clin. N. Am. 2014, 26, 1–12. [Google Scholar] [CrossRef]
- Goules, A.V.; Tzioufas, A.G. Primary Sjogren’s syndrome: Clinical phenotypes, outcome and the development of biomarkers. Autoimmun. Rev. 2016, 15, 695–703. [Google Scholar] [CrossRef]
- Harris, V.M.; Scofield, R.H.; Sivils, K.L. Genetics in Sjogren’s syndrome: Where we are and where we go. Clin. Exp. Rheumatol. 2019, 37, S234–S239. [Google Scholar]
- Liang, Y.; Yang, Z.X.; Qin, B.D.; Zhong, R.Q. Primary Sjogren’s syndrome and malignancy risk: A systematic review and meta-analysis. Ann. Rheum. Dis. 2014, 73, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Nezos, A.; Gravani, F.; Tassidou, A.; Kapsogeorgou, E.K.; Voulgarelis, M.; Koutsilieris, M.; Crow, M.K.; Mavragani, C.P. Type I and II interferon signatures in Sjogren’s syndrome pathogenesis: Contributions in distinct clinical phenotypes and Sjogren’s related lymphomagenesis. J. Autoimmun. 2015, 63, 47–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nocturne, G.; Pontarini, E.; Bombardieri, M.; Mariette, X. Lymphomas complicating primary Sjogren’s syndrome: From autoimmunity to lymphoma. Rheumatology 2019, 60, 3513–3521. [Google Scholar] [CrossRef] [PubMed]
- Killedar, S.J.; Eckenrode, S.E.; McIndoe, R.A.; She, J.X.; Nguyen, C.Q.; Peck, A.B.; Cha, S. Early pathogenic events associated with Sjogren’s syndrome (SjS)-like disease of the NOD mouse using microarray analysis. Lab. Investig. 2006, 86, 1243–1260. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.Q.; Sharma, A.; Lee, B.H.; She, J.X.; McIndoe, R.A.; Peck, A.B. Differential gene expression in the salivary gland during development and onset of xerostomia in Sjogren’s syndrome-like disease of the C57BL/6.NOD-Aec1Aec2 mouse. Arthritis Res. Ther. 2009, 11, R56. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.Q.; Sharma, A.; She, J.X.; McIndoe, R.A.; Peck, A.B. Differential gene expressions in the lacrimal gland during development and onset of keratoconjunctivitis sicca in Sjogren’s syndrome (SJS)-like disease of the C57BL/6.NOD-Aec1Aec2 mouse. Exp. Eye Res. 2009, 88, 398–409. [Google Scholar] [CrossRef] [Green Version]
- Peck, A.B.; Nguyen, C.Q. Transcriptome analysis of the interferon-signature defining the autoimmune process of Sjogren’s syndrome. Scand. J. Immunol. 2012, 76, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Delaleu, N.; Nguyen, C.Q.; Tekle, K.M.; Jonsson, R.; Peck, A.B. Transcriptional landscapes of emerging autoimmunity: Transient aberrations in the targeted tissue’s extracellular milieu precede immune responses in Sjogren’s syndrome. Arthritis Res. Ther. 2013, 15, R174. [Google Scholar] [CrossRef] [Green Version]
- Peck, A.B.; Nguyen, C.Q.; Ambrus, J. Early Covert Appearance of Marginal Zone B Cells in Salivary Glands of Sjogren’s Syndrome-Susceptible Mice: Initiators of Subsequent Overt Clinical Disease. Int. J. Mol. Sci. 2021, 22, 1919. [Google Scholar] [CrossRef]
- Peck, A.B.; Nguyen, C.Q.; Ambrus, J.L. Upregulated Chemokine and Rho-GTPase Genes Define Immune Cell Emigration into Salivary Glands of Sjogren’s Syndrome-Susceptible C57BL/6.NOD-Aec1Aec2 Mice. Int. J. Mol. Sci. 2021, 22, 7176. [Google Scholar] [CrossRef]
