
Homo sapiens May Incorporate Daily Acute Cycles of “Conditioning–Deconditioning” to Maintain Musculoskeletal Integrity: Need to Integrate with Biological Clocks and Circadian Rhythm Mediators
Abstract
:1. Introduction
1.1. The Purpose of This Review
1.2. Background
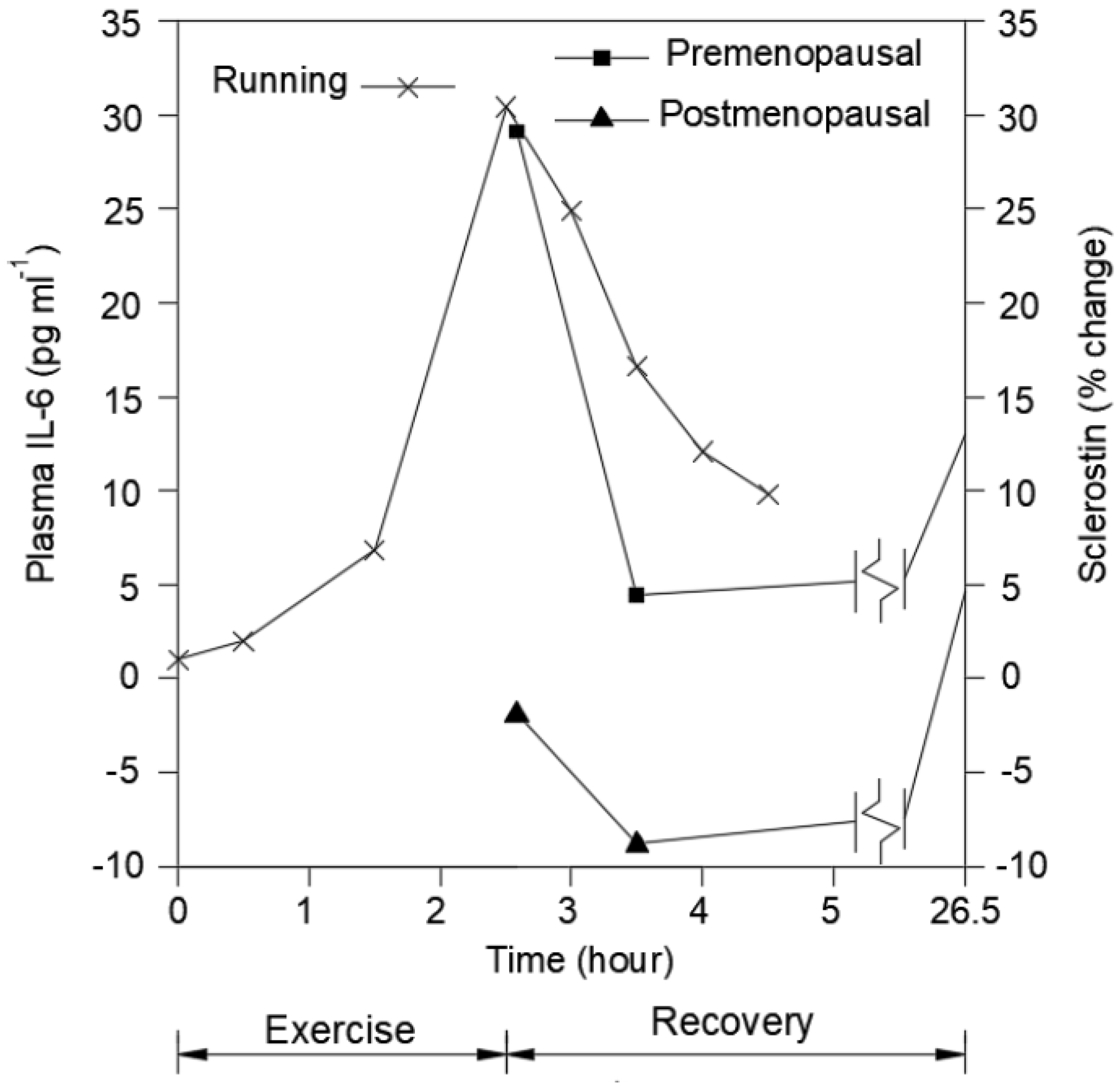
2. The Potential Role of Mediators and Receptor Expression Generated by Physical Activity and Engagement of GRFs to Set the Stage for Subsequent Effector Processes
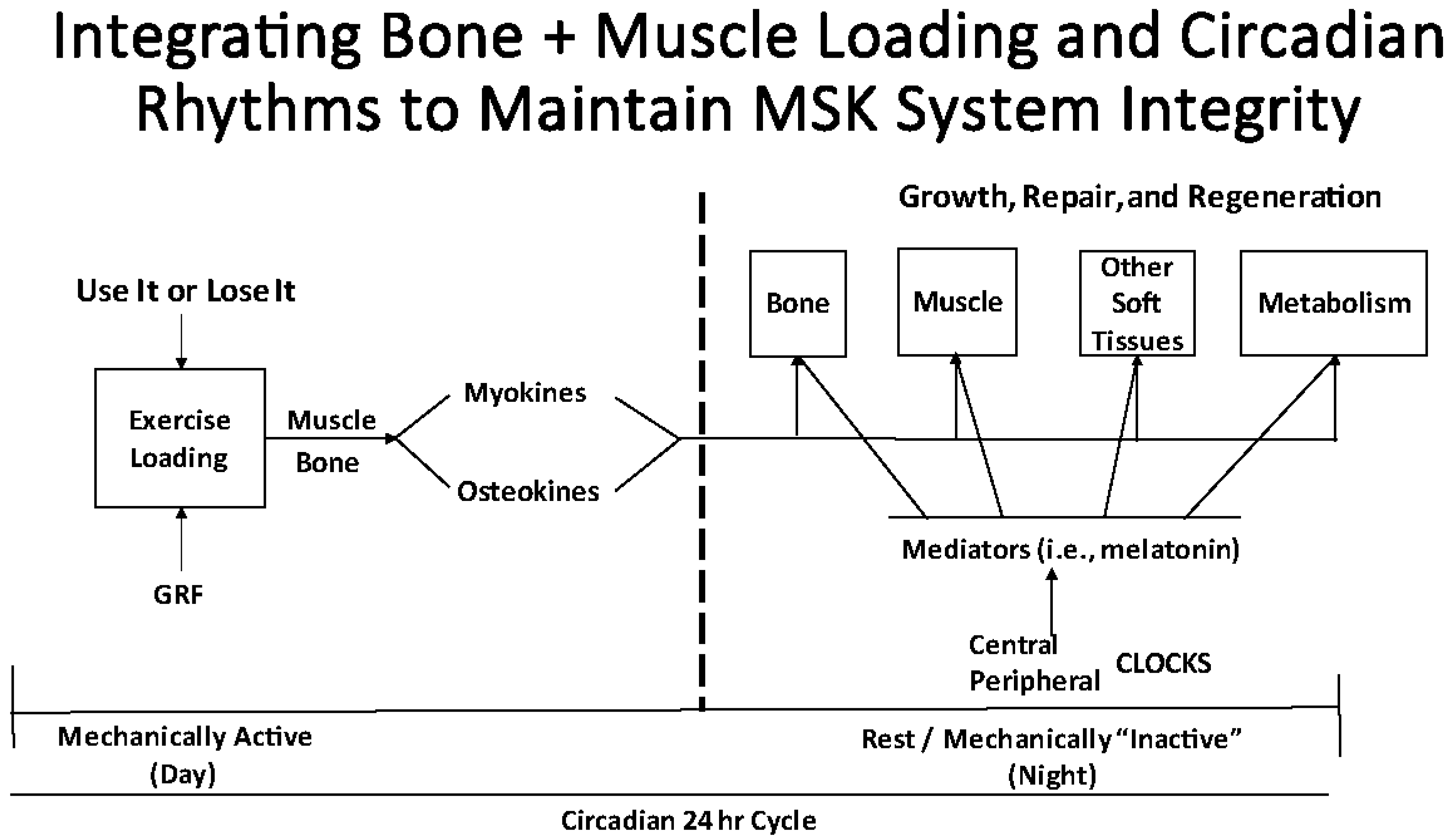
3. The Need to Integrate Activity-Generated Mediators with Circadian Rhythm Variations during Periods of Acute “Deconditioning”
3.1. Background
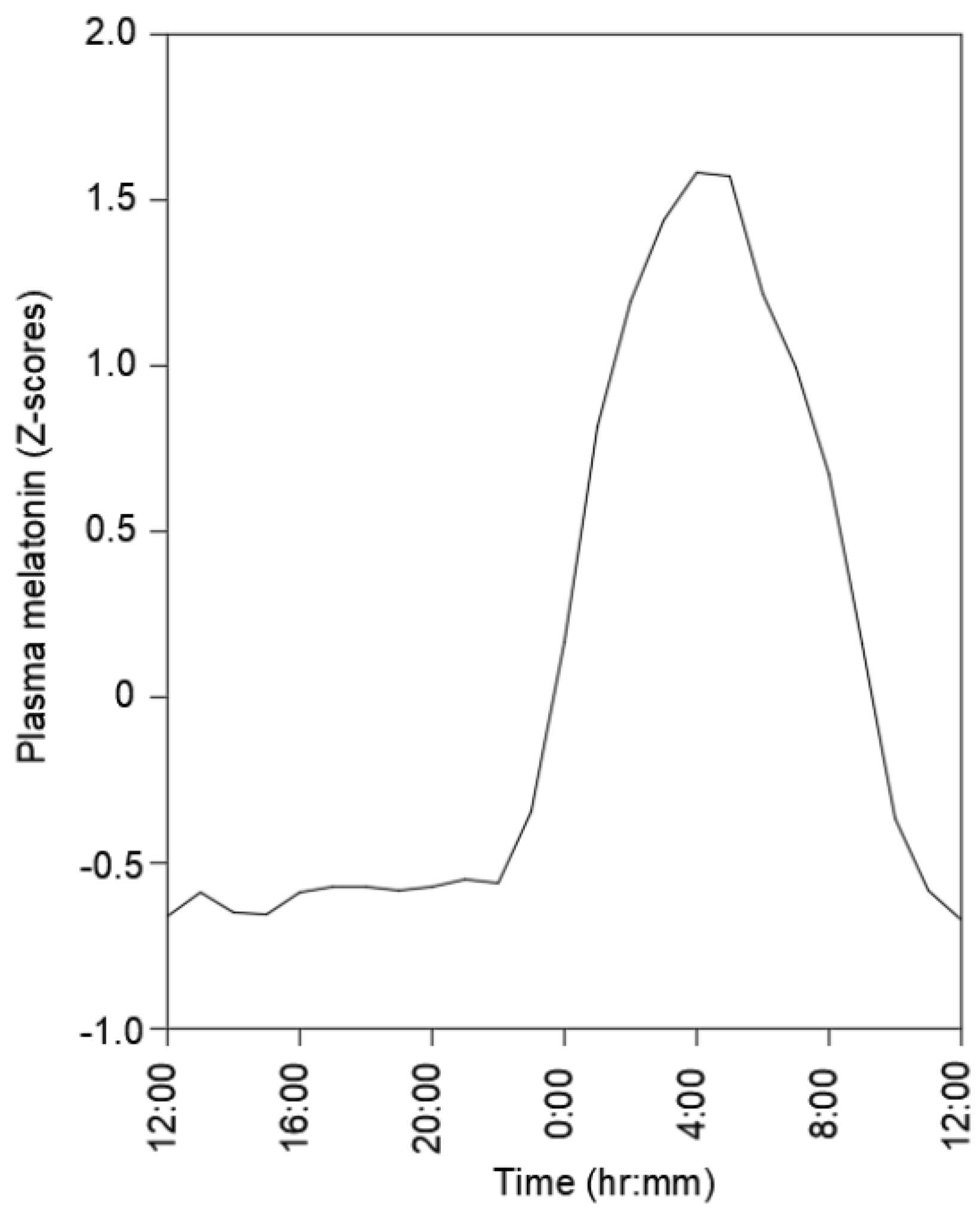
3.2. Biological Clocks, Circadian Rhythms, and Regulation of MSK Tissue Homeostasis and Repair
4. Indirect Evidence for Conditioning–Deconditioning in Regulation of Connective Tissue Growth and Maintenance
5. Why Might Cycles of Acute “Conditioning–Deconditioning” Be Important in Regulation of Lower Extremity Connective Tissues?
6. Are Responses to GRFs Mechanistically Unique Due to Exposure to Them Late in Evolution?
7. Further Testing the Acute “Conditioning/Deconditioning” Hypothesis
8. Author Opinion
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sherratt, M.J.; Hopkinson, L.; Naven, M.; Hibbert, S.A.; Ozols, M.; Eckersley, A.; Newton, V.L.; Bell, M.; Meng, Q.J. Circadian rhythms in skin and other elastic tissues. Matrix Biol. 2019, 84, 97–110. [Google Scholar] [CrossRef]
- Basti, A.; Yalcin, M.; Herms, D.; Hesse, J.; Aboumanify, O.; Li, Y.; Aretz, Z.; Garmshausen, J.; El-Athman, R.; Hastermann, M.; et al. Diurnal variations in the expression of core-clock genes correlate with resting muscle properties and predict fluctuations in exercise performance across the day. BMJ Open Sport Exerc. Med. 2021, 7, e000876. [Google Scholar] [CrossRef]
- Gabriel, B.M.; Zierath, J.R. Circadian rhythms and exercise—Re-setting the clock in metabolic disease. Nat. Rev. Endocrinol. 2019, 15, 197–206. [Google Scholar] [CrossRef]
- Hart, D.A.; Zernicke, R.F. Optimal human functioning requires exercise across the lifespan: Mobility in a 1 g environment is intrinsic to the integrity of multiple biological systems. Front. Physiol. 2020, 11, 156. [Google Scholar] [CrossRef]
- Dijk, D.J.; Shanahan, T.L.; Duffy, J.F.; Ronda, J.M.; Czeisler, C.A. Variation of electroencephalographic activity during non-rapid eye movement and rapid eye movement sleep with phase of circadian melatonin rhythm in humans. J. Physiol. 1997, 505, 851–858. [Google Scholar] [CrossRef]
- Wei, K.; Wang, Q.; Gan, J.; Zhang, S.; Ye, M.; Gragnoli, C.; Wu, R. Mapping genes for drug chronotherapy. Drug Discov. Today 2018, 23, 1883–1888. [Google Scholar] [CrossRef]
- Scherholz, M.L.; Schlesinger, N.; Androulakis, J.P. Chronopharmacology of glucocorticoids. Adv. Drug. Deliv. Rev. 2019, 151–152, 245–261. [Google Scholar] [CrossRef]
- Gaspar, L.S.; Alvaro, A.R.; Carmo-Silva, S.; Mendes, A.F.; Relogio, A.; Cavadas, C. The importance of determining circadian parameters in pharmacological studies. Br. J. Pharmacol. 2019, 176, 2827–2847. [Google Scholar] [CrossRef]
- Rogers, E.H.; Hunt, J.A.; Pekovic-Vaughan, V. Adult stem cell maintenance and tissue regeneration around the clock: Do impaired stem cell clocks drive age-associated tissue degeneration? Biogerontology 2018, 19, 497–517. [Google Scholar] [CrossRef]
- Ray, E.K. Introduction: Are aging and space effects similar? Exp. Gerontol. 1991, 26, 123–129. [Google Scholar] [CrossRef]
- Mallis, M.M.; DeRosha, C.W. Circadian rhythms, sleep, and performance in space. Aviat. Space Environ. Med. 2005, 6 (Suppl. 6), B94–B107. [Google Scholar]
- Guo, J.H.; Qu, W.M.; Chen, S.G.; Chen, X.P.; Lv, K.; Huang, Z.L.; Wu, Y.L. Keeping the right time in space: Importance of circadian clock and sleep for physiology and performance of astronauts. Mil. Med. Res. 2014, 1, 23. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Febbraio, M.A. Muscles, exercise and obesity: Skeletal muscle as a secretory organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef]
- Morgan, P.T.; Smeuninx, B.; Breen, L. Exploring the impact of obesity on skeletal muscle function in older age. Front. Nutr. 2020, 7, 569904. [Google Scholar] [CrossRef]
- Guo, A.; Li, K.; Xiao, Q. Sarcopenic obesity: Myokines as potential diagnostic biomarkers and therapeutic targets? Exp. Gerontol. 2020, 139, 111022. [Google Scholar] [CrossRef]
- Collins, K.H.; Herzog, W.; MacDonald, G.Z.; Reimer, R.A.; Rios, J.L.; Smith, I.C.; Zernicke, R.F.; Hart, D.A. Obesity, metabolic syndrome, and musculoskeletal disease: Common inflammatory pathways suggest a central role for loss of muscle integrity. Front. Physiol. 2018, 9, 112. [Google Scholar] [CrossRef]
- Kirk, B.; Feehan, J.; Lombardi, G.; Duque, G. Muscle, bone, and fat crosstalk: The biological role of myokines, osteokines, and adipokines. Curr. Osteoporos. Rep. 2020, 18, 388–400. [Google Scholar] [CrossRef]
- Dai, Z.; Wu, F.; Yeung, E.W.; Li, Y. IGF-IEc expression, regulation and biological function in different tissues. Growth Horm. IGF Res. 2010, 20, 275–281. [Google Scholar] [CrossRef]
- Janssen, A.A.M.J.L. Impact of physical exercise on endocrine aging. Front. Horm. Res. 2016, 47, 68–81. [Google Scholar] [CrossRef]
- Chang, J.S.; Kim, T.H.; Nguyen, T.T.; Park, K.S.; Kim, N.; Kong, I.D. Circulating irisin levels as a predictive biomarker for sarcopenia: A cross-sectional community-based study. Geriatr. Gerontol. Int. 2017, 17, 2266–2273. [Google Scholar] [CrossRef]
- Kim, H.J.; So, B.; Choi, M.; Kang, D.; Song, W. Resistance exercise training increases the expression of irisin concomitant with improved muscle function in aging mice and humans. Exp. Gerontol. 2015, 70, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Estell, E.G.; Le, P.T.; Vegting, Y.; Kim, H.; Wrann, C.; Bouxsein, M.L.; Nagano, K.; Baron, R.; Spiegelman, B.M.; Rosen, C.J. Irisin directly stimulates osteoclastogenesis and bone resorption in vitro and in vivo. eLife 2020, 9, e58172. [Google Scholar] [CrossRef] [PubMed]
- Battafarano, G.; Rossi, M.; Marampon, F.; Minisola, S.; Del Fattore, A. Bone control of muscle function. Int. J. Mol. Sci. 2020, 21, 1178. [Google Scholar] [CrossRef]
- Zhang, J.; Valverde, P.; Zhu, X.; Murray, D.; Wu, Y.; Yu, L.; Jiang, H.; Dard, M.M.; Huang, J.; Xu, Z.; et al. Exercise-induced irisin in bone and systemic irisin administration reveal new regulatory mechanisms of bone metabolism. Bone Res. 2017, 5, 16056. [Google Scholar] [CrossRef]
- Serbest, S.; Tiftikci, U.; Tosun, H.B.; Kisa, U. The irisin hormone profile and expression in bone tissue in the bone healing process in patients. Med. Sci. Monit. 2017, 23, 4278–4283. [Google Scholar] [CrossRef] [PubMed]
- Steensberg, A.; Toft, A.D.; Schjerling, P.; Halkjaer-Kristensen, J.; Pedersen, B.K. Plasma interleukin-6 during strenuous exercise: Role of epinephrine. Am. J. Physiol. Cell Physiol. 2001, 28, C1001–C1004. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.; Kouvelioti, R.; Theocharidis, A.; Falk, B.; Tiidus, P.; Klentrou, P. Osteokines and bone markers at rest and following plyometric exercise in pre- and postmenopausal women. BioMed Res. Int. 2020, 2020, 7917309. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gil, A.M.; Elizondo-Montemayor, L. The role of exercise in the interplay between myokines, hepatokines, osteokines, adipokines and modulation of inflammation for energy substrate redistribution and fat mass loss: A review. Nutrients 2020, 12, 1899. [Google Scholar] [CrossRef]
- Herrmann, M.; Engelke, K.; Ebert, R.; Muller-Deubert, S.; Rudert, M.; Ziouti, F.; Jundt, F.; Felsenberg, D.; Jakob, F. Interactions between muscles and bone-where physics meets biology. Biomolecules 2020, 10, 432. [Google Scholar] [CrossRef]
- Tagliaferri, C.; Wittrant, Y.; Davicco, M.J.; Walrand, S.; Coxam, V. Muscle and bone, two interconnected tissues. Aging Res. Rev. 2015, 21, 55–70. [Google Scholar] [CrossRef]
- Kaji, H. Effects of myokines on bone. BoneKEy Rep. 2016, 5, 826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cariati, I.; Bonanni, R.; Onorato, F.; Mastrogregori, A.; Rossi, D.; Iundusi, R.; Gasbarra, E.; Tancredi, V.; Tarantino, U. Role of physical activity in bone-muscle crosstalk: Biological aspects and clinical implications. J. Funct. Morphol. Kinesiol. 2021, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- He, C.; He, W.; Hou, J.; Chen, K.; Huang, M.; Yang, M.; Luo, X.; Li, C. Bone and muscle crosstalk in aging. Front. Cell Dev. Biol. 2020, 8, 585644. [Google Scholar] [CrossRef] [PubMed]
- Trajanoska, K.; Rivadeneira, F.; Kiel, D.P.; Karasik, D. Genetics of bone and muscle interactions in humans. Curr. Osteoporos. Rep. 2019, 17, 86–95. [Google Scholar] [CrossRef]
- Lau, P.; Vico, L.; Rittweger, J. Dissociation of bone resorption and formation in spaceflight and simulated microgravity: Potential role of myokines and osteokines? Biomedicines 2022, 10, 342. [Google Scholar] [CrossRef]
- Baptista, S.; Piloto, N.; Reis, F.; Teixeira-de-Lemos, E.; Garrido, A.P.; Dias, A.; Lourenco, M.; Palmeiro, A.; Ferrer-Antunes, C.; Teixeira, F. Treadmill runing and swimming imposes distinct cardiovascular physiological adaptations in the rat: Focus on serotonergic and sympathetic nervous system modulation. Acta Physiol. Hung. 2008, 95, 365–381. [Google Scholar] [CrossRef]
- Gamble, K.L.; Berry, R.; Frank, S.J.; Young, M.E. Circadian clock control of endocrine factors. Nat. Rev. Endocrinol. 2014, 10, 466–475. [Google Scholar] [CrossRef]
- Deibel, S.H.; Zelinski, E.L.; Keeley, R.J.; Kovalchuk, O.; McDonald, R.J. Epigenetic alterations in the suprachiasmatic nucleus and hippocampus contribute to age-related cognitive decline. Oncotarget 2015, 6, 23181–23203. [Google Scholar] [CrossRef]
- Lu, Q.; Kim, J.Y. Mammalian circadian networks mediated by the suprachiasmatic nucleus. FEBS J. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Van Drunen, R.; Eckel-Mahan, K. Circadian rhythms of the hypothalamus: From function to physiology. Clocks Sleep 2021, 3, 189–226. [Google Scholar] [CrossRef]
- Ashton, A.; Foster, R.G.; Jagannath, A. Photic entrainment of the circadian system. Int. J. Med. Sci. 2022, 23, 729. [Google Scholar] [CrossRef] [PubMed]
- Spitschan, M.; Santhi, N. Individual differences and diversity in human physiological responses to light. EBioMedicine 2022, 75, 103640. [Google Scholar] [CrossRef] [PubMed]
- Leliavski, A.; Dumbell, R.; Ott, V.; Oster, H. Adrenal clocks and the role of adrenal hormones in the regulation of circadian physiology. J. Biol. Rhythm. 2015, 30, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Swarbrick, M.; Zhou, H.; Seibel, M. Mechanisms in Endocrinology: Local and systemic effects of glucocorticoids on metabolism: New lessons from animal models. Eur. J. Endocrinol. 2021, 185, R113–R129. [Google Scholar] [CrossRef] [PubMed]
- Dietrich-Zagonel, F.; Aspenberg, P.; Eliasson, P. Dexamethasone enhances Achille’s tendon healing in an animal injury model, and the effects are dependent on dose, administration time, and mechanical loading stimulation. Am. J. Sports Med. 2022, 50, 1306–1316. [Google Scholar] [CrossRef]
- Steyn, F.J.; Ngo, S.T.; Sullivan, S. Endocrine rhythms of growth hormone release: Insights from animal studies. Best Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 521–533. [Google Scholar] [CrossRef]
- Nicolaides, N.C.; Chrousos, G.P. Sex differences in circadian endocrine rhythms: Clinical implications. Eur. J. Neurosci. 2020, 52, 2575–2585. [Google Scholar] [CrossRef]
- Maria, S.; Witt-Enderby, P.A. Melatonin effects on bone: Potential use for the prevention and treatment for osteopenia, osteoporosis, and periodontal disease and for use in bone-grafting procedures. J. Pineal Res. 2014, 56, 115–125. [Google Scholar] [CrossRef]
- Suzuki, N.; Somei, M.; Seki, A.; Reiter, R.J.; Hattori, A. Novel bromomelatonin derivatives as potentially effective drugs to treat bone diseases. J. Pineal Res. 2008, 45, 2290234. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Ladizesky, M.G.; Boggio, V.; Cutrena, R.A.; Mautalen, C. Melatonin effects on bone: Experimental facts and clinical perspectives. J. Pineal Res. 2003, 34, 81–87. [Google Scholar] [CrossRef]
- Amstrup, A.K.; Sikjaer, T.; Mosekilde, L.; Rejmark, L. Melatonin and the skeleton. Osteoporos. Int. 2013, 24, 2919–2927. [Google Scholar] [CrossRef]
- Igarashi-Migitaka, J.; Seki, A.; Ikegame, M.; Honda, M.; Sekiguchi, T.; Mishima, H.; Shimizu, N.; Matsubara, H.; Srivastav, A.K.; Hirayama, J.; et al. Oral administration of melatonin contained in drinking water increased bone strength in naturally aged mice. Acta Histochem. 2020, 122, 151596. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; You, W.; Shan, T. The regulatory role of melatonin in skeletal muscle. J. Muscle Res. Cell Motil. 2020, 41, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Yu, S.; Chen, G.; Zheng, W.; Peng, J.; Huang, X.; Chen, L. Insights into the roles of melatonin in bone tissue and bone-related diseases (Review). Int. J. Mol. Sci. 2021, 47, 82. [Google Scholar] [CrossRef]
- Histing, T.; Anton, C.; Scheuer, C.; Garcia, P.; Holstein, J.H.; Klein, M.; Matthys, R.; Pohlemann, T.; Menger, M.D. Melatonin impairs fracture healing by suppressing RANKL-mediated bone remodeling. J. Surg. Res. 2012, 173, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Kose, D.; Kose, A.; Halici, Z.; Ali Gurbuz, M.; Aydin, A.; Ugan, R.A.; Karaman, A.; Toktay, E. Do peripheral melatonin agonists improve bone fracture healing? The effects of agomelatine and ramelteon on experimental bone fracture. Eur. J. Pharmacol. 2020, 887, 173577. [Google Scholar] [CrossRef] [PubMed]
- Arabaci, T.; Kermen, E.; Ozkanlar, S.; Kose, O.; Kara, A.; Kozldag, A.; Duman, S.B.; Ibisoglu, E. Therapeutic effects of melatonin on alveolar bone resorption after experimental periodontitis in rats: A biochemical and immunohistochemical study. J. Periodontol. 2015, 86, 874–881. [Google Scholar] [CrossRef]
- Pei, M.; He, F.; Wei, L.; Rawson, A. Melatonin enhances cartilage matrix synthesis by procine articular chondrocytes. J. Pineal Res. 2009, 46, 181–187. [Google Scholar] [CrossRef]
- Song, W.; Ma, Z.; Wang, C.; Li, H.; He, Y. Pro-chondrogenic and immunomodulatory melatonin-loaded electrospun membranes for tendon-to-bone healing. J. Mater. Chem. B 2019, 7, 6564–6575. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, J.; Liu, J.; Yan, B.; Yan, B.; Huang, M.; Zhang, Z.; Wang, L. Melatonin promotes heterotopic ossification through regulation of endothelial-mesenchymal transition in injured Achilles tendons of rats. Front. Cell Dev. Biol. 2021, 9, 629274. [Google Scholar] [CrossRef]
- Yao, Z.; Qian, Y.; Jin, Y.; Wang, S.; Li, J.; Yuan, W.E.; Fan, C. Biomimetic multilayer polycaprolactone/sodium alginate hydrogel scaffolds loaded with melatonin facilitate tendon regeneration. Carbohydr. Polym. 2022, 277, 118865. [Google Scholar] [CrossRef]
- Liu, X.; Xu, Y.; Chen, S.; Tan, Z.; Xiong, K.; Li, Y.; Ye, Y.; Luo, Z.P.; He, F.; Gong, Y. Rescue of proinflammatory cytokine-inhibited chondrogenesis by the antiarthritic effect of melatonin in synovium mesenchymal stem cells via suppression of reactive oxygen species and matrix metalloproteinases. Free Radic. Biol. Med. 2014, 68, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, F.; Canonico, B.; Bartolini, D.; Arcangeletti, M.; Ciffolili, S.; Murdolo, G.; Piroddi, M.; Papa, S.; Reiter, R.J.; Galli, F. Melatonin regulates mesenchymal stem cell differentiation: A review. J. Pineal Res. 2014, 56, 382–397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, S.; Li, Y.; Liu, Y. Melatonin as a promising agent of regulating stem cell biology and its application in disease therapy. Pharmacol. Res. 2017, 117, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Jung, Y.H.; Oh, S.Y.; Yun, S.P.; Han, H.J. Melatonin enhances the human mesenchymal stem cells motility via melatonin receptor 2 coupling with Gaq in skin wound healing. J. Pineal Res. 2014, 57, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Lin, M.; Liang, A.; Zhang, L.; Chen, C.; Liang, G.; Xu, C.; Peng, Y.; Chen, C.; Huang, D.; et al. Melatonin enhances chondrogenic differentiation of human mesenchymal stem cells. J. Pineal Res. 2014, 56, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.H.; Lu, P.W.A.; Lu, E.W.H.; Tang, C.H.; Su, S.C.; Lin, C.W.; Yang, S.F. The potential remedy of melatonin on osteoarthritis. J. Pineal Res. 2021, 71, e12762. [Google Scholar] [CrossRef]
- Leung, M.; Tranmer, J.; Hung, E.; Korsiak, J.; Day, A.G.; Aronson, K.J. Shift work, chrontype, and melatonin patterns among female hospital employees on day and night shifts. Cancer Epidemiol. Biomark. Prev. 2016, 25, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Fuller, C. /A.; Hoban-Higgins, T.M.; Griffin, D.W.; Murakani, D.M. Influence of gravity on the circadian timing system. Adv. Space Res. 1994, 14, 399–408. [Google Scholar] [CrossRef]
- Tosini, G.; Aquzzi, J. Effect of space flight on circadian rhythms. Adv. Space Biol. Med. 2005, 10, 165–174. [Google Scholar] [CrossRef]
- Goel, N.; Bale, T.L.; Epperson, C.N.; Kornstein, S.G.; Leon, G.R.; Palinkas, L.A.; Stuster, J.W.; Dinges, D.F. Effects of sex and gender on adaptation to space: Behavioral health. J. Women’s Health 2014, 23, 975–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikegame, M.; Hattori, A.; Tabata, M.; Kitamura, K.I.; Tabouchi, Y.; Furusawa, Y.; Manuyama, Y.; Yamamoto, T.; Sekiguchi, T.; Matsuoka, R.; et al. Melatonin is a potential drug for the prevention of bone loss during space flight. J. Pineal Res. 2019, 67, e12594. [Google Scholar] [CrossRef] [PubMed]
- Chase, J.E.; Gidal, B.E. Melatonin: Therapeutic use in sleep disorders. Ann. Pharmacother. 1997, 31, 1218–1226. [Google Scholar] [CrossRef] [PubMed]
- Marqueze, E.C.; Nogueira, L.F.R.; Vetter, C.; Skene, D.J.; Cipolla-Neto, J.; Mareno, C.R.C. Exogenous melatonin decreases circadian misalignment and body weight among early types. J. Pineal Res. 2021, 71, e12750. [Google Scholar] [CrossRef] [PubMed]
- Foley, H.M.; Steel, A.E. Adverse events associated with oral administration of melatonin: A critical systematic review of clinical evidence. Complement. Ther. Med. 2019, 42, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Frost, H.M. Could some biomechanical effects of growth hormone help to explain its effects on bone formation and resorption? Bone 1998, 23, 395–398. [Google Scholar] [CrossRef]
- Hart, D.A.; Natsu-ume, T.; Sciore, P.; Tasevski, V.; Frank, C.B.; Shrive, N.G. Mechanobiology: Similarities and differences between in vivo and in vitro analysis at the functional and molecular levels. Recent Res. Dev. Biophys. Biochem. 2002, 2, 153–177. [Google Scholar]
- Kos, O.; Hughson, R.L.; Hart, D.A.; Clement, G.; Frings-Meuthen, P.; Linnarsson, D.; Paloski, W.H.; Rittweger, J.; Wuyts, F.; Zange, J.; et al. Elevated serum soluble CD200 and CD200R as surrogate markers of bone loss under bed rest conditions. Bone 2014, 60, 33–40. [Google Scholar] [CrossRef]
- Custaud, M.A.; Amaud, S.B.; Monk, R.H.; Claustrat, B.; Gharib, C.; Gauquelin-Koch, G. Hormonal changes during 17 days of head-down bedrest. Life Sci. 2003, 72, 1001–1014. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Fuentes-Broto, L. Melatonin: A multitasking molecule. Prog. Brain Res. 2010, 181, 127–151. [Google Scholar] [CrossRef]
- Chan, K.H.; Wong, Y.H. A molecular and chemical perspective in defining melatonin receptor subtype selectivity. Int. J. Mol. Sci. 2013, 14, 18385–18406. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Clough, S.J.; Hutchison, A.J.; Adamah-Biassi, E.B.; Popovska-Gorrevski, M.; Dubocovich, M.L. MT1 and MT2 melatonin receptors: A therapeutic perspective. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 361–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, S.; Frank, C.; Hart, D. Immobilization alters cell metabolism in an immature ligament. Clin. Orthop. Relat. Res. 1992, 277, 277–288. [Google Scholar] [CrossRef]
- Walsh, S.; Frank, C.; Shrive, N.; Hart, D. Knee immobilization inhibits biomechanical maturation of the rabbit medial collateral ligament. Clin. Orthop. Relat. Res. 1993, 297, 253–261. [Google Scholar] [CrossRef]
- Walsh, S. The Effects of Immobilization on the Growth and Maturation of the Rabbit Medial Collateral Ligament. Master’s Thesis, University of Calgary, Calgary, AB, Canada, 1988. [Google Scholar]
- Weir, T. Recovery of a Growing Ligament Following Immobilization in a Rabbit Model Immobilization. Master’s Thesis, University of Calgary, Calgary, AB, Canada, 1991. [Google Scholar]
- Dudek, M.; Meng, Q.J. Running on time: The role of circadian clocks in the musculoskeletal system. Biochem. J. 2014, 463, 1–8. [Google Scholar] [CrossRef]
- Yang, N.; Meng, Q.J. Circadian clocks in articular cartilage and bone: A compass in the sea of matrices. J. Biol. Rhythms 2016, 31, 415–427. [Google Scholar] [CrossRef]
- Pferdehirt, L.; Damato, A.R.; Dudek, M.; Meng, Q.J.; Herzog, E.D.; Guilak, F. Synthetic gene circuits for preventing disruption of the circadian clock due to interleukin-1-induced inflammation. Sci. Adv. 2022, 8, eabj8892. [Google Scholar] [CrossRef]
- Dudek, M.; Gossan, N.; Yang, N.; Im, H.J.; Ruckshanthi, J.P.D.; Yoshitane, H.; Li, X.; Jin, D.; Wang, P.; Boudiffa, M.; et al. The chondrocyte clock gene Bmal1 controls cartilage homeostasis and integrity. J. Clin. Investig. 2016, 126, 365–376. [Google Scholar] [CrossRef]
- Pourcet, B.; Duez, H. Circadian control of inflammasome pathways: Implications for circadian medicine. Front. Immunol. 2020, 11, 1630. [Google Scholar] [CrossRef]
- Wang, M.; Yu, D.; Zheng, L.; Hong, B.; Li, H.; Hu, X.; Zhang, K.; Mou, Y. Mechanical stress affects circadian rhythm in skeletal muscle (C2C12 myoblasts) by reduc ing Per/Cry gene expression and increasing Bmal1 gene expression. Med. Sci. Monit. 2021, 27, e928359. [Google Scholar] [CrossRef]
- Xue, Y.; Liu, P.; Wang, H.; Xiao, C.; Lin, C.; Liu, J.; Dong, D.; Fu, T.; Yang, Y.; Wang, Z.; et al. Modulation of circadian rhythms affects corneal epithelium renewal and repair in mice. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1865–1874. [Google Scholar] [CrossRef]
- Fawcett, S.; Al Kassas, R.; Dykes, I.M.; Hughes, A.T.; Ghali, F.; Ross, K. A time to heal: MicroRNA and circadian dynamics in cutaneous wound repair. Clin. Sci. 2022, 136, 579–597. [Google Scholar] [CrossRef] [PubMed]
- Ruby, C.L.; Major, R.J.; Hinrichsen, R.D. Regulation of tissue regeneration by the circadian clock. Eur. J. Neurosci. 2021, 53, 3576–3597. [Google Scholar] [CrossRef] [PubMed]
- Janich, P.; Meng, Q.J.; Benitah, S.A. Circadian control of tissue homeostasis and adult stem cells. Curr. Opin. Cell Biol. 2014, 31, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Kunimoto, T.; Okubo, N.; Minami, Y.; Fujiwara, H.; Hosokawa, T.; Asada, M.; Oda, R.; Kubo, T.; Yagita, K. A PTH-responsive circadian clock operates in ex vivo mouse femur fracture healing site. Sci. Rep. 2016, 6, 22409. [Google Scholar] [CrossRef]
- Bouchard, A.L.; Dsouza, C.; Julien, C.; Rummler, M.; Gaumond, M.H.; Cermakian, N.; Willie, B.M. Bone adaptation to mechanical loading in mice is affected by circadian rhythms. Bone 2022, 154, 116218. [Google Scholar] [CrossRef]
- Song, X.; Zhao, M.; Tang, J.; Ma, T.; Bai, H.; Wang, X.; Liu, L.; Li, T.; Xu, X.; Sheng, X.; et al. Dark-light cycle disrupts bone metabolism and suppresses joint deterioration in osteoarthritic rats. Arthritis Res. Ther. 2022, 24, 158. [Google Scholar] [CrossRef]
- Shao, P.; Ohtsuka-Isoya, M.; Shinoda, H. Circadian rhythms in serum bone markers and their relation to the effect of etidronate in rats. Chronobiol. Int. 2003, 20, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Generali, D.; Dovio, A.; Tampellini, M.; Tucci, M.; Tedoldi, S.; Torta, M.; Bonardi, S.; Allevi, G.; Aguggini, S.; Milani, M.; et al. Changes of bone marrow turnover markers and serum PTH after night or morning administration of zoledronic acid in breast cancer patients with bone metastases. Br. J. Cancer 2008, 98, 1753–1758. [Google Scholar] [CrossRef]
- Brown, S.A. Circadian clock-mediated control of stem cell division and differentiation: Beyond night and day. Development 2014, 141, 3105–3111. [Google Scholar] [CrossRef]
- Naven, M.A.; Zeef, L.A.H.; Li, S.; Humphreys, P.A.; Smith, C.A.; Pathirange, D.; Cain, S.; Woods, S.; Bates, N.; Au, M.; et al. Development of human cartilage circadian rhythm in a stem cell-chondrogenesis model. Theranostics 2022, 12, 3963–3976. [Google Scholar] [CrossRef] [PubMed]
- Kc, R.; Li, X.; Forsyth, C.B.; Voigt, R.M.; Summa, K.C.; Vitaterna, M.H.; Tryniszewska, B.; Keshavarzian, A.; Turek, F.W.; Meng, Q.J.; et al. Osteoarthritis-like pathologic changes in the knee joint induced by environmental disruption of circadian rhythms is potentiated by a high-fat diet. Sci. Rep. 2015, 5, 16896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, S.L.; Zhang, T.W.; Zhang, Q.C.; Ding, W.; Li, Z.F.; Han, G.J.; Bai, J.S.; Li, X.L.; Dong, J.; Wang, H.R.; et al. Excessive mechanical strain accelerates intervertebral disc degeneration by disrupting intrinsic circadian rhythm. Exp. Mol. Med. 2021, 53, 1911–1923. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.C.; Marin, T.C.; Azevedo, L.; Rosa-Silva, J.M.; Shea, S.A.; Thosar, S.S. Chronobiology of exercise: Evaluating the best time to exercise for greater cardiovascular and metabolic benefits. Compr. Physiol. 2022, 12, 3621–3639. [Google Scholar] [CrossRef]
- McClean, C.; Davison, G.W. Circadian clocks, redox homeostasis, and exercise: Time to connect the dots? Antioxidants 2022, 11, 256. [Google Scholar] [CrossRef]
- Mansingh, S. Handschin, C. Time to train: The involvement of the molecular clock in exercise adaptation of skeletal muscle. Front. Physiol. 2022, 13, 902031. [Google Scholar] [CrossRef]
- Chow, L.S.; Gerszten, R.E.; Raylor, J.M.; Pedersen, B.K.; van Praag, H.; Trappe, S.; Febbraio, M.A.; Galis, Z.S.; Gao, Y.; Haus, J.M.; et al. Exerkines in health, relilience and disease. Nat. Rev. Endocrinol. 2022, 18, 273–289. [Google Scholar] [CrossRef]
- Frankova, S.; Parizkova, J.; Mikulecka, A. Spontaneous wheel running in laboratory rats genetically selected for activity level in a novel environment. Act. Nerv. Super. 1987, 29, 83–91. [Google Scholar]
- Morishima-Yamato, M.; Hisaoka, F.; Shinomiya, S.; Harada, N.; Matoba, H.; Takahashi, A.; Nakaya, Y. Clioning and establishment of a line of rats for high levels of voluntary wheel running. Life Sci. 2005, 775, 551–561. [Google Scholar] [CrossRef]
- Acosta, W.; Meek, T.H.; Schultz, H.; Dlugosz, E.M.; Garland, T., Jr. Preference for Western diet coadapts in high runner mice and affects voluntary exercise and spontaneous physical activity in a genotype-dependent manner. Behav. Process. 2017, 135, 56–65. [Google Scholar] [CrossRef]
- Vaanholt, L.M.; Jonas, I.; Doornbos, M.; Schubert, K.A.; Nyakas, C.; Garland, T.; Visser, G.H.; van Dijk, G. Metabolic and behavioral responses to high-fat feeding in mice selectively bred for high wheel-running activity. Int. J. Obes. 2008, 32, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Greendale, G.A.; Witt-Erderby, P.; Karlamargla, A.S.; Munmin, F.; Crawford, S.; Huang, M.H.; Santoro, N. Melatonin patterns and levels during the human menstrual cycle and after menopause. J. Endocr. Soc. 2020, 4, bvaa115. [Google Scholar] [CrossRef] [PubMed]
- Paula, A.B.R.; de Coutinho Miranda, D.; Nogueira, F.T.; de Lauro Castrucci, A.M.; Isoldi, M.C. Does a high-fat diet affect the circadian clock, or is it the other way around? A systematic review. Nutr. Res. 2020, 84, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Vieira, E.; Mirizio, G.G.; Barin, G.R.; de Andrade, R.V.; Nimer, N.F.S.; La Sala, L. Clock genes, inflammation and the immune system-implications for diabetwes, obesity and neurodegenerative diseases. Int. J. Mol. Sci. 2020, 21, 9743. [Google Scholar] [CrossRef]
- Sato, T.; Sassone-Corsi, P. Nutrition, metabolism, and epigenetics: Pathways of circadian reprogramming. EMBO Rep. 2022, 23, e52412. [Google Scholar] [CrossRef] [PubMed]
- Rios, J.L.; Bomhof, M.R.; Reimer, R.A.; Hart, D.A.; Collins, K.H.; Herzog, W. Protective effect of prebiotic and exercise intervention on knee health in a rat model of diet-induced obesity. Sci. Rep. 2019, 9, 3893. [Google Scholar] [CrossRef]
- Rios, J.L.; Hart, D.A.; Reimer, R.A.; Herzog, W. Prebiotic and exercise do not alter knee osteoarthritis in a rat model of established obesity. Cartilage 2021, 13 (Suppl. 2), 1456S–1466S. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Wisor, J.P. Multi-modal regulation of circadian physiology by interactive features of biological clocks. Biology 2021, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Boivin, D.B.; Boudreau, P.; Kosmadopoulos, A. Disturbance of the circadian system in shift work and its health impact. J. Biol. Rhythms 2022, 37, 3–28. [Google Scholar] [CrossRef]
- Lanyon, L.E.; Rubin, C.T. Static vs dynamic loads as an influence on bone remodelling. J. Biomech. 1984, 17, 897–905. [Google Scholar] [CrossRef]
- Rubin, C.T.; Lanyon, L.E. Kappa Delta Award paper. Osteoregulatory nature of mechanical stimuli: Function as a determinant for adaptive remodelling in bone. J. Orthop. Res. 1987, 5, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kordsmeier, J.; Xiong, J. New advances in osteocyte mechanotransduction. Curr. Osteoporos. Rep. 2021, 19, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Li, M.C.M.; Chow, S.K.H.; Wong, R.M.Y.; Qin, L.; Cheung, W.H. The role of osteocytes-specific molecular mechanism in regulation of mechanotransduction- a systematic review. J. Orthop. Transl. 2021, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Frost, H.M. From Wolff’s law to the mechanstat: A new “face” of physiology. J. Orthop. Sci. 1998, 3, 282–286. [Google Scholar] [CrossRef]
- Frost, H.M. Changing concepts in skeletal physiology: Wolff’s Law, the mechanostat, and the “Utah Paradigm”. Am. J. Hum. Biol. 1998, 10, 599–605. [Google Scholar] [CrossRef]
- Frost, H.M. Bone’s mechanostat: A 2003 update. Anat. Rec. Discov. Mol. Cell Evol. Biol. 2003, 275, 1081–1101. [Google Scholar] [CrossRef]
- Martin, R.B.; Burr, D.B.; Sharkey, N.A.; Fyhrie, D.P. Mechanical adaptability of the skeleton. In Skeletal Tissue Mechanics, 2nd ed.; Springer: New York, NY, USA, 2015; pp. 275–354. [Google Scholar]
- Cavanagh, P.R.; Licata, A.A.; Rice, A.J. Exercise and pharmacological countermeasures for bone loss during long-duration space flight. Gravit. Space Biol. Bull. 2005, 18, 39–58. [Google Scholar]
- Cavanagh, P.R.; Genc, K.O.; Gopalakrishnan, R.; Kuklis, M.M.; Maender, C.C.; Rice, A.J. Foot forces during typical days on the international space station. J. Biomech. 2010, 43, 182–188. [Google Scholar] [CrossRef]
- McClanahan, B.S.; Harmon-Clayton, K.; Ward, K.D.; Klesges, R.C.; Vudadinovich, C.M.; Cantler, E.D. Side-to-side comparison of bone mineral density in upper and lower limbs of collegiate athletes. J. Strength Cond. Res. 2002, 16, 586–590. [Google Scholar]
- Scofield, K.L.; Hecht, S. Bone health in endurance athletes: Runners, cyclists and swimmers. Curr. Sports Med. Rep. 2012, 11, 328–334. [Google Scholar] [CrossRef]
- Taylor, W.R.; Schytz, P.; Bergmann, G.; List, R.; Rostolka, B.; Hitz, M.; Dymke, J.; Damm, P.; Duda, G.; Gerber, H. A comprehensive assessment of the musculoskeletal system: The CANS-Knee data set. J. Biomech. 2017, 65, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Aboulossein, A.; Awad, M.; Maqbool, H.F.; Crisp, C.; Stewart, T.D.; Messenger, N.; Richardson, R.C.; Dehghani-Sanij, A.A.; Bradley, D. Foot trajectories and loading rates in a transfemoral amputee for six different commercial prosthetic knees: An indication of adaptability. Med. Eng. Phys. 2019, 68, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Polk, J.D.; Blumenfeld, J.; Ahluwalia, K. Knee posture predicted from subchondral apparent density in the distal femur: An experimental validation. Anat. Rec. 2008, 291, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Colaianni, G.; Mongelli, T.; Cuscito, C.; Pignataro, P.; Lippo, L.; Spiro, G.; Notarnicola, A.; Seven, I.; Passeri, G.; Mori, G.; et al. Irisin prevents and restores bone loss and muscle atrophy in hind-limb suspended mice. Sci. Rep. 2017, 7, 2811. [Google Scholar] [CrossRef]
- Trudel, G.; Kilborn, S.H.; Uhthoff, H.K. Bone growth increases the knee flexion contracture angle: A study using rats. Arch. Phys. Med. Rehabil. 2001, 82, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Lowrance, E.W. Roentgenographic record of skeletal growth in relation to age and body weight of the rabbit; calcaneous and tibia. Growth 1953, 17, 183–189. [Google Scholar]
- Masoud, I.; Shapiro, F.; Kent, R.; Moses, A. A longitudinal study of the growth of the New Zealand White rabbit cumulative and bioweekly incremental growth rates for body length, body weight, femoral length and tibial length. J. Orthop. Res. 1986, 4, 221–231. [Google Scholar] [CrossRef]
- Stavnichuk, M.; Mikolajewicz, N.; Corlett, T.; Morris, M.; Komarova, S.V. A systematic review and meta-analysis of bone loss in space travellers. NPJ Microgravity 2020, 6, 13. [Google Scholar] [CrossRef]
- Gross, T.S.; Poliachik, S.L.; Ausk, B.J.; Sanford, D.A.; Becker, B.A.; Srinivasan, S. Why rest stimulates bone formation: A hypothesis based on complex adaptive phenomenon. Exerc. Sport Sci. Rev. 2004, 32, 9–13. [Google Scholar] [CrossRef]
- Srinivasan, S.; Ausk, B.J.; Bain, S.D.; Gardiner, E.M.; Kwon, R.Y.; Gross, T.S. Rest intervals reduce the number of loading bouts required to enhance bone formation. Med. Sci. Sports Exerc. 2015, 47, 1095–1103. [Google Scholar] [CrossRef]
- Srinivasan, S.; Ausk, B.J.; Poliachik, S.L.; Warner, S.E.; Richardson, T.S.; Gross, T.S. Rest-inserted loading rapidly amplifies the response of bone to small increases in strain and load cycles. J. Appl. Physiol. 2007, 102, 1945–1952. [Google Scholar] [CrossRef] [PubMed]
- LaMothe, J.M.; Zernicke, R.F. Rest insertion combined with high-frequency loading enhances osteogenesis. J. Appl. Physiol. 2004, 96, 1788–1793. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, H.M.; Kontulainen, S.A.; Khan, K.M.; McKay, H.A. Is a school-based physical activity intervention effective for increasing tibial bone strength in boys and girls? J. Bone Miner. Res. 2007, 22, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Harding, A.T.; Weeks, B.K.; Lambert, C.; Watson, S.L.; Weis, L.J.; Beck, B.R. A comparison of bone-targeted exercise strategies to reduce fracture risk in middle-aged and older men with osteopenia and osteoporosis: LIFTMOR-M semi-randomized controlled trial. J. Bone Miner. Res. 2020, 35, 1404–1414. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Meng, H.; Inamder, S.; Das, B.; Gupta, H.; Wang, W.; Thompson, C.L.; Knight, M.M. Activation of TRPV4 by mechanical, osmotic or pharmacological stimulation is anti-inflammatory blocking IL-1B mediated articular cartilage matrix destruction. Osteoarthr. Cartil. 2021, 29, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Li, X.; Li, J.; Wang, X.; Liu, D.; Zhai, L.; Ding, B.; Li, G.; Sun, Y.; Yokota, H.; et al. Mechanical loading mitigates osteoarthritis symptoms by regulating the inflammatory microenvironment in a mouse model. Ann. N. Y. Acad. Sci. 2022, 1512, 141–153. [Google Scholar] [CrossRef]
- Torzilli, P.A.; Bhargava, M.; Park, S.; Chen, C.T.C. Mechanical load inhibits IL-1 induced matrix degradation in articular cartilage. Osteoarthr. Cartil. 2010, 18, 97–105. [Google Scholar] [CrossRef]
- Torzilli, P.A.; Bhargava, M.; Chen, C.T. Mechanical loading of articular cartilage reduces IL-1 induced enzyme expression. Cartilage 2011, 2, 364–373. [Google Scholar] [CrossRef]
- McNulty, A.L.; Estes, B.T.; Wilusz, R.E.; Weinberg, J.B.; Guilak, F. Dynamic loading enhances integrative meniscal repair in the presence of interleukin. Osteoarthr. Cartil. 2010, 18, 830–838. [Google Scholar] [CrossRef]
- Radin, E.L.; Burr, D.B.; Caterson, B.; Fyhrie, D.; Brown, T.D.; Boyd, R.D. Mechanical determinants of osteoarthrosis. Semin. Arthritis Rheum. 1991, 3 (Suppl. 2), 12–21. [Google Scholar] [CrossRef]
- Frank, C.B.; Shrive, N.G.; Boorman, R.S.; Lo, I.K.Y.; Hart, D.A. New perspectives on bioengineering of joint tissues: Joint adaptation creates a moving target for engineering replacement tissues. Ann. Biomed. Eng. 2004, 32, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Natsu-ume, T.; Majima, T.; Reno, C.; Shrive, N.G.; Frank, C.B.; Hart, D.A. Menisci of the rabbit knee require mechanical loading to maintain homeostasis: Cyclic hydrostatic compression in vitro prevents de-repression of catabolic genes. J. Orthop. Sci. 2005, 10, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Laskou, F.; Patel, H.P.; Cooper, C.; Dennison, E. A pas de deux of osteoporosis and sarcopenia: Osteosarcopenia. Climacteric 2022, 25, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Buvinic, S.; Balanta-Melo, J.; Kupczik, K.; Vasquez, W.; Beato, C.; Toro-Ibacache, V. Muscle-bone crosstalk in the masticatory system: From biomechanical to molecular interactions. Front. Endocrinol. 2021, 11, 606947. [Google Scholar] [CrossRef] [PubMed]
- McDermott, M.M.; Ferrucci, L.; Gonzalez-Freire, M.; Kosmac, K.; Leeuwenburgh, C.; Peterson, C.A.; Saini, S.; Sufit, R. Skeletal muscle pathology in peripheral artery disease: A brief review. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2577–2585. [Google Scholar] [CrossRef]
- McDermott, M.M.; Spring, B.; Tian, L.; Treat-Jacobson, D.; Ferrucci, L.; Lloyd-Jones, D.; Zhao, L.; Polonsky, T.; Kibbe, M.R.; Bazzano, L.; et al. Effect of low-intensity vs high-intensity home-based walking exercise on walk distance in patients with peripheral artery disease: The LITE randomized clinical trial. JAMA 2021, 325, 1266–1276. [Google Scholar] [CrossRef]
- Biewener, A.A. Musculoskeletal design in relation to body size. J. Biomech. 1991, 24 (Suppl. 1), 19–29. [Google Scholar] [CrossRef]
- Rittweger, J.; Gunga, H.C.; Felsenberg, D.; Kirsch, K.A. Muscle and bone-aging and space. J. Gravit. Physiol. 1999, 6, P133–P136. [Google Scholar]
- Ferretti, J.L.; Cointry, G.R.; Capozza, R.F.; Frost, H.M. Bone mass, bone strength, muscle-bone interactions, osteopenia and osteoporosis. Mech. Ageing Dev. 2003, 24, 269–279. [Google Scholar] [CrossRef]
- Hood, D.M.; Wagner, I.P.; Taylor, D.D.; Brumbaugh, G.W.; Chaffin, M.K. Voluntary limb-load distribution in horses with acute and chronic laminitis. Am. J. Vet. Res. 2001, 62, 1393–1398. [Google Scholar] [CrossRef]
- Hughes, J.M.; Petit, M.A. Biological underpinnings of Frost’s mechanostat thresholds: The important role of osteocytes. J. Musculoskelet. Neuronal Interact. 2010, 10, 128–135. [Google Scholar] [PubMed]
- Goodship, A.E.; Cunningham, J.L.; Oganov, V.; Darling, J.; Miles, A.W.; Owen, G.W. Bone loss during long term space flight is prevented by the application of a short term implusive mechanical stimulus. Acta Astronaut. 1998, 43, 65–75. [Google Scholar] [CrossRef]
- James, F.O.; Cermakian, N.; Boivin, D.B. Circadian rhythms of melatonin, cortisol, and clock gene expression during simulated night shift work. Sleep 2007, 30, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Ritonja, J.; Aronson, K.J.; Leung, M.; Flaten, L.; Topouza, D.G.; Duan, Q.L.; Durocher, F.; Tranmer, J.E.; Bhatti, P. Investigating the relationship between melatonin patterns and methylation in circadian genes among day shift and night shift workers. Occup. Environ. Med. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Stevens, R.G.; Zhu, Y. Electric light, particularly at night, disrupts human circadian rhythmicity: Is that a problem? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140120. [Google Scholar] [CrossRef]
- Page, A.J. Gastrointestinal vagal afferents and food intake: Relevance of circadian rhythms. Nutrients 2021, 13, 844. [Google Scholar] [CrossRef]
- Basolo, A.; Genzano, S.B.; Piaggi, P.; Krakoff, J.; Santini, F. Energy balance and control of body weight: Possible effects of meal timing and circadian rhythm dysregulation. Nutrients 2021, 13, 3276. [Google Scholar] [CrossRef]
- Sinturel, F.; Spaleniak, W.; Dibner, C. Circadian rhythm of lipid metabolism. Biochem. Soc. Trans. 2022, 50, 1191–1204. [Google Scholar] [CrossRef]
- Li, Y.; Ma, J.; Yao, K.; Su, W.; Tan, B.; Wu, X.; Huang, X.; Li, T.; Yin, Y.; Tosini, G.; et al. Circadian rhythms and obesity: Timekeeping governs lipid metabolism. J. Pineal Res. 2020, 69, e12682. [Google Scholar] [CrossRef]
- Kawai, M. Disruption of the circadian rhythms and its relationship with pediatric obesity. Pediatr. Int. 2022, 64, e14992. [Google Scholar] [CrossRef]
- Grosshans, M.; Vollmert, C.; Vollstaedt-Klein, S.; Nolte, I.; Schwarz, E.; Wagner, X.; Leweke, M.; Mutschler, J.; Kiefer, F.; Bumb, J.M. The association of pineal gland volume and body mass in obese and normal weight individuals: A pilot study. Psychiatr. Danub. 2016, 28, 220–224. [Google Scholar] [PubMed]
- Davis, I.S.; Rice, H.M.; Wearing, S.C. Why forefoot striking in minimal shoes might positively change the course of running injuries. J. Sport Health Sci. 2017, 6, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Diamantopoulou, Z.; Castro-Giner, F.; Schwab, F.D.; Foerster, C.; Saini, M.; Budinjas, S.; Strittmatter, K.; Krol, I.; Seifert, B.; Heinzelmann-Schwarz, V.; et al. The metastatic spread of breast cancer accelerates during sleep. Nature 2022, 607, 156–162. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Tissue/Condition | Species | Type | Year | Citation |
|---|---|---|---|---|
| Skeletal Muscle | General | Review | 2020 | Chen et al. [53] |
| Bone | General | Review | 2021 | Lu et al. [54] |
| Mice | Original | 2012 | Histing et al. [55] | |
| Rats | Original | 2020 | Kose et al. [56] | |
| Rats | Original | 2015 | Arabaci et al. [57] | |
| Cartilage | Pig | Original | 2009 | Pei et al. [58] |
| Tendon | Rat | Original | 2019 | Song et al. [59] |
| Rat | Original | 2021 | Zhang et al. [60] | |
| Rat | Original | 2022 | Yao et al. [61] | |
| Adult Stem Cells | Human | Original | 2014 | Liu et al. [62] |
| General | Review | 2014 | Luchetti et al. [63] | |
| General | Review | 2017 | Zhang et al. [64] | |
| Human | Original | 2014 | Lee et al. [65] | |
| Human | Original | 2014 | Gao et al. [66] | |
| Wound Healing | Human/Mice | Original | 2014 | Lee et al. [65] |
| Osteoarthritis | General | Review | 2021 | Lu et al. [67] |
| Tissue/Condition | Species | Type | Year | Citation |
|---|---|---|---|---|
| Corneal Repair | Mice | Original | 2017 | Xue et al. [93] |
| Skin Wounds | General | Review | 2022 | Fawcett et al. [94] |
| Tissue Regeneration | General | Review | 2021 | Ruby et al. [95] |
| Tissue Homeostasis | General | Review | 2014 | Janich et al. [96] |
| Fracture Healing | Mice | Original | 2016 | Kunimoto et al. [97] |
| Bone Adaptation | Mice | Original | 2022 | Bouchard et al. [98] |
| Bone Turnover | Rats | Original | 2022 | Song et al. [99] |
| Bisphosphonates | Rats | Original | 2003 | Shao et al. [100] |
| Bisphosphonates | Patients | Clinical Trial | 2008 | Generali et al. [101] |
| Adult Stem Cells | General | Review | 2014 | Brown [102] |
| General | Review | 2014 | Janich et al. [96] | |
| Embryonic Stem Cells | Human | Original | 2022 | Naven et al. [103] |
| High Fat Diet/OA | Mice | Original | 2015 | Kc et al. [104] |
| Post-Traumatic OA | Rats | Original | 2022 | Song et al. [99] |
| IVD Degeneration | Rats | Original | 2021 | Ding et al. [105] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hart, D.A.; Zernicke, R.F.; Shrive, N.G. Homo sapiens May Incorporate Daily Acute Cycles of “Conditioning–Deconditioning” to Maintain Musculoskeletal Integrity: Need to Integrate with Biological Clocks and Circadian Rhythm Mediators. Int. J. Mol. Sci. 2022, 23, 9949. https://doi.org/10.3390/ijms23179949
Hart DA, Zernicke RF, Shrive NG. Homo sapiens May Incorporate Daily Acute Cycles of “Conditioning–Deconditioning” to Maintain Musculoskeletal Integrity: Need to Integrate with Biological Clocks and Circadian Rhythm Mediators. International Journal of Molecular Sciences. 2022; 23(17):9949. https://doi.org/10.3390/ijms23179949
Chicago/Turabian StyleHart, David A., Ronald F. Zernicke, and Nigel G. Shrive. 2022. "Homo sapiens May Incorporate Daily Acute Cycles of “Conditioning–Deconditioning” to Maintain Musculoskeletal Integrity: Need to Integrate with Biological Clocks and Circadian Rhythm Mediators" International Journal of Molecular Sciences 23, no. 17: 9949. https://doi.org/10.3390/ijms23179949