
Cytokines and Immune Cells Profile in Different Tissues of Rodents Induced by Environmental Enrichment: Systematic Review
 , , and
, , and
Abstract
:1. Introduction
2. Methods
2.1. Search Strategy
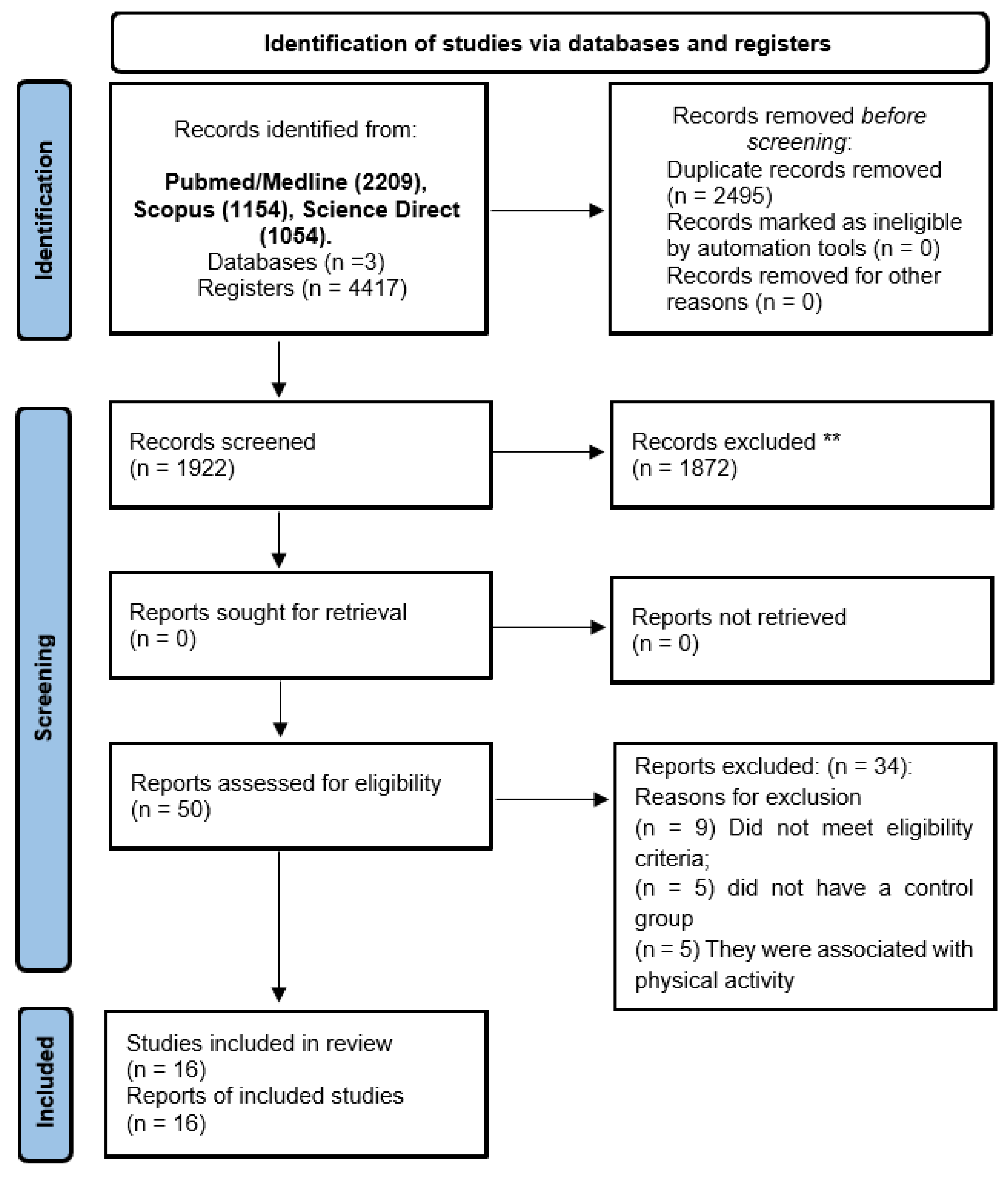
2.2. Selection of Articles
2.3. Quality Assessment
3. Results and Discussion
3.1. Characterization of Included Studies
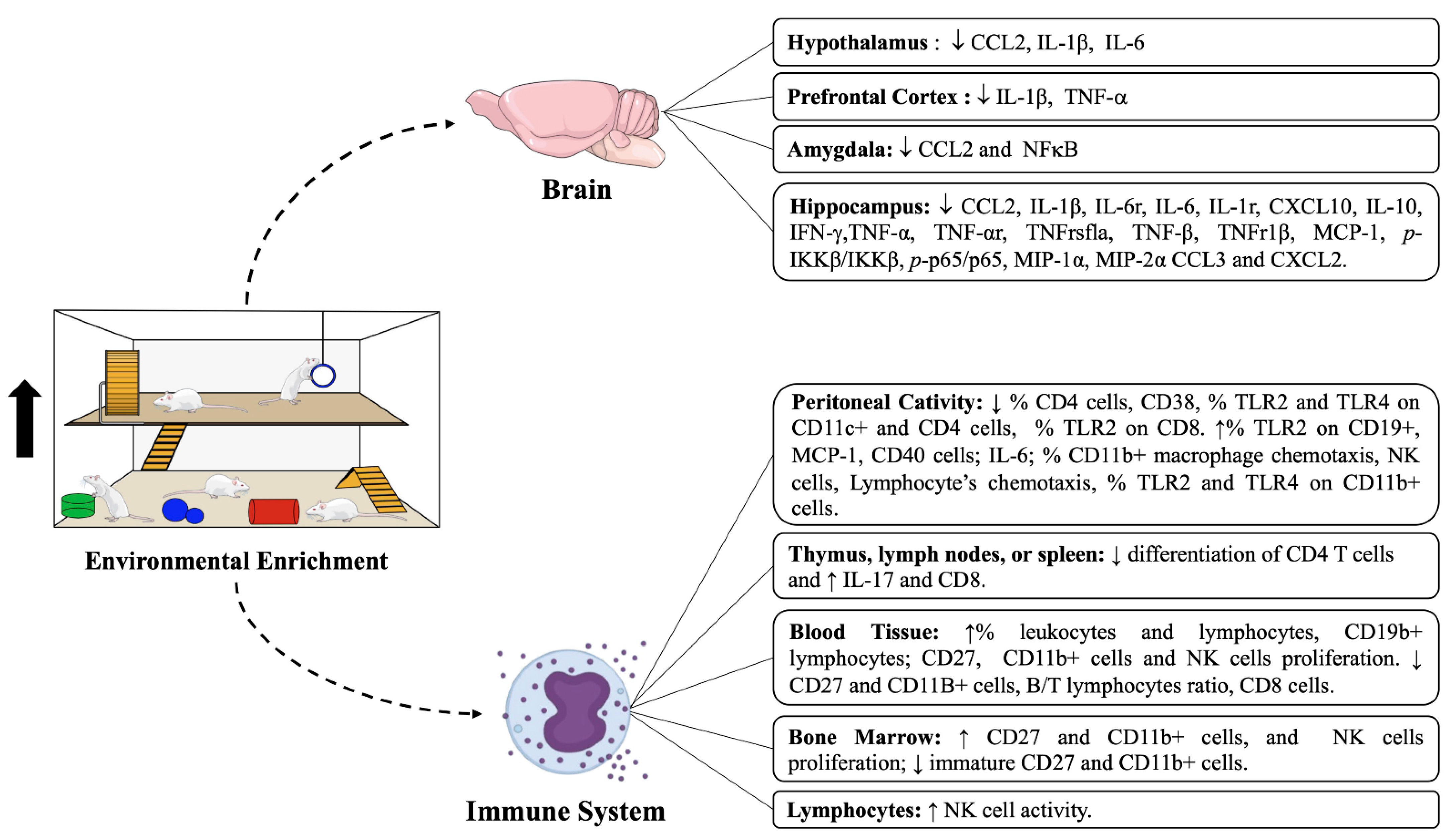
3.2. Cytokine Outcomes from Cells or Tissues after EE Intervention
3.2.1. Pro/Anti-Inflammatory Cytokines Levels and Their Receptors in Different Brain Regions
3.2.2. Pro/Anti-Inflammatory Cytokines Levels in the Peritoneal Cavity, Spleen, and Lymphocytes
3.3. Immune Cell Compartment after Intervention with EE in Different Tissues
3.3.1. Hippocampus and Peritoneal Cavity
3.3.2. Blood
3.3.3. Bone Marrow and Lymphoid Tissues
3.4. Methodological Quality of Studies
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hebb, D.O. The effects of early experience on problem-solving at maturity. Am. Psychol. 1947, 2, 306–307. [Google Scholar]
- Crofton, E.J.; Zhang, Y.; Green, T.A. Inoculation stress hypothesis of environmental enrichment. Neurosci. Biobehav. Rev. 2015, 49, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Ismail, T.R.; Yap, C.G.; Naidu, R.; Pamidi, N. Enrichment Protocol for Rat Models. Curr. Protoc. 2021, 1, e152. [Google Scholar] [CrossRef]
- Webster, A.J. Environmental stress and the physiology, performance and health of ruminants. J. Anim. Sci. 1983, 57, 1584–1593. [Google Scholar] [CrossRef]
- Ashokan, A.; Hegde, A.; Balasingham, A.; Mitra, R. Housing environment influences stress-related hippocampal substrates and depression-like behavior. Brain Res. 2018, 1683, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Zefferino, R.; Di Gioia, S.; Conese, M. Molecular links between endocrine, nervous and immune system during chronic stress. Brain Behav. 2021, 11, e01960. [Google Scholar] [CrossRef] [PubMed]
- Parkin, J.; Cohen, B. An overview of the immune system. Lancet 2001, 357, 1777–1789. [Google Scholar] [CrossRef]
- McComb, S.; Thiriot, A.; Akache, B.; Krishnan, L.; Stark, F. Introduction to the Immune System. Methods Mol. Biol. 2019, 2024, 1–24. [Google Scholar]
- Sattler, S. The Role of the Immune System Beyond the Fight Against Infection. Adv. Exp. Med. Biol. 2017, 1003, 3–14. [Google Scholar]
- Takai, D.; Abe, A.; Miura, H.; Tanaka, S.; Komura, J.I. Minimum environmental enrichment is effective in activating antitumor immunity to transplanted tumor cells in mice. Exp. Anim. 2019, 68, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Woo, C.C.; Leon, M. Environmental enrichment as an effective treatment for autism: A randomized controlled trial. Behav. Neurosci. 2013, 127, 487–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Giorgio, A. the roles of motor activity and environmental enrichment in intellectual disability. Somatosens. Mot. Res. 2017, 34, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Liu, X.; Queen, N.J.; Patel, R.S.; Wilkins, R.K.; Mo, X.; Cao, L. Long-term environmental enrichment affects microglial morphology in middle age mice. Aging 2019, 11, 2388. [Google Scholar] [CrossRef]