- Peck, A.B.; Nguyen, C.Q.; Ambrus, J.L. A MZB Cell Activation Profile Present in the Lacrimal Glands of Sjogren’s Syndrome-Susceptible C57BL/6.NOD-Aec1Aec2 Mice Defined by Global RNA Transcriptomic Analyses. Int. J. Mol. Sci. 2022, 23, 6106. [Google Scholar] [CrossRef] [PubMed]
- Jentho, E.; Weis, S. DAMPs and Innate Immune Training. Front. Immunol. 2021, 12, 699563. [Google Scholar] [CrossRef] [PubMed]
- Zorro, M.M.; Aguirre-Gamboa, R.; Mayassi, T.; Ciszewski, C.; Barisani, D.; Hu, S.; Weersma, R.K.; Withoff, S.; Li, Y.; Wijmenga, C.; et al. Tissue alarmins and adaptive cytokine induce dynamic and distinct transcriptional responses in tissue-resident intraepithelial cytotoxic T lymphocytes. J. Autoimmun. 2020, 108, 102422. [Google Scholar] [CrossRef] [PubMed]
- Giovannetti, A.; Straface, E.; Rosato, E.; Casciaro, M.; Pioggia, G.; Gangemi, S. Role of Alarmins in the Pathogenesis of Systemic Sclerosis. Int. J. Mol. Sci. 2020, 21, 4985. [Google Scholar] [CrossRef]
- Sarrand, J.; Baglione, L.; Parisis, D.; Soyfoo, M. The Involvement of Alarmins in the Pathogenesis of Sjogren’s Syndrome. Int. J. Mol. Sci. 2022, 23, 5671. [Google Scholar] [CrossRef]
- Moss, S.E.; Morgan, R.O. The annexins. Genome Biol. 2004, 5, 219. [Google Scholar] [CrossRef] [Green Version]
- Gerke, V.; Creutz, C.E.; Moss, S.E. Annexins: Linking Ca2+ signalling to membrane dynamics. Nat. Rev. Mol. Cell Biol. 2005, 6, 449–461. [Google Scholar] [CrossRef]
- Mirsaeidi, M.; Gidfar, S.; Vu, A.; Schraufnagel, D. Annexins family: Insights into their functions and potential role in pathogenesis of sarcoidosis. J. Transl. Med. 2016, 14, 89. [Google Scholar] [CrossRef] [Green Version]
- Grewal, T.; Rentero, C.; Enrich, C.; Wahba, M.; Raabe, C.A.; Rescher, U. Annexin Animal Models-From Fundamental Principles to Translational Research. Int. J. Mol. Sci. 2021, 22, 3439. [Google Scholar] [CrossRef]
- Moreno-Angarita, A.; Aragon, C.C.; Tobon, G.J. Cathelicidin LL-37: A new important molecule in the pathophysiology of systemic lupus erythematosus. J. Transl. Autoimmun. 2020, 3, 100029. [Google Scholar] [CrossRef]
- Martin, M.; Leffler, J.; Blom, A.M. Annexin A2 and A5 serve as new ligands for C1q on apoptotic cells. J. Biol. Chem. 2012, 287, 33733–33744. [Google Scholar] [CrossRef] [PubMed]
- Pianta, A.; Drouin, E.E.; Crowley, J.T.; Arvikar, S.; Strle, K.; Costello, C.E.; Steere, A.C. Annexin A2 is a target of autoimmune T and B cell responses associated with synovial fibroblast proliferation in patients with antibiotic-refractory Lyme arthritis. Clin. Immunol. 2015, 160, 336–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruschi, M.; Petretto, A.; Vaglio, A.; Santucci, L.; Candiano, G.; Ghiggeri, G.M. Annexin A1 and Autoimmunity: From Basic Science to Clinical Applications. Int. J. Mol. Sci. 2018, 19, 1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, K.; Li, Y.; Zhao, Q.; Fan, L.; Tan, B.; Gu, J. Downregulation of Annexin A11 (ANXA11) Inhibits Cell Proliferation, Invasion, and Migration via the AKT/GSK-3beta Pathway in Gastric Cancer. Med. Sci. Monit. 2018, 24, 149–160. [Google Scholar] [CrossRef]
- Lim, H.I.; Hajjar, K.A. Annexin A2 in Fibrinolysis, Inflammation and Fibrosis. Int. J. Mol. Sci. 2021, 22, 6836. [Google Scholar] [CrossRef]
- Iaccarino, L.; Ghirardello, A.; Canova, M.; Zen, M.; Bettio, S.; Nalotto, L.; Punzi, L.; Doria, A. Anti-annexins autoantibodies: Their role as biomarkers of autoimmune diseases. Autoimmun. Rev. 2011, 10, 553–558. [Google Scholar] [CrossRef]
- van Venrooij, W.J.; Wodzig, K.W.; Habets, W.J.; de Rooij, D.J.; van de Putte, L.B. Anti-56K: A novel, frequently occurring autoantibody specificity in connective tissue disease. Clin. Exp. Rheumatol. 1989, 7, 277–282. [Google Scholar]
- Rodriguez-Garcia, M.I.; Fernandez, J.A.; Rodriguez, A.; Fernandez, M.P.; Gutierrez, C.; Torre-Alonso, J.C. Annexin V autoantibodies in rheumatoid arthritis. Ann. Rheum. Dis. 1996, 55, 895–900. [Google Scholar] [CrossRef] [Green Version]
- Gheita, T.A.; El Sisi, R.W.; Raafat, H.A.; Khalil, H.M. Anti-annexin V antibodies in primary fibromyalgia syndrome: Relation to associated Sjogren’s syndrome. J. Clin. Immunol. 2013, 33, 311–312. [Google Scholar] [CrossRef]
- Dema, B.; Charles, N. Autoantibodies in SLE: Specificities, Isotypes and Receptors. Antibodies 2016, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Elzakra, N.; Xu, S.; Xiao, G.G.; Yang, Y.; Hu, S. Investigation of three potential autoantibodies in Sjogren’s syndrome and associated MALT lymphoma. Oncotarget 2017, 8, 30039–30049. [Google Scholar] [CrossRef] [PubMed]
- Scholz, P.; Auler, M.; Ruthard, J.; Brachvogel, B.; Klatt, A.R.; Streichert, T. Detection of annexin A8 antibodies in serum of patients with antiphospholipid syndrome. Biochem. Med. 2018, 28, 030703. [Google Scholar] [CrossRef] [PubMed]
- Finamore, F.; Cecchettini, A.; Ceccherini, E.; Signore, G.; Ferro, F.; Rocchiccioli, S.; Baldini, C. Characterization of Extracellular Vesicle Cargo in Sjogren’s Syndrome through a SWATH-MS Proteomics Approach. Int. J. Mol. Sci. 2021, 22, 4864. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.; Nagashima, H.; Brown, V.B.; Peck, A.B.; Humphreys-Beher, M.G. Two NOD Idd-associated intervals contribute synergistically to the development of autoimmune exocrinopathy (Sjogren’s syndrome) on a healthy murine background. Arthritis Rheum. 2002, 46, 1390–1398. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.Q.; Peck, A.B. Unraveling the pathophysiology of Sjogren syndrome-associated dry eye disease. Ocul. Surf. 2009, 7, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Horvath, S.; Nazmul-Hossain, A.N.; Pollard, R.P.; Kroese, F.G.; Vissink, A.; Kallenberg, C.G.; Spijkervet, F.K.; Bootsma, H.; Michie, S.A.; Gorr, S.U.; et al. Systems analysis of primary Sjogren’s syndrome pathogenesis in salivary glands identifies shared pathways in human and a mouse model. Arthritis Res. Ther. 2012, 14, R238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peck, A.B.; Nguyen, C.Q. What can Sjogren’s syndrome-like disease in mice contribute to human Sjogren’s syndrome? Clin. Immunol. 2017, 182, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Bond, S.R.; Naus, C.C. The pannexins: Past and present. Front. Physiol. 2014, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Peck, A.B.; Saylor, B.T.; Nguyen, L.; Sharma, A.; She, J.X.; Nguyen, C.Q.; McIndoe, R.A. Gene expression profiling of early-phase Sjögren’s syndrome in C57BL/6.NOD-Aec1Aec2 mice identifies focal adhesion maturation associated with infiltrating leukocytes. Investig. Ophthalmol. Vis. Sci. 2011, 29, 5647–5655. [Google Scholar] [CrossRef] [Green Version]
