
Therapeutic Potential of Mesenchymal Stem Cell-Secreted Factors on Delay in Corneal Wound Healing by Nitrogen Mustard
 , and
, and
Abstract
:1. Introduction
2. Results
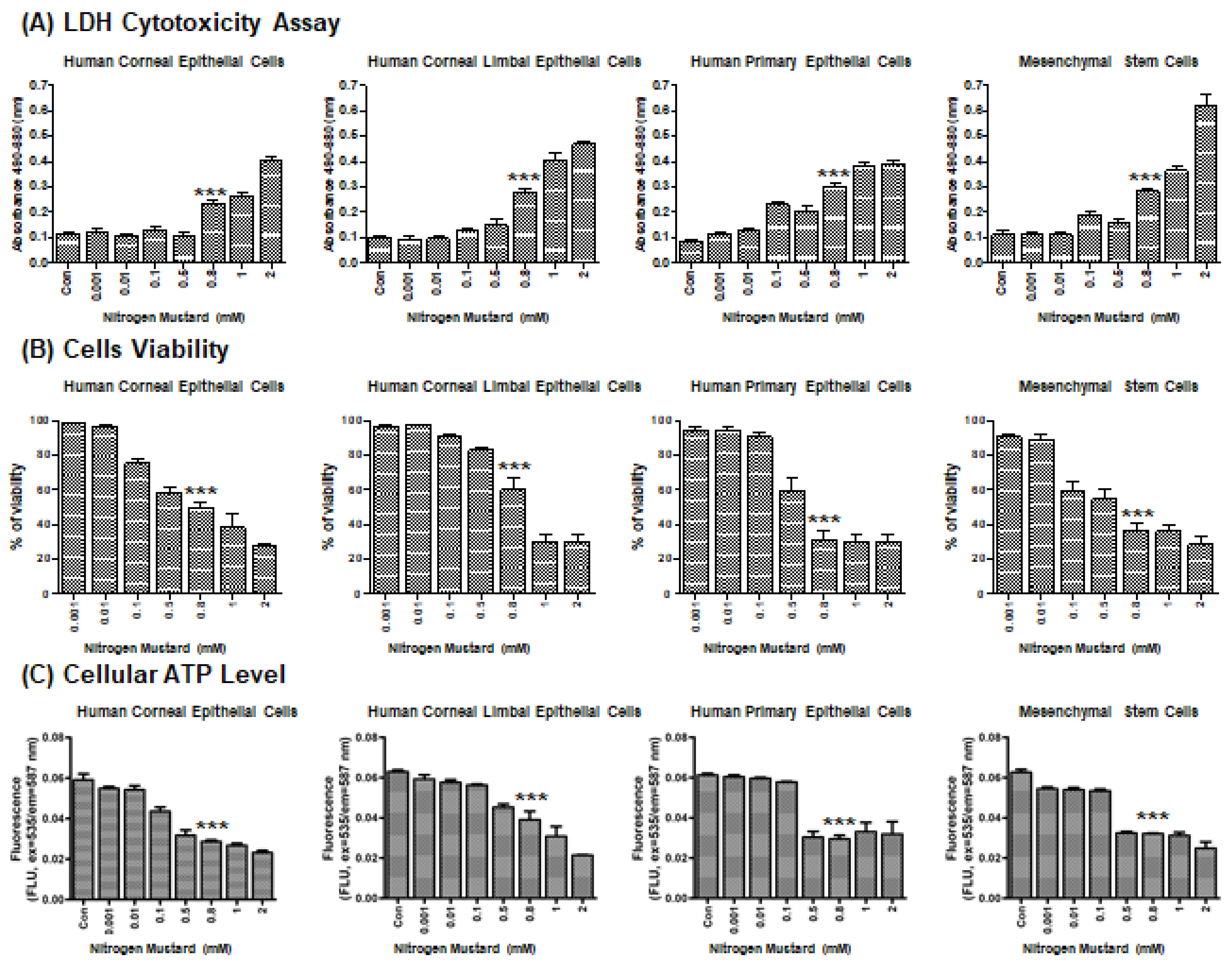
2.1. Effects of Nitrogen Mustard on Viability In Vitro
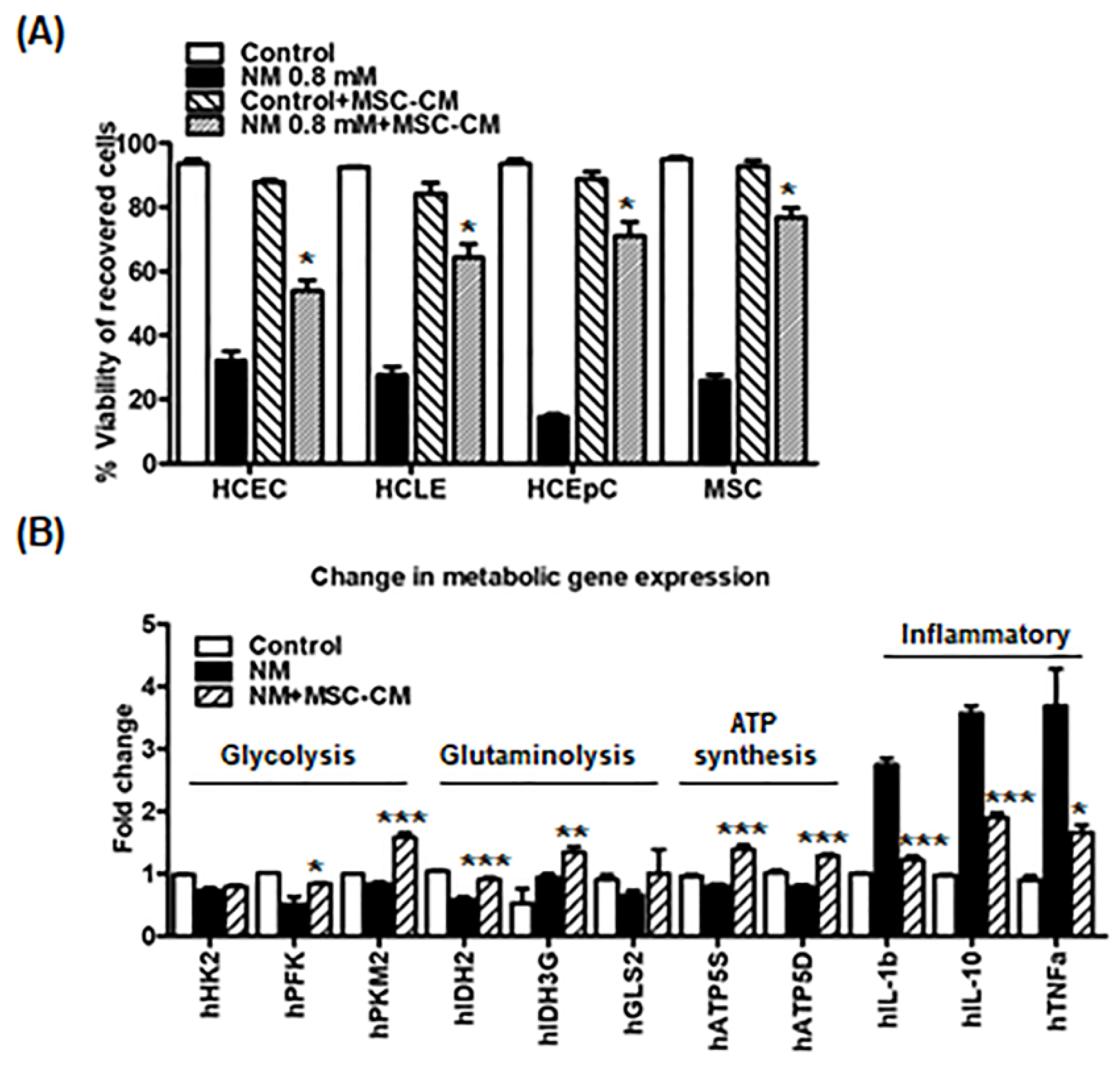
2.2. MSC-CM Rescues Immediate Cytotoxic Effects and Enhances Wound Healing in Corneal Epithelial Cells Exposed to NM
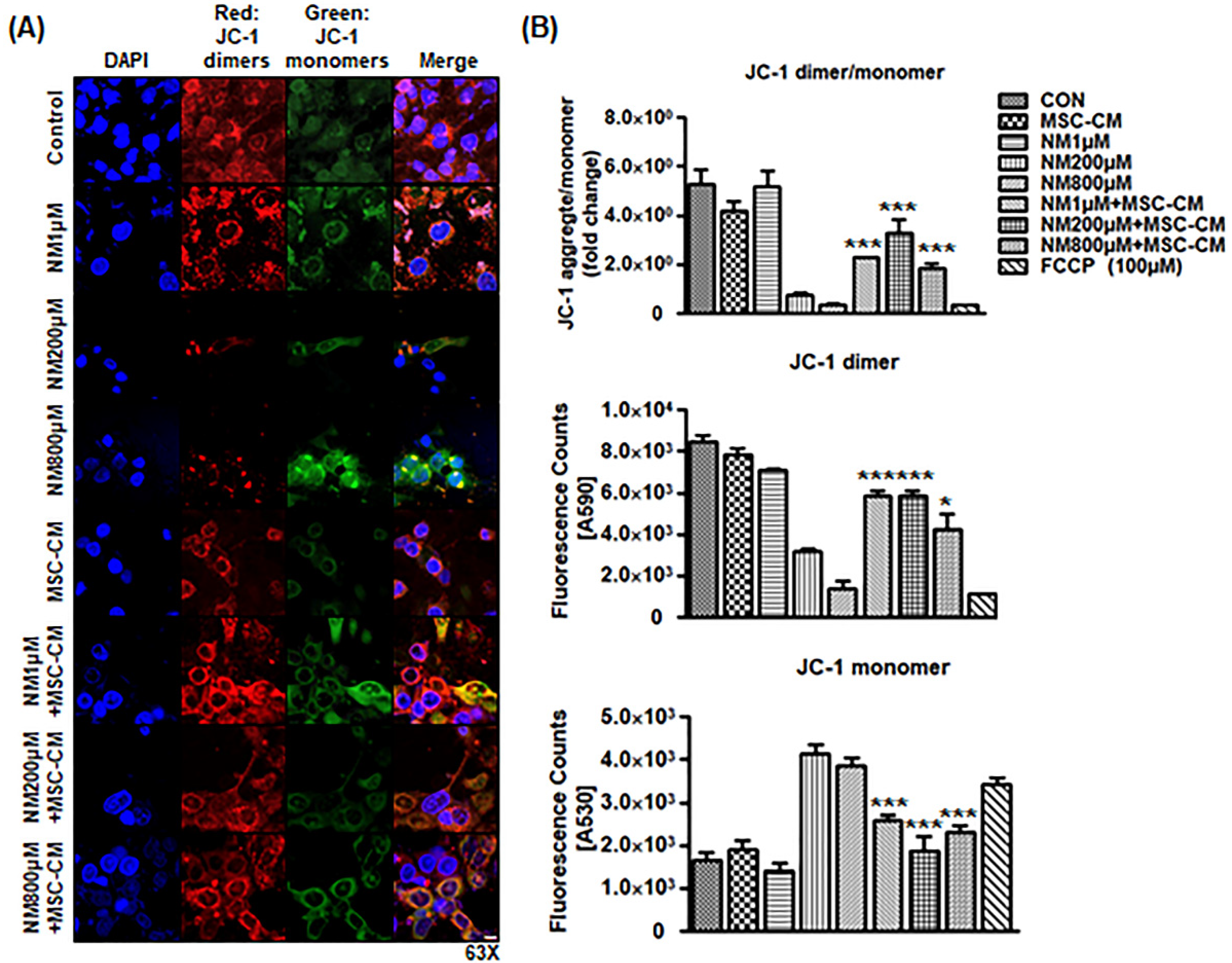
2.3. MSC-CM Attenuates NM-Induced Change in Mitochondrial Membrane Potential
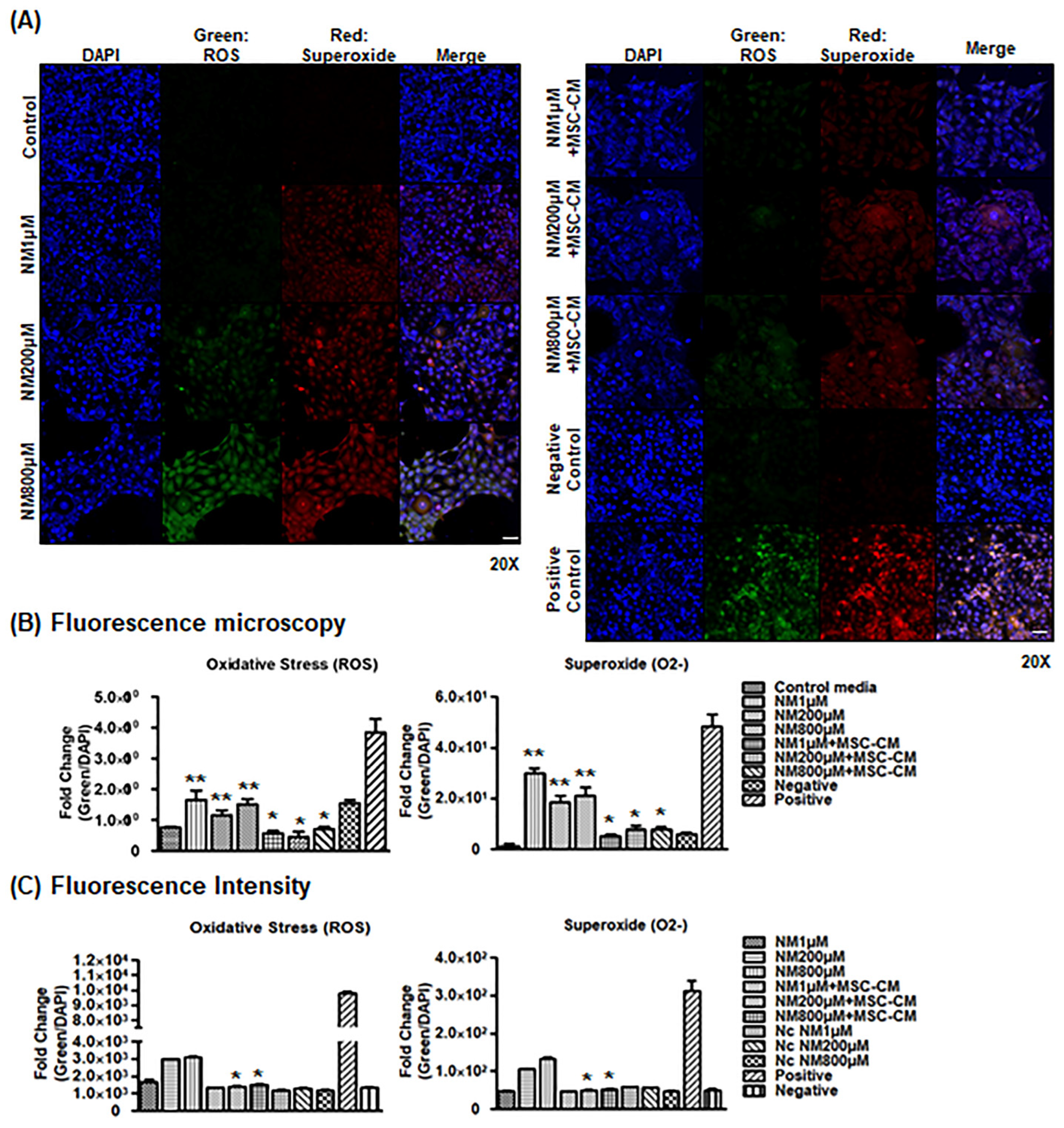
2.4. MSC-CM Decreases Cellular ROS Generation
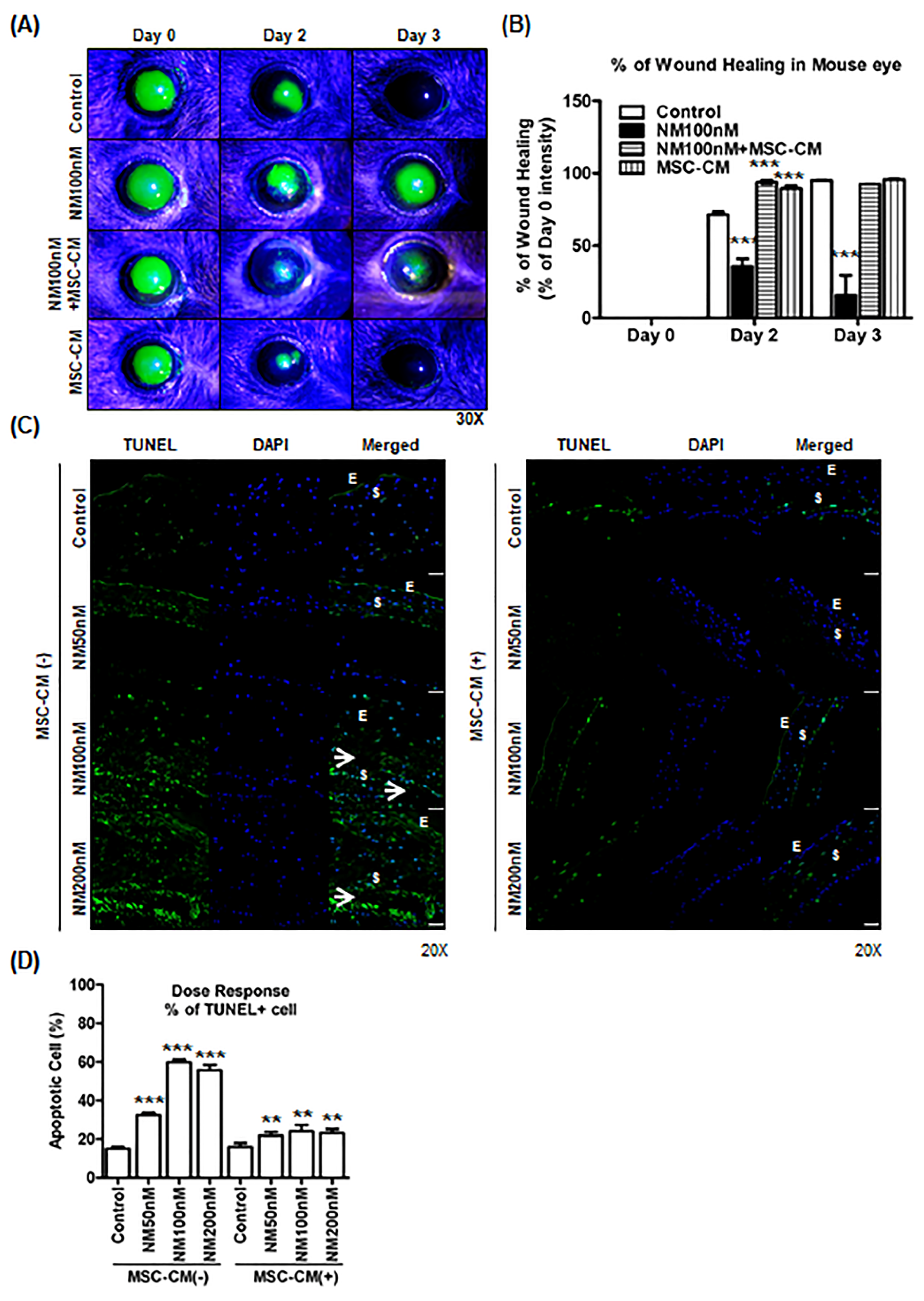
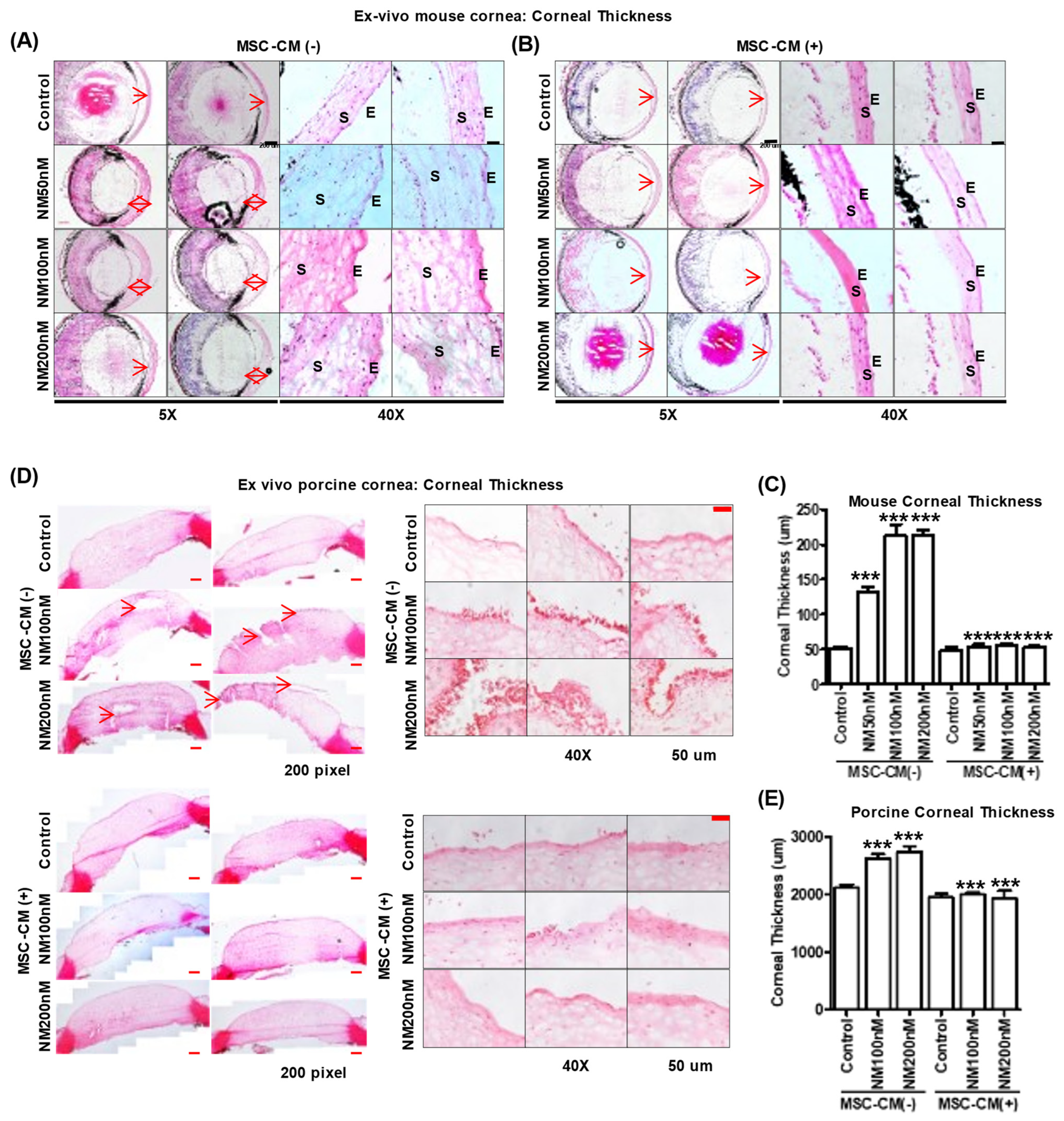
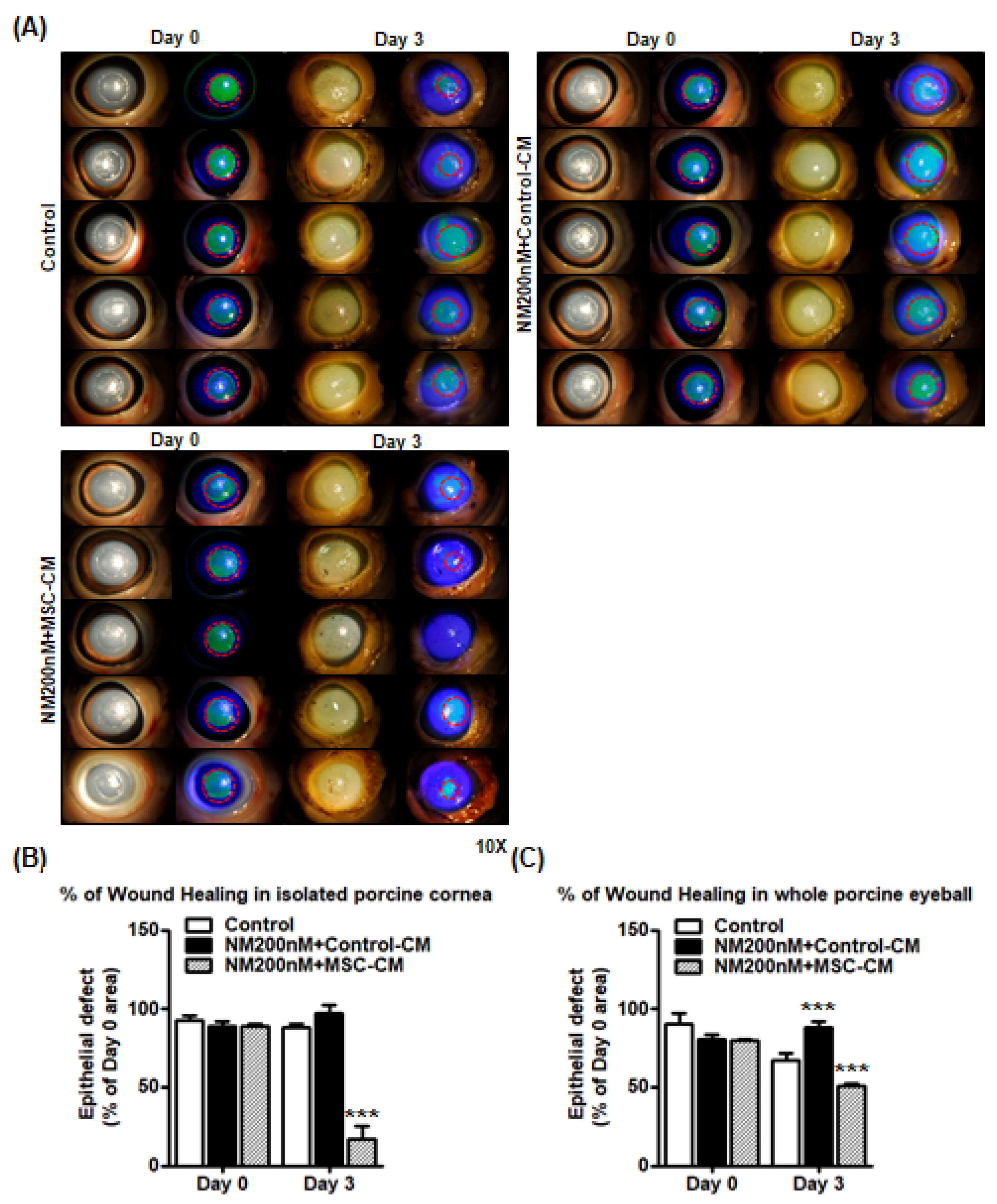
2.5. Effect of MSC-CM on Murine and Porcine Corneal Wound Healing after NM Injury
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of Nitrogen Mustards and Treatment
4.3. Cell Viability
4.4. MSC-CM
4.5. LDH Toxicity Assay
4.6. ROS Measurements
4.7. JC-1 Immunostaining
4.8. ATP Measurements
4.9. Apoptosis Detection Kit
4.10. Porcine Cornea Model of Corneal Epithelial Wound Healing
4.11. Mouse Model of Corneal Epithelial Wound Healing
4.12. Histology
4.13. In Vitro Scratch Assay
4.14. Macrophage Polarization and Flow Cytometry
4.15. Real-Time PCR
4.16. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gordon, M.K.; Desantis, A.; Deshmukh, M.; Lacey, C.J.; Hahn, R.A.; Beloni, J.; Anumolu, S.S.; Schlager, J.J.; Gallo, M.A.; Gerecke, D.R.; et al. Doxycycline hydrogels as a potential therapy for ocular vesicant injury. J. Ocul. Pharmacol. Ther. 2010, 26, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.G.; Kumar, D.; Tewari-Singh, N.; Orlicky, D.J.; Jain, A.K.; Kant, R.; Rancourt, R.C.; Dhar, D.; Inturi, S.; Agarwal, C.; et al. Topical nitrogen mustard exposure causes systemic toxic effects in mice. Exp. Toxicol. Pathol. 2015, 67, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.G.; Tewari-Singh, N.; Dhar, D.; Kumar, D.; Agarwal, C.; Ammar, D.A.; Kant, R.; Enzenauer, R.W.; Petrash, J.M.; Agarwal, R. Nitrogen Mustard-Induced Corneal Injury Involves DNA Damage and Pathways Related to Inflammation, Epithelial-Stromal Separation, and Neovascularization. Cornea 2016, 35, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Loeber, R.; Michaelson, E.; Fang, Q.; Campbell, C.; Pegg, A.E.; Tretyakova, N. Cross-linking of the DNA repair protein Omicron6-alkylguanine DNA alkyltransferase to DNA in the presence of antitumor nitrogen mustards. Chem. Res. Toxicol. 2008, 21, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Jan, Y.H.; Heck, D.E.; Laskin, D.L.; Laskin, J.D. DNA damage signaling in the cellular responses to mustard vesicants. Toxicol. Lett. 2020, 326, 78–82. [Google Scholar] [CrossRef]
- Mol, M.A.; van der Schans, G.P.; Lohman, P.H. Quantification of sulfur mustard-induced DNA interstrand cross-links and single-strand breaks in cultured human epidermal keratinocytes. Mutat. Res. 1993, 294, 235–245. [Google Scholar] [CrossRef]
- Her, J.; Bunting, S.F. How cells ensure correct repair of DNA double-strand breaks. J. Biol. Chem. 2018, 293, 10502–10511. [Google Scholar] [CrossRef]
- Kadar, T.; Dachir, S.; Cohen, L.; Sahar, R.; Fishbine, E.; Cohen, M.; Turetz, J.; Gutman, H.; Buch, H.; Brandeis, R.; et al. Ocular injuries following sulfur mustard exposure--pathological mechanism and potential therapy. Toxicology 2009, 263, 59–69. [Google Scholar] [CrossRef]
- Charkoftaki, G.; Jester, J.V.; Thompson, D.C.; Vasiliou, V. Nitrogen mustard-induced corneal injury involves the sphingomyelin-ceramide pathway. Ocul. Surf. 2018, 16, 154–162. [Google Scholar] [CrossRef]
- Naderi, M.; Jadidi, K.; Falahati, F.; Alavi, S.A. The effect of sulfur mustard and nitrogen mustard on corneal collagen degradation induced by the enzyme collagenase. Cutan. Ocul. Toxicol. 2010, 29, 234–240. [Google Scholar] [CrossRef]
- Tewari-Singh, N.; Rana, S.; Gu, M.; Pal, A.; Orlicky, D.J.; White, C.W.; Agarwal, R. Inflammatory biomarkers of sulfur mustard analog 2-chloroethyl ethyl sulfide-induced skin injury in SKH-1 hairless mice. Toxicol. Sci. 2009, 108, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.K.; Tewari-Singh, N.; Orlicky, D.J.; White, C.W.; Agarwal, R. 2-Chloroethyl ethyl sulfide causes microvesication and inflammation-related histopathological changes in male hairless mouse skin. Toxicology 2011, 282, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Tewari-Singh, N.; Jain, A.K.; Inturi, S.; Ammar, D.A.; Agarwal, C.; Tyagi, P.; Kompella, U.B.; Enzenauer, R.W.; Petrash, J.M.; Agarwal, R. Silibinin, dexamethasone, and doxycycline as potential therapeutic agents for treating vesicant-inflicted ocular injuries. Toxicol. Appl. Pharmacol. 2012, 264, 23–31. [Google Scholar] [CrossRef]
- Kaluzhny, Y.; Kinuthia, M.W.; Lapointe, A.M.; Truong, T.; Klausner, M.; Hayden, P. Oxidative stress in corneal injuries of different origin: Utilization of 3D human corneal epithelial tissue model. Exp. Eye Res. 2020, 190, 107867. [Google Scholar] [CrossRef] [PubMed]
- Laskin, J.D.; Black, A.T.; Jan, Y.H.; Sinko, P.J.; Heindel, N.D.; Sunil, V.; Heck, D.E.; Laskin, D.L. Oxidants and antioxidants in sulfur mustard-induced injury. Ann. N. Y. Acad. Sci. 2010, 1203, 92–100. [Google Scholar] [CrossRef]
- Eveleth, D.D.; Eveleth, J.J.; Subramaniam, A.; Hahn, R.; Zhou, P.; Gordon, M.K.; Bradshaw, R.A. An Engineered Human Fibroblast Growth Factor-1 Derivative, TTHX1114, Ameliorates Short-term Corneal Nitrogen Mustard Injury in Rabbit Organ Cultures. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4720–4730. [Google Scholar] [CrossRef]
- Javadi, M.A.; Baradaran-Rafii, A. Living-related conjunctival-limbal allograft for chronic or delayed-onset mustard gas keratopathy. Cornea 2009, 28, 51–57. [Google Scholar] [CrossRef]
- Javadi, M.A.; Yazdani, S.; Sajjadi, H.; Jadidi, K.; Karimian, F.; Einollahi, B.; Ja’farinasab, M.R.; Zare, M. Chronic and delayed-onset mustard gas keratitis: Report of 48 patients and review of literature. Ophthalmology 2005, 112, 617–625. [Google Scholar] [CrossRef]
- Javadi, M.A.; Yazdani, S.; Kanavi, M.R.; Mohammadpour, M.; Baradaran-Rafiee, A.; Jafarinasab, M.R.; Einollahi, B.; Karimian, F.; Zare, M.; Naderi, M.; et al. Long-term outcomes of penetrating keratoplasty in chronic and delayed mustard gas keratitis. Cornea 2007, 26, 1074–1078. [Google Scholar] [CrossRef]
- Safarinejad, M.R.; Moosavi, S.A.; Montazeri, B. Ocular injuries caused by mustard gas: Diagnosis, treatment, and medical defense. Mil. Med. 2001, 166, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Dahl, H.; Gluud, B.; Vangsted, P.; Norn, M. Eye lesions induced by mustard gas. Acta Ophthalmol. Suppl. 1985, 173, 30–31. [Google Scholar] [CrossRef] [PubMed]
- Solberg, Y.; Alcalay, M.; Belkin, M. Ocular injury by mustard gas. Surv. Ophthalmol. 1997, 41, 461–466. [Google Scholar] [CrossRef]
- Beigi Harchegani, A.; Khor, A.; Tahmasbpour, E.; Ghatrehsamani, M.; Bakhtiari Kaboutaraki, H.; Shahriary, A. Role of oxidative stress and antioxidant therapy in acute and chronic phases of sulfur mustard injuries: A review. Cutan Ocul. Toxicol. 2019, 38, 9–17. [Google Scholar] [CrossRef]
- Vallabh, N.A.; Romano, V.; Willoughby, C.E. Mitochondrial dysfunction and oxidative stress in corneal disease. Mitochondrion 2017, 36, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, M.; Kumar, N.V.A.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Fokou, P.V.T.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Vojvodic, V.; Milosavljevic, Z.; Boskovic, B.; Bojanic, N. The protective effect of different drugs in rats poisoned by sulfur and nitrogen mustards. Fundam Appl. Toxicol. 1985, 5 Pt 2, S160–S168. [Google Scholar] [CrossRef]
- Vogt, R.F., Jr.; Dannenberg, A.M., Jr.; Schofield, B.H.; Hynes, N.A.; Papirmeister, B. Pathogenesis of skin lesions caused by sulfur mustard. Toxicol. Sci. 1984, 4 Pt 2, S71–S83. [Google Scholar] [CrossRef]
- Oh, J.Y.; Kim, M.K.; Shin, M.S.; Lee, H.J.; Ko, J.H.; Wee, W.R.; Lee, J.H. The anti-inflammatory and anti-angiogenic role of mesenchymal stem cells in corneal wound healing following chemical injury. Stem Cells 2008, 26, 1047–1055. [Google Scholar] [CrossRef]
- Hoogduijn, M.J.; Popp, F.; Verbeek, R.; Masoodi, M.; Nicolaou, A.; Baan, C.; Dahlke, M.H. The immunomodulatory properties of mesenchymal stem cells and their use for immunotherapy. Int. Immunopharmacol. 2010, 10, 1496–1500. [Google Scholar] [CrossRef]
- Motegi, S.I.; Sekiguchi, A.; Uchiyama, A.; Uehara, A.; Fujiwara, C.; Yamazaki, S.; Perera, B.; Nakamura, H.; Ogino, S.; Yokoyama, Y.; et al. Protective effect of mesenchymal stem cells on the pressure ulcer formation by the regulation of oxidative and endoplasmic reticulum stress. Sci. Rep. 2017, 7, 17186. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.T.; Tsai, M.J.; Hsieh, N.; Lo, M.J.; Lee, M.J.; Cheng, H.; Huang, W.C. The superiority of conditioned medium derived from rapidly expanded mesenchymal stem cells for neural repair. Stem Cell Res. Ther. 2019, 10, 390. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.J.; Liou, D.Y.; Lin, Y.R.; Weng, C.F.; Huang, M.C.; Huang, W.C.; Tseng, F.W.; Cheng, H. Attenuating Spinal Cord Injury by Conditioned Medium from Bone Marrow Mesenchymal Stem Cells. J. Clin. Med. 2018, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, Y.; Bi, Y.; Gong, M.; Jiang, W.; Wei, X.; Li, T.; Chen, J. Mesenchymal stromal cell neuroprotection of hydrogen peroxide-challenged pheochromocytoma cells through reducing apoptosis and releasing cytokines. Cytotherapy 2012, 14, 954–966. [Google Scholar] [CrossRef] [PubMed]
- Voulgari-Kokota, A.; Fairless, R.; Karamita, M.; Kyrargyri, V.; Tseveleki, V.; Evangelidou, M.; Delorme, B.; Charbord, P.; Diem, R.; Probert, L. Mesenchymal stem cells protect CNS neurons against glutamate excitotoxicity by inhibiting glutamate receptor expression and function. Exp. Neurol. 2012, 236, 161–170. [Google Scholar] [CrossRef]
- Navas, A.; Magana-Guerrero, F.S.; Dominguez-Lopez, A.; Chavez-Garcia, C.; Partido, G.; Graue-Hernandez, E.O.; Sanchez-Garcia, F.J.; Garfias, Y. Anti-Inflammatory and Anti-Fibrotic Effects of Human Amniotic Membrane Mesenchymal Stem Cells and Their Potential in Corneal Repair. Stem Cells Transl. Med. 2018, 7, 906–917. [Google Scholar] [CrossRef]
- Jabbehdari, S.; Yazdanpanah, G.; Kanu, L.N.; Chen, E.; Kang, K.; Anwar, K.N.; Ghassemi, M.; Hematti, P.; Rosenblatt, M.I.; Djalilian, A.R. Therapeutic Effects of Lyophilized Conditioned-Medium Derived from Corneal Mesenchymal Stromal Cells on Corneal Epithelial Wound Healing. Curr. Eye Res. 2020, 45, 1490–1496. [Google Scholar] [CrossRef]
- Brayboy, L.M.; Clark, H.; Knapik, L.O.; Schnirman, R.E.; Wessel, G.M. Nitrogen mustard exposure perturbs oocyte mitochondrial physiology and alters reproductive outcomes. Reprod. Toxicol. 2018, 82, 80–87. [Google Scholar] [CrossRef]
- Kumar, D.; Tewari-Singh, N.; Agarwal, C.; Jain, A.K.; Inturi, S.; Kant, R.; White, C.W.; Agarwal, R. Nitrogen mustard exposure of murine skin induces DNA damage, oxidative stress and activation of MAPK/Akt-AP1 pathway leading to induction of inflammatory and proteolytic mediators. Toxicol. Lett. 2015, 235, 161–171. [Google Scholar] [CrossRef]
- Yu, W.; Dong, X.; Dan, G.; Ye, F.; Cheng, J.; Zhao, Y.; Chen, M.; Sai, Y.; Zou, Z. Vitamin D3 protects against nitrogen mustard-induced apoptosis of the bronchial epithelial cells via activating the VDR/Nrf2/Sirt3 pathway. Toxicol. Lett. 2022, 354, 14–23. [Google Scholar] [CrossRef]
- Banin, E.; Morad, Y.; Berenshtein, E.; Obolensky, A.; Yahalom, C.; Goldich, J.; Adibelli, F.M.; Zuniga, G.; DeAnda, M.; Pe’er, J.; et al. Injury induced by chemical warfare agents: Characterization and treatment of ocular tissues exposed to nitrogen mustard. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2966–2972. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Adachi, W.; Shimizu-Matsumoto, A.; Kinoshita, S.; Mizuno, K.; Matsubara, K.; Okubo, K. A gene expression profile of human corneal epithelium and the isolation of human keratin 12 cDNA. Investig. Ophthalmol. Vis. Sci. 1996, 37, 1800–1809. [Google Scholar]
- Kuipers, D.P.; Scripture, J.P.; Gunnink, S.M.; Salie, M.J.; Schotanus, M.P.; Ubels, J.L.; Louters, L.L. Differential regulation of GLUT1 activity in human corneal limbal epithelial cells and fibroblasts. Biochimie 2013, 95, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, H.; Ogando, D.G.; Li, S.; Feng, M.; Price, F.W., Jr.; Tennessen, J.M.; Joseph, A.; Bonanno, J.A. Glutaminolysis is Essential for Energy Production and Ion Transport in Human Corneal Endothelium. EBioMedicine 2017, 16, 292–301. [Google Scholar] [CrossRef]
- Gidfar, S.; Milani, F.Y.; Milani, B.Y.; Shen, X.; Eslani, M.; Putra, I.; Huvard, M.J.; Sagha, H.; Djalilian, A.R. Rapamycin Prolongs the Survival of Corneal Epithelial Cells in Culture. Sci. Rep. 2017, 7, 40308. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Li, X.; Yin, Y.; Wu, R.; Xu, X.; Chen, F. The effects of conditioned media generated by polarized macrophages on the cellular behaviours of bone marrow mesenchymal stem cells. J. Cell Mol. Med. 2018, 22, 1302–1315. [Google Scholar] [CrossRef]
- Jabbehdari, S.; Yazdanpanah, G.; Kanu, L.N.; Anwar, K.N.; Shen, X.; Rabiee, B.; Putra, I.; Eslani, M.; Rosenblatt, M.I.; Hematti, P.; et al. Reproducible Derivation and Expansion of Corneal Mesenchymal Stromal Cells for Therapeutic Applications. Transl. Vis. Sci. Technol. 2020, 9, 26. [Google Scholar] [CrossRef]
- Eslani, M.; Putra, I.; Shen, X.; Hamouie, J.; Tadepalli, A.; Anwar, K.N.; Kink, J.A.; Ghassemi, S.; Agnihotri, G.; Reshetylo, S.; et al. Cornea-Derived Mesenchymal Stromal Cells Therapeutically Modulate Macrophage Immunophenotype and Angiogenic Function. Stem Cells 2018, 36, 775–784. [Google Scholar] [CrossRef]
- Movahedan, A.; Afsharkhamseh, N.; Sagha, H.M.; Shah, J.R.; Milani, B.Y.; Milani, F.Y.; Logothetis, H.D.; Chan, C.C.; Djalilian, A.R. Loss of Notch1 disrupts the barrier repair in the corneal epithelium. PLoS ONE 2013, 8, e69113. [Google Scholar] [CrossRef]
- Eslani, M.; Movahedan, A.; Afsharkhamseh, N.; Sroussi, H.; Djalilian, A.R. The role of toll-like receptor 4 in corneal epithelial wound healing. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6108–6115. [Google Scholar] [CrossRef]
- Fernandes-Cunha, G.M.; Na, K.S.; Putra, I.; Lee, H.J.; Hull, S.; Cheng, Y.C.; Blanco, I.J.; Eslani, M.; Djalilian, A.R.; Myung, D. Corneal Wound Healing Effects of Mesenchymal Stem Cell Secretome Delivered within a Viscoelastic Gel Carrier. Stem Cells Transl. Med. 2019, 8, 478–489. [Google Scholar] [CrossRef]
- Saika, S.; Shiraishi, A.; Liu, C.Y.; Funderburgh, J.L.; Kao, C.W.; Converse, R.L.; Kao, W.W. Role of lumican in the corneal epithelium during wound healing. J. Biol. Chem. 2000, 275, 2607–2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samaeekia, R.; Rabiee, B.; Putra, I.; Shen, X.; Park, Y.J.; Hematti, P.; Eslani, M.; Djalilian, A.R. Effect of Human Corneal Mesenchymal Stromal Cell-derived Exosomes on Corneal Epithelial Wound Healing. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5194–5200. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, D.A.; Kunzmann, B.C.; Steinhorst, N.A.; Spitzer, M.S.; Schultheiss, M. A Porcine Corneal Endothelial Organ Culture Model Using Split Corneal Buttons. J. Vis. Exp. 2019, 152, e60171. [Google Scholar] [CrossRef] [PubMed]
- Shafiq, M.A.; Gemeinhart, R.A.; Yue, B.Y.; Djalilian, A.R. Decellularized human cornea for reconstructing the corneal epithelium and anterior stroma. Tissue Eng. Part C Methods 2012, 18, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Nien, C.J.; Massei, S.; Lin, G.; Liu, H.; Paugh, J.R.; Liu, C.Y.; Kao, W.W.; Brown, D.J.; Jester, J.V. The development of meibomian glands in mice. Mol. Vis. 2010, 16, 1132–1140. [Google Scholar] [PubMed]
- Movahedan, A.; Majdi, M.; Afsharkhamseh, N.; Sagha, H.M.; Saadat, N.S.; Shalileh, K.; Milani, B.Y.; Ying, H.; Djalilian, A.R. Notch inhibition during corneal epithelial wound healing promotes migration. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7476–7483. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| HK2 | CAAAGTGACAGTGGGTGTGG | GCCAGGTCCTTCACTGTCTC |
| PFK | GTACCTGGCGCTGGTATCTG | CCTCTCACACATGAAGTTCTCC |
| PKM2 | AAGGGTGTGAACCTTCCTGG | GCTCGACCCCAAACTTCAGA |
| IDH2 | CGCCACTATGCCGACAAAAG | ACTGCCAGATAATACGGGTCA |
| IDH3G | ACATCGAAACCAACCATAACCTG | GGCTCTTACAGTGGATGACGTT |
| GLS2 | TGCCTATAGTGGCGATGTCTCA | GTTCCATATCCATGGCTGACAA |
| ATP5S | AGCAGTTGTGTGGCGTAAAGA | CTGATGCGATCATAATCCACCTT |
| ATP5D | ACTCTTCGGTGCAGTTGTTGG | GCCTCGATTCGGATCTGGAT |
| IL-1β | CTGCTCTGGGATTCTCTTCAG | ATCTTCCTCAGCTTGTCCATG |
| IL-10 | TCTCCGAGATGCCTTCAGCAGA | TCAGACAAGGCTTGGCAACCCA |
| TNF-α | CTTCTCCTTCCTGATCGTGG | GCTGGTTATCTCTCAGCTCCA |
| GAPDH | GCCAAAAGGGTCATCATCTC | GTAGAGGCAGGGATGATGTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, S.; Shen, X.; Anwar, K.; Ashraf, M.; Lee, H.; Koganti, R.; Ghassemi, M.; Djalilian, A.R. Therapeutic Potential of Mesenchymal Stem Cell-Secreted Factors on Delay in Corneal Wound Healing by Nitrogen Mustard. Int. J. Mol. Sci. 2022, 23, 11510. https://doi.org/10.3390/ijms231911510
An S, Shen X, Anwar K, Ashraf M, Lee H, Koganti R, Ghassemi M, Djalilian AR. Therapeutic Potential of Mesenchymal Stem Cell-Secreted Factors on Delay in Corneal Wound Healing by Nitrogen Mustard. International Journal of Molecular Sciences. 2022; 23(19):11510. https://doi.org/10.3390/ijms231911510
Chicago/Turabian StyleAn, Seungwon, Xiang Shen, Khandaker Anwar, Mohammadjavad Ashraf, Hyungjo Lee, Raghuram Koganti, Mahmood Ghassemi, and Ali R. Djalilian. 2022. "Therapeutic Potential of Mesenchymal Stem Cell-Secreted Factors on Delay in Corneal Wound Healing by Nitrogen Mustard" International Journal of Molecular Sciences 23, no. 19: 11510. https://doi.org/10.3390/ijms231911510