- Arranz, L.; De Castro, N.M.; Baeza, I.; Maté, I.; Viveros, M.P.; De la Fuente, M. Environmental enrichment improves age-related immune system impairment: Long-term exposure since adulthood increases life span in mice. Rejuvenation Res. 2010, 13, 415–428. [Google Scholar] [CrossRef] [Green Version]
- Brod, S.; Gobbetti, T.; Gittens, B.; Ono, M.; Perretti, M.; D’Acquisto, F. The impact of environmental enrichment on the murine inflammatory immune response. JCI Insight 2017, 2, e90723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griñan-Ferré, C.; Puigoriol-Illamola, D.; Palomera-Ávalos, V.; Pérez-Cáceres, D.; Companys-Alemany, J.; Camins, A.; Ortuño-Sahagún, D.; Rodrigo, M.T.; Pallàs, M. Environmental enrichment modified epigenetic mechanisms in SAMP8 mouse hippocampus by reducing oxidative stress and inflammaging and achieving neuroprotection. Front. Aging Neurosci. 2016, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Gurfein, B.T.; Hasdemir, B.; Milush, J.M.; Touma, C.; Palme, R.; Nixon, D.F.; Darcel, N.; Hecht, F.M.; Bhargava, A. Enriched environment and stress exposure influence splenic B lymphocyte composition. PLoS ONE 2017, 12, e0180771. [Google Scholar] [CrossRef] [Green Version]
- Keymoradzadeh, A.; Ch, M.H.; Abedinzade, M.; Gazor, R.; Rostampour, M.; Taleghani, B.K. Enriched environment effect on lipopolysaccharide-induced spatial learning, memory impairment and hippocampal inflammatory cytokine levels in male rats. Behav. Brain Res. 2020, 394, 112814. [Google Scholar] [CrossRef]
- Marashi, V.; Barnekow, A.; Ossendorf, E.; Sachser, N. Effects of different forms of environmental enrichment on behavioral, endocrinological, and immunological parameters in male mice. Horm. Behav. 2003, 43, 281–292. [Google Scholar] [CrossRef]
- McQuaid, R.J.; Audet, M.C.; Jacobson-Pick, S.; Anisman, H. Environmental enrichment influences brain cytokine variations elicited by social defeat in mice. Psychoneuroendocrinology 2013, 38, 987–996. [Google Scholar] [CrossRef]
- Meng, Z.; Liu, T.; Song, Y.; Wang, Q.; Xu, D.; Jiang, J.; Li, M.; Qiao, J.; Luo, X.; Gu, J.; et al. Exposure to an enriched environment promotes the terminal maturation and proliferation of natural killer cells in mice. Brain Behav. Immun. 2019, 77, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Rattazzi, L.; Piras, G.; Brod, S.; Smith, K.; Ono, M.; D’Acquisto, F. Impact of enriched environment on murine T cell differentiation and gene expression profile. Front. Immunol. 2016, 7, 381. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Cao, M.; Pu, T.; Huang, H.; Marshall, C.; Xiao, M. Enriched physical environment attenuates spatial and social memory impairments of aged socially isolated mice. Int. J. Neuropsychopharmacol. 2018, 21, 1114–1127. [Google Scholar] [CrossRef] [PubMed]