- Humphreys-Beher, M.G.; Peck, A.B.; Dang, H.; Talal, N. The role of apoptosis in the initiation of the autoimmune response in Sjogren’s syndrome. Clin. Exp. Immunol. 1999, 116, 383–387. [Google Scholar] [CrossRef]
- Ernst, S.; Lange, C.; Wilbers, A.; Goebeler, V.; Gerke, V.; Rescher, U. An annexin 1 N-terminal peptide activates leukocytes by triggering different members of the formyl peptide receptor family. J. Immunol. 2004, 172, 7669–7676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavins, F.N.; Hickey, M.J. Annexin A1 and the regulation of innate and adaptive immunity. Front. Immunol. 2012, 3, 354. [Google Scholar] [CrossRef] [PubMed]
- Perretti, M.; D’Acquisto, F. Annexin A1 and glucocorticoids as effectors of the resolution of inflammation. Nat. Rev. Immunol. 2009, 9, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Hannon, R.; Croxtall, J.D.; Getting, S.J.; Roviezzo, F.; Yona, S.; Paul-Clark, M.J.; Gavins, F.N.; Perretti, M.; Morris, J.F.; Buckingham, J.C.; et al. Aberrant inflammation and resistance to glucocorticoids in annexin 1-/- mouse. FASEB J. 2003, 17, 253–255. [Google Scholar] [CrossRef] [Green Version]
- Arora, S.; Lim, W.; Bist, P.; Perumalsamy, R.; Lukman, H.M.; Li, F.; Welker, L.B.; Yan, B.; Sethi, G.; Tambyah, P.A.; et al. Influenza A virus enhances its propagation through the modulation of Annexin-A1 dependent endosomal trafficking and apoptosis. Cell Death Differ. 2016, 23, 1243–1256. [Google Scholar] [CrossRef]
- Nguyen, C.Q.; Peck, A.B. The Interferon-Signature of Sjogren’s Syndrome: How Unique Biomarkers Can Identify Underlying Inflammatory and Immunopathological Mechanisms of Specific Diseases. Front. Immunol. 2013, 4, 142. [Google Scholar] [CrossRef] [Green Version]
- Seidah, N.G.; Poirier, S.; Denis, M.; Parker, R.; Miao, B.; Mapelli, C.; Prat, A.; Wassef, H.; Davignon, J.; Hajjar, K.A.; et al. Annexin A2 is a natural extrahepatic inhibitor of the PCSK9-induced LDL receptor degradation. PLoS ONE 2012, 7, e41865. [Google Scholar] [CrossRef] [Green Version]
- Bharadwaj, A.; Bydoun, M.; Holloway, R.; Waisman, D. Annexin A2 heterotetramer: Structure and function. Int. J. Mol. Sci. 2013, 14, 6259–6305. [Google Scholar] [CrossRef] [Green Version]
- Rescher, U.; Gerke, V. S100A10/p11: Family, friends and functions. Pflug. Arch. 2008, 455, 575–578. [Google Scholar] [CrossRef]
- Hessner, F.; Dlugos, C.P.; Chehab, T.; Schaefer, C.; Homey, B.; Gerke, V.; Weide, T.; Pavenstadt, H.; Rescher, U. CC chemokine receptor 10 cell surface presentation in melanocytes is regulated by the novel interaction partner S100A10. Sci. Rep. 2016, 6, 22649. [Google Scholar] [CrossRef]
- Zhang, S.; Yu, M.; Guo, Q.; Li, R.; Li, G.; Tan, S.; Li, X.; Wei, Y.; Wu, M. Annexin A2 binds to endosomes and negatively regulates TLR4-triggered inflammatory responses via the TRAM-TRIF pathway. Sci. Rep. 2015, 5, 15859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, C.Q.; Hu, M.H.; Li, Y.; Stewart, C.; Peck, A.B. Salivary gland tissue expression of interleukin-23 and interleukin-17 in Sjogren’s syndrome: Findings in humans and mice. Arthritis Rheum. 2008, 58, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Tomas, A.; Moss, S.E. Calcium- and cell cycle-dependent association of annexin 11 with the nuclear envelope. J. Biol. Chem. 2003, 278, 20210–20216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomas, A.; Futter, C.; Moss, S.E. Annexin 11 is required for midbody formation and completion of the terminal phase of cytokinesis. J. Cell Biol. 2004, 165, 813–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutboul, D.; Vince, N.; Mahevas, M.; Bories, J.C.; Fieschi, C.; Def, I.S.G. TNFA, ANXA11 and BTNL2 Polymorphisms in CVID Patients with Granulomatous Disease. J. Clin. Immunol. 2016, 36, 110–112. [Google Scholar] [CrossRef]
- Zhou, H.; Diao, M.; Zhang, M. The Association between ANXA11 Gene Polymorphisms and Sarcoidosis: A Meta-Analysis and systematic review. Sarcoidosis Vasc. Diffus. Lung Dis. 2016, 33, 102–111. [Google Scholar]
- Hofmann, S.; Franke, A.; Fischer, A.; Jacobs, G.; Nothnagel, M.; Gaede, K.I.; Schurmann, M.; Muller-Quernheim, J.; Krawczak, M.; Rosenstiel, P.; et al. Genome-wide association study identifies ANXA11 as a new susceptibility locus for sarcoidosis. Nat. Genet. 2008, 40, 1103–1106. [Google Scholar] [CrossRef]
- Grewal, T.; Hoque, M.; Conway, J.R.W.; Reverter, M.; Wahba, M.; Beevi, S.S.; Timpson, P.; Enrich, C.; Rentero, C. Annexin A6-A multifunctional scaffold in cell motility. Cell Adhes. Migr. 2017, 11, 288–304. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Kien, F.; Maniere, M.; Zhang, Y.; Lagarde, N.; Tse, K.S.; Poon, L.L.; Nal, B. Human annexin A6 interacts with influenza a virus protein M2 and negatively modulates infection. J. Virol. 2012, 86, 1789–1801. [Google Scholar] [CrossRef] [Green Version]
- Musiol, A.; Gran, S.; Ehrhardt, C.; Ludwig, S.; Grewal, T.; Gerke, V.; Rescher, U. Annexin A6-balanced late endosomal cholesterol controls influenza A replication and propagation. mBio 2013, 4, e00608-13. [Google Scholar] [CrossRef] [Green Version]
- Kuhnl, A.; Musiol, A.; Heitzig, N.; Johnson, D.E.; Ehrhardt, C.; Grewal, T.; Gerke, V.; Ludwig, S.; Rescher, U. Late Endosomal/Lysosomal Cholesterol Accumulation Is a Host Cell-Protective Mechanism Inhibiting Endosomal Escape of Influenza A Virus. mBio 2018, 9, e01345-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, D.M.; Moss, S.E.; Wright, N.A.; Crumpton, M.J. Expression of annexin VI (p68, 67 kDa-calelectrin) in normal human tissues: Evidence for developmental regulation in B- and T-lymphocytes. Histochemistry 1991, 96, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Enrich, C.; Rentero, C.; de Muga, S.V.; Reverter, M.; Mulay, V.; Wood, P.; Koese, M.; Grewal, T. Annexin A6-Linking Ca2+ signaling with cholesterol transport. Biochim. Biophys. Acta 2011, 1813, 935–947. [Google Scholar] [CrossRef]
- Li, B.; Dedman, J.R.; Kaetzel, M.A. Intron disruption of the annexin IV gene reveals novel transcripts. J. Biol. Chem. 2003, 278, 43276–43283. [Google Scholar] [CrossRef] [Green Version]
- Heinick, A.; Pluteanu, F.; Hermes, C.; Klemme, A.; Domnik, M.; Husser, X.; Gerke, V.; Schmitz, W.; Muller, F.U. Annexin A4 N-terminal peptide inhibits adenylyl cyclase 5 and limits beta-adrenoceptor-mediated prolongation of cardiac action potential. FASEB J. 2020, 34, 10489–10504. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Enomoto, T.; Serada, S.; Yoshino, K.; Nagamori, S.; Morimoto, A.; Yokoyama, T.; Kim, A.; Kimura, T.; Ueda, Y.; et al. Annexin A4-conferred platinum resistance is mediated by the copper transporter ATP7A. Int. J. Cancer 2014, 134, 1796–1809. [Google Scholar] [CrossRef] [PubMed]
- Kuijpers, G.A.; Lee, G.; Pollard, H.B. Immunolocalization of synexin (annexin VII) in adrenal chromaffin granules and chromaffin cells: Evidence for a dynamic role in the secretory process. Cell Tissue Res. 1992, 269, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Gerelsaikhan, T.; Vasa, P.K.; Chander, A. Annexin A7 and SNAP23 interactions in alveolar type II cells and in vitro: A role for Ca2+ and PKC. Biochim. Biophys. Acta 2012, 1823, 1796–1806. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Liu, S.; Greenaway, F.; Sun, M.Z. Potential role of annexin A7 in cancers. Clin. Chim. Acta 2013, 423, 83–89. [Google Scholar] [CrossRef]
- Lin, Q.S.; Wang, W.X.; Lin, Y.X.; Lin, Z.Y.; Yu, L.H.; Kang, Y.; Kang, D.Z. Annexin A7 induction of neuronal apoptosis via effect on glutamate release in a rat model of subarachnoid hemorrhage. J. Neurosurg. 2019, 132, 777–787. [Google Scholar] [CrossRef]
- Li, H.; Liu, N.; Wang, S.; Wang, L.; Zhao, J.; Su, L.; Zhang, Y.; Zhang, S.; Xu, Z.; Zhao, B.; et al. Identification of a small molecule targeting annexin A7. Biochim. Biophys. Acta 2013, 1833, 2092–2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boersma, H.H.; Kietselaer, B.L.; Stolk, L.M.; Bennaghmouch, A.; Hofstra, L.; Narula, J.; Heidendal, G.A.; Reutelingsperger, C.P. Past, present, and future of annexin A5: From protein discovery to clinical applications. J. Nucl. Med. 2005, 46, 2035–2050. [Google Scholar] [PubMed]
- Dubois, T.; Mira, J.P.; Feliers, D.; Solito, E.; Russo-Marie, F.; Oudinet, J.P. Annexin V inhibits protein kinase C activity via a mechanism of phospholipid sequestration. Biochem. J. 1998, 330 Pt 3, 1277–1282. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, T.E.; Das, D.; Young, B.; Moss, S.E. DT40 cells lacking the Ca2+-binding protein annexin 5 are resistant to Ca2+-dependent apoptosis. Proc. Natl. Acad. Sci. USA 2002, 99, 8054–8059. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Xu, J.; Kirsch, T. Annexin-mediated Ca2+ influx regulates growth plate chondrocyte maturation and apoptosis. J. Biol. Chem. 2003, 278, 3762–3769. [Google Scholar] [CrossRef] [Green Version]
- Monastyrskaya, K.; Babiychuk, E.B.; Hostettler, A.; Rescher, U.; Draeger, A. Annexins as intracellular calcium sensors. Cell Calcium 2007, 41, 207–219. [Google Scholar] [CrossRef]
- Faria, D.; Dahimene, S.; Alessio, L.; Scott-Ward, T.; Schreiber, R.; Kunzelmann, K.; Amaral, M.D. Effect of Annexin A5 on CFTR: Regulated traffic or scaffolding? Mol. Membr. Biol. 2011, 28, 14–29. [Google Scholar] [CrossRef]
- Ghislat, G.; Aguado, C.; Knecht, E. Annexin A5 stimulates autophagy and inhibits endocytosis. J. Cell Sci. 2012, 125 Pt 1, 92–107. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Munoz, L.E.; Mallavarapu, M.; Herrmann, M.; Finnemann, S.C. Annexin A5 regulates surface αvβ5 integrin for retinal clearance phagocytosis. J. Cell Sci. 2019, 132, jcs232439. [Google Scholar] [CrossRef]
