
Allethrin Promotes Apoptosis and Autophagy Associated with the Oxidative Stress-Related PI3K/AKT/mTOR Signaling Pathway in Developing Rat Ovaries

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
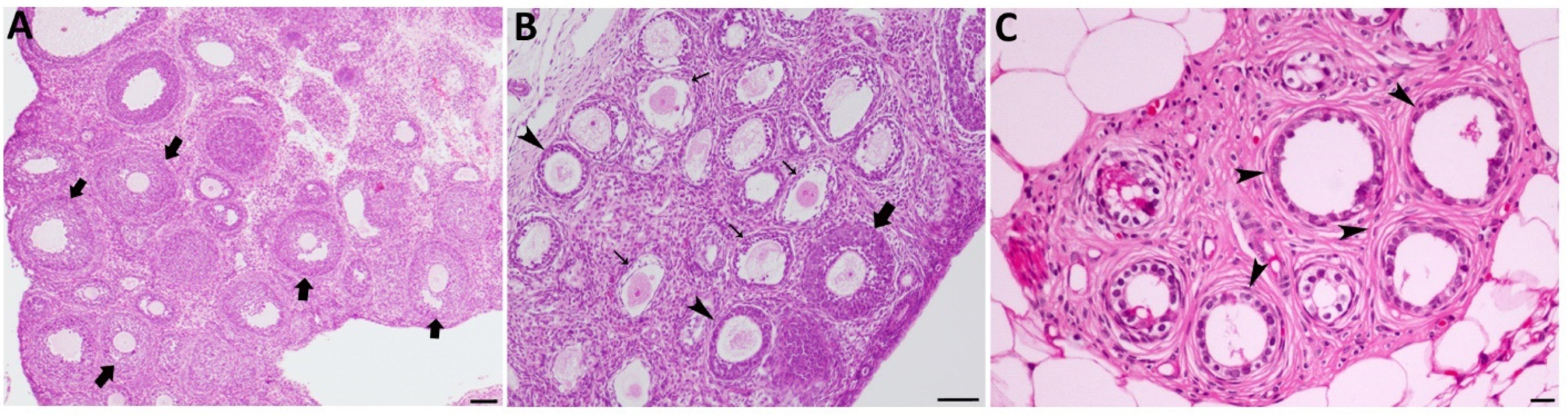
2.1. Allethrin Caused Histopathological Damage in Rat Ovaries
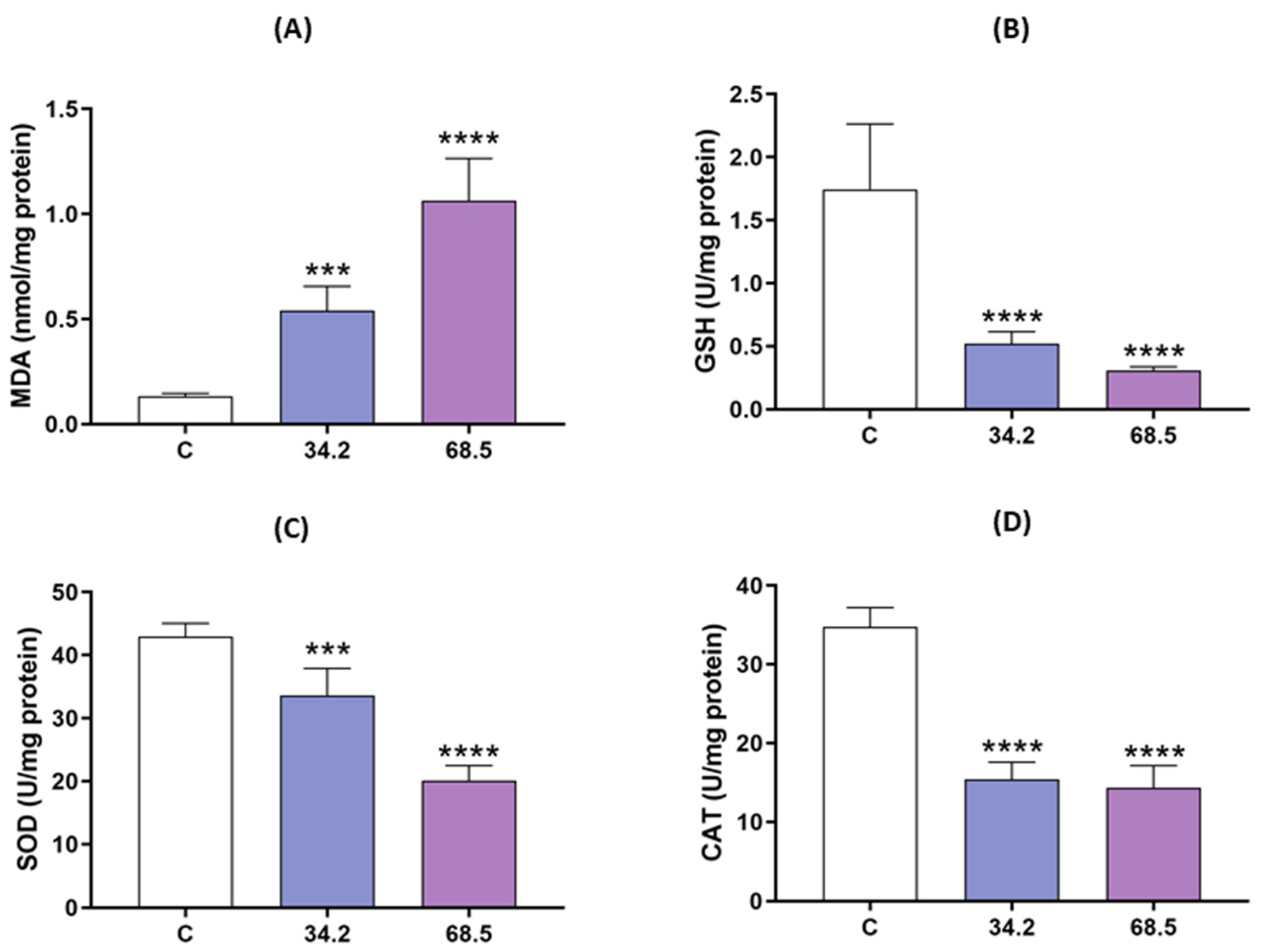
2.2. Allethrin Induced Oxidative Stress in Rat Ovaries
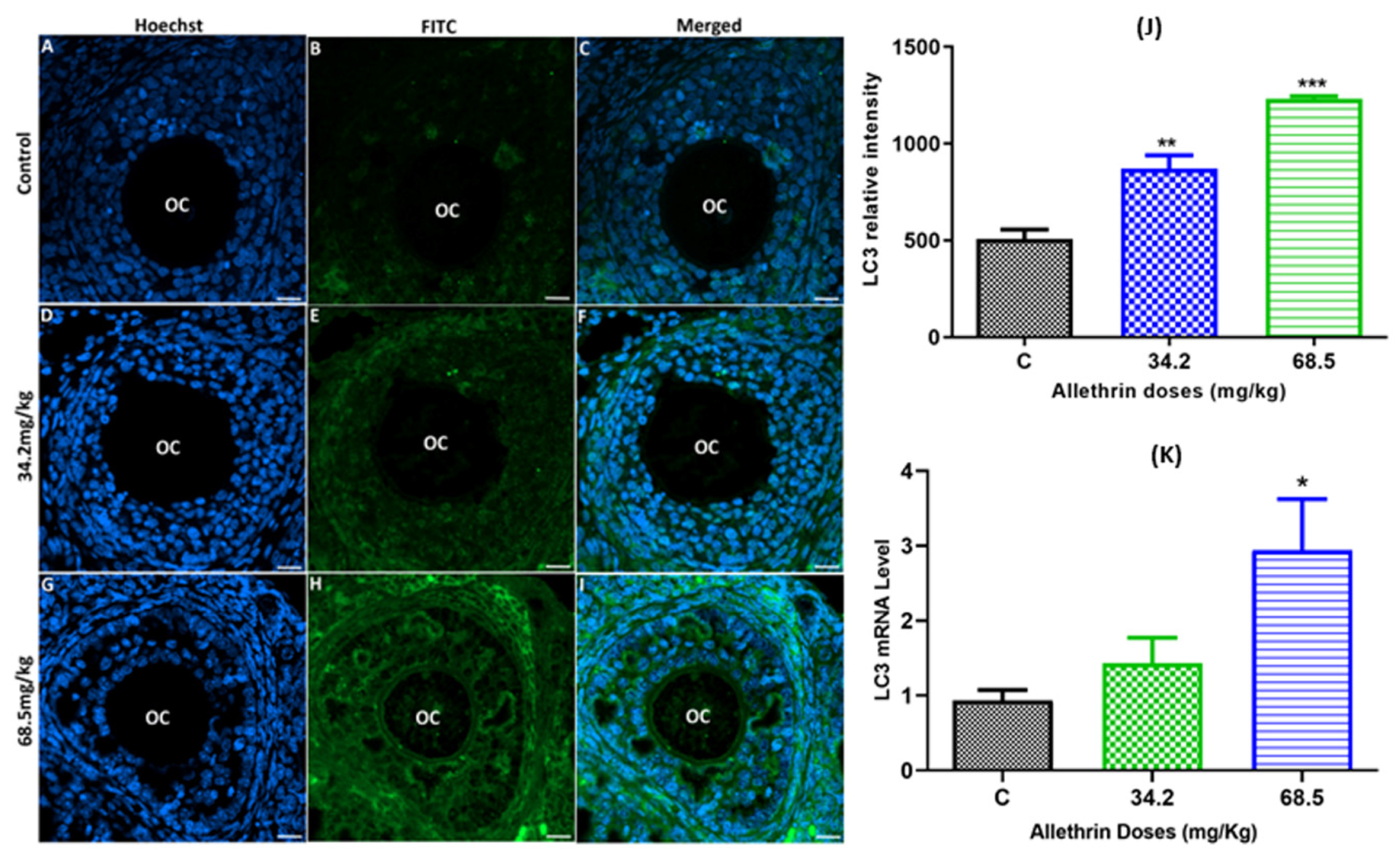
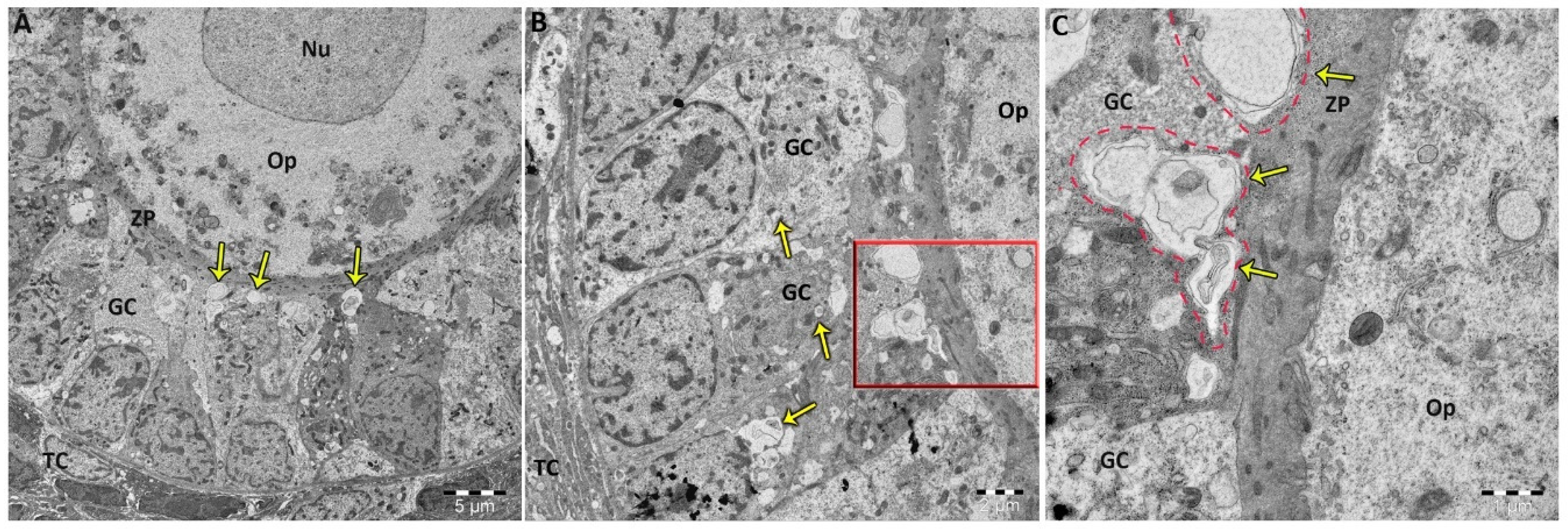
2.3. Allethrin Induced Autophagic and Apoptotic Marker Expression
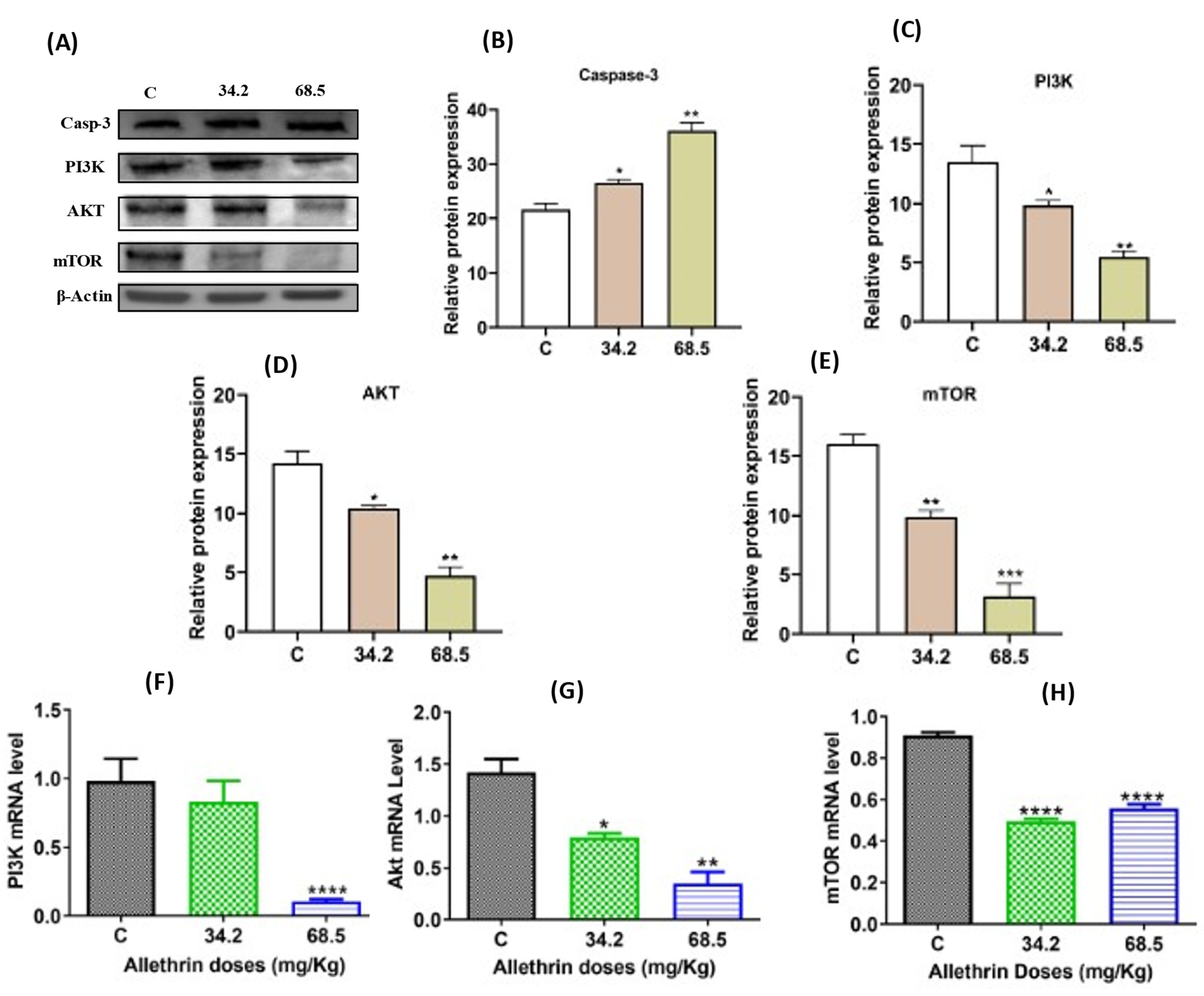
2.4. Allethrin Inhibited the PI3K/AKT/mTOR Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Animal Treatment and Sampling
4.2. Histological Preparation
4.3. Oxidative Stress Measurement
4.4. Western Blot Analysis
4.5. Immunofluorescence Staining and Confocal Microscopy
4.6. Ultrastructure Evaluation
4.7. Analysis of Gene Expression (RT–PCR)
4.8. Statistics
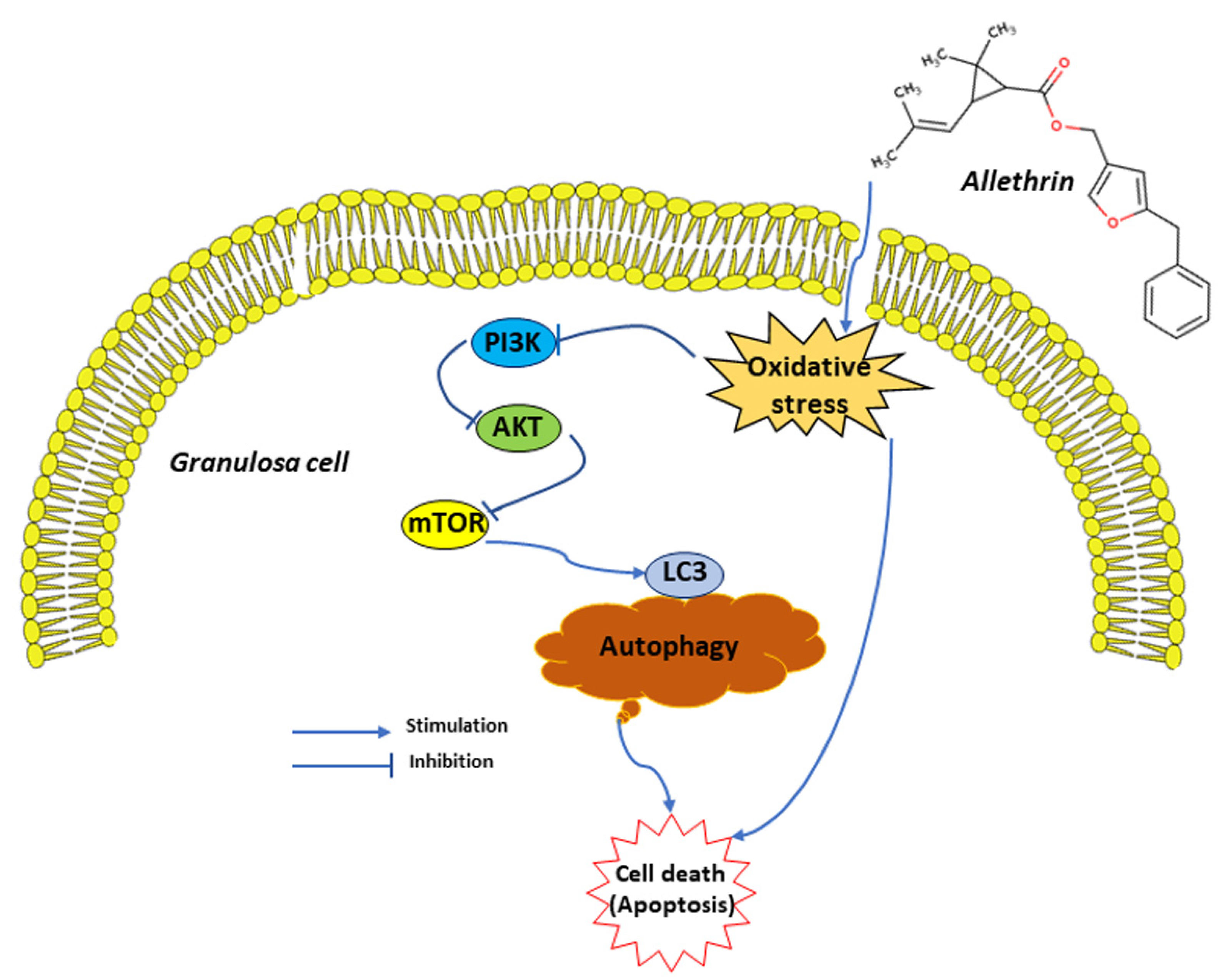
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D. Mothers, Babies, and Disease in Later Life; BMJ Publishing Group: London, UK, 1994. [Google Scholar]
- Osmond, C.; Barker, D.J. Fetal, infant, and childhood growth are predictors of coronary heart disease, diabetes, and hypertension in adult men and women. Environ. Health Perspect. 2000, 108 (Suppl. 3), 545–553. [Google Scholar] [CrossRef] [PubMed]
- Tamashiro, K.L.; Moran, T.H. Perinatal environment and its influences on metabolic programming of offspring. Physiol. Behav. 2010, 100, 560–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Boo, H.A.; Harding, J.E. The developmental origins of adult disease (Barker) hypothesis. Aust. N. Z. J. Obstet. Gynaecol. 2006, 46, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Ortega, H.H.; Salvetti, N.R.; Padmanabhan, V. Developmental programming: Prenatal androgen excess disrupts ovarian steroid receptor balance. Reproduction 2009, 137, 865. [Google Scholar] [CrossRef] [Green Version]
- Padmanabhan, V.; Sarma, H.N.; Savabieasfahani, M.; Steckler, T.L.; Veiga-Lopez, A. Developmental reprogramming of reproductive and metabolic dysfunction in sheep: Native steroids vs. environmental steroid receptor modulators. Int. J. Androl. 2010, 33, 394–404. [Google Scholar] [CrossRef] [Green Version]