- Williamson, L.L.; Chao, A.; Bilbo, S.D. Environmental enrichment alters glial antigen expression and neuroimmune function in the adult rat hippocampus. Brain Behav. Immun. 2012, 26, 500–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarif, H.; Nicolas, S.; Guyot, M.; Hosseiny, S.; Lazzari, A.; Canali, M.M.; Cazareth, J.; Brau, F.; Golzné, V.; Dourneau, E.; et al. CD8+ T cells are essential for the effects of enriched environment on hippocampus-dependent behavior, hippocampal neurogenesis and synaptic plasticity. Brain Behav. Immun. 2018, 69, 235–254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xie, H.; Wang, Y.; Li, D.; Du, L.; Wu, Y.; Yang, G.Y. Enriched environment improves behavioral performance and attenuates inflammatory response induced by TNF-α in healthy adult mice. Eur. J. Inflamm. 2017, 15, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Otaki, M.; Hirano, T.; Yamaguchi, Y.; Kaida, K.; Koshika, S.; Nagata, K.; Nishimura, M.; Kakinuma, S.; Shimada, Y.; Kobayashi, Y. Changes in the function and phenotype of resident peritoneal macrophages after housing in an enriched environment. Int. Immunopharmacol. 2018, 65, 44–49. [Google Scholar] [CrossRef]
- Xiao, R.; Ali, S.; Caligiuri, M.A.; Cao, L. Enhancing Effects of Environmental Enrichment on the Functions of Natural Killer Cells in Mice. Front Immunol. 2021, 28, 695859. [Google Scholar] [CrossRef]
- Stephenson, J.; Nutma, E.; van der Valk, P.; Amor, S. Inflammation in CNS neurodegenerative diseases. Immunology 2018, 154, 204–219. [Google Scholar] [CrossRef] [Green Version]
- Hammond, T.R.; Marsh, S.E.; Stevens, B. Immune Signaling in Neurodegeneration. Immunity 2019, 50, 955–974. [Google Scholar] [CrossRef] [Green Version]
- Sabatino, J.J., Jr.; Pröbstel, A.K.; Zamvil, S.S. B cells in autoimmune and neurodegenerative central nervous system diseases. Nat. Rev. Neurosci. 2019, 20, 728–745. [Google Scholar]
- Huang, Z.B.; Sheng, G.Q. Interleukin-1β with learning and memory. Neurosci. Bull. 2010, 26, 455–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohman, R.A.; Rhodes, J.S. Neurogenesis, inflammation and behavior. Brain Behav. Immun. 2013, 27, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozben, T.; Ozben, S. Neuro-inflammation and anti-inflammatory treatment options for Alzheimer’s disease. Clin. Biochem. 2019, 72, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Sung, P.S.; Lin, P.Y.; Liu, C.H.; Su, H.C.; Tsai, K.J. Neuroinflammation and Neurogenesis in Alzheimer’s Disease and Potential Therapeutic Approaches. Int. J. Mol. Sci. 2020, 21, 701. [Google Scholar] [CrossRef] [Green Version]
- Okabe, Y. Immune Niche Within the Peritoneal Cavity. Curr. Top Microbiol. Immunol. 2021, 434, 123–134. [Google Scholar]
- Delibegovic, S. Pathophysiological changes in peritonitis. Med. Arh. 2007, 61, 109–113. [Google Scholar]
- Aufricht, C.; Neuhofer, W.; Topley, N.; Wörnle, M. Peritoneal infection and inflammation. Mediat. Inflamm. 2012, 2012, 456985. [Google Scholar] [CrossRef]
- Queen, N.; Deng, H.; Huang, W.; Mo, X.; Wilkins, R.K.; Zhu, T.; Wu, X.; Cao, L. Environmental Enrichment Mitigates Age-Related Metabolic Decline and Lewis Lung Carcinoma Growth in Aged Female Mice. Cancer Prev. Res. 2021, 14, 1075–1088. [Google Scholar] [CrossRef]
- Perry, V.H.; Nicoll, J.A.; Holmes, C. Microglia in neurodegenerative disease. Nat. Rev. Neurol. 2010, 6, 193–201. [Google Scholar] [CrossRef]
- Li, Q.; Barres, B.A. Microglia and macrophages in brain homeostasis and disease. Nat. Rev. Immunol. 2018, 18, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Bartels, T.; De Schepper, S.; Hong, S. Microglia modulate neurodegeneration in Alzheimer’s and Parkinson’s diseases. Science 2020, 370, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Leng, F.; Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: Where do we go from here ? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Moir, S.; Chun, T.W.; Fauci, A.S. Pathogenic mechanisms of HIV disease. Annu. Rev. Pathol. 2011, 6, 223–248. [Google Scholar] [CrossRef]
- Sattentau, Q.J.; Moore, J.P. The role of CD4 in HIV binding and entry. Philos. Trans. R Soc. Lond. B Biol. Sci. 1993, 342, 59–66. [Google Scholar] [PubMed]
- Sadighi Akha, A.A. Aging and the immune system: An overview. J. Immunol. Methods 2018, 463, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Di Benedetto, S.; Pawelec, G. The immune system and its dysregulation with Aging. Subcell Biochem. 2019, 91, 21–43. [Google Scholar] [PubMed]
- Schink, A.; Neumann, J.; Leifke, A.L.; Ziegler, K.; Fröhlich-Nowoisky, J.; Cremer, C.; Thines, E.; Weber, B.; Pöschl, U.; Schuppan, D.; et al. Screening of herbal extracts for TLR2- and TLR4-dependent anti-inflammatory effects. PLoS ONE 2018, 13, e0203907. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.C.; Khan, S.Q.; Kaneda, M.M.; Pathria, P.; Shepard, R.; Louis, T.L.; Anand, S.; Woo, G.; Leem, C.; Faridi, M.H.; et al. Integrin CD11b activation drives anti-tumor innate immunity. Nat. Commun. 2018, 9, 5379. [Google Scholar] [CrossRef] [Green Version]
- Starzer, A.M.; Berghoff, A.S. New emerging targets in cancer immunotherapy: CD27 (TNFRSF7). ESMO Open 2020, 4, e000629. [Google Scholar] [CrossRef] [Green Version]
- Myers, J.A.; Miller, J.S. Exploring the NK cell platform for cancer immunotherapy. Nat. Rev. Clin. Oncol. 2021, 18, 85–100. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Database | Code Line |
|---|---|
| Pubmed/Medline Scopus | (((Environmental Enrichment) OR (Enriched Environment)) AND ((((((((Immune System) OR (Immune Systems)) OR (System, Immune)) OR (Inflammation)) OR (Inflammations)) OR (Innate Inflammatory Response)) OR (Cytokine)) OR (Cytokines)). |
| ScienceDirect | (((“Exercise”) OR (“Physical Activity”)) OR (“Physical Exercise”)) AND ((((“Endoplasmic Reticulum Stress”) OR (“Endoplasmic Reticulum Stresses”)) OR (“Stress, Endoplasmic Reticulum”)) |
| Inclusion Criteria | Exclusion Criteria | |
|---|---|---|
| Population | Rodents | Humans and other species |
| Intervention | Environmental Enrichment | Absence of the Environmental Enrichment |
| Comparator | Animals that did not undergo Environmental Enrichment | Absence of the control group |
| Outcomes | Cytokines and immune cells | No Cytokines and immune cells |
| Study design | Experimental | Observational, reviews, case reports, and scientific abstracts |