- Bode, K.; Bujupi, F.; Link, C.; Hein, T.; Zimmermann, S.; Peiris, D.; Jaquet, V.; Lepenies, B.; Weyd, H.; Krammer, P.H. Dectin-1 Binding to Annexins on Apoptotic Cells Induces Peripheral Immune Tolerance via NADPH Oxidase-2. Cell Rep. 2019, 29, 4435–4446.e9. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, A.; Raguenes-Nicol, C.; Favier-Perron, B.; Mesonero, J.; Huber, R.; Russo-Marie, F.; Lewit-Bentley, A. The annexin A3-membrane interaction is modulated by an N-terminal tryptophan. Biochemistry 2000, 39, 7712–7721. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Liu, B.; Zhou, L.; Wang, D.; He, X.; Xu, X.; Zhang, L.; Niu, C.; Liu, S. Downregulation of annexin A3 inhibits tumor metastasis and decreases drug resistance in breast cancer. Cell Death Dis. 2018, 9, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.Z.; Pan, K.; Wang, Q.J.; Weng, D.S.; Zhao, J.J.; Zheng, H.X.; Zhang, X.F.; Jiang, S.S.; Lv, L.; Tang, Y.; et al. Annexin A3 as a potential target for immunotherapy of liver cancer stem-like cells. Stem Cells 2015, 33, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Ito, Y.; Sato, A.; Hosono, T.; Niimi, S.; Ariga, T.; Seki, T. Annexin A3 as a negative regulator of adipocyte differentiation. J. Biochem. 2012, 152, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Meadows, S.M.; Cleaver, O. Annexin A3 Regulates Early Blood Vessel Formation. PLoS ONE 2015, 10, e0132580. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Crist, A.M.; Patel, N.R.; Blanks, A.; Carter, K.; Cleaver, O.; Meadows, S.M. Annexin A3 is necessary for parallel artery-vein alignment in the mouse retina. Dev. Dyn. 2020, 249, 666–678. [Google Scholar] [CrossRef]
- Tong, M.; Che, N.; Zhou, L.; Luk, S.T.; Kau, P.W.; Chai, S.; Ngan, E.S.; Lo, C.M.; Man, K.; Ding, J.; et al. Efficacy of annexin A3 blockade in sensitizing hepatocellular carcinoma to sorafenib and regorafenib. J. Hepatol. 2018, 69, 826–839. [Google Scholar] [CrossRef]
- Zhang, Z.; Deng, M.; Huang, J.; Wu, J.; Li, Z.; Xing, M.; Wang, J.; Guo, Q.; Zou, W. Microglial annexin A3 downregulation alleviates bone cancer-induced pain through inhibiting the Hif-1alpha/vascular endothelial growth factor signaling pathway. Pain 2020, 161, 2750–2762. [Google Scholar] [CrossRef]
- Malkov, M.I.; Lee, C.T.; Taylor, C.T. Regulation of the Hypoxia-Inducible Factor (HIF) by Pro-Inflammatory Cytokines. Cells 2021, 10, 2340. [Google Scholar] [CrossRef]
- Xing, J.; Lu, J. HIF-1alpha Activation Attenuates IL-6 and TNF-alpha Pathways in Hippocampus of Rats Following Transient Global Ischemia. Cell. Physiol. Biochem. 2016, 39, 511–520. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B-Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peck, A.B.; Ambrus, J.L., Jr. A Temporal Comparative RNA Transcriptome Profile of the Annexin Gene Family in the Salivary versus Lacrimal Glands of the Sjögren’s Syndrome-Susceptible C57BL/6.NOD-Aec1Aec2 Mouse. Int. J. Mol. Sci. 2022, 23, 11709. https://doi.org/10.3390/ijms231911709
Peck AB, Ambrus JL Jr. A Temporal Comparative RNA Transcriptome Profile of the Annexin Gene Family in the Salivary versus Lacrimal Glands of the Sjögren’s Syndrome-Susceptible C57BL/6.NOD-Aec1Aec2 Mouse. International Journal of Molecular Sciences. 2022; 23(19):11709. https://doi.org/10.3390/ijms231911709
Chicago/Turabian StylePeck, Ammon B., and Julian L. Ambrus, Jr. 2022. "A Temporal Comparative RNA Transcriptome Profile of the Annexin Gene Family in the Salivary versus Lacrimal Glands of the Sjögren’s Syndrome-Susceptible C57BL/6.NOD-Aec1Aec2 Mouse" International Journal of Molecular Sciences 23, no. 19: 11709. https://doi.org/10.3390/ijms231911709