- Hanson, B.; Johnstone, E.; Dorais, J.; Silver, B.; Peterson, C.M.; Hotaling, J. Female infertility, infertility-associated diagnoses, and comorbidities: A review. J. Assist. Reprod. Genet. 2017, 34, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Saillenfait, A.-M.; Ndiaye, D.; Sabaté, J.-P. Pyrethroids: Exposure and health effects—An update. Int. J. Hyg. Environ. Health 2015, 218, 281–292. [Google Scholar] [CrossRef]
- Todd, D.; Wohlers, D.; Citra, M. Agency for Toxic Stubstances and Disease Registry: Toxicological Profile for Pyrethrins and Pyrethroids; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2003.
- Bao, W.; Liu, B.; Simonsen, D.W.; Lehmler, H.-J. Association between exposure to pyrethroid insecticides and risk of all-cause and cause-specific mortality in the general US adult population. JAMA Intern. Med. 2020, 180, 367–374. [Google Scholar] [CrossRef]
- Sereda, B.; Bouwman, H.; Kylin, H. Comparing water, bovine milk, and indoor residual spraying as possible sources of DDTand pyrethroid residues in breast milk. J. Toxicol. Environ. Health A 2009, 72, 842–851. [Google Scholar] [CrossRef]
- Jurewicz, J.; Radwan, P.; Wielgomas, B.; Radwan, M.; Karwacka, A.; Kałużny, P.; Piskunowicz, M.; Dziewirska, E.; Hanke, W. Exposure to pyrethroid pesticides and ovarian reserve. Environ. Int. 2020, 144, 106028. [Google Scholar] [CrossRef]
- Fei, J.; Qu, J.-H.; Ding, X.-L.; Xue, K.; Lu, C.-C.; Chen, J.-F.; Song, L.; Xia, Y.-K.; Wang, S.-L.; Wang, X.-R. Fenvalerate inhibits the growth of primary cultured rat preantral ovarian follicles. Toxicology 2010, 267, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kotil, T.; Yön, N.D. The effects of permethrin on rat ovarian tissue morphology. Exp. Toxicol. Pathol. 2015, 67, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, K.W.; Baird, D.D.; Steiner, A.Z.; Bornman, R.M.; Travlos, G.S.; Wilson, R.E.; Longnecker, M.P. Antimüllerian hormone and lifestyle, reproductive, and environmental factors among women in rural South Africa. Epidemiology 2015, 26, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Ma, X.; Li, F.; Liu, J. Exposure to multiple pyrethroid insecticides affects ovarian follicular development via modifying microRNA expression. Sci. Total Environ. 2022, 828, 154384. [Google Scholar] [CrossRef] [PubMed]
- Ravula, A.R.; Yenugu, S. Effect of oral administration of a mixture of pyrethroids at doses relevant to human exposure on the general and male reproductive physiology in the rat. Ecotoxicol. Environ. Saf. 2021, 208, 111714. [Google Scholar] [CrossRef] [PubMed]
- Moya-Quiles, M.R.; Muñoz-Delgado, E.; Vidal, C.J. Effect of the pyrethroid insecticide allethrin on membrane fluidity. Biochem. Mol. Biol. Int. 1995, 36, 1299–1308. [Google Scholar]
- Taiwo Idowu, E.; Aimufua, O.J.; Yomi-Onilude, E.; Akinsanya, B.; Adetoro Otubanjo, O. Toxicological effects of prolonged and intense use of mosquito coil emission in rats and its implications on malaria control. Rev. Biol. Trop. 2013, 61, 1463–1473. [Google Scholar]
- Al-Mamun, M.A.; Rahman, M.A.; Rahman, M.H.; Hoque, K.; Ferdousi, Z.; Matin, M.N.; Reza, M.A. Biochemical and histological alterations induced by the smoke of allethrin based mosquito coil on mice model. BMC Clin. Pathol. 2017, 17, 19. [Google Scholar]
- Naz, M.; Rehman, N.; Ansari, M.N.; Kamal, M.; Ganaie, M.A.; Awaad, A.S.; Alqasoumi, S.I. Comparative study of subchronic toxicities of mosquito repellents (coils, mats and liquids) on vital organs in Swiss albino mice. Saudi Pharm. J. 2019, 27, 348–353. [Google Scholar] [CrossRef]
- Divakar, D.; Sasmal, D.; Kumar, A.; Sharma, N. Prophylactic Role of Piperine and Curcumin in Allethrin Altered Hematological and Biochemical Parameters in Swiss Albino Mice. Pharmacologia 2015, 6, 396–412. [Google Scholar]
- Mehrpour, O.; Karrari, P.; Zamani, N.; Tsatsakis, A.M.; Abdollahi, M. Occupational exposure to pesticides and consequences on male semen and fertility: A review. Toxicol. Lett. 2014, 230, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Madhubabu, G.; Yenugu, S. Allethrin induces oxidative stress, apoptosis and calcium release in rat testicular carcinoma cells (LC540). Toxicol. In Vitro 2014, 28, 1386–1395. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Tiwari, M.; Pandey, A.N.; Shrivastav, T.G.; Chaube, S.K. Impact of stress on oocyte quality and reproductive outcome. J. Biomed. Sci. 2016, 23, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kükürt, A.; Karapehlivan, M. Protective effect of astaxanthin on experimental ovarian damage in rats. J. Biochem. Mol. Toxicol. 2021, 36, e22966. [Google Scholar] [CrossRef]
- Agarwal, A.; Aponte-Mellado, A.; Premkumar, B.J.; Shaman, A.; Gupta, S. The effects of oxidative stress on female reproduction: A review. Reprod. Biol. Endocrinol. 2012, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Gao, X.; Zhu, J.; Cheng, K.; Tang, M. Mechanisms involved in reproductive toxicity caused by nickel nanoparticle in female rats. Environ. Toxicol. 2016, 31, 1674–1683. [Google Scholar] [CrossRef]
- Singh, M.; Verma, Y.; Rana, S.V.S. Attributes of oxidative stress in the reproductive toxicity of nickel oxide nanoparticles in male rats. Environ. Sci. Pollut. Res. 2022, 29, 5703–5717. [Google Scholar] [CrossRef]
- Liu, T.; Di, Q.-N.; Sun, J.-H.; Zhao, M.; Xu, Q.; Shen, Y. Effects of nonylphenol induced oxidative stress on apoptosis and autophagy in rat ovarian granulosa cells. Chemosphere 2020, 261, 127693. [Google Scholar] [CrossRef]
- An, J.; Yin, L.; Shang, Y.; Zhong, Y.; Zhang, X.; Wu, M.; Yu, Z.; Sheng, G.; Fu, J.; Huang, Y. The combined effects of BDE47 and BaP on oxidatively generated DNA damage in L02 cells and the possible molecular mechanism. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2011, 721, 192–198. [Google Scholar] [CrossRef]
- Siddique, S.; Sadeu, J.C.; Foster, W.G.; Feng, Y.l.; Zhu, J. In vitro exposure to cigarette smoke induces oxidative stress in follicular cells of F1 hybrid mice. J. Appl. Toxicol. 2014, 34, 224–226. [Google Scholar] [CrossRef]
- Tsai-Turton, M.; Nakamura, B.N.; Luderer, U. Induction of apoptosis by 9, 10-dimethyl-1, 2-benzanthracene in cultured preovulatory rat follicles is preceded by a rise in reactive oxygen species and is prevented by glutathione. Biol. Reprod. 2007, 77, 442–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, G.I.; Robles, R.; Knudson, C.M.; Flaws, J.A.; Korsmeyer, S.J.; Tilly, J.L. Prolongation of ovarian lifespan into advanced chronological age by Bax-deficiency. Nat. Genet. 1999, 21, 200–203. [Google Scholar] [CrossRef]
