
From Molecules to Behavior in Long-Term Inorganic Mercury Intoxication: Unraveling Proteomic Features in Cerebellar Neurodegeneration of Rats

 , , , ,
, , , ,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
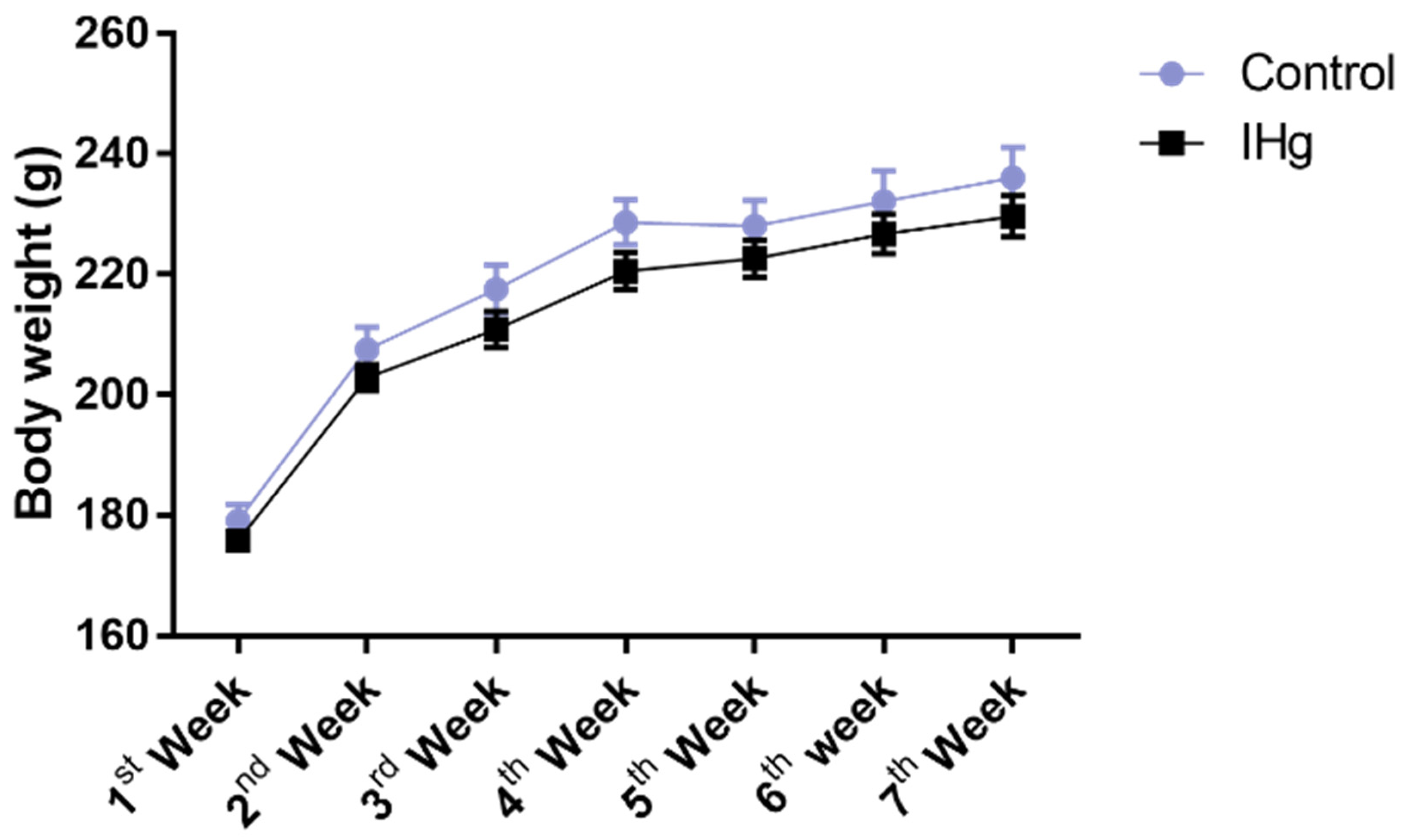
2.1. Long-Term Exposure to IHg Did Not Impair Mass Body Gain of Adult Rats
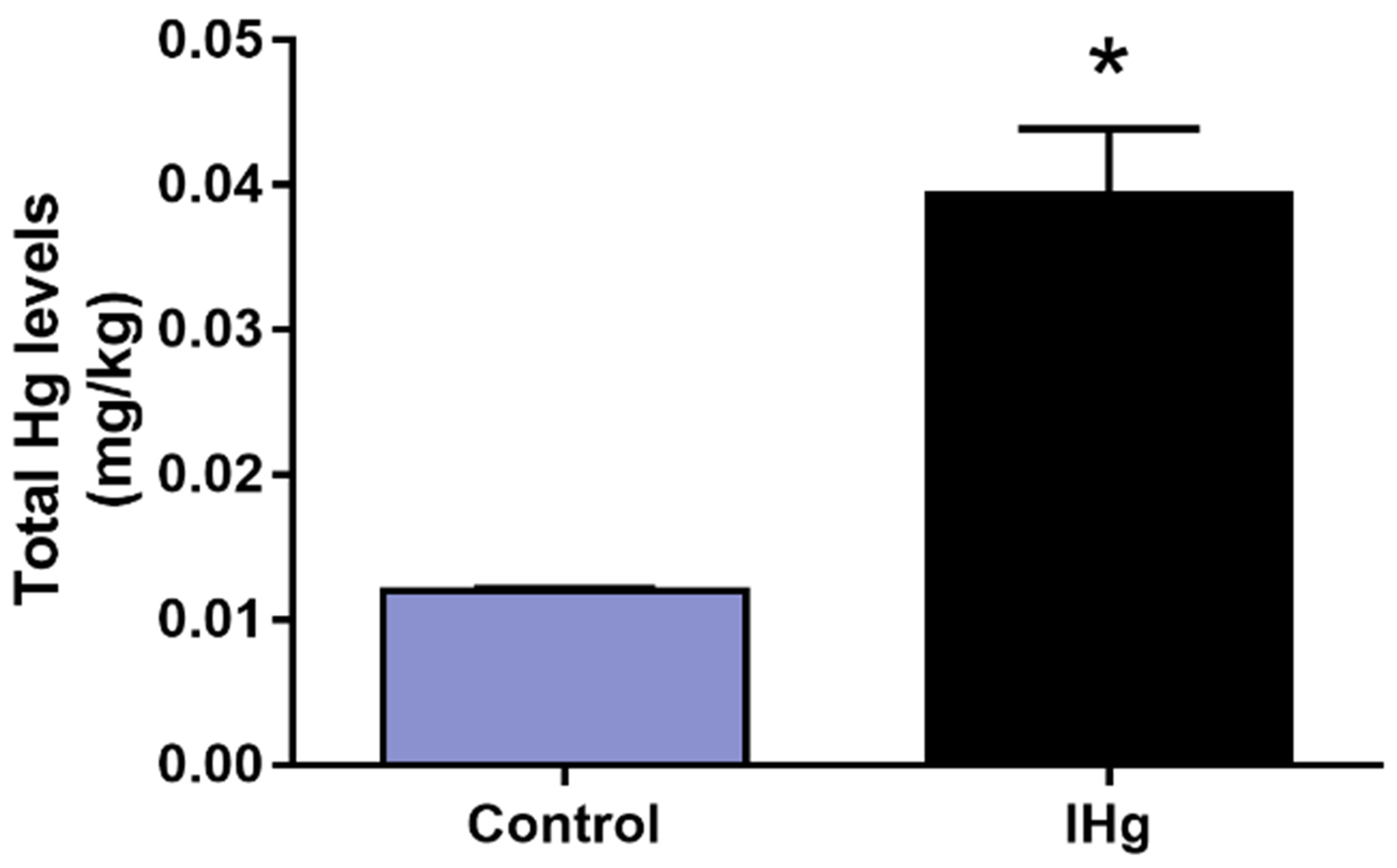
2.2. Prolonged IHg Exposure Increased the Total Mercury Content in Cerebellar Parenchyma of Adult Rats
2.3. IHg Long-Term Exposure Triggered Oxidative Stress in the Cerebellum of Adult Rats
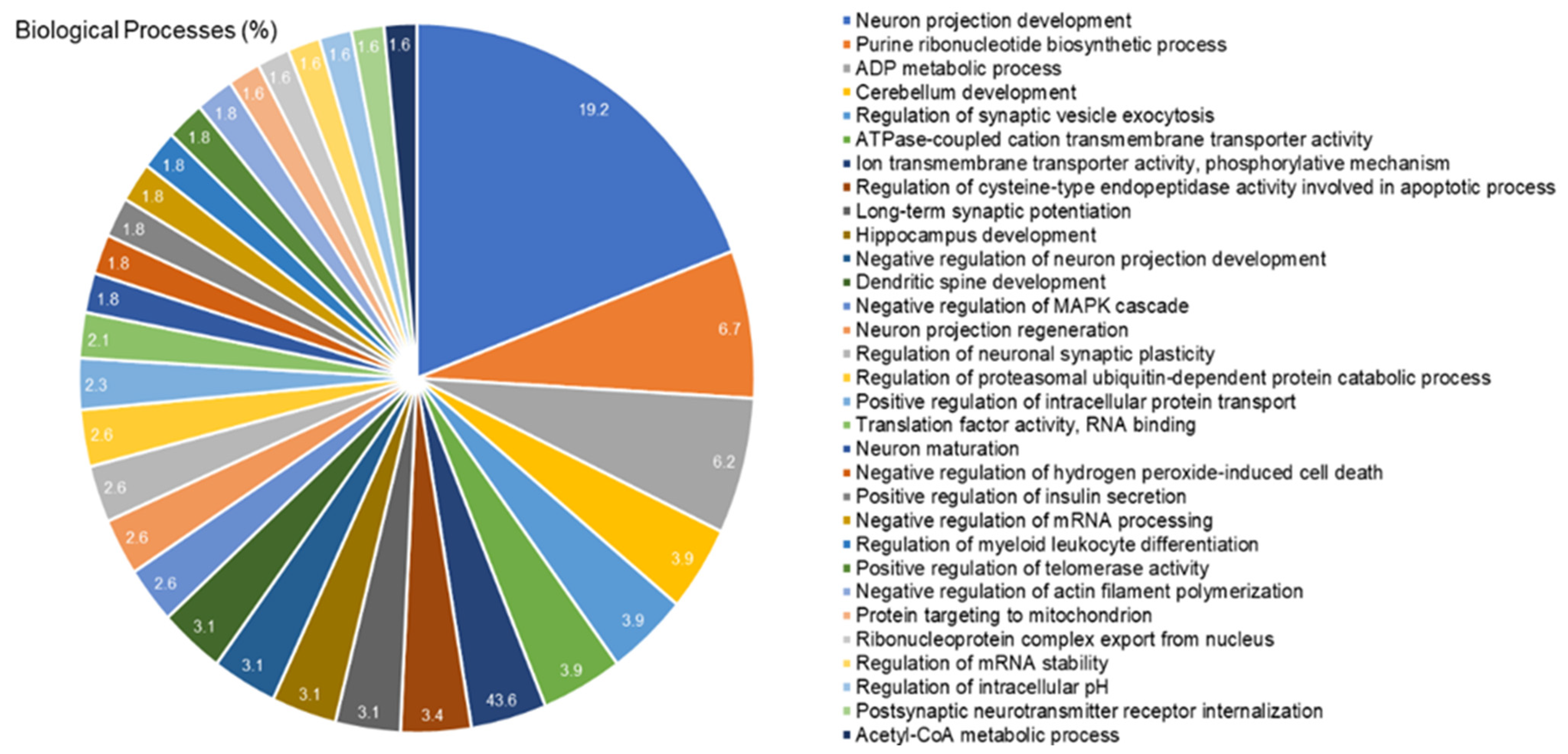
2.4. The Cerebellar Proteomic Profile Was Significantly Modulated after Exposure to IHg Rats
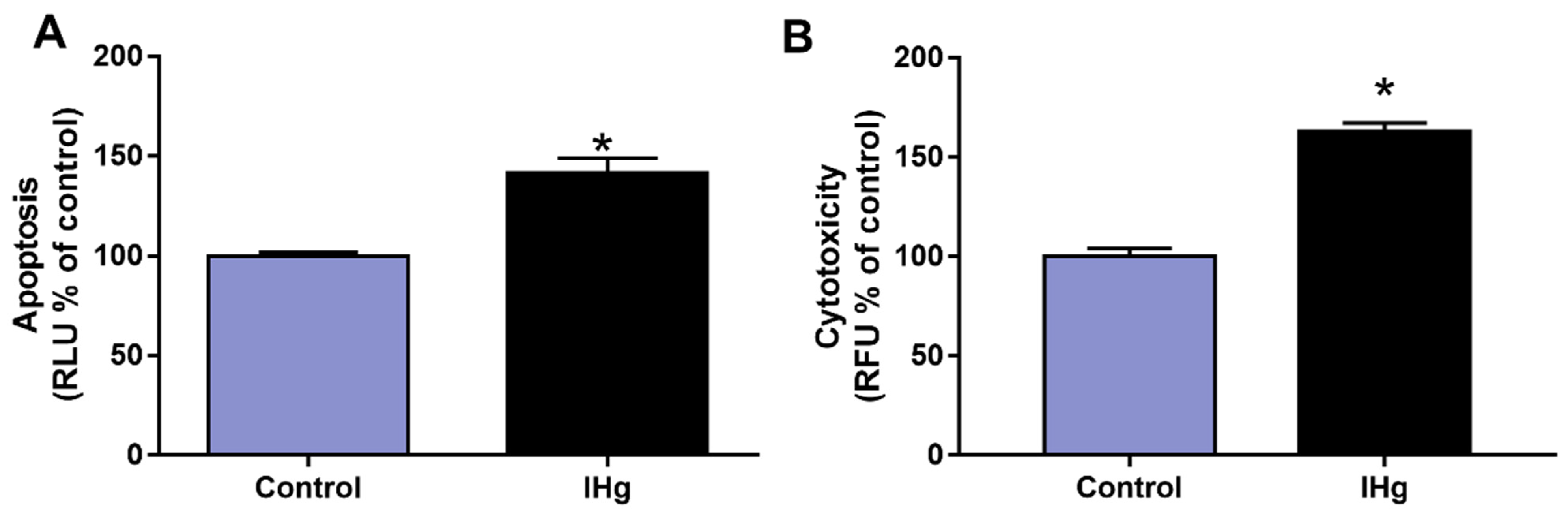
2.5. The IHg Exposure Increased Cell Death by Cytotoxicity and Apoptosis in the Cerebellum of Adult Rats
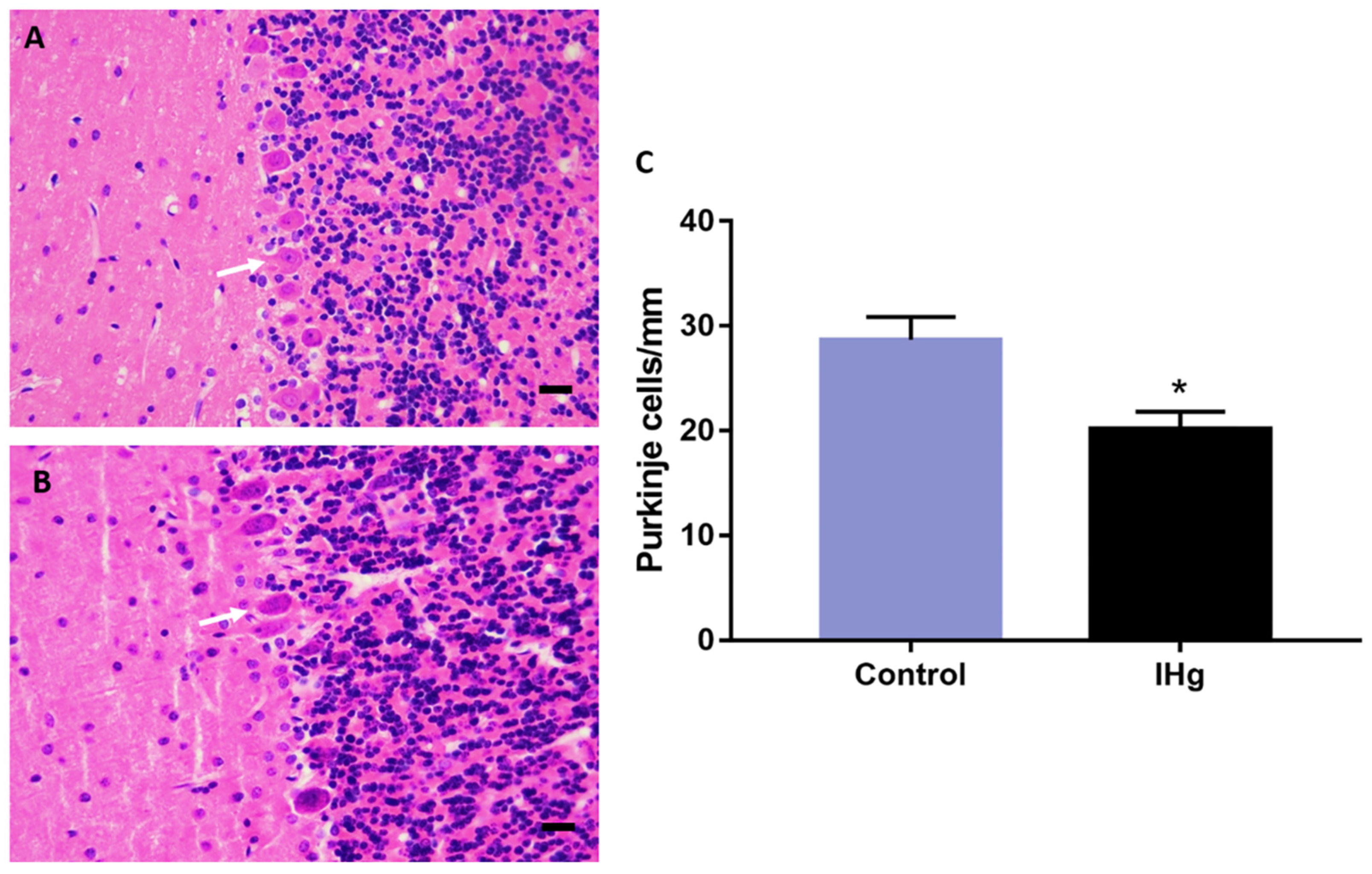
2.6. The IHg Intoxication Reduces Purkinje Cells Density in Adult Rats
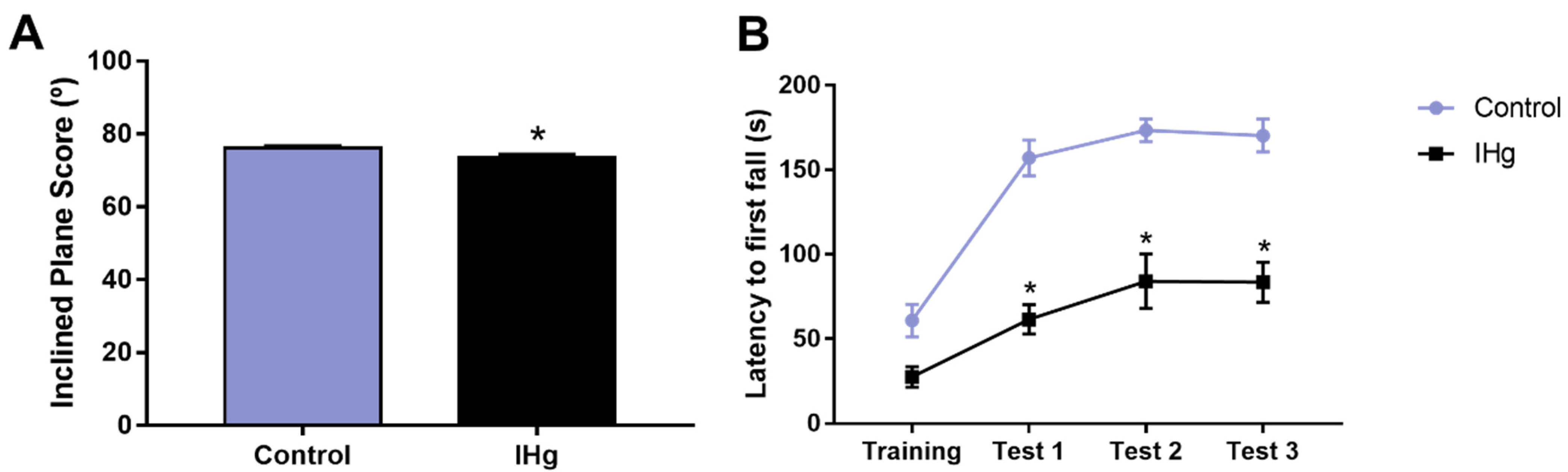
2.7. The Successive IHg-Induced Damages on the Cerebellum of Rats Led to a Poor Motor Function
3. Discussion
4. Materials and Methods
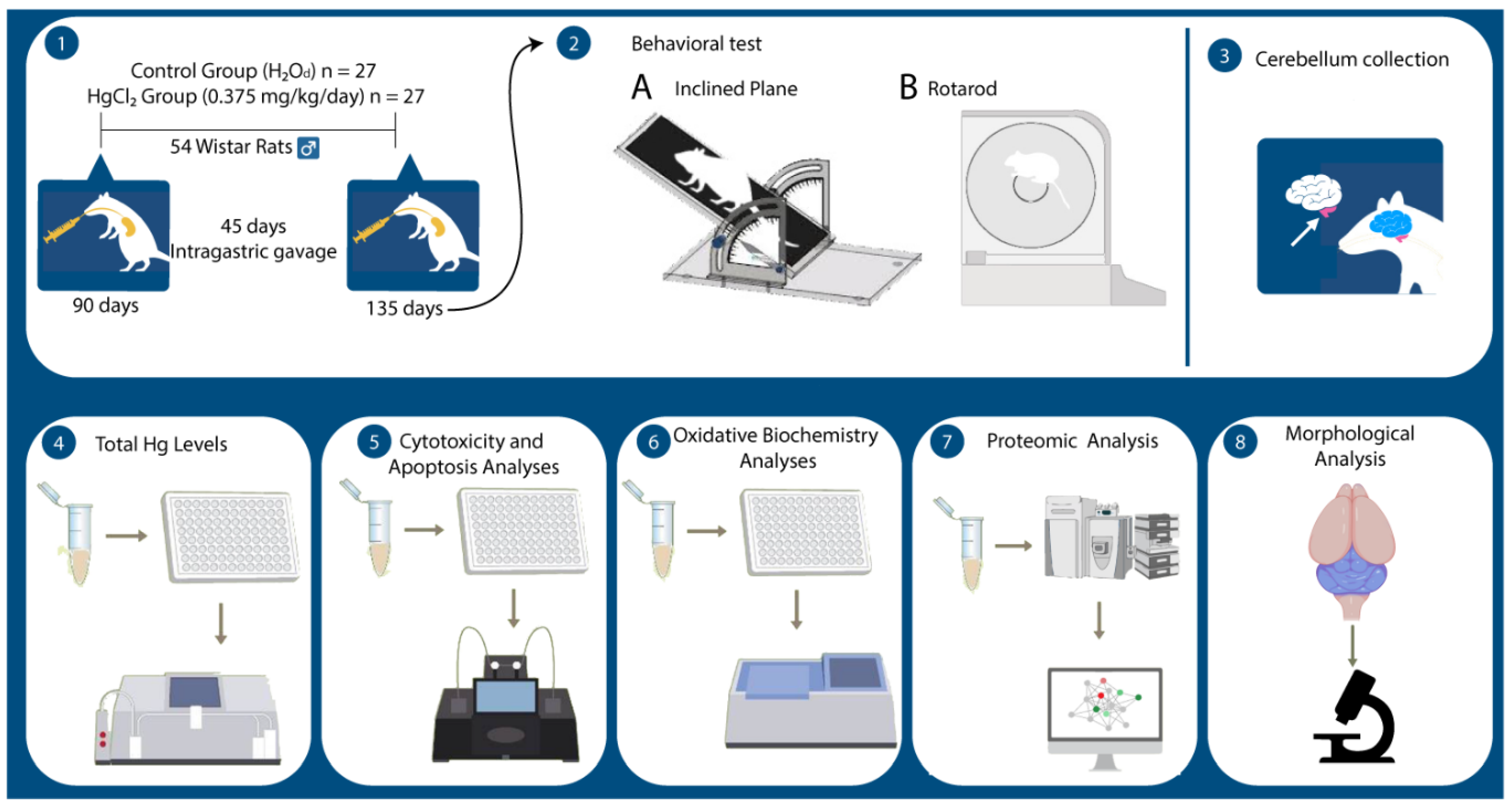
4.1. Ethical Statement and Experimental Protocol
4.2. Behavioral Assessment
4.2.1. Inclined Plane Test
4.2.2. Rotarod Test
4.3. Sample Collection and Preparation
4.4. Mercury Measurements
4.5. Oxidative Biochemistry Analyses
4.5.1. Antioxidant Capacity against Peroxyl Radicals (ACAPs)
4.5.2. Lipid Peroxidation (LPO)
4.6. Proteomic Approach and Bioinformatic Analyses
4.7. Apoptosis and Cytotoxicity Assays
4.8. Morphological Analysis
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IHg | Inorganic Mercury |
| CNS | Central Nervous System |
| i.p. | Intraperitoneal |
| w/v | weight/volume |
| ABAP | 2,2′-azobis-2-methylpropionamidine dihydrochloride |
| LPO | Lipid Peroxidation |
| MDA | Malondialdehyde |
| UPLC | Ultra performance liquid chromatography |
| PLGS | Protein Lynx Global SERVER |
| RLU | Relative Luminescence Unit |
| RFU | Relative Fluorescence Unit |
| ROS | Reactive Oxygen Species |
| HSP | Heat Shock Proteins |
References
- Ha, E.; Basu, N.; Bose-O’Reilly, S.; Dórea, J.G.; McSorley, E.; Sakamoto, M.; Chan, H.M. Current progress on understanding the impact of mercury on human health. Environ. Res. 2017, 152, 419–433. [Google Scholar] [CrossRef] [Green Version]
- Orr, S.E.; Barnes, M.C.; Joshee, L.; Uchakina, O.; McKallip, R.J.; Bridges, C.C. Potential mechanisms of cellular injury following exposure to a physiologically relevant species of inorganic mercury. Toxicol. Lett. 2019, 304, 13–20. [Google Scholar] [CrossRef]
- Crespo-Lopez, M.E.; Augusto-Oliveira, M.; Lopes-Araújo, A.; Santos-Sacramento, L.; Yuki Takeda, P.; Macchi, B.M.; do Nascimento, J.L.M.; Maia, C.S.F.; Lima, R.R.; Arrifano, G.P. Mercury: What can we learn from the Amazon? Environ. Int. 2021, 146, 106223. [Google Scholar] [CrossRef] [PubMed]
- Santos-Sacramento, L.; Arrifano, G.P.; Lopes-Araújo, A.; Augusto-Oliveira, M.; Albuquerque-Santos, R.; Takeda, P.Y.; Souza-Monteiro, J.R.; Macchi, B.M.; do Nascimento, J.L.M.; Lima, R.R.; et al. Human neurotoxicity of mercury in the Amazon: A scoping review with insights and critical considerations. Ecotoxicol. Environ. Saf. 2021, 208, 111686. [Google Scholar] [CrossRef]
- Nevado, J.J.B.; Martín-Doimeadios, R.C.R.; Moreno, M.J.; do Nascimento, J.L.M.; Herculano, A.M.; Crespo-López, M.E. Mercury speciation analysis on cell lines of the human central nervous system to explain genotoxic effects. Microchem. J. 2009, 93, 12–16. [Google Scholar] [CrossRef]
- Rodríguez Martín-Doimeadios, R.C.; Berzas Nevado, J.J.; Guzmán Bernardo, F.J.; Jiménez Moreno, M.; Arrifano, G.P.; Herculano, A.M.; do Nascimento, J.L.; Crespo-López, M.E. Comparative study of mercury speciation in commercial fishes of the Brazilian Amazon. Environ. Sci. Pollut. Res. Int. 2014, 21, 7466–7479. [Google Scholar] [CrossRef]
- Machado, C.L.R.; Crespo-Lopez, M.E.; Augusto-Oliveira, M.; Arrifano, G.P.; Macchi, B.M.; Lopes-Araújo, A.; Santos-Sacramento, L.; Souza-Monteiro, J.R.; Alvarez-Leite, J.I.; Souza, C.B.A. Eating in the Amazon: Nutritional Status of the Riverine Populations and Possible Nudge Interventions. Foods 2021, 10, 1015. [Google Scholar] [CrossRef]
- Park, J.D.; Zheng, W. Human exposure and health effects of inorganic and elemental mercury. J. Prev. Med. Public Health Yebang Uihakhoe Chi 2012, 45, 344–352. [Google Scholar] [CrossRef]
- Ahmad, S.; Mahmood, R. Mercury chloride toxicity in human erythrocytes: Enhanced generation of ROS and RNS, hemoglobin oxidation, impaired antioxidant power, and inhibition of plasma membrane redox system. Environ. Sci. Pollut. Res. Int. 2019, 26, 5645–5657. [Google Scholar] [CrossRef]
- Pamphlett, R.; Kum Jew, S. Inorganic mercury in human astrocytes, oligodendrocytes, corticomotoneurons and the locus ceruleus: Implications for multiple sclerosis, neurodegenerative disorders and gliomas. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2018, 31, 807–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risher, J.F.; De Rosa, C.T. Inorganic: The other mercury. J. Environ. Health 2007, 70, 9–16; discussion 40. [Google Scholar]
- Mariën, P.; Borgatti, R. Language and the cerebellum. Handb. Clin. Neurol. 2018, 154, 181–202. [Google Scholar] [CrossRef]
- Schmahmann, J.D.; Guell, X.; Stoodley, C.J.; Halko, M.A. The Theory and Neuroscience of Cerebellar Cognition. Annu. Rev. Neurosci. 2019, 42, 337–364. [Google Scholar] [CrossRef]
- Stoodley, C.J. The Cerebellum and Neurodevelopmental Disorders. Cerebellum 2016, 15, 34–37. [Google Scholar] [CrossRef]
- Koeppen, A.H. The neuropathology of the adult cerebellum. Handb. Clin. Neurol. 2018, 154, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Chemelo, V.; Bittencourt, L.O.; Aragão, W.A.B.; Dos Santos, S.M.; Souza-Rodrigues, R.D.; Ribeiro, C.; Monteiro, M.C.; Lima, R.R. Long-Term Exposure to Inorganic Mercury Leads to Oxidative Stress in Peripheral Blood of Adult Rats. Biol. Trace Elem. Res. 2021, 199, 2992–3000. [Google Scholar] [CrossRef]
- Aragão, W.A.B.; Teixeira, F.B.; Fagundes, N.C.F.; Fernandes, R.M.; Fernandes, L.M.P.; da Silva, M.C.F.; Amado, L.L.; Sagica, F.E.S.; Oliveira, E.H.C.; Crespo-Lopez, M.E.; et al. Hippocampal Dysfunction Provoked by Mercury Chloride Exposure: