
Zinc Deficiency Induces Autophagy in HT-22 Mouse Hippocampal Neuronal Cell Line

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
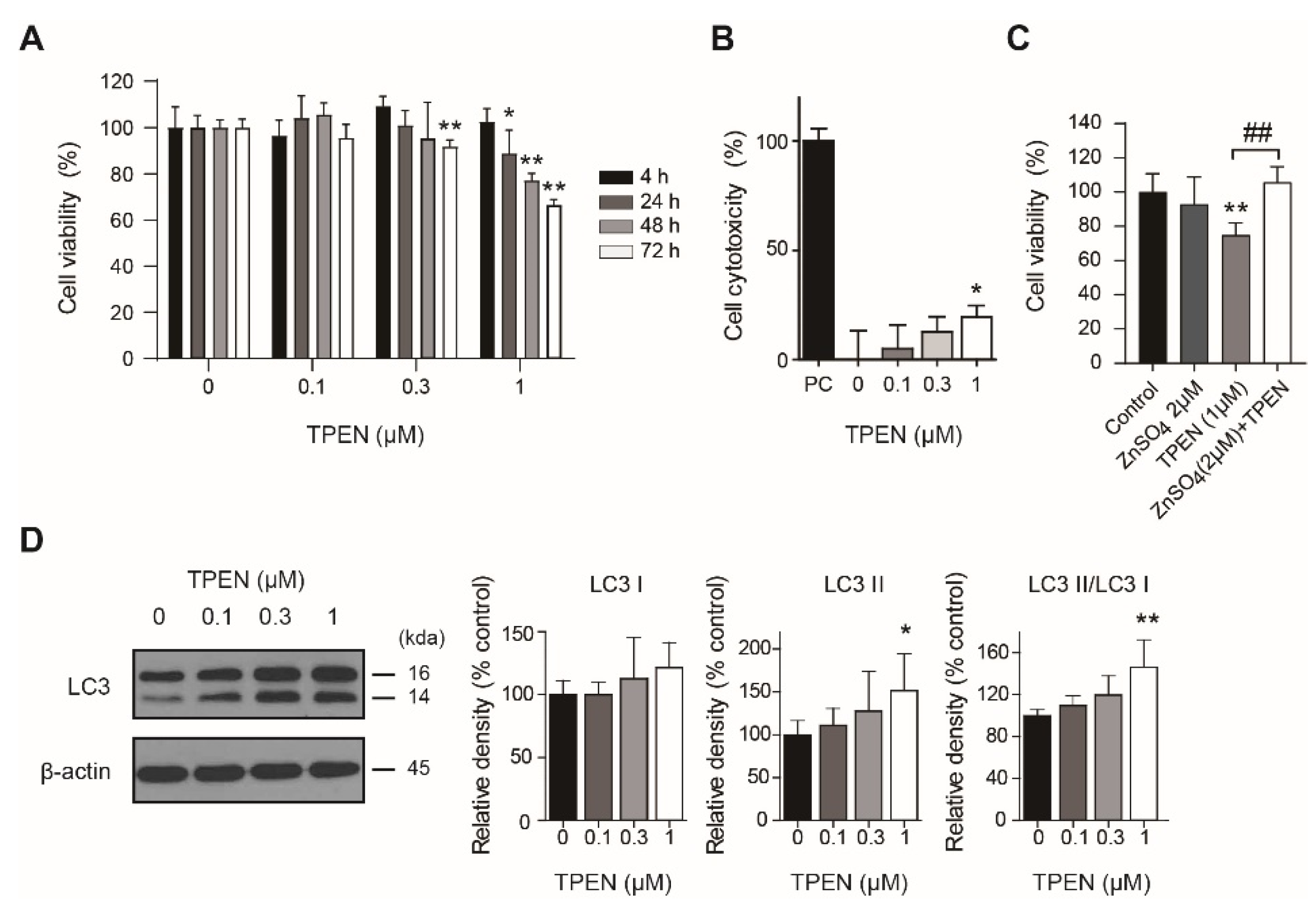
2.1. Zinc Deficiency by TPEN Treatment Decreased HT-22 Cell Viability and Induced Autophagy
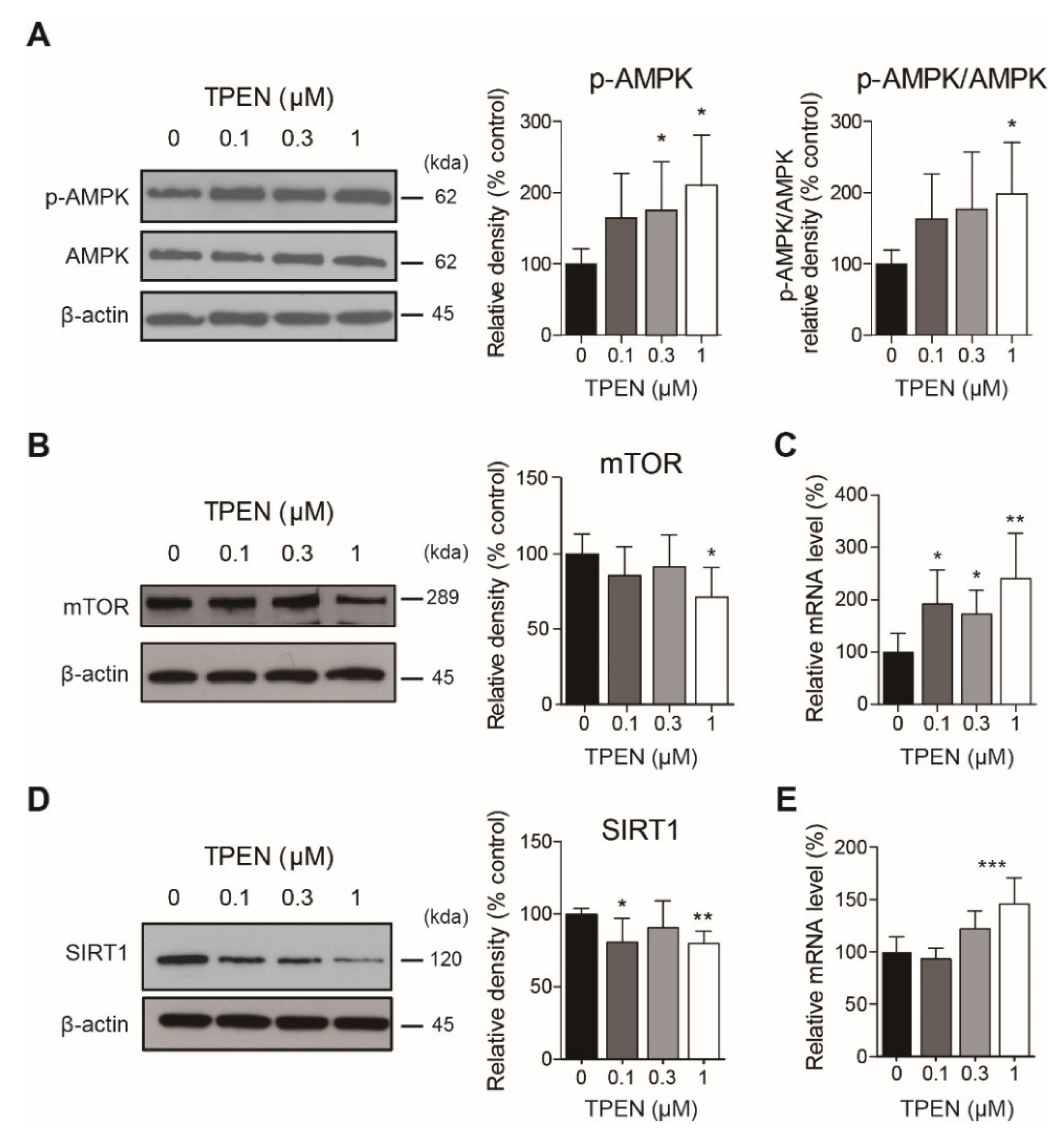
2.2. Zinc Deficiency by TPEN Treatment Changed Autophagy-Related Signaling
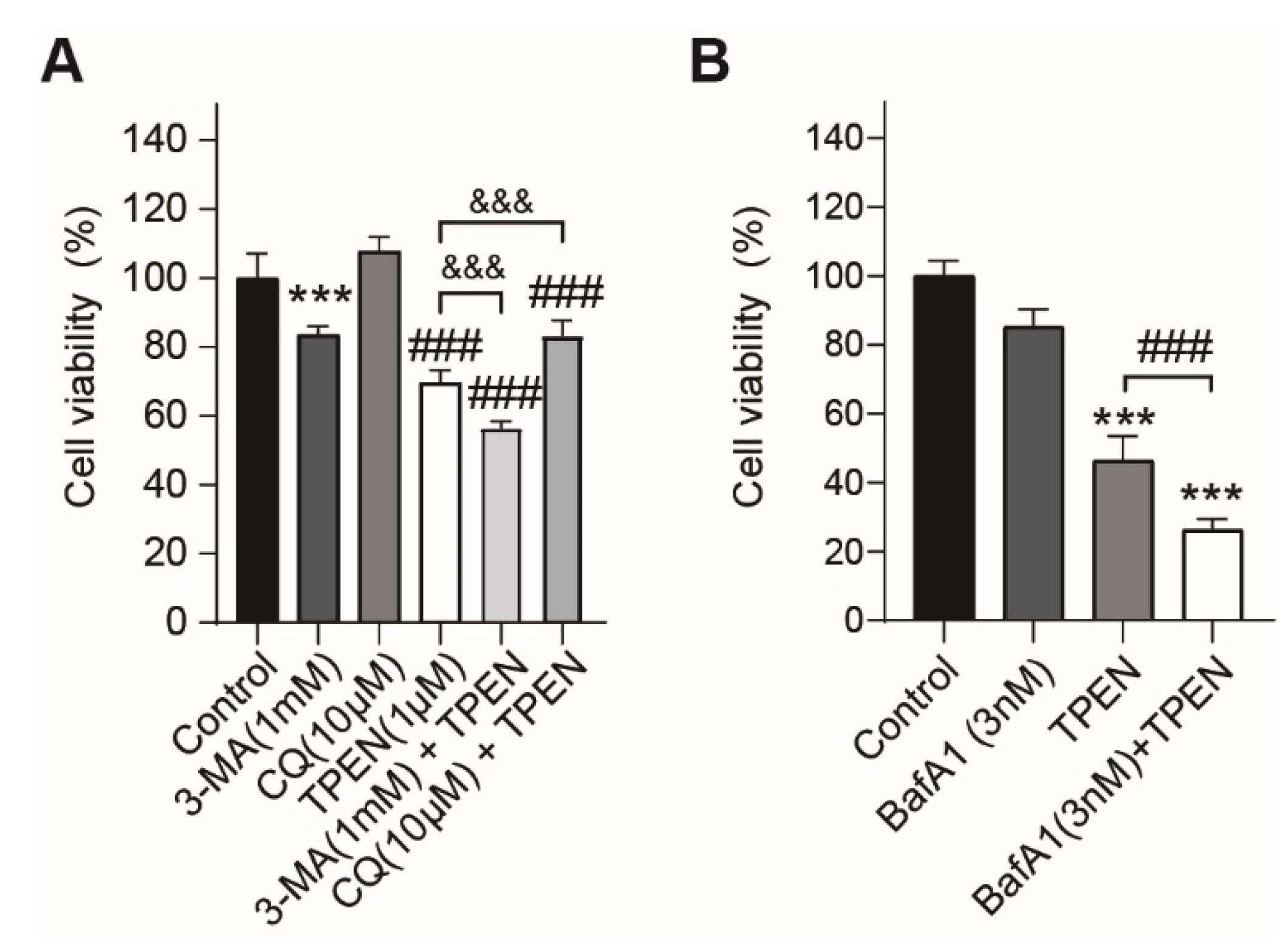
2.3. Inhibition of TPEN-Induced Autophagy by Chemical Inhibitors (3-MA, Chloroquine, Bafilomycin A1) Altered TPEN-Induced Cytotoxicity in HT-22 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Drug Treatments
4.3. Cell Viability Assay
4.4. Lactate Dehydrogenase (LDH) Assay
4.5. Quantitative Real-Time PCR (qRT-PCR) Analysis
4.6. Western Blot Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, H.Y.; Gale, J.R.; Reynolds, I.J.; Weiss, J.H.; Aizenman, E. The Multifaceted Roles of Zinc in Neuronal Mitochondrial Dysfunction. Biomedicines 2021, 9, 489. [Google Scholar] [CrossRef]
- Pérez-Clausell, J.; Danscher, G. Intravesicular Localization of Zinc in Rat Telencephalic Boutons. A Histochemical Study. Brain Res. 1985, 337, 91–98. [Google Scholar] [CrossRef]
- Wuehler, S.E.; Peerson, J.M.; Brown, K.H. Use of National Food Balance Data to Estimate the Adequacy of Zinc in National Food Supplies: Methodology and Regional Estimates. Public Health Nutr. 2005, 8, 812–819. [Google Scholar] [CrossRef]
- Warthon-Medina, M.; Moran, V.H.; Stammers, A.L.; Dillon, S.; Qualter, P.; Nissensohn, M.; Serra-Majem, L.; Lowe, N.M. Zinc Intake, Status and Indices of Cognitive Function in Adults and Children: A Systematic Review and Meta-Analysis. Eur. J. Clin. Nutr. 2015, 69, 649–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massaro, T.F.; Mohs, M.; Fosmire, G. Effects of Moderate Zinc Deficiency on Cognitive Performance in Young Adult Rats. Physiol. Behav. 1982, 29, 117–121. [Google Scholar] [CrossRef]
- Tahmasebi Boroujeni, S.; Naghdi, N.; Shahbazi, M.; Farrokhi, A.; Bagherzadeh, F.; Kazemnejad, A.; Javadian, M. The Effect of Severe Zinc Deficiency and Zinc Supplement on Spatial Learning and Memory. Biol. Trace Elem. Res. 2009, 130, 48–61. [Google Scholar] [CrossRef]
- Halas, E.S.; Eberhardt, M.J.; Diers, M.A.; Sandstead, H.H. Learning and Memory Impairment in Adult Rats Due to Severe Zinc Deficiency during Lactation. Physiol. Behav. 1983, 30, 371–381. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of Metabolism and Mitochondrial Homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.L. AMPK and MTOR Regulate Autophagy through Direct Phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Guan, K.L. MTOR: A Pharmacologic Target for Autophagy Regulation. J. Clin. Investig. 2015, 125, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamata, T.; Horie, T.; Matsunami, M.; Sasaki, M.; Ohsumi, Y. Zinc Starvation Induces Autophagy in Yeast. J. Biol. Chem. 2017, 292, 8520–8530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Li, Y.; Li, C.M. Autophagy Plays a Positive Role in Zinc-Induced Apoptosis in Intestinal Porcine Epithelial Cells. Toxicol. Vitr. 2017, 44, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Lee, S.J.; Byun, H.; Kim, Y.; Oh, Y.J.; Koh, J.Y.; Hwang, J.J. Clioquinol Induces Autophagy in Cultured Astrocytes and Neurons by Acting as a Zinc Ionophore. Neurobiol. Dis. 2011, 42, 242–251. [Google Scholar] [CrossRef]
- Lee, S.J.; Cho, K.S.; Koh, J.Y. Oxidative Injury Triggers Autophagy in Astrocytes: The Role of Endogenous Zinc. Glia 2009, 57, 1351–1361. [Google Scholar] [CrossRef]
- Liuzzi, J.P.; Yoo, C. Role of Zinc in the Regulation of Autophagy during Ethanol Exposure in Human Hepatoma Cells. Biol. Trace Elem. Res. 2013, 156, 350–356. [Google Scholar] [CrossRef]
- Bucci, M.D.; Weisenhorn, E.; Haws, S.; Yao, Z.; Zimmerman, G.; Gannon, M.; Taggart, J.; Lee, T.; Klionsky, D.J.; Russell, J.; et al. An Autophagy-Independent Role for ATG41 in Sulfur Metabolism during Zinc Deficiency. Genetics 2018, 208, 1115–1130. [Google Scholar] [CrossRef] [Green Version]
- Liuzzi, J.P.; Pazos, R. Interplay Between Autophagy and Zinc. J. Trace Elem. Med. Biol. 2020, 62, 126636. [Google Scholar] [CrossRef]
- Nixon, R.A. The Role of Autophagy in Neurodegenerative Disease. Nat. Med. 2013, 19, 983–997. [Google Scholar] [CrossRef]
- Jung, S.; Jeong, H.; Yu, S.W. Autophagy as a Decisive Process for Cell Death. Exp. Mol. Med. 2020, 52, 921–930. [Google Scholar] [CrossRef]
- Ullio, C.; Brunk, U.T.; Urani, C.; Melchioretto, P.; Bonelli, G.; Baccino, F.M.; Autelli, R. Autophagy of Metallothioneins Prevents TNF-Induced Oxidative Stress and Toxicity in Hepatoma Cells. Autophagy 2015, 11, 2184–2198. [Google Scholar] [CrossRef] [Green Version]
- Choudhuri, S.; McKim, J.M.; Klaassen, C.D. Role of Hepatic Lysosomes in the Degradation of Metallothionein. Toxicol. Appl. Pharmacol. 1992, 115, 64–71. [Google Scholar] [CrossRef]
- Aydemir, T.B.; Liuzzsi, J.P.; McClellan, S.; Cousins, R.J. Zinc Transporter ZIP8 (SLC39A8) and Zinc Influence IFN-γ Expression in Activated Human T Cells. J. Leukoc. Biol. 2009, 86, 337–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiselyov, K.; Colletti, G.A.; Terwilliger, A.; Ketchum, K.; Lyons, C.W.P.; Quinn, J.; Muallem, S. TRPML: Transporters of Metals in Lysosomes Essential for Cell Survival? Cell Calcium 2011, 50, 288–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinozaki, D.; Merkulova, E.A.; Naya, L.; Horie, T.; Kanno, Y.; Seo, M.; Ohsumi, Y.; Masclaux-Daubresse, C.; Yoshimoto, K. Autophagy Increases Zinc Bioavailability to Avoid Light-Mediated Reactive Oxygen Species Production under Zinc Deficiency. Plant Physiol. 2020, 182, 1284–1296. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Moyer, A.; Peng, B.; Wu, J.; Hannafon, B.N.; Ding, W.Q. Chloroquine Is a Zinc Ionophore. PLoS ONE 2014, 9, e109180. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, Y.C.; Fang, C.; Russell, R.C.; Kim, J.H.; Fan, W.; Liu, R.; Zhong, Q.; Guan, K.L. Differential Regulation of Distinct Vps34 Complexes by AMPK in Nutrient Stress and Autophagy. Cell 2013, 152, 290–303. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wang, W.; Sun, X.; Xu, D.; Wang, C.; Zhang, Q.; Wang, H.; Luo, W.; Chen, Y.; Chen, H.; et al. AMPK Regulates Autophagy by Phosphorylating BECN1 at Threonine 388. Autophagy 2016, 12, 1447–1459. [Google Scholar] [CrossRef] [Green Version]
- Jing, K.; Song, K.; Shin, S.; Kim, N.; Jeong, S.; Oh, H.-R.; Park, J.; Seo, K.; Heo, J.-Y.; Han, J.; et al. Docosahexaenoic Acid Induces Autophagy through P53/AMPK/MTOR Signaling and Promotes Apoptosis in Human Cancer Cells Harboring Wild-Type P53. Autophagy 2011, 7, 1348–1358. [Google Scholar] [CrossRef] [Green Version]
- Jinpiao, Z.; Zongze, Z.; Qiuyue, Y.; Peng, F.; Qi, Z.; Yanlin, W.; Chang, C. Metformin Attenuates Sevoflurane-Induced Neurocognitive Impairment through AMPK-ULK1-Dependent Autophagy in Aged Mice. Brain Res. Bull. 2020, 157, 18–25. [Google Scholar] [CrossRef]
- Follo, C.; Vidoni, C.; Morani, F.; Ferraresi, A.; Seca, C.; Isidoro, C. Amino Acid Response by Halofuginone in Cancer Cells Triggers Autophagy through Proteasome Degradation of MTOR. Cell Commun. Signal. 2019, 17, 39. [Google Scholar] [CrossRef] [Green Version]
- Cohen, H.Y.; Miller, C.; Bitterman, K.J.; Wall, N.R.; Hekking, B.; Kessler, B.; Howitz, K.T.; Gorospe, M.; De Cabo, R.; Sinclair, D.A. Calorie Restriction Promotes Mammalian Cell Survival by Inducing the SIRT1 Deacetylase. Science 2004, 305, 390–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, X.; Lee, M.R.; Huang, X.; Messina-Graham, S.; Broxmeyer, H.E. SIRT1 Positively Regulates Autophagy and Mitochondria Function in Embryonic Stem Cells under Oxidative Stress. Stem Cells 2014, 32, 1183–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Z.Q.; Zhang, X.; Zhen, Y.; He, X.Y.; Zhao, S.; Li, X.F.; Yang, B.; Gao, F.; Guo, F.Y.; Fu, L.; et al. A Novel Small-Molecule Activator of Sirtuin-1 Induces Autophagic Cell Death/Mitophagy as a Potential Therapeutic Strategy in Glioblastoma Article. Cell Death Dis. 2018, 9, 767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- In, H.L.; Cao, L.; Mostoslavsky, R.; Lombard, D.B.; Liu, J.; Bruns, N.E.; Tsokos, M.; Alt, F.W.; Finkel, T. A Role for the NAD-Dependent Deacetylase Sirt1 in the Regulation of Autophagy. Proc. Natl. Acad. Sci. USA 2008, 105, 3374–3379. [Google Scholar] [CrossRef] [Green Version]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Crosstalk between Oxidative Stress and SIRT1: Impact on the Aging Process. Int. J. Mol. Sci. 2013, 14, 3834–3859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Wang, L.; Fozouni, P.; Evjen, G.; Chandra, V.; Jiang, J.; Lu, C.; Nicastri, M.; Bretz, C.; Winkler, J.D.; et al. SIRT1 Is Downregulated by Autophagy in Senescence and Ageing. Nat. Cell Biol. 2020, 22, 1170–1179. [Google Scholar] [CrossRef]
- Arslan, P.; Di Virgilio, F.; Beltrame, M. Cytosolic Ca2+ Homeostasis in Ehrlich and Yoshida Carcinomas. A New, Membrane-Permeant Chelator of Heavy Metals Reveals That These Ascites Tumor Cell Lines Have Normal Cytosolic Free Ca2+. J. Biol. Chem. 1985, 260, 2719–2727. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-Y.; Lee, J.-H.; Kim, S.-A. Zinc Deficiency Induces Autophagy in HT-22 Mouse Hippocampal Neuronal Cell Line. Int. J. Mol. Sci. 2022, 23, 8811. https://doi.org/10.3390/ijms23158811
Kim S-Y, Lee J-H, Kim S-A. Zinc Deficiency Induces Autophagy in HT-22 Mouse Hippocampal Neuronal Cell Line. International Journal of Molecular Sciences. 2022; 23(15):8811. https://doi.org/10.3390/ijms23158811
Chicago/Turabian StyleKim, Si-Yeon, Jung-Ho Lee, and Soon-Ae Kim. 2022. "Zinc Deficiency Induces Autophagy in HT-22 Mouse Hippocampal Neuronal Cell Line" International Journal of Molecular Sciences 23, no. 15: 8811. https://doi.org/10.3390/ijms23158811