
Caffeine Inhibits NLRP3 Inflammasome Activation by Downregulating TLR4/MAPK/NF-κB Signaling Pathway in an Experimental NASH Model
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
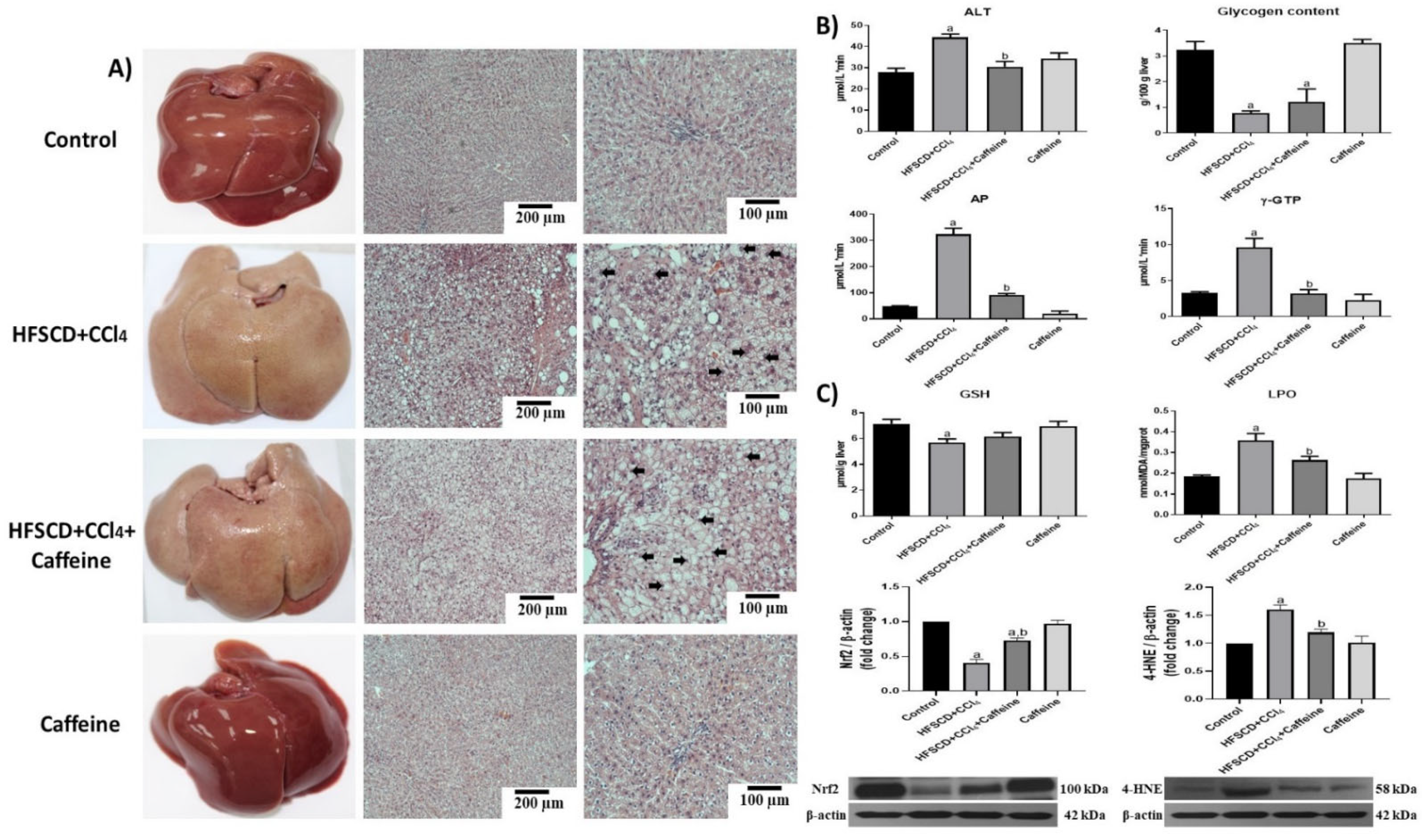
2.1. Caffeine Prevents Liver Damage and Oxidative Stress in the NASH Experimental Model
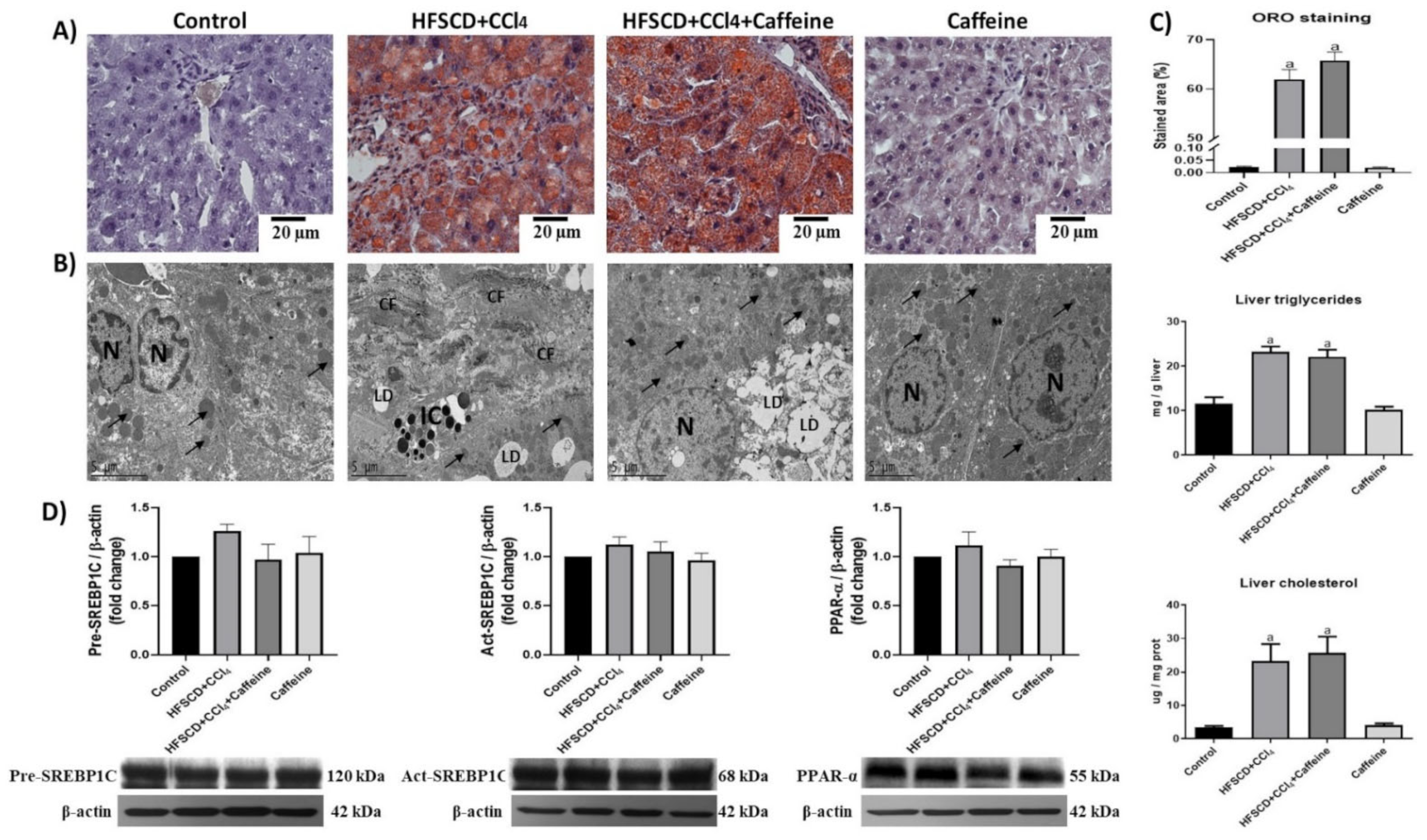
2.2. Effect of Caffeine on Liver Steatosis in the NASH Experimental Model
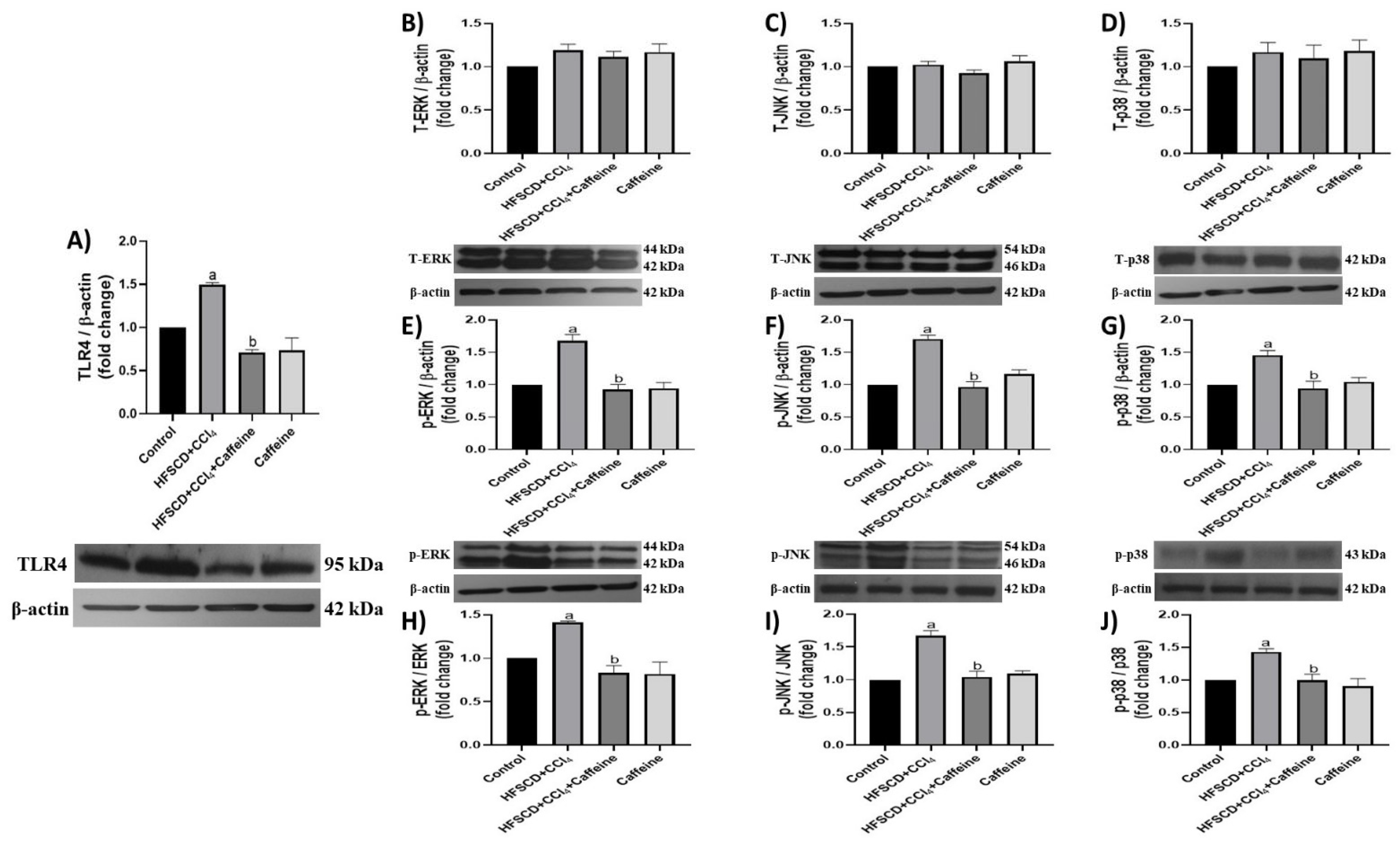
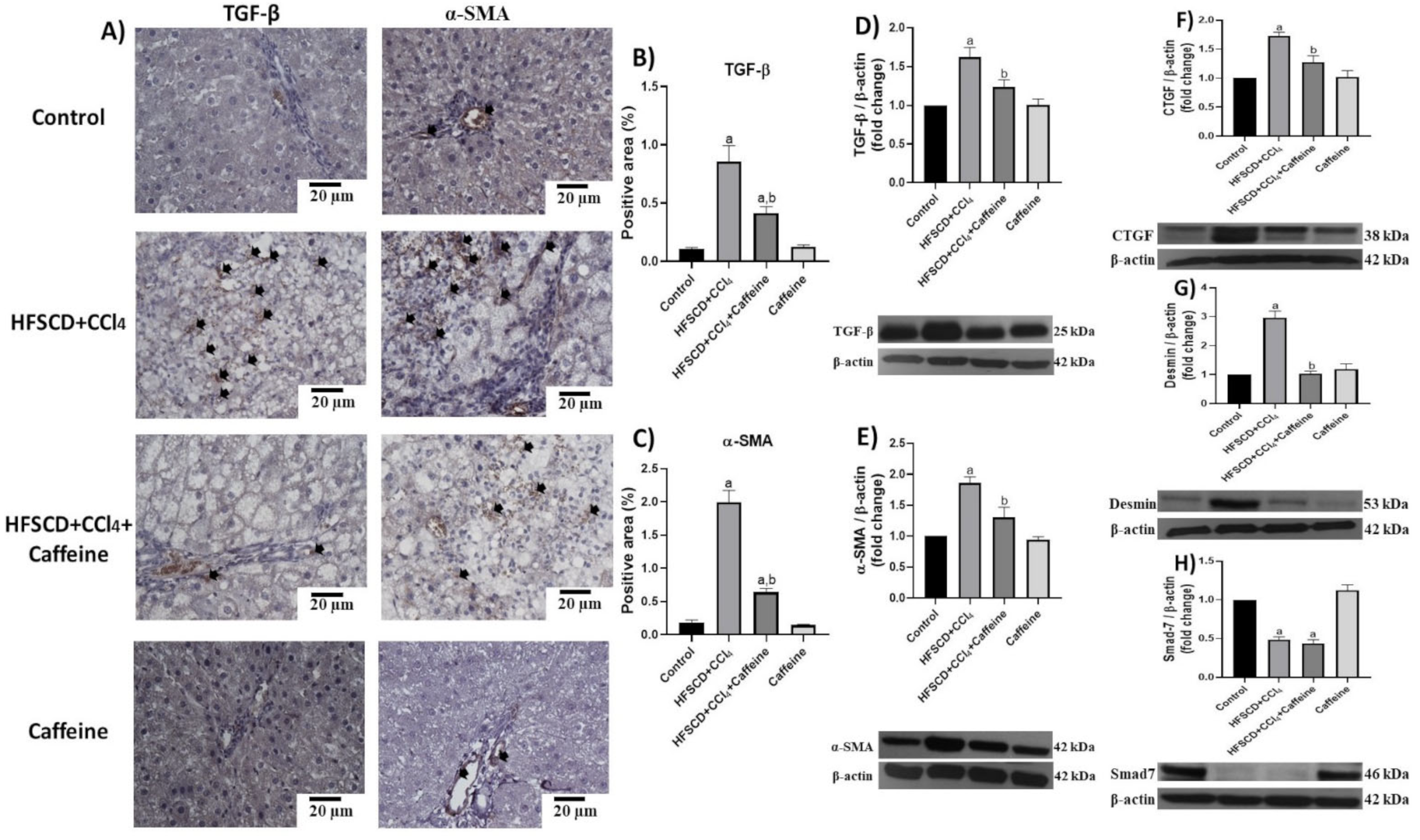
2.3. Caffeine Effect on the Proinflammatory and Fibrogenic TLR4-MAPK Pathway
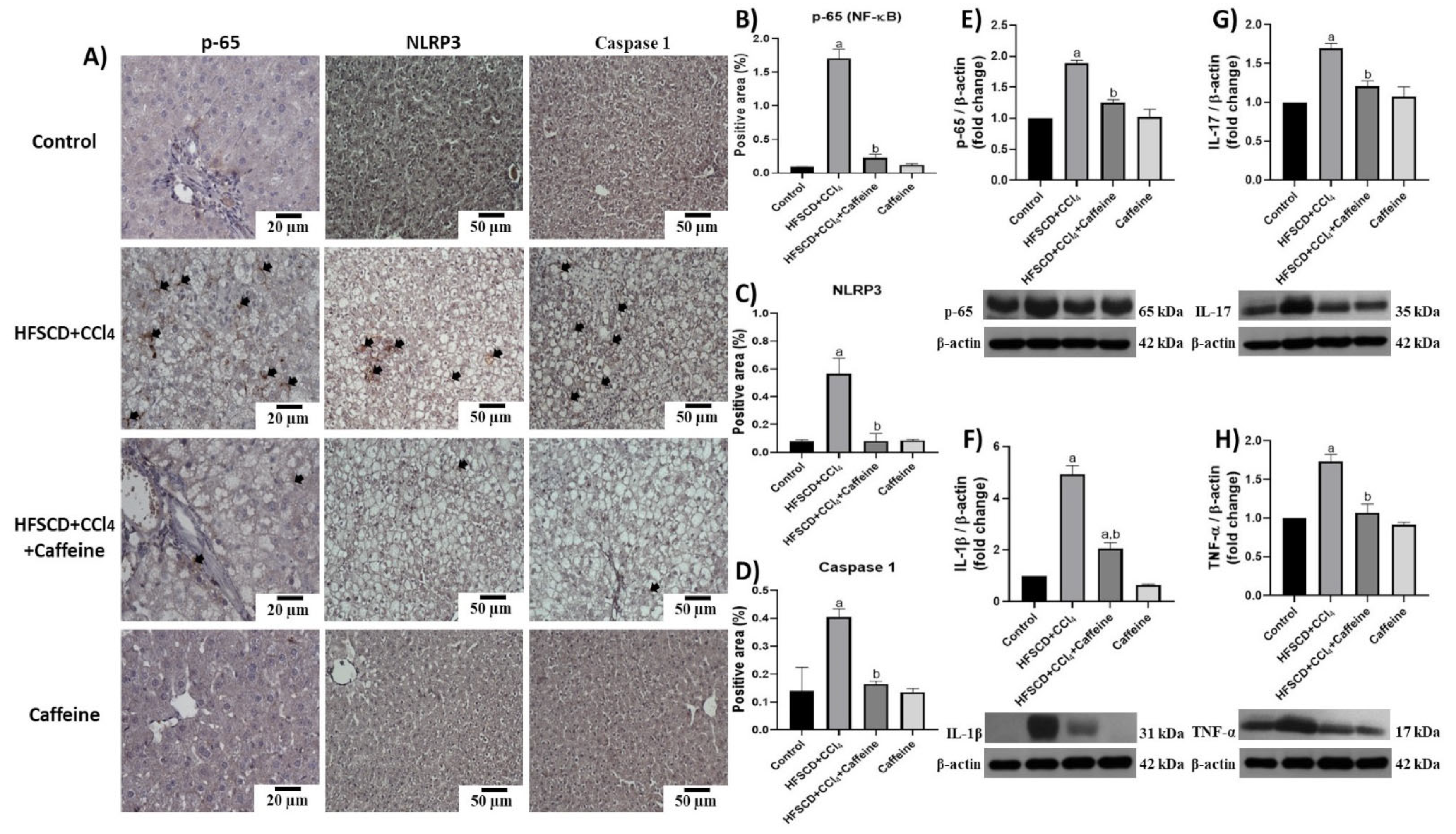
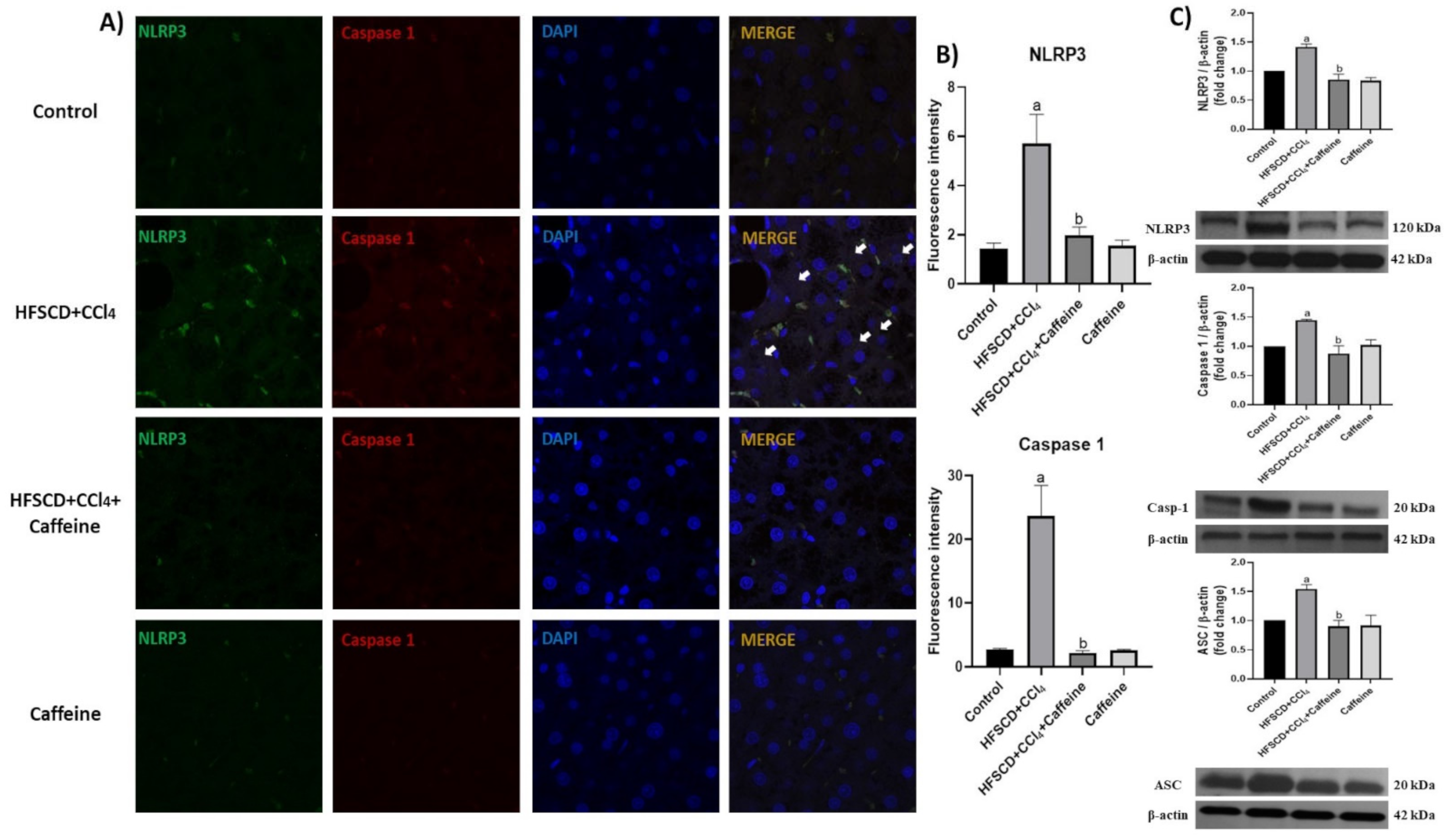
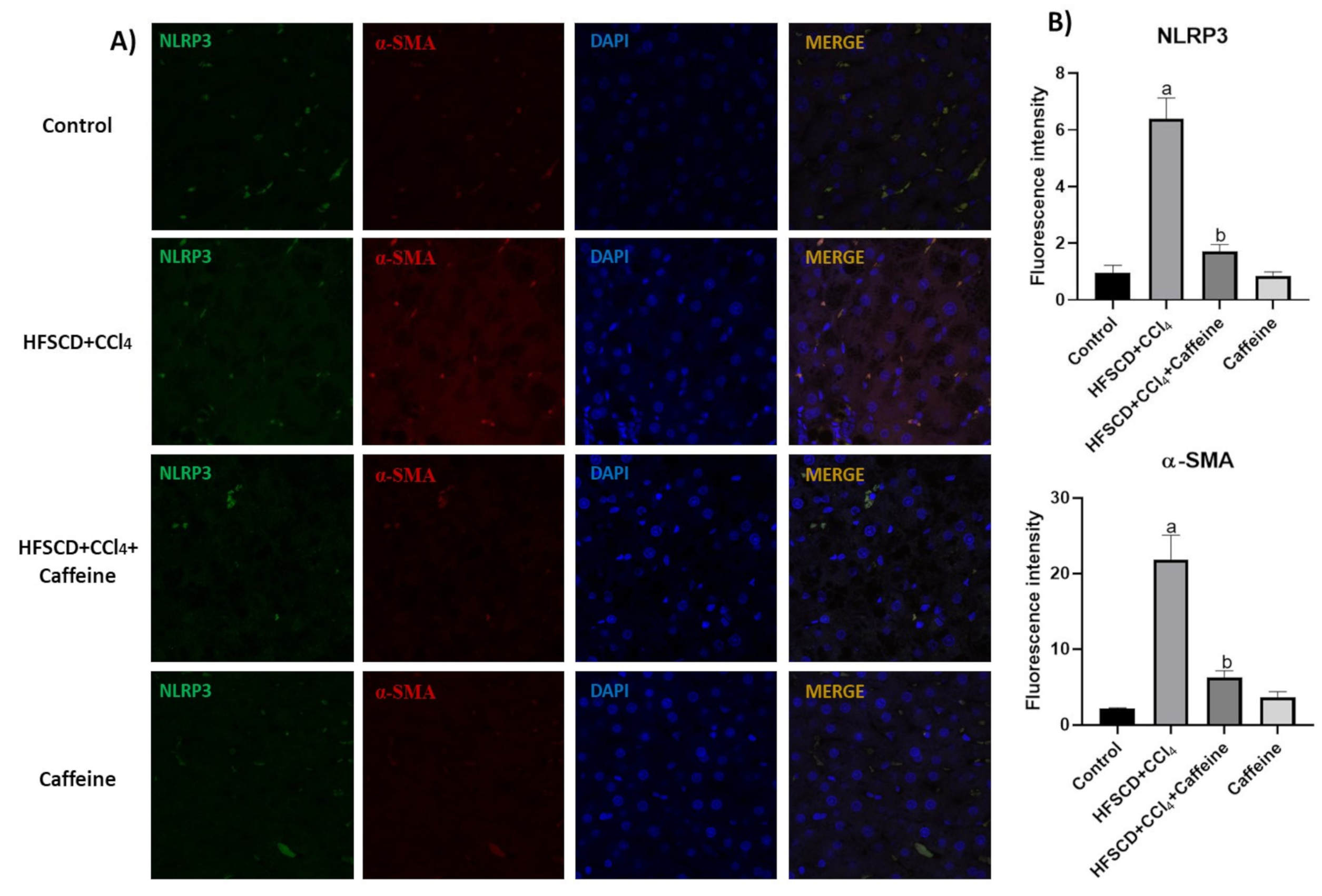
2.4. Caffeine Prevents NASH by Blocking the Activation of the NLRP3 Inflammasome Signaling Pathway
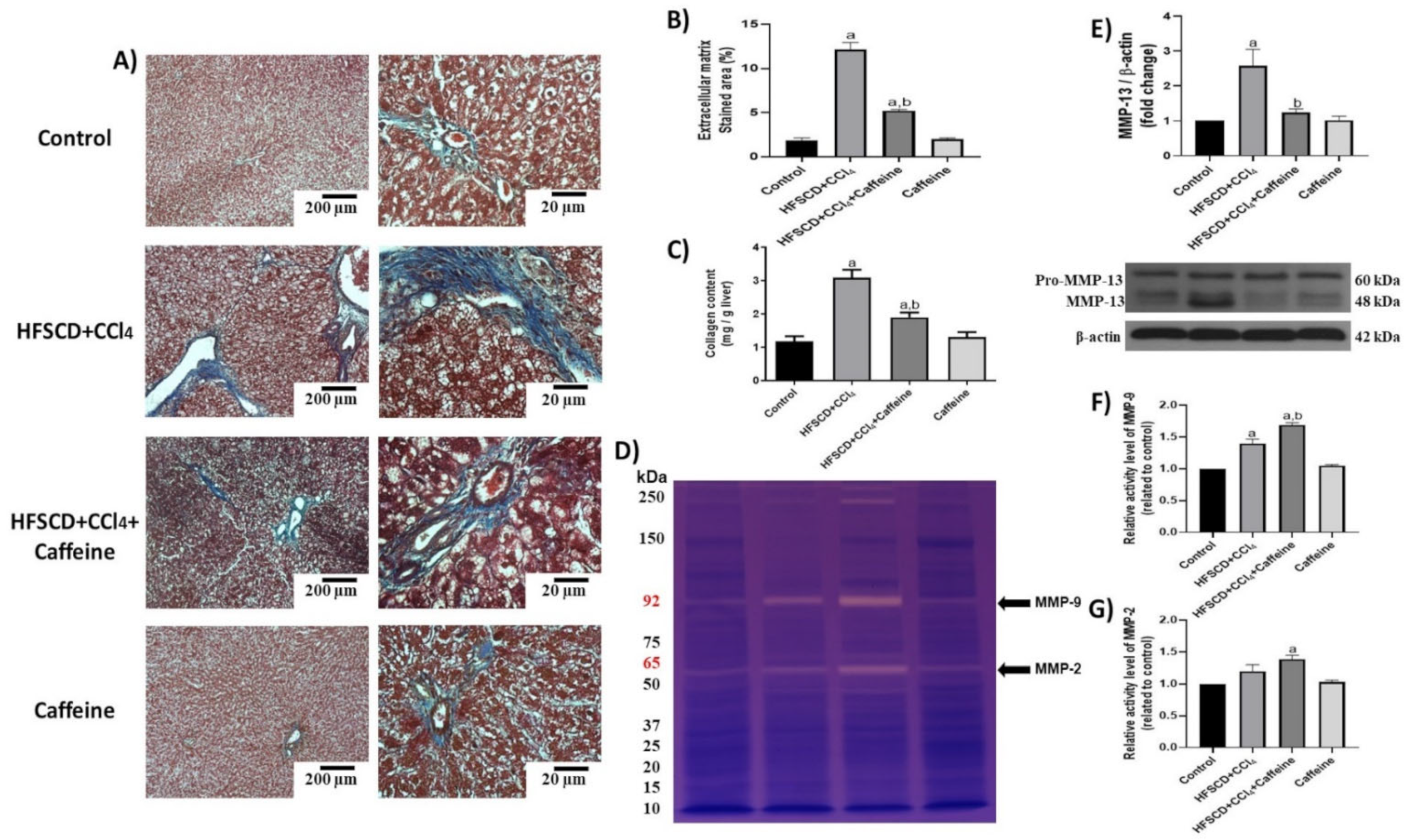
2.5. Caffeine Attenuates Fibrosis by Mitigating Hepatic Stellate Cell Activation
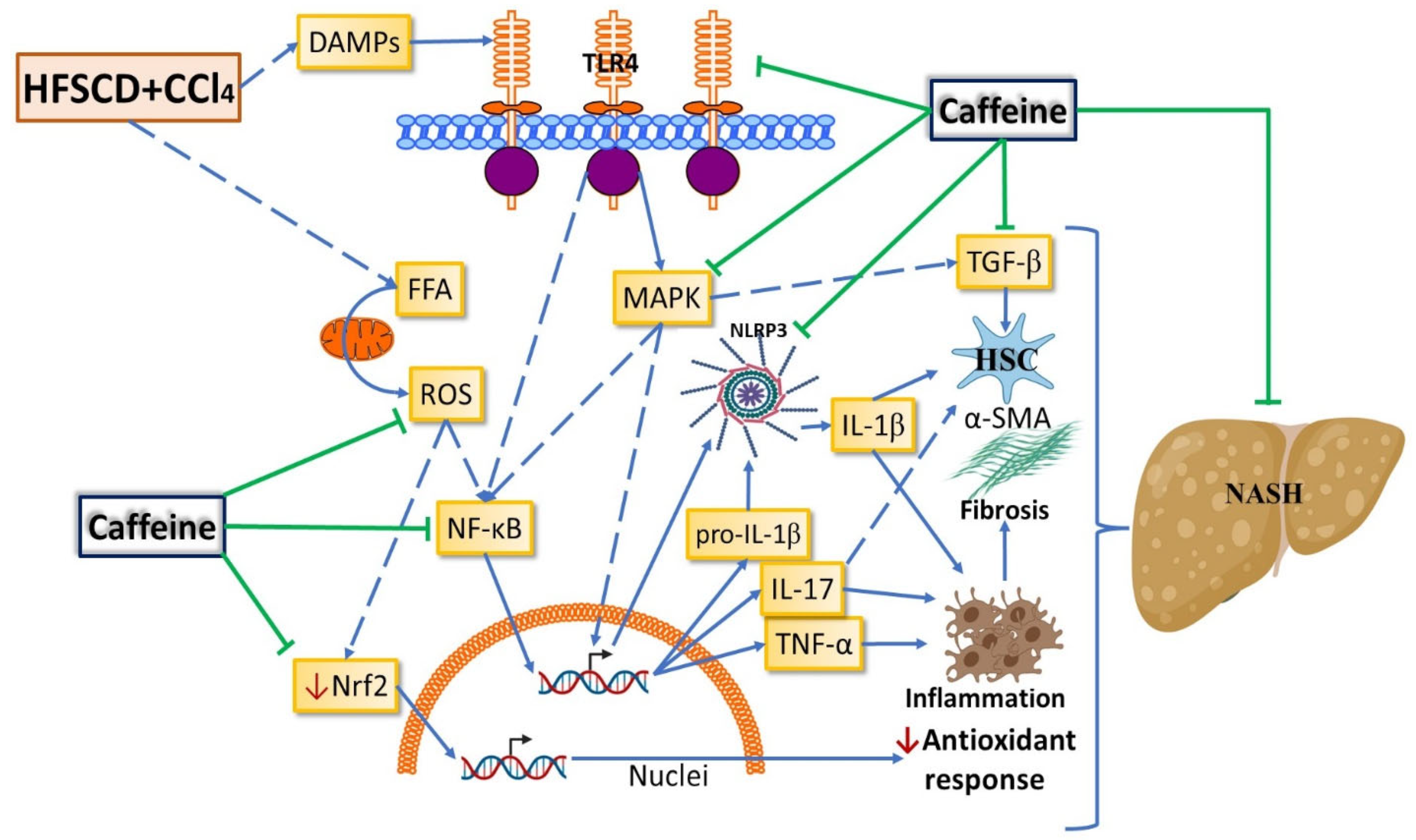
3. Discussion
4. Materials and Methods
4.1. Animal Treatments
Non-Alcoholic Steatohepatitis Induction and Caffeine Treatment
4.2. Reagents
4.3. Antibodies
4.4. Biochemical Analyses
4.5. Histology Determinations
4.6. Immunohistochemistry Assays
4.7. Transmission Electron Microscopy
4.8. Immunofluorescence
4.9. Western Blotting Analysis
4.10. Zymography
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muriel, P. (Ed.) The Liver: General Aspects and Epidemiology. In Liver Pathophysiology: Therapies and Antioxidants; Elsevier: Waltham, MA, USA, 2017; pp. 3–22. [Google Scholar] [CrossRef]
- Kucera, O.; Cervinkova, Z. Experimental models of non-alcoholic fatty liver disease in rats. World J. Gastroenterol. 2014, 20, 8364–8376. [Google Scholar] [CrossRef] [PubMed]
- Sheka, A.C.; Adeyi, O.; Thompson, J.; Hameed, B.; Crawford, P.A.; Ikramuddin, S. Nonalcoholic steatohepatitis: A review. JAMA 2020, 323, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, R.F.; Tabas, I.; Pajvani, U.B. Mechanisms of fibrosis development in nonalcoholic steatohepatitis. Gastroenterology 2020, 158, 1913–1928. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Fernández-Galilea, M.; Martínez-Fernández, L.; González-Muniesa, P.; Pérez-Chávez, A.; Martínez, J.A.; Moreno-Aliaga, M.J. Oxidative stress and non-alcoholic fatty liver disease: Effects of omega-3 fatty acid supplementation. Nutrients 2019, 11, 872. [Google Scholar] [CrossRef]