| Study | Q1 | Q2 | Q3 | Q4 | Q5 | Q6 | Q7 | Q8 | Q9 | Q10 | Score |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ali et al., 2019 [13] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Arranz et al., 2010 [14] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Brod et al., 2017 [15] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Ferré et al., 2016 [16] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Gurfein et al., 2017 [17] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Keymoradzadeh et al., 2020 [18] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Marashi et al., 2003 [19] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| McQuaid et al., 2013 [20] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Meng et al., 2018 [21] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Rattazi et al., 2016 [22] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Takai et al., 2019 [10] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Wang et al., 2018 [23] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Williamson et al., 2012 [24] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Zarif et al., 2017 [25] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Zhang et al., 2017 [26] | U | Y | Y | Y | N | Y | U | U | Y | Y | 6 |
| Author, Year | Species/Sex and Age | Environmental Enrichment Protocol, Animals Data, and Housing Dimensions (Length, Width, and Depth) | Exposure Time to Environmental Enrichment | Brain Outcomes |
|---|---|---|---|---|
| Ali, 2019 [13] | C57BL/6N mice, Female, and 10 months old | Wheels, tunnels, igloos, huts, retreats, wood toys, and a maze in addition to standard chow and water. n = 5 and 63 cm × 49 cm × 44 cm | 10 months | Hypothalamus: ↓ CCL2, IL-1β, IL-6. Amygdala: ↓ CCL2 e NFkbia/= IL-1β, IL-6, SOCS3. |
| Arranz, 2010 [14] | ICR/CD-1 mice, Female, Adults: 44 ± 4 weeks, Old: 69 ± 4 weeks, Very Old: 92 ± 4 weeks | Range bucket, jolly ball, hoop, holed ball, yellow tunnel, rough red object, yellow billiard ball, and silver ball. Uninformed and Uniformed. | 6–8 weeks | - |
| Brod, 2017 [15] | CD1 mice, male and 6 weeks old | Wheel, nest house, tunnel, ample nesting material, and wood chip bedding to a 5 cm depth. Uninformed and 50 × 38 cm × 21 cm. | 2 weeks | - |
| Ferré, 2016 [16] | SAMP8 mice, Male and 3 months old | Plastic tubes (20 cm long and 2.5 cm in diameter) were placed in EE cardboard cages, moreover to plastic dolls or toys, which were added, removed, or changed every week. Uninformed and Uninformed. | Uninformed | Hippocampus: ↓ IL-6; CXCL10/= TNF-α |
| Gurfein, 2017 [17] | BALB/c mice, Male and 6 weeks old | Uninformed, n = 10 and 257 mm × 483 mm × 152 mm = 980 cm2 | 9 weeks | - |
| Keymoradzadeh, 2020 [18] | Wistar Rats, Male and 7 weeks old | Running wheels, a tunnel, a small compartment, stairs, and many other colorful objects (e.g., colorful plastic plates, wooden disks of varied colors and sizes, plastic cups, and hanging cubes). n = 7 and 96 cm × 49 cm × 38 cm. | 3 weeks | Hippocampus: ↓ IL-6; CXCL10/= TNF-α. |
| Marashi, 2003 [19] | Strain CS of the inbred strain ABG mice, Male and uniformed | Passable enriched cage, extra plains, plastic stairs, wooden footpaths, hemp ropes, and a climbing tree. n = 4 and 100 cm × 40 cm × 34.5 cm | Uninformed | Hippocampus: = IL-1β and IL-10. |
| Mcquaid, 2013 [20] | BALB/cByJ mice, male and 6–8 weeks old | Two running wheels, one red polypropylene shelter, one orange polypropylene shelter with an angled running wheel, as well as three yellow polypropylene tunnels and two cotton nestlets. Uninformed and 50 cm × 40 cm × 20 cm. | 4 weeks | Hippocampus: = IL-6, IL-1b, TNF-α. ↓ IL-6r; IL-1r/= TNF-αr. Prefrontal Córtex: = IL-6r; IL-1r and TNF-αr. |
| Meng, 2018 [21] | C57BL/6N mice/Male/3 or 10 weeks old | Running wheels, wood toys, plastic tunnels, ladders, huts, and nesting materials. Toys were changed every 3–4 days. n = 12 and 61 cm × 43 cm × 21 cm. | 5 weeks | - |
| Otaki, 2018 [27] | B6C3F1/Cr1 mice/Male and 3 weeks | Running wheels, tunnel, bio-hut, wood gnawing block, shelter, and nesting sheet. n = 5 and Uniformed. | 6–8 weeks | - |
| Rattazi, 2016 [22] | C57BL/6N mice/Male and 6 weeks | Colored transparent plastic nest-box, fabric tube, running wheel, and one wood hamster swing. n = 6 and 40 cm × 25 cm × 20 cm. | 2 weeks | - |
| Takai, 2019 [10] | B6C3F1/mice/Female and 6 weeks old | Uninformed, n = 8–24 and Uninformed. | 6 weeks | - |
| Wang, 2018 [23] | C57BL/6N mice/Male and 18 months old | Uninformed, n = 4 and 47 cm × 30 cm × 23 cm | 2 months | Hippocampus: ↓ IL-1b; TNF-α; =IL-6; =pro-IL1b; ↓ p-IKKβ/IKKβ and p-P65/P65. Prefrontal Cortex: ↓ IL-1b; TNF-α; = IL-6; = pro-IL1β/= p-IKKβ/IKKβ and p-P65/P65. |
| Williamson, 2012 [24] | Sprague -Dawley | Quarter-inch corn-cob bedding (identical to home cage controls), a running wheel, a PVC tube, and various small objects and toys. n = 6–8; 55.9 cm × 35.6 cm × 30.5 cm | 7 weeks | Hippocampus: ↓ IL-1β; TNF-α; TNF-rsfla; TNF-r1β; TNF-β; ↓ MCP-1/CCL2; MIP-1α/CCL3; MIP-2α/CXCL2. |
| Zarif, 2017 [25] | C57BL/6N mice/Female and 4 weeks | Nesting material, houses, running wheels, hammocks, scales, plastic toys, and tunnels. n = 5–6, 120 cm × 76 cm × 21 cm | 1–4 weeks | Hippocampus: ↓ TNF-α; = IFN-γ. |
| Zhang, 2017 [26] | C57BL/6 N mice and 10 weeks old | Plastic tunnels, wooden climbing frame, platforms, hiding shelters, house, exercise wheel, chew toys, and other novel objects designed specifically for small animals = 20 and 56 cm × 40 cm × 22 cm. | 3 weeks | Hippocampus: = IL-1β; TNF-α; ↓ CD68+ |
| Author, Year | Species/Sex and Age | Environmental Enrichment Protocol, Animals Data, and Housing Dimensions (Length, Width, and Depth) | Exposure Time to Environmental Enrichment | Peripheral Outcomes |
|---|---|---|---|---|
| Ali, 2019 [13] | C57BL/6N mice, Female, and 10 months old | Wheels, tunnels, igloos, huts, retreats, wood toys, and a maze in addition to standard chow and water. n = 5 and 63 cm × 49 cm × 44 cm | 10 months | Peritoneal Cavity: ↓ % CD4 cells in all groups. ↑ % CD11b+ in very old mice (VO). ↑ macrophage chemotaxis in VO. ↑ phagocytic index in old and VO; ↑ phagocytic efficacy in VO. ↑ NK cells proliferation in adult and VO. ↑ NK cells activity in VO. ↑ Lymphocyte’s chemotaxis in old and VO. ↑ % TLR2 and TLR4 on CD11b+ cells in VO. ↓ % TLR2 and TLR4 on CD11c+ and CD4 cells in VO. ↓ % TLR2 on CD8 cells in VO. ↑% TLR2 on CD19+ in VO. |
| Arranz, 2010 [14] | ICR/CD-1 mice, Female, Adults: 44 ± 4 weeks, Old: 69 ± 4 weeks, Very Old: 92 ± 4 weeks | Range bucket, jolly ball, hoop, holed ball, yellow tunnel, rough red object, yellow billiard ball, and silver ball. Uninformed and Uniformed. | 6–8 weeks | Blood Tissue: ↑ % leukocytes and lymphocytes. Peritoneal Cavity: ↑ IL-2/↑ TNF-α, = IL-6, MCP-1 and TNF-α. |
| Brod, 2017 [15] | CD1 mice, Male and 6 weeks old | Wheel, nest house, tunnel, ample nesting material, and wood chip bedding to a 5 cm depth. Uninformed and 50 cm × 38 cm × 21 cm. | 2 weeks | - |
| Ferré, 2016 [16] | SAMP8 mice, Male and 3 months old | Plastic tubes (20 cm long and 2.5 cm in diameter) were placed in EE cardboard cages, and plastic dolls or toys, which were added, removed, or changed every week. Uninformed and Uninformed. | Uninformed | Blood Tissue: ↑CD19b+ lymphocytes; = Thy1.2+T lymphocytes; = CD4+T lymphocytes; = CD8+ lymphocytes; = CD11b+ monocytes; CD49b+ NK cells; Ly-6g+ neutrophils; ↓ B/T lymphocytes ratio. |
| Gurfein, 2017 [17] | BALB/c mice, Male and 6 weeks old | Uninformed, n = 10 and 257 mm × 483 mm × 152 mm = 980 cm2 | 9 weeks | - |
| Keymoradzadeh, 2020 [18] | Wistar Rats, Male and 7 weeks old | Running wheels, a tunnel, a small compartment, stairs, and many other colorful objects (e.g., colorful plastic plates, wooden disks of varied colors and sizes, plastic cups, and hanging cubes). n = 7 and 96 cm × 49 cm × 38 cm. | 3 weeks | Blood Tissue: = CD4/CD8 ratio and CD4; ↓ CD8 cells. Spleen: = IL-2; IL-4; IL-10 and IFN-γ/↓ IFN-γ/IL-10; ↓IL-2/IL-10. |
| Marashi, 2003 [19] | Strain CS of the inbred strain ABG mice, Male and uniformed | Passable enriched cage, extra plains, plastic stairs, wooden footpaths, hemp ropes, and a climbing tree. n = 4 and 100 cm × 40 cm × 34.5 cm | Uninformed | - |
| Mcquaid, 2013 [20] | BALB/cByJ mice, male and 6–8 weeks old | Two running wheels, one red polypropylene shelter, one orange polypropylene shelter with an angled running wheel, as well as three yellow polypropylene tunnels and two cotton nestles. Uninformed and 50 cm × 40 cm × 20 cm. | 4 weeks | Bone marrow: = NK cells; ↑ mature CD27 and CD11b+ cells; ↓ immature CD27 and CD11b+ cells. ↑ NK cells proliferation. Spleen: ↑ % NK cells; ↑ mature CD27 and CD11b+ cells; ↓immature CD27 and CD11b+ cells. Blood Tissue: = NK cells; ↑ CD27 and CD11b+ cells; ↓ immature CD27 and CD11B+ cells. ↑ NK cells proliferation. |
| Meng, 2018 [21] | C57BL/6N mice/Male/3 or 10 weeks old | Running wheels, wood toys, plastic tunnels, ladders, huts, and nesting materials. Toys were changed every 3–4 days. n = 12 and 61 cm × 43 cm × 21 cm. | 5 weeks | Peritoneal Cavity: ↑ macrophage phagocytosis. = MIP-2; ↑ MCP-1; ↑ Phagocytosis of apoptotic neutrophils. M1 markers: ↑ CD40 cells; ↑ IL-6; =NOS2 and ↓ CD38. |
| Otaki, 2018 [27] | B6C3F1/Cr1 mice/Male and 3 weeks | Running wheels, tunnel, bio-hut, wood gnawing block, shelter, and nesting sheet. n = 5 and Uniformed. | 6–8 weeks | Thymus, lymph nodes, or spleen: = CD3 and CD4 T cells; ↓ differentiation of CD4 T cells in response to IFN-γ and ↑ IL-17. Lymphocytes: = IL-4; IL-10; ↑ IL-17; ↓ IFN-γ |
| Rattazi, 2016 [22] | C57BL/6N mice/Male and 6 weeks | Colored transparent plastic nest-box, fabric tube, running wheel, and one wood hamster swing. n = 6 and 40 cm × 25 cm × 20 cm. | 2 weeks | Lymphocytes: ↑ NK cell activity; = NK cells. |
| Takai, 2019 [10] | B6C3F1/mice/Female and 6 weeks old | Uninformed, n = 8–24 and Uninformed. | 6 weeks | - |
| Wang, 2018 [21] | C57BL/6N mice/Male and 18 months old | Uninformed, n = 4 and 47 cm × 30 cm × 23 cm | 2 months | - |
| Williamson, 2012 [24] | Sprague -Dawley | Quarter-inch corn-cob bedding (identical to home cage controls), a running wheel, a PVC tube, and various small objects and toys. n = 6–8; 55.9 cm × 35.6 cm × 30.5 cm | 7 weeks | Spleen: = CD8+ T cells; = CD8/CD4; ↑ CD8 cell proliferation |
| Zarif, 2017 [25] | C57BL/6N mice/Female and 4 weeks | Nesting material, houses, running wheels, hammocks, scales, plastic toys, and tunnels. n = 5–6, 120 cm × 76 cm × 21 cm | 1–4 weeks | Peritoneal Cavity: ↓ % CD4 cells in all groups. ↑ % CD11b+ in very old mice (VO). ↑ macrophage chemotaxis in VO. ↑ phagocytic index in old and VO; ↑ phagocytic efficacy in VO. ↑ NK cells proliferation in adult and VO. ↑ NK cells activity in VO. ↑ Lymphocyte’s chemotaxis in old and VO. ↑ % TLR2 and TLR4 on CD11b+ cells in VO. ↓ % TLR2 and TLR4 on CD11c+ and CD4 cells in VO. ↓ % TLR2 on CD8 cells in VO. ↑% TLR2 on CD19+ in VO. |
| Zhang, 2017 [26] | C57BL/6N mice/Male and 10 weeks old | Plastic tunnels, wooden climbing frame, platforms, hiding shelters, house, exercise wheel, chew toys, and other novel objects designed specifically for small animals = 20 and 56 cm × 40 cm × 22 cm. | 3 weeks | Blood Tissue: ↑ % leukocytes and lymphocytes. Peritoneal Cavity: ↑ IL-2/↑ TNF-α, = IL-6, MCP-1 and TNF-α. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sousa Fernandes, M.S.; Santos, G.C.J.; Filgueira, T.O.; Gomes, D.A.; Barbosa, E.A.S.; dos Santos, T.M.; Câmara, N.O.S.; Castoldi, A.; Souto, F.O. Cytokines and Immune Cells Profile in Different Tissues of Rodents Induced by Environmental Enrichment: Systematic Review. Int. J. Mol. Sci. 2022, 23, 11986. https://doi.org/10.3390/ijms231911986
de Sousa Fernandes MS, Santos GCJ, Filgueira TO, Gomes DA, Barbosa EAS, dos Santos TM, Câmara NOS, Castoldi A, Souto FO. Cytokines and Immune Cells Profile in Different Tissues of Rodents Induced by Environmental Enrichment: Systematic Review. International Journal of Molecular Sciences. 2022; 23(19):11986. https://doi.org/10.3390/ijms231911986
Chicago/Turabian Stylede Sousa Fernandes, Matheus Santos, Gabriela Carvalho Jurema Santos, Tayrine Ordonio Filgueira, Dayane Aparecida Gomes, Elias Almeida Silva Barbosa, Tony Meireles dos Santos, Niels Olsen Saraiva Câmara, Angela Castoldi, and Fabricio Oliveira Souto. 2022. "Cytokines and Immune Cells Profile in Different Tissues of Rodents Induced by Environmental Enrichment: Systematic Review" International Journal of Molecular Sciences 23, no. 19: 11986. https://doi.org/10.3390/ijms231911986