- Bristol-Gould, S.K.; Kreeger, P.K.; Selkirk, C.G.; Kilen, S.M.; Cook, R.W.; Kipp, J.L.; Shea, L.D.; Mayo, K.E.; Woodruff, T.K. Postnatal regulation of germ cells by activin: The establishment of the initial follicle pool. Dev. Biol. 2006, 298, 132–148. [Google Scholar] [CrossRef] [Green Version]
- Madden, J.A.; Hoyer, P.B.; Devine, P.J.; Keating, A.F. Acute 7, 12-dimethylbenz[a]anthracene exposure causes differential concentration-dependent follicle depletion and gene expression in neonatal rat ovaries. Toxicol. Appl. Pharmacol. 2014, 276, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.-Y.; Jiang, X.-X.; Zhu, X.; He, W.-Y.; Kuang, Y.-L.; Ren, K.; Lin, Y.; Gou, X. ROS activates JNK-mediated autophagy to counteract apoptosis in mouse mesenchymal stem cells in vitro. Acta Pharmacol. Sin. 2015, 36, 1473–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Levine, B. Autosis and autophagic cell death: The dark side of autophagy. Cell Death Differ. 2015, 22, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self-killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741–752. [Google Scholar] [CrossRef]
- Zhu, S.; Zhou, J.; Zhou, Z.; Zhu, Q. Abamectin induces apoptosis and autophagy by inhibiting reactive oxygen species-mediated PI3K/AKT signaling in MGC803 cells. J. Biochem. Mol. Toxicol. 2019, 33, e22336. [Google Scholar] [CrossRef]
- An, J.; Zhou, Q.; Wu, M.; Wang, L.; Zhong, Y.; Feng, J.; Shang, Y.; Chen, Y. Interactions between oxidative stress, autophagy and apoptosis in A549 cells treated with aged black carbon. Toxicol. In Vitro 2019, 54, 67–74. [Google Scholar] [CrossRef]
- Ylä-Anttila, P.; Vihinen, H.; Jokitalo, E.; Eskelinen, E.L. Monitoring autophagy by electron microscopy in Mammalian cells. Methods Enzymol. 2009, 452, 143–164. [Google Scholar]
- Lou, Y.; Yu, W.; Han, L.; Yang, S.; Wang, Y.; Ren, T.; Yu, J.; Zhao, A. ROS activates autophagy in follicular granulosa cells via mTOR pathway to regulate broodiness in goose. Anim. Reprod. Sci. 2017, 185, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Duan, P.; Hu, C.; Quan, C.; Yu, T.; Zhou, W.; Yuan, M.; Shi, Y.; Yang, K. 4-Nonylphenol induces apoptosis, autophagy and necrosis in Sertoli cells: Involvement of ROS-mediated AMPK/AKT-mTOR and JNK pathways. Toxicology 2016, 341, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Gump, J.M.; Thorburn, A. Autophagy and apoptosis: What is the connection? Trends Cell Biol. 2011, 21, 387–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maizels, E.T.; Cottom, J.; Jones, J.C.; Hunzicker-Dunn, M. Follicle stimulating hormone (FSH) activates the p38 mitogen-activated protein kinase pathway, inducing small heat shock protein phosphorylation and cell rounding in immature rat ovarian granulosa cells. Endocrinology 1998, 139, 3353–3356. [Google Scholar] [CrossRef]
- Law, N.C.; Donaubauer, E.M.; Zeleznik, A.J.; Hunzicker-Dunn, M. How protein kinase A activates canonical tyrosine kinase signaling pathways to promote granulosa cell differentiation. Endocrinology 2017, 158, 2043–2051. [Google Scholar] [CrossRef]
- Ravikumar, B.; Sarkar, S.; Davies, J.E.; Futter, M.; Garcia-Arencibia, M.; Green-Thompson, Z.W.; Jimenez-Sanchez, M.; Korolchuk, V.I.; Lichtenberg, M.; Luo, S. Regulation of mammalian autophagy in physiology and pathophysiology. Physiol. Rev. 2010, 90, 1383–1435. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Guan, K.-L. mTOR: A pharmacologic target for autophagy regulation. J. Clin. Investig. 2015, 125, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Sun, B.; Dong, C.; Lei, H.; Gong, Y.; Li, M.; Zhang, Y.; Zhang, H.; Sun, L. Knockdown of inhibitor of differentiation 1 suppresses proliferation and induces apoptosis by inactivating PI3K/Akt/mTOR signaling in hemangioma-derived endothelial cells. Biomed. Pharmacother. 2019, 111, 236–243. [Google Scholar] [CrossRef]
- Heras-Sandoval, D.; Pérez-Rojas, J.M.; Hernández-Damián, J.; Pedraza-Chaverri, J. The role of PI3K/AKT/mTOR pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration. Cell. Signal. 2014, 26, 2694–2701. [Google Scholar] [CrossRef]
- Lin, C.-J.; Chen, T.-L.; Tseng, Y.-Y.; Wu, G.-J.; Hsieh, M.-H.; Lin, Y.-W.; Chen, R.-M. Honokiol induces autophagic cell death in malignant glioma through reactive oxygen species-mediated regulation of the p53/PI3K/Akt/mTOR signaling pathway. Toxicol. Appl. Pharmacol. 2016, 304, 59–69. [Google Scholar] [CrossRef]
- Huang, W.; Cao, Z.; Zhang, J.; Ji, Q.; Li, Y. Aflatoxin B1 promotes autophagy associated with oxidative stress-related PI3K/AKT/mTOR signaling pathway in mice testis. Environ. Pollut. 2019, 255, 113317. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Quan, C.; Duan, P.; Tang, S.; Chen, W.; Yang, K. Nonylphenol induced apoptosis and autophagy involving the Akt/mTOR pathway in prepubertal Sprague-Dawley male rats in vivo and in vitro. Toxicology 2016, 373, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, H.; Liu, Y.; Yang, L.; Sun, L.; Tian, Y.; Zhao, B.; Lu, H. The PI3K/Akt/mTOR signaling pathway plays a role in regulating aconitine-induced autophagy in mouse liver. Res. Vet. Sci. 2019, 124, 317–320. [Google Scholar] [CrossRef] [PubMed]
- WHO. The World Health Organization Guidelines to Classification of Pesticides by Hazard; WHO: Geneva, Switzerland, 1996. [Google Scholar]
- Hossain, M.M.; Suzuki, T.; Sato, I.; Takewaki, T.; Suzuki, K.; Kobayashi, H. Neuromechanical effects of pyrethroids, allethrin, cyhalothrin and deltamethrin on the cholinergic processes in rat brain. Life Sci. 2005, 77, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Madhubabu, G.; Yenugu, S. Allethrin induced toxicity in the male reproductive tract of rats contributes to disruption in the transcription of genes involved in germ cell production. Environ. Toxicol. 2014, 29, 1330–1345. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Clark, J.M.; Sheets, L.P.; Mullin, L.S.; Piccirillo, V.J.; Sargent, D.; Stevens, J.T.; Weiner, M.L. Mechanisms of pyrethroid neurotoxicity: Implications for cumulative risk assessment. Toxicology 2002, 171, 3–59. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. [30] Microsomal lipid peroxidation. In Methods Enzymol; Elsevier: Amsterdam, The Netherlands, 1978; Volume 52, pp. 302–310. [Google Scholar]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. In Methods Enzymol; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Aldawood, N.; Alrezaki, A.; Alanazi, S.; Amor, N.; Alwasel, S.; Sirotkin, A.; Harrath, A.H. Acrylamide impairs ovarian function by promoting apoptosis and affecting reproductive hormone release, steroidogenesis and autophagy-related genes: An in vivo study. Ecotoxicol. Environ. Saf. 2020, 197, 110595. [Google Scholar] [CrossRef]
- Harrath, A.H.; Alrezaki, A.; Jalouli, M.; Aldawood, N.; Aldahmash, W.; Mansour, L.; Alwasel, S. Ethylbenzene exposure disrupts ovarian function in Wistar rats via altering folliculogenesis and steroidogenesis-related markers and activating autophagy and apoptosis. Ecotoxicol. Environ. Saf. 2022, 229, 113081. [Google Scholar] [CrossRef]
- Zhihan, T.; Xinyi, M.; Qingying, L.; Rufei, G.; Yan, Z.; Xuemei, C.; Yanqing, G.; Yingxiong, W.; Junlin, H. Autophagy participates in cyst breakdown and primordial folliculogenesis by reducing reactive oxygen species levels in perinatal mouse ovaries. J. Cell. Physiol. 2019, 234, 6125–6135. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Sequences |
|---|---|
| LC3 | F: TGTTAGGCTTGCTCTTTTGG R: GCAGAGGAAATGACCACAGAT |
| PI3K | F: GATGTCTGCGTTAGGGCTTACC R: TCAGCATCATGGAGAACAGGAT |
| AKT | F: CTCATTCCAGACCCACGAC R: ACAGCCCGAAGTCCGTTA |
| mTOR | F: TGCCTTCACAGATACCCAGTAC R: AGGTAGACCTTAAACTCGGAC |
| GAPDH | F: GCATCTTCTTGTGCAGTGCC R: GATGGTGATGGGTTTCCCGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jalouli, M.; Mofti, A.; Elnakady, Y.A.; Nahdi, S.; Feriani, A.; Alrezaki, A.; Sebei, K.; Bizzarri, M.; Alwasel, S.; Harrath, A.H. Allethrin Promotes Apoptosis and Autophagy Associated with the Oxidative Stress-Related PI3K/AKT/mTOR Signaling Pathway in Developing Rat Ovaries. Int. J. Mol. Sci. 2022, 23, 6397. https://doi.org/10.3390/ijms23126397
Jalouli M, Mofti A, Elnakady YA, Nahdi S, Feriani A, Alrezaki A, Sebei K, Bizzarri M, Alwasel S, Harrath AH. Allethrin Promotes Apoptosis and Autophagy Associated with the Oxidative Stress-Related PI3K/AKT/mTOR Signaling Pathway in Developing Rat Ovaries. International Journal of Molecular Sciences. 2022; 23(12):6397. https://doi.org/10.3390/ijms23126397
Chicago/Turabian StyleJalouli, Maroua, Afoua Mofti, Yasser A. Elnakady, Saber Nahdi, Anouar Feriani, Abdelkarem Alrezaki, Khaled Sebei, Mariano Bizzarri, Saleh Alwasel, and Abdel Halim Harrath. 2022. "Allethrin Promotes Apoptosis and Autophagy Associated with the Oxidative Stress-Related PI3K/AKT/mTOR Signaling Pathway in Developing Rat Ovaries" International Journal of Molecular Sciences 23, no. 12: 6397. https://doi.org/10.3390/ijms23126397