
Evogliptin Directly Inhibits Inflammatory and Fibrotic Signaling in Isolated Liver Cells

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
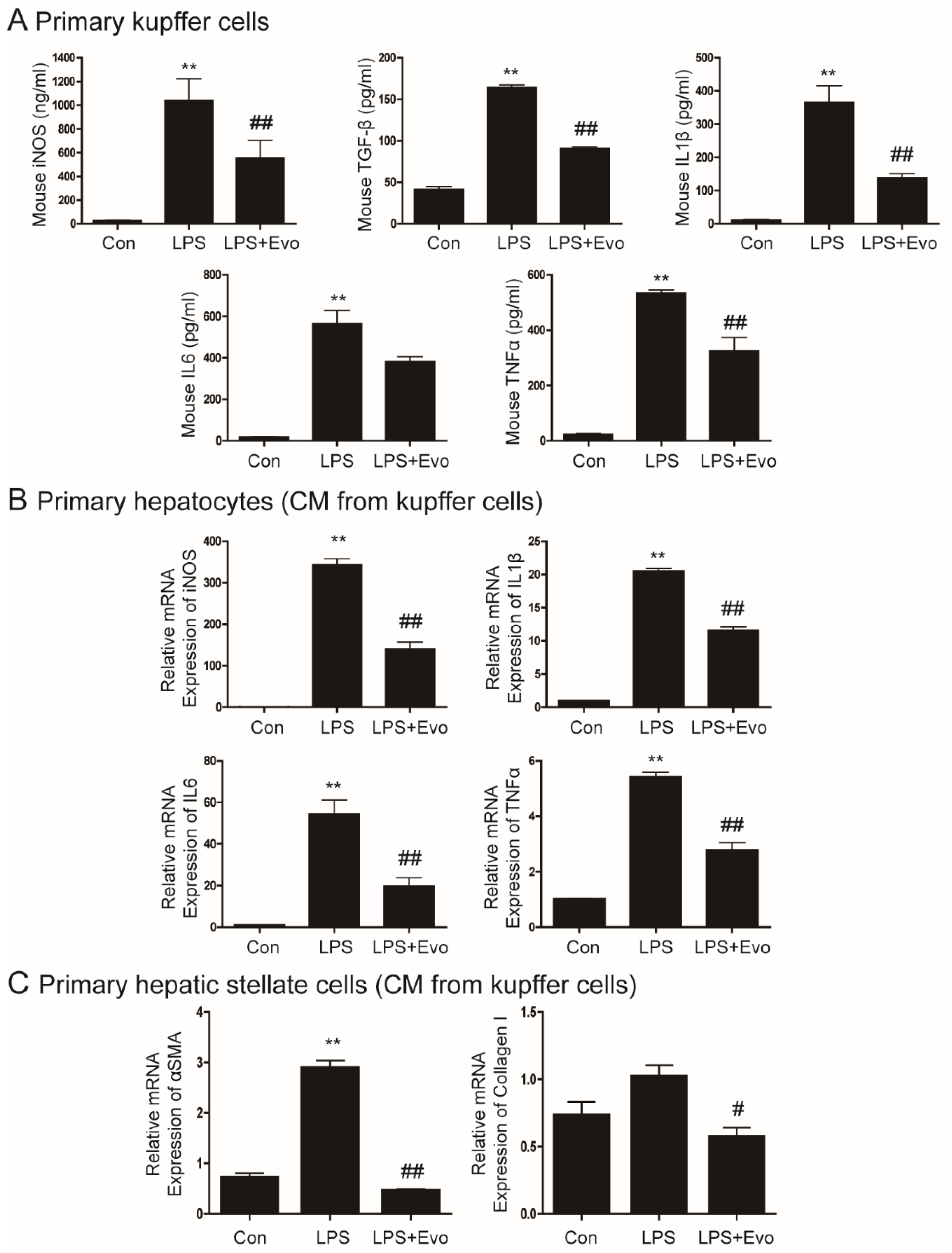
2.1. Evogliptin Inhibited Cytokine Secretion from Kupffer Cells
2.2. Evogliptin Inhibited Hepatocyte Inflammation and Hepatic Stellate Cell Activation Caused by KC-Secreted Cytokines
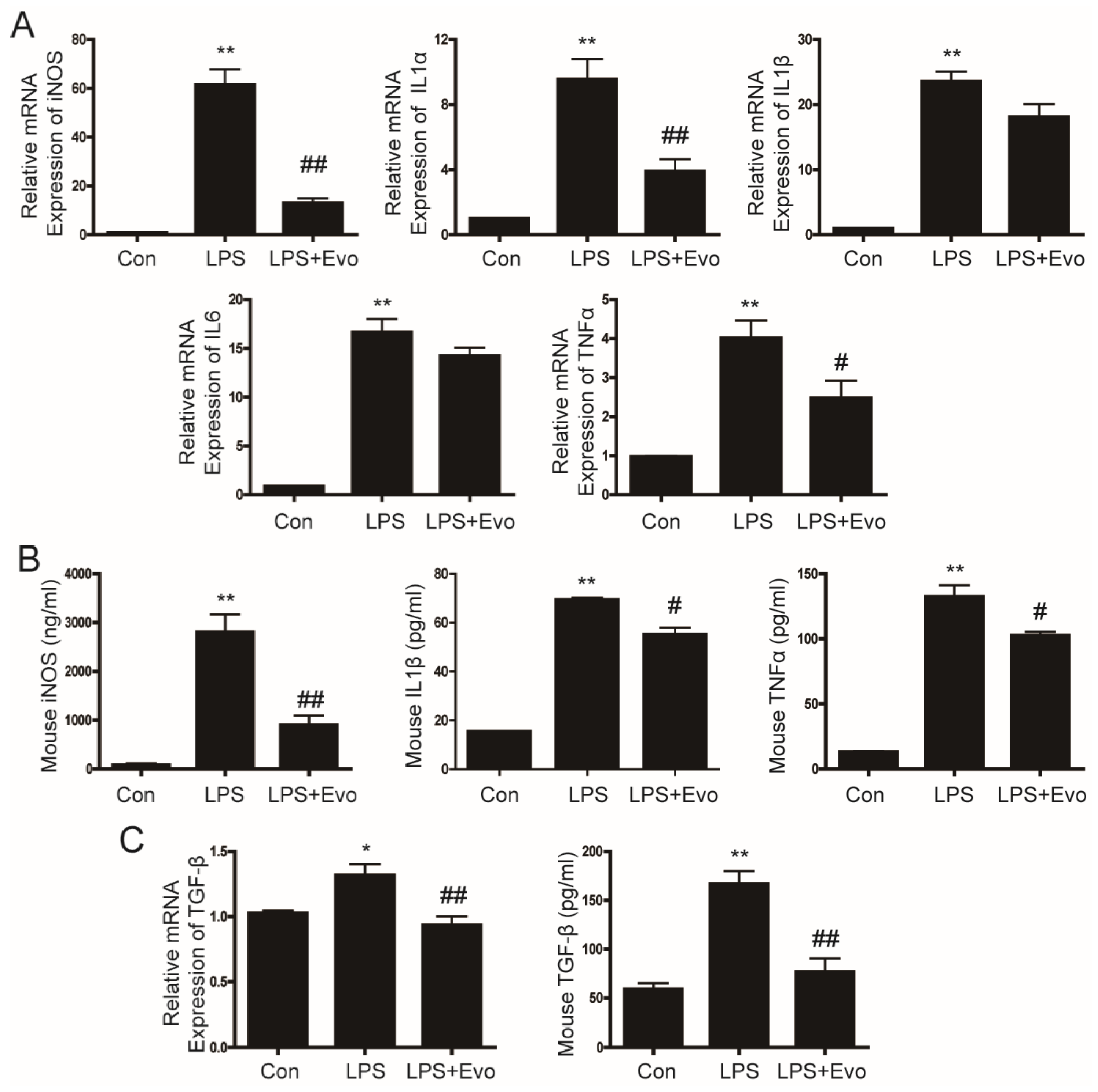
2.3. Evogliptin Inhibited LPS-Stimulated Inflammatory Cytokine Levels in Primary Hepatocytes
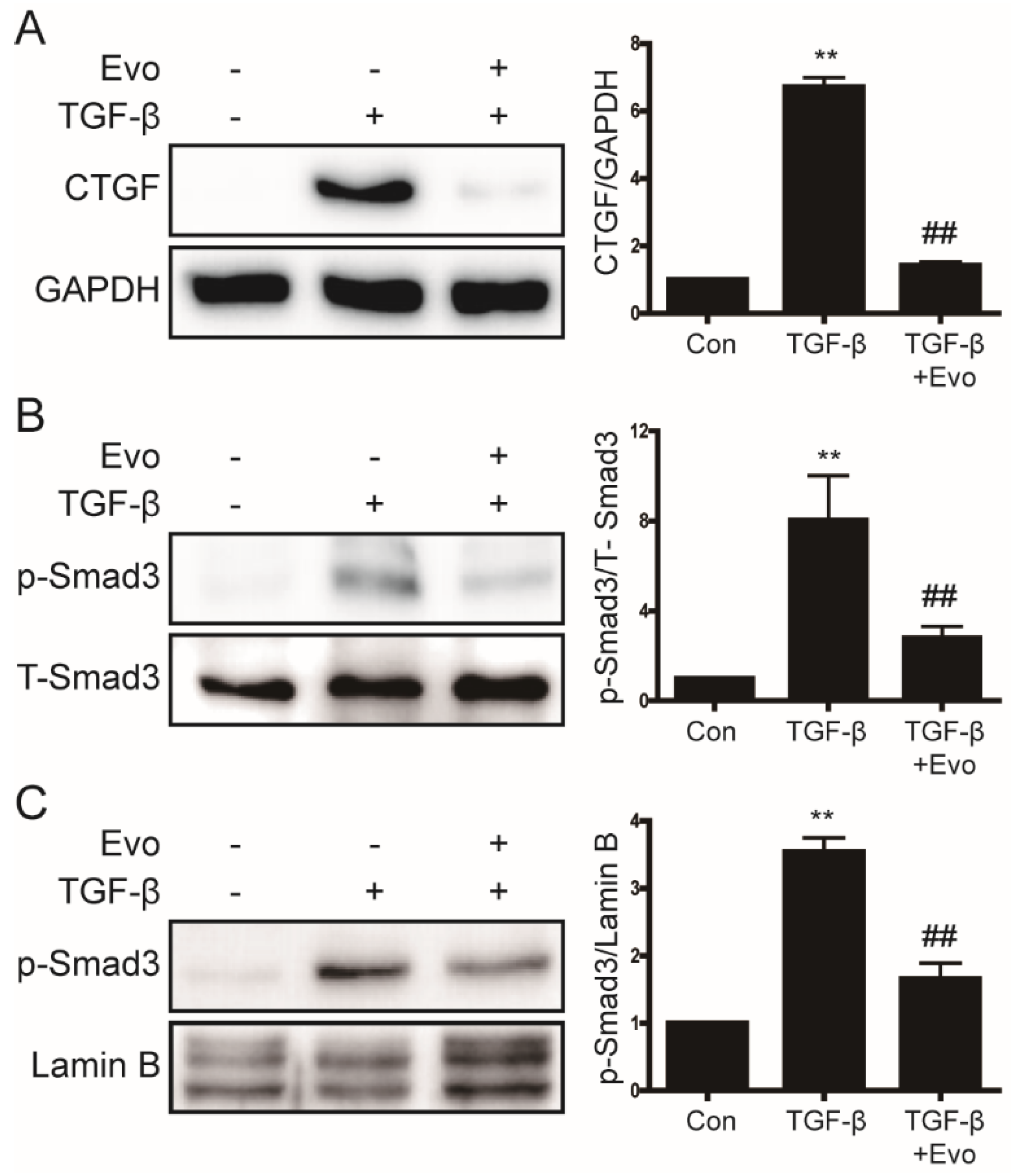
2.4. Evogliptin Inhibited TGF-β-Induced CTGF Expression and Smad3 Phosphorylation
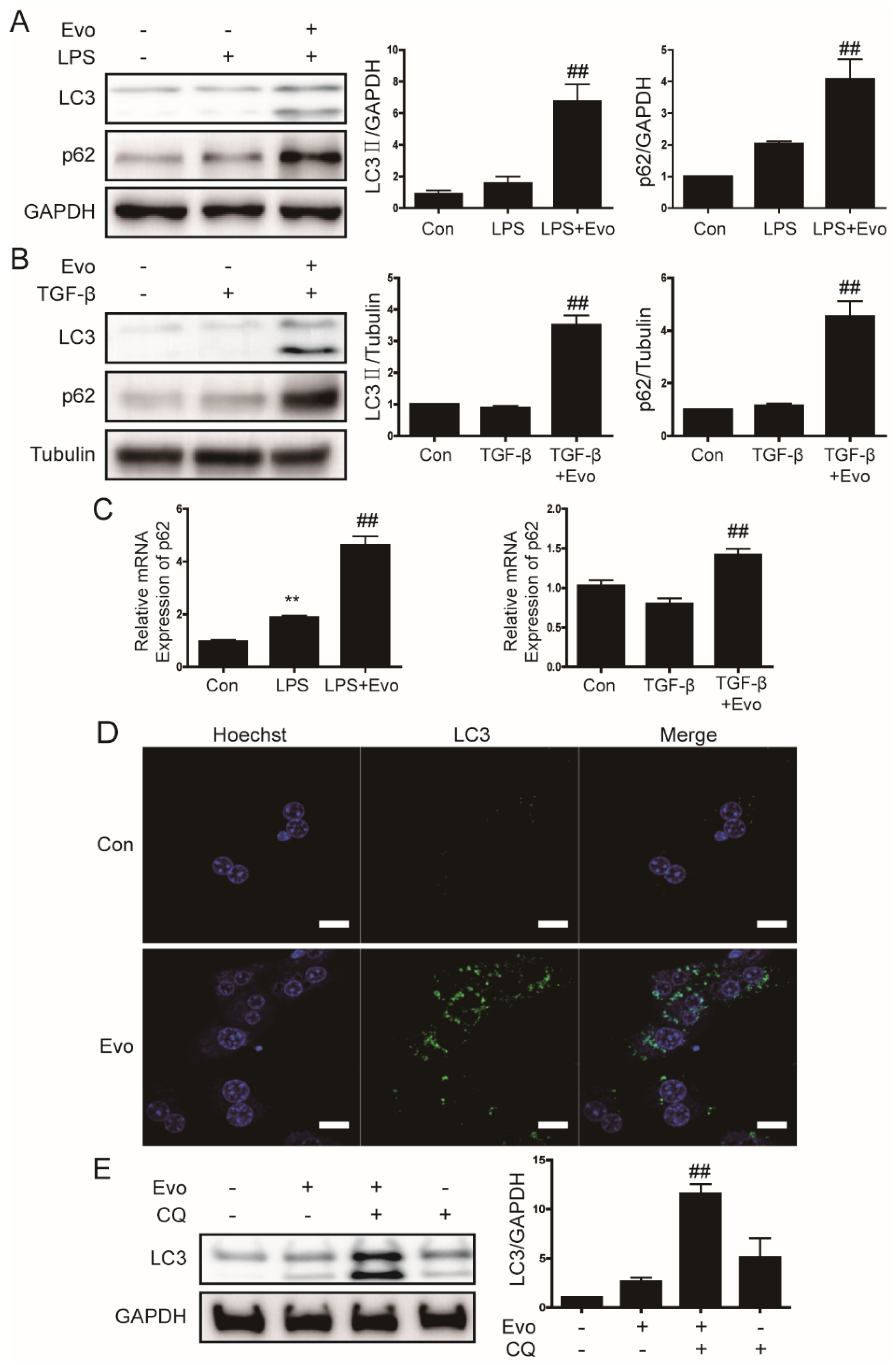
2.5. Evogliptin Increased Autophagy Flux
2.6. Inhibition of Inflammatory Signaling by Evogliptin Did Not Require Modulation of the Nrf2/HO-1 Pathway
2.7. Evogliptin Prevents Inflammatory and Fibrotic Signaling through Autophagy Induction
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Isolation of Primary KC and Primary Hepatocytes
4.3. Isolation of Primary HSC
4.4. Quantitative Real-Time PCR
4.5. Determination of Cytokine Levels
4.6. Western Blot Analysis
4.7. ATG7f/f-Albumin-Cre+ Mice
4.8. Immunofluorescence Analysis
4.9. siRNA Targeting Nrf2
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seki, E.; Schwabe, R.F. Hepatic inflammation and fibrosis: Functional links and key pathways. Hepatology 2015, 61, 1066–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouwens, L.; Baekeland, M.; de Zanger, R.; Wisse, E. Quantitation, tissue distribution and proliferation kinetics of kupffer cells in normal rat liver. Hepatology 1986, 6, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Benkdane, M.; Teixeira-Clerc, F.; Bonnafous, S.; Louvet, A.; Lafdil, F.; Pecker, F.; Tran, A.; Gual, P.; Mallat, A.; et al. M2 Kupffer cells promote M1 Kupffer cell apoptosis: A protective mechanism against alcoholic and nonalcoholic fatty liver disease. Hepatology 2014, 59, 130–142. [Google Scholar] [CrossRef]
- Seki, E.; Tsutsui, H.; Nakano, H.; Tsuji, N.M.; Hoshino, K.; Adachi, O.; Adachi, K.; Futatsugi, S.; Kuida, K.; Takeuchi, O.; et al. Lipopolysaccharide-induced IL-18 secretion from murine kupffer cells independently of myeloid differentiation factor 88 That is critically involved in induction of production of IL-12 and IL-1β. J. Immunol. 2001, 166, 2651–2657. [Google Scholar] [CrossRef] [Green Version]
- Zeng, T.; Zhang, C.-L.; Xiao, M.; Yang, R.; Xie, K.-Q. Critical Roles of Kupffer Cells in the Pathogenesis of Alcoholic Liver Disease: From Basic Science to Clinical Trials. Front. Immunol. 2016, 7, 538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Bansal, M.B. Role of Kupffer Cells in Driving Hepatic Inflammation and Fibrosis in HIV Infection. Front. Immunol. 2020, 11, 1086. [Google Scholar] [CrossRef] [PubMed]
- Kolios, G.; Valatas, V.; Kouroumalis, E. Role of Kupffer cells in the pathogenesis of liver disease. World J. Gastroenterol. 2006, 12, 7413–7420. [Google Scholar] [CrossRef]
- Canbay, A.; Feldstein, A.E.; Higuchi, H.; Werneburg, N.; Grambihler, A.; Bronk, S.F.; Gores, G.J. Kupffer cell engulfment of apoptotic bodies stimulates death ligand and cytokine expression. Hepatology 2003, 38, 1188–1198. [Google Scholar] [CrossRef]
- Liu, C.; Tao, Q.; Sun, M.; Wu, J.Z.; Yang, W.; Jian, P.; Peng, J.; Hu, Y.; Liu, C.; Liu, P. Kupffer cells are associated with apoptosis, inflammation and fibrotic effects in hepatic fibrosis in rats. Lab. Investig. 2010, 90, 1805–1816. [Google Scholar] [CrossRef] [Green Version]
- Rachfal, A. Connective tissue growth factor (CTGF/CCN2) in hepatic fibrosis. Hepatol. Res. 2003, 26, 1–9. [Google Scholar] [CrossRef]
- Frazier, K.; Williams, S.; Kothapalli, D.; Klapper, H.; Grotendorst, G.R. Stimulation of Fibroblast Cell Growth, Matrix Production, and Granulation Tissue Formation by Connective Tissue Growth Factor. J. Investig. Dermatol. 1996, 107, 404–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leivonen, S.-K.; Häkkinen, L.; Liu, D.; Kähäri, V.-M. Smad3 and Extracellular Signal-Regulated Kinase 1/2 Coordinately Mediate Transforming Growth Factor-β-Induced Expression of Connective Tissue Growth Factor in Human Fibroblasts. J. Investig. Dermatol. 2005, 124, 1162–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohrborn, D.; Wronkowitz, N.; Eckel, J. DPP4 in Diabetes. Front. Immunol. 2015, 6, 386. [Google Scholar] [CrossRef] [Green Version]
- Lamers, D.; Famulla, S.; Wronkowitz, N.; Hartwig, S.; Lehr, S.; Ouwens, D.M.; Eckardt, K.; Kaufman, J.M.; Ryden, M.; Müller, S.; et al. Dipeptidyl peptidase 4 is a novel adipokine potentially linking obesity to the metabolic syndrome. Diabetes 2011, 60, 1917–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirakawa, H.; Zempo, H.; Ogawa, M.; Watanabe, R.; Suzuki, J.-I.; Akazawa, H.; Komuro, I.; Isobe, M. A DPP-4 inhibitor suppresses fibrosis and inflammation on experimental autoimmune myocarditis in mice. PLoS ONE 2015, 10, e0119360. [Google Scholar] [CrossRef]
- Shinjo, T.; Nakatsu, Y.; Iwashita, M.; Sano, T.; Sakoda, H.; Ishihara, H.; Kushiyama, A.; Fujishiro, M.; Fukushima, T.; Tsuchiya, Y.; et al. DPP-IV inhibitor anagliptin exerts anti-inflammatory effects on macrophages, adipocytes, and mouse livers by suppressing NF-κB activation. Am. J. Physiol. Metab. 2015, 309, E214–E223. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, J.; Sugiyama, S.; Akiyama, E.; Iwashita, S.; Kurokawa, H.; Ohba, K.; Maeda, H.; Fujisue, K.; Yamamoto, E.; Kaikita, K.; et al. Dipeptidyl peptidase-4 inhibitor, sitagliptin, improves endothelial dysfunction in association with its anti-inflammatory effects in patients with coronary artery disease and uncontrolled diabetes. Circ. J. 2013, 77, 1337–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Tada, Y.; Gladson, S.; Nishimura, R.; Shimomura, I.; Karasawa, S.; Tatsumi, K.; West, J. Vildagliptin ameliorates pulmonary fibrosis in lipopolysaccharide-induced lung injury by inhibiting endothelial-to-mesenchymal transition. Respir. Res. 2017, 18, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumeier, C.; Schlüter, L.; Saussenthaler, S.; Laeger, T.; Rödiger, M.; Alaze, S.A.; Fritsche, L.; Häring, H.-U.; Stefan, N.; Fritsche, A.; et al. Elevated hepatic DPP4 activity promotes insulin resistance and non-alcoholic fatty liver disease. Mol. Metab. 2017, 6, 1254–1263. [Google Scholar] [CrossRef]
- Kawakubo, M.; Tanaka, M.; Ochi, K.; Watanabe, A.; Saka-Tanaka, M.; Kanamori, Y.; Yoshioka, N.; Yamashita, S.; Goto, M.; Itoh, M.; et al. Dipeptidyl peptidase-4 inhibition prevents nonalcoholic steatohepatitis-associated liver fibrosis and tumor development in mice independently of its anti-diabetic effects. Sci. Rep. 2020, 10, 983. [Google Scholar] [CrossRef] [Green Version]
- Klein, T.; Fujii, M.; Sandel, J.; Shibazaki, Y.; Wakamatsu, K.; Mark, M.; Yoneyama, H. Linagliptin alleviates hepatic steatosis and inflammation in a mouse model of non-alcoholic steatohepatitis. Med. Mol. Morphol. 2014, 47, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Kaji, K.; Yoshiji, H.; Ikenaka, Y.; Noguchi, R.; Aihara, Y.; Douhara, A.; Moriya, K.; Kawaratani, H.; Shirai, Y.; Yoshii, J.; et al. Dipeptidyl peptidase-4 inhibitor attenuates hepatic fibrosis via suppression of activated hepatic stellate cell in rats. J. Gastroenterol. 2014, 49, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Khalil, R.; Shata, A.; El-Kader, E.M.A.; Sharaf, H.; Abdo, W.S.; Amin, N.A.; Saber, S. Vildagliptin, a DPP-4 inhibitor, attenuates carbon tetrachloride-induced liver fibrosis by targeting ERK1/2, p38α, and NF-κB signaling. Toxicol. Appl. Pharmacol. 2020, 407, 115246. [Google Scholar] [CrossRef]
- Park, J.; Park, S.W.; Yoon, K.H.; Kim, S.R.; Ahn, K.J.; Lee, J.H.; Mok, J.O.; Chung, C.H.; Han, K.A.; Koh, G.P.; et al. Efficacy and safety of evogliptin monotherapy in patients with type 2 diabetes and moderately elevated glycated haemoglobin levels after diet and exercise. Diabetes, Obes. Metab. 2017, 19, 1681–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, E.; Huh, J.H.; Lee, E.Y.; Bae, J.C.; Chun, S.W.; Yu, S.H.; Kwak, S.H.; Park, K.S.; Lee, B. Efficacy and safety of evogliptin in patients with type 2 diabetes and non-alcoholic fatty liver disease: A multicentre, double-blind, randomized, comparative trial. Diabetes Obes. Metab. 2022, 24, 752–756. [Google Scholar] [CrossRef]
- Sun, L.; Xiu, M.; Wang, S.; Brigstock, D.R.; Li, H.; Qu, L.; Gao, R. Lipopolysaccharide enhances TGF -β1 signalling pathway and rat pancreatic fibrosis. J. Cell. Mol. Med. 2018, 22, 2346–2356. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Liu, C.; Zhou, D.; Zhang, L. TGF-β/SMAD Pathway and Its Regulation in Hepatic Fibrosis. J. Histochem. Cytochem. 2016, 64, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Zhou, J.; Song, S.; Kong, W.; Xia, W.; Chen, L.; Zeng, T. Dipeptidyl-Peptidase 4 Inhibitor Sitagliptin Ameliorates Hepatic Insulin Resistance by Modulating Inflammation and Autophagy in ob/ob Mice. Int. J. Endocrinol. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, M.; Kurokawa, H.; Waguri, S.; Taguchi, K.; Kobayashi, A.; Ichimura, Y.; Sou, Y.-S.; Ueno, I.; Sakamoto, A.; Tong, K.I.; et al. The selective autophagy substrate p62 activates the stress responsive transcription factor Nrf2 through inactivation of Keap1. Nat. Cell Biol. 2010, 12, 213–223. [Google Scholar] [CrossRef]
- Guo, K.; Jin, F. Dipeptidyl Peptidase-4 (DPP-4) Inhibitor Saxagliptin Alleviates Lipopolysaccharide-Induced Acute Lung Injury via Regulating the Nrf-2/HO-1 and NF-κB Pathways. J. Investig. Surg. 2021, 34, 695–702. [Google Scholar] [CrossRef]
- Si, J.; Meng, R.; Gao, P.; Hui, F.; Li, Y.; Liu, X.; Yang, B. Linagliptin protects rat carotid artery from balloon injury and activates the NRF2 antioxidant pathway. Exp. Anim. 2019, 68, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, C.R. Pro- and Anti-fibrogenic Functions of Gram-Negative Bacterial Lipopolysaccharide in the Liver. Front. Med. 2020, 7, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, Y.; Brenner, D.A. Liver inflammation and fibrosis. J. Clin. Investig. 2017, 127, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Kim, E.-Y.; Kim, J.-E.; Oh, S.; Park, S.-O.; Kim, S.-M.; Choi, H.; Song, J.-K.; Chang, E.-J. Evogliptin Suppresses Calcific Aortic Valve Disease by Attenuating Inflammation, Fibrosis, and Calcification. Cells 2021, 10, 57. [Google Scholar] [CrossRef]
- Nguyen, P.A.; Won, J.S.; Rahman, K.; Bae, E.J.; Cho, M.K. Modulation of Sirt1/NF-κB interaction of evogliptin is attributed to inhibition of vascular inflammatory response leading to attenuation of atherosclerotic plaque formation. Biochem. Pharmacol. 2019, 168, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Ba, Y.-D.; Sun, J.-H.; Zhao, X.-X. Evogliptin attenuates bleomycin-induced lung fibrosis via inhibiting TGF-β/Smad signaling in fibroblast. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10790–10798. [Google Scholar]
- Kim, M.-J.; Kim, N.-Y.; Jung, Y.-A.; Lee, S.; Jung, G.-S.; Kim, J.-G.; Lee, I.-K.; Lee, S.; Choi, Y.-K.; Park, K.-G. Evogliptin, a Dipeptidyl Peptidase-4 Inhibitor, Attenuates Renal Fibrosis Caused by Unilateral Ureteral Obstruction in Mice. Diabetes Metab. J. 2020, 44, 186–192. [Google Scholar] [CrossRef]
- Zhong, J.; Rajagopalan, S. Dipeptidyl Peptidase-4 Regulation of SDF-1/CXCR4 Axis: Implications for Cardiovascular Disease. Front. Immunol. 2015, 6, 477. [Google Scholar] [CrossRef] [Green Version]
- Diesen, D.L.; Kuo, P.C. Nitric Oxide and Redox Regulation in the Liver: Part II. Redox Biology in Pathologic Hepatocytes and Implications for Intervention. J. Surg. Res. 2011, 167, 96–112. [Google Scholar] [CrossRef] [Green Version]
- Dooley, S.; ten Dijke, P. TGF-β in progression of liver disease. Cell Tissue Res. 2012, 347, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wang, Y.; McNutt, M.A.; Zhu, W.-G. Autophagy process is associated with anti-neoplastic function. Acta Biochim. Biophys. Sin. 2011, 43, 425–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, M.; Fang, X.; Wang, X. Autophagy and inflammation. Clin. Transl. Med. 2017, 6, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zhang, C.; Ma, H.; Huang, Z.; Li, J.; Lou, J.; Li, B.; Tu, Q.; Gao, W. Detrimental Effect of Sitagliptin Induced Autophagy on Multiterritory Perforator Flap Survival. Front. Pharmacol. 2020, 11, 951. [Google Scholar] [CrossRef] [PubMed]
- Allaire, M.; Rautou, P.-E.; Codogno, P.; Lotersztajn, S. Autophagy in liver diseases: Time for translation? J. Hepatol. 2019, 70, 985–998. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.-Y.; Lee, S.-H.; Lee, J.-H.; Na Kang, Y.; Hwang, J.S.; Park, K.-G.; Kim, M.K.; Jang, B.K. Src Inhibition Attenuates Liver Fibrosis by Preventing Hepatic Stellate Cell Activation and Decreasing Connective Tissue Growth Factor. Cells 2020, 9, 558. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Jeong, Y.T.; Oh, H.; Kim, S.H.; Cho, J.M.; Kim, Y.-N.; Kim, S.S.; Kim, D.H.; Hur, K.Y.; Kim, H.K.; et al. Autophagy deficiency leads to protection from obesity and insulin resistance by inducing Fgf21 as a mitokine. Nat. Med. 2013, 19, 83–92. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, H.-Y.; Lee, S.-H.; Han, E.; Hwang, J.S.; Han, S.; Kim, M.K.; Jang, B.K. Evogliptin Directly Inhibits Inflammatory and Fibrotic Signaling in Isolated Liver Cells. Int. J. Mol. Sci. 2022, 23, 11636. https://doi.org/10.3390/ijms231911636
Seo H-Y, Lee S-H, Han E, Hwang JS, Han S, Kim MK, Jang BK. Evogliptin Directly Inhibits Inflammatory and Fibrotic Signaling in Isolated Liver Cells. International Journal of Molecular Sciences. 2022; 23(19):11636. https://doi.org/10.3390/ijms231911636
Chicago/Turabian StyleSeo, Hye-Young, So-Hee Lee, Eugene Han, Jae Seok Hwang, Sol Han, Mi Kyung Kim, and Byoung Kuk Jang. 2022. "Evogliptin Directly Inhibits Inflammatory and Fibrotic Signaling in Isolated Liver Cells" International Journal of Molecular Sciences 23, no. 19: 11636. https://doi.org/10.3390/ijms231911636