- Weiskirchen, R.; Tacke, F. Liver fibrosis: From pathogenesis to novel therapies. Dig. Dis. 2016, 34, 410–422. [Google Scholar] [CrossRef]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef]
- Mridha, A.R.; Wree, A.; Robertson, A.A.; Yeh, M.M.; Johnson, C.D.; Van Rooyen, D.M.; Haczeyni, F.; Teoh, N.C.-H.; Savard, C.; Ioannou, G.N.; et al. NLRP3 inflammasome blockade reduces liver inflammation and fibrosis in experimental NASH in mice. J. Hepatol. 2017, 66, 1037–1046. [Google Scholar] [CrossRef]
- Wu, X.; Dong, L.; Lin, X.; Li, J. Relevance of the NLRP3 inflammasome in the pathogenesis of chronic liver disease. Front. Immunol. 2017, 8, 1728. [Google Scholar] [CrossRef]
- Alegre, F.; Pelegrin, P.; Feldstein, A.E. Inflammasomes in liver fibrosis. Semin. Liver Dis. 2017, 37, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Vargas-Pozada, E.E.; Muriel, P. Herbal medicines for the liver: From bench to bedside. Eur. J. Gastroenterol. Hepatol. 2020, 32, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Tovar, E.; Muriel, P. Hepatoprotective Effect of Coffee. In Coffee Consumption and Health Implications; Farah, A., Ed.; Royal Society of Chemistry: London, UK, 2019; pp. 211–233. [Google Scholar]
- Sinha, R.A.; Farah, B.L.; Singh, B.K.; Siddique, M.M.; Li, Y.; Wu, Y.; Ilkayeva, O.R.; Gooding, J.; Ching, J.; Zhou, J.; et al. Caffeine stimulates hepatic lipid metabolism by the autophagy-lysosomal pathway in mice. Hepatology 2014, 59, 1366–1380. [Google Scholar] [CrossRef] [PubMed]
- Molloy, J.W.; Calcagno, C.J.; Williams, C.D.; Jones, F.J.; Torres, D.M.; Harrison, S.A. Association of coffee and caffeine consumption with fatty liver disease, nonalcoholic steatohepatitis, and degree of hepatic fibrosis. Hepatology 2012, 55, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Rodriguez, A.C.; Shiani, A.; Lipka, S.; Shahzad, G.; Kumar, A.; Mustacchia, P. Association between caffeine consumption and nonalcoholic fatty liver disease: A systemic review and meta-analysis. Ther. Adv. Gastroenterol. 2016, 9, 113–120. [Google Scholar] [CrossRef]
- Murosaki, S.; Lee, T.R.; Muroyama, K.; Shin, E.S.; Cho, S.Y.; Yamamoto, Y.; Lee, S.J. A combination of caffeine, arginine, soy isoflavones, and L-carnitine enhances both lipolysis and fatty acid oxidation in 3T3-L1 and HepG2 cells in vitro and in KK mice in vivo. J. Nutr. 2007, 137, 2252–2257. [Google Scholar] [CrossRef]
- Sugiura, C.; Nishimatsu, S.; Moriyama, T.; Ozasa, S.; Kawada, T.; Sayama, K. Catechins and caffeine inhibit fat accumulation in mice through the improvement of hepatic lipid metabolism. J. Obes. 2012, 2012, 520510. [Google Scholar] [CrossRef] [PubMed]
- Birerdinc, A.; Stepanova, M.; Pawloski, L.; Younossi, Z.M. Caffeine is protective in patients with non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2012, 35, 76–82. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Bättig, K.; Holmén, J.; Nehlig, A.; Zvartau, E.E. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol. Rev. 1999, 51, 83–133. [Google Scholar]
- Tanda, G.; Goldberg, S.R. Alteration of the behavioral effects of nicotine by chronic caffeine exposure. Pharmacol. Biochem. Behav. 2000, 66, 47–64. [Google Scholar] [CrossRef]
- Gasior, M.; Jaszyna, M.; Munzar, P.; Witkin, J.M.; Goldberg, S.R. Caffeine potentiates the discriminative-stimulus effects of nicotine in rats. Psychopharmacology 2002, 162, 385–395. [Google Scholar] [CrossRef]
- Arauz, J.; Zarco, N.; Segovia, J.; Shibayama, M.; Tsutsumi, V.; Muriel, P. Caffeine prevents experimental liver fibrosis by blocking the expression of TGF-β. Eur. J. Gastroenterol. Hepatol. 2014, 26, 164–173. [Google Scholar] [CrossRef]
- Du, Y.; Melchert, H.U.; Knopf, H.; Braemer-Hauth, M.; Gerding, B.; Pabel, E. Association of serum caffeine concentrations with blood lipids in caffeine-drug users and nonusers—Results of German National Health Surveys from 1984 to 1999. Eur. J. Epidemiol. 2005, 20, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Marangon, A.F.C.; Helou, T.; Gonzalez, D.V. Effect of caffeine on lipid profile in ciclism practitioners. J. Int. Soc. Sports Nutr. 2012, 9 (Suppl. 1), P20. [Google Scholar] [CrossRef]
- González, S.; Salazar, N.; Ruiz-Saavedra, S.; Gómez-Martín, M.; de Los Reyes-Gavilán, C.G.; Gueimonde, M. Long-term coffee consumption is associated with fecal microbial composition in humans. Nutrients 2020, 12, 1287. [Google Scholar] [CrossRef] [PubMed]
- Gurwara, S.; Dai, A.; Ajami, N.; El-Serag, H.B.; Graham, D.Y.; Jiao, L. Caffeine consumption and the colonic mucosa-associated gut microbiota. Off. J. Am. Coll. Gastroenterol. 2019, 114, S119–S120. [Google Scholar] [CrossRef]
- van Dam, R.M.; Hu, F.B.; Willett, W.C. Coffee, caffeine, and health. N. Engl. J. Med. 2020, 383, 369–378. [Google Scholar] [CrossRef]
- Amer, M.G.; Mazen, N.F.; Mohamed, A.M. Caffeine intake decreases oxidative stress and inflammatory biomarkers in experimental liver diseases induced by thioacetamide: Biochemical and histological study. Int. J. Immunopathol. Pharmacol. 2017, 30, 13–24. [Google Scholar] [CrossRef]
- Gordillo-Bastidas, D.; Oceguera-Contreras, E.; Salazar-Montes, A.; González-Cuevas, J.; Hernández-Ortega, L.D.; Armendáriz-Borunda, J. Nrf2 and Snail-1 in the prevention of experimental liver fibrosis by caffeine. World J. Gastroenterol. 2013, 19, 9020–9033. [Google Scholar] [CrossRef]
- Cachón, A.U.; Quintal-Novelo, C.; Medina-Escobedo, G.; Castro-Aguilar, G.; Moo-Puc, R.E. Hepatoprotective effect of low doses of caffeine on CCl4-induced liver damage in rats. J. Diet. Suppl. 2017, 14, 158–172. [Google Scholar] [CrossRef]
- Ramos-Tovar, E.; Muriel, P. Molecular mechanisms that link oxidative stress, inflammation, and fibrosis in the liver. Antioxidants 2020, 9, 1279. [Google Scholar] [CrossRef]
- Arauz, J.; Zarco, N.; Hernández-Aquino, E.; Galicia-Moreno, M.; Favari, L.; Segovia, J.; Muriel, P. Coffee consumption prevents fibrosis in a rat model that mimics secondary biliary cirrhosis in humans. Nutr. Res. 2017, 40, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Rezaie, A.; Pashmforosh, M.; Karamallah, M.H.; Fazlara, A.; Haghighat, N.; Shahriari, A. Hepatoprotective effect of caffeine on diethylnitrosamine-induced liver injury in rats. Bulg. J. Vet. Med. 2014, 17, 183–190. [Google Scholar]
- Casas-Grajales, S.; Muriel, P. Antioxidants in liver health. World J. Gastrointest. Pharmacol. Ther. 2015, 6, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Azam, S.; Hadi, N.; Khan, N.U.; Hadi, S.M. Antioxidant and prooxidant properties of caffeine, theobromine and xanthine. Med. Sci. Monit. 2003, 9, BR325–BR330. [Google Scholar]
- Szabo, G.; Petrasek, J. Inflammasome activation and function in liver disease. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 387–400. [Google Scholar] [CrossRef]
- Knorr, J.; Wree, A.; Tacke, F.; Feldstein, A.E. The NLRP3 inflammasome in alcoholic and nonalcoholic steatohepatitis. Semin. Liver Dis. 2020, 40, 298–306. [Google Scholar] [CrossRef]
- Horn, P.; Newsome, P.N. Emerging therapeutic targets for NASH: Key innovations at the preclinical level. Expert Opin. Ther. Targets 2020, 24, 175–186. [Google Scholar] [CrossRef]
- Zhao, W.; Ma, L.; Cai, C.; Gong, X. Caffeine inhibits NLRP3 inflammasome activation by suppressing MAPK/NF-κB and A2aR signaling in LPS-induced THP-1 macrophages. Int. J. Biol. Sci. 2019, 15, 1571–1581. [Google Scholar] [CrossRef]
- Kovács, E.; Alatshan, A.; Budai, M.; Czimmerer, Z.; Bíró, E.; Benkő, S. Caffeine has different immunomodulatory effect on the cytokine expression and NLRP3 inflammasome function in various human macrophage subpopulations. Nutrients 2021, 13, 2409. [Google Scholar] [CrossRef]
- Chen, S.; Wu, Q.; Zhong, D.; Li, C.; Du, L. Caffeine prevents hyperoxia-induced lung injury in neonatal mice through NLRP3 inflammasome and NF-κB pathway. Respir. Res. 2020, 21, 140. [Google Scholar] [CrossRef]
- Wang, H.-Q.; Song, K.-Y.; Feng, J.-Z.; Huang, S.-Y.; Guo, X.-M.; Zhang, L.; Zhang, G.; Huo, Y.-C.; Zhang, R.-R.; Ma, Y.; et al. Caffeine inhibits activation of the NLRP3 inflammasome via autophagy to attenuate microglia-mediated neuroinflammation in experimental autoimmune encephalomyelitis. J. Mol. Neurosci. 2022, 72, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yu, X.; Zhang, Y.; Liu, N.; Xue, X.; Fu, J. Caffeine treatment started before injury reduces hypoxic-ischemic white-matter damage in neonatal rats by regulating phenotypic microglia polarization. Pediatr. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Boaru, S.G.; Borkham-Kamphorst, E.; Tihaa, L.; Haas, U.; Weiskirchen, R. Expression analysis of inflammasomes in experimental models of inflammatory and fibrotic liver disease. J. Inflamm. 2012, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [PubMed]
- Mills, K.H.; Dungan, L.S.; Jones, S.A.; Harris, J. The role of inflammasome-derived IL-1 in driving IL-17 responses. J. Leukoc. Biol. 2013, 93, 489–497. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef]
- Yang, J.; Wise, L.; Fukuchi, K.I. TLR4 cross-talk with NLRP3 inflammasome and complement signaling pathways in Alzheimer’s disease. Front. Immunol. 2020, 11, 724. [Google Scholar] [CrossRef]
- Meng, Y.; Yu, C.-H.; Li, W.; Li, T.; Luo, W.; Huang, S.; Wu, P.-S.; Cai, S.-X.; Li, X. Angiotensin-converting enzyme 2/angiotensin-(1-7)/Mas axis protects against lung fibrosis by inhibiting the MAPK/NF-κB pathway. Am. J. Respir. Cell Mol. Biol. 2014, 50, 723–736. [Google Scholar] [CrossRef]
- Khan, S.; Choi, R.J.; Shehzad, O.; Kim, H.P.; Islam, N.; Choi, J.S.; Kim, Y.S. Molecular mechanism of capillarisin-mediated inhibition of MyD88/TIRAP inflammatory signaling in in vitro and in vivo experimental models. J. Ethnopharmacol. 2013, 145, 626–637. [Google Scholar] [CrossRef]
- Cheng, Y.-C.; Ding, Y.-M.; Hueng, D.-Y.; Chen, J.-Y.; Chen, Y. Caffeine suppresses the progression of human glioblastoma via cathepsin B and MAPK signaling pathway. J. Nutr. Biochem. 2016, 33, 63–72. [Google Scholar] [CrossRef]
- Wang, H.; Guan, W.; Yang, W.; Wang, Q.; Zhao, H.; Yang, F.; Lv, X.; Li, J. Caffeine inhibits the activation of hepatic stellate cells induced by acetaldehyde via adenosine A2A receptor mediated by the cAMP/PKA/SRC/ERK1/2/P38 MAPK signal pathway. PLoS ONE 2014, 9, e92482. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Pozada, E.E.; Ramos-Tovar, E.; Acero-Hernández, C.; Cardoso-Lezama, I.; Galindo-Gómez, S.; Tsutsumi, V.; Muriel, P. Caffeine mitigates experimental nonalcoholic steatohepatitis and the progression of thioacetamide-induced liver fibrosis by blocking the MAPK and TGF-β/Smad3 signaling pathways. Ann. Hepatol. 2022, 27, 100671. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Chisaka, T.; Kubomura, Y.; Yamahara, J.; Sawada, T.; Fujimura, H.; Kimura, H.; Hasegawa, T. Effects of crude drugs on experimental hypercholesterolemia. I. Tea and its active principles. J. Ethnopharmacol. 1986, 17, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Glossmann, H.; Neville, D.M. Gamma-Glutamyltransferase in kidney brush border membranes. FEBS Lett. 1972, 19, 340–344. [Google Scholar] [CrossRef]
- Bergmeyer, H.U.; Grabl, M.; Walter, H.E. Enzymes. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Grabl, M., Eds.; Verlag-Chemie: Weinheim, Germany, 1983; pp. 269–270. ISBN 3527260455. [Google Scholar]
- Seifter, S.; Dayton, S. The estimation of glycogen with the anthrone reagent. Arch. Biochem. 1950, 25, 191–200. [Google Scholar]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Chen, T.-C.; Lee, R.A.; Tsai, S.L.; Kanamaluru, D.; Gray, N.E.; Yiv, N.; Cheang, R.T.; Tan, J.H.; Lee, J.Y.; Fitch, M.D.; et al. An ANGPTL4-ceramide-protein kinase Cζ axis mediates chronic glucocorticoid exposure-induced hepatic steatosis and hypertriglyceridemia in mice. J. Biol. Chem. 2019, 294, 9213–9224. [Google Scholar] [CrossRef]
- Domínguez-Pérez, M.; Simoni-Nieves, A.; Rosales, P.; Nuño-Lámbarri, N.; Rosas-Lemus, M.; Souza, V.; Miranda, R.U.; Bucio, L.; Carvajal, S.U.; Marquardt, J.U.; et al. Cholesterol burden in the liver induces mitochondrial dynamic changes and resistance to apoptosis. J. Cell. Physiol. 2019, 234, 7213–7223. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Prockop, D.J.; Udenfriend, S. A specific method for the analysis of hydroxyproline in tissues and urine. Anal. Biochem. 1960, 1, 228–239. [Google Scholar] [CrossRef]
- Ramos-Tovar, E.; Hernández-Aquino, E.; Casas-Grajales, S.; Buendia-Montaño, L.D.; Galindo-Gómez, S.; Camacho, J.; Tsutsumi, V.; Muriel, P. Stevia prevents acute and chronic liver injury induced by carbon tetrachloride by blocking oxidative stress through Nrf2 Upregulation. Oxid. Med. Cell. Longev. 2018, 2018, 3823426. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Aquino, E.; Quezada-Ramírez, M.A.; Silva-Olivares, A.; Casas-Grajales, S.; Ramos-Tovar, E.; Flores-Beltrán, R.E.; Segovia, J.; Shibayama, M.; Muriel, P. Naringenin attenuates the progression of liver fibrosis via inactivation of hepatic stellate cells and profibrogenic pathways. Eur. J. Pharmacol. 2019, 865, 172730. [Google Scholar] [CrossRef]
- Rodriguez-Callejas, J.D.; Fuchs, E.; Perez-Cruz, C. Increased oxidative stress, hyperphosphorylation of tau, and dystrophic microglia in the hippocampus of aged Tupaia belangeri. Glia 2020, 68, 1775–1793. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Brand (Location) | Catalog | WB Dilution | IHC Dilution | IF Dilution |
|---|---|---|---|---|---|
| MMP-13 | Merck-Millipore® (Burlington, MA, USA) | MAB13426 | 1:500 | ||
| TGF-β | Merck-Millipore® (Burlington, MA, USA) | MAB1032 | 1:500 | 1:250 | |
| NF-κB (p65) | Merck Millipore® (Burlington, MA, USA) | MAB3026 | 1:500 | 1:250 | |
| JNK | Cell Signaling Technology® (Danvers, MA, USA) | 9252S | 1:500 | ||
| pp38 | Cell Signaling Technology® (Danvers, MA, USA) | 9211S | 1:500 | ||
| p38 | Abcam® (Cambridge, UK) | AB31828 | 1:500 | ||
| Smad7 | Abcam® (Cambridge, UK) | AB90086 | 1:500 | ||
| pJNK | Abcam® (Cambridge, UK) | AB131499 | 1:500 | ||
| 4-HNE | Abcam® (Cambridge, UK) | AB46545 | 1:500 | ||
| IL-1β | Abcam® (Cambridge, UK) | AB18329 | 1:500 | ||
| Nrf2 | Abcam® (Cambridge, UK) | AB31163 | 1:500 | ||
| SREBP1C | Abcam® (Cambridge, UK) | AB28481 | 1:500 | ||
| α-SMA | Sigma-Aldrich® (St. Louis, MI, USA) | A-5691 | 1:500 | 1:250 | 1:250 |
| Desmin | Sigma-Aldrich® (St. Louis, MI, USA) | 243M-1 | 1:500 | ||
| pERK | Santa Cruz Biotechnology® (Santa Cruz, CA, USA) | SC-136521 | 1:500 | ||
| ERK | Santa Cruz Biotechnology® (Santa Cruz, CA, USA) | SC-292838 | 1:500 | ||
| CTGF | Santa Cruz Biotechnology® (Santa Cruz, CA, USA) | SC-365970 | 1:500 | ||
| IL-17 | Santa Cruz Biotechnology® (Dallas, TX, USA) | SC-374218 | 1:500 | ||
| TNF-α | Santa Cruz Biotechnology® (Dallas, TX, USA) | SC-52746 | 1:500 | ||
| ASC | Santa Cruz Biotechnology® (Dallas, TX, USA) | SC-514414 | 1:500 | ||
| Caspase 1 | Santa Cruz Biotechnology® (Dallas, TX, USA) | SC-392736 | 1:500 | 1:250 | 1:250 |
| PPAR-α | Santa Cruz Biotechnology® (Dallas, TX, USA) | SC-398394 | 1:500 | ||
| NLRP3 | Novus biologicals® (Littleton, CO, USA) | NBP2-12446 | 1:500 | 1:250 | 1:250 |
| TLR4 | Thermo Fisher® (Waltham, MA, USA) | 48-2300 | 1:500 | ||
| β-actin | Thermo Fisher® (Waltham, MA, USA) | AM4302 | 1:500 | ||
| Secondary Antibodies | |||||
| Anti-Rabbit | Merck Millipore® (Burlington, MA, USA) | A0545 | 1:3000 | 1:1000 | |
| Anti-Mouse | Thermo Fisher® (Waltham, MA, USA) | 62-6520 | 1:5000 | 1:1000 | |
| Alexa fluor 594 | Thermo Fisher® (Waltham, MA, USA) | Z25007 | 1:1000 | ||
| Alexa fluor 488 | Thermo Fisher® (Waltham, MA, USA) | Z25302 | 1:1000 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargas-Pozada, E.E.; Ramos-Tovar, E.; Rodriguez-Callejas, J.D.; Cardoso-Lezama, I.; Galindo-Gómez, S.; Talamás-Lara, D.; Vásquez-Garzón, V.R.; Arellanes-Robledo, J.; Tsutsumi, V.; Villa-Treviño, S.; et al. Caffeine Inhibits NLRP3 Inflammasome Activation by Downregulating TLR4/MAPK/NF-κB Signaling Pathway in an Experimental NASH Model. Int. J. Mol. Sci. 2022, 23, 9954. https://doi.org/10.3390/ijms23179954
Vargas-Pozada EE, Ramos-Tovar E, Rodriguez-Callejas JD, Cardoso-Lezama I, Galindo-Gómez S, Talamás-Lara D, Vásquez-Garzón VR, Arellanes-Robledo J, Tsutsumi V, Villa-Treviño S, et al. Caffeine Inhibits NLRP3 Inflammasome Activation by Downregulating TLR4/MAPK/NF-κB Signaling Pathway in an Experimental NASH Model. International Journal of Molecular Sciences. 2022; 23(17):9954. https://doi.org/10.3390/ijms23179954
Chicago/Turabian StyleVargas-Pozada, Eduardo E., Erika Ramos-Tovar, Juan D. Rodriguez-Callejas, Irina Cardoso-Lezama, Silvia Galindo-Gómez, Daniel Talamás-Lara, Verónica Rocío Vásquez-Garzón, Jaime Arellanes-Robledo, Víctor Tsutsumi, Saúl Villa-Treviño, and et al. 2022. "Caffeine Inhibits NLRP3 Inflammasome Activation by Downregulating TLR4/MAPK/NF-κB Signaling Pathway in an Experimental NASH Model" International Journal of Molecular Sciences 23, no. 17: 9954. https://doi.org/10.3390/ijms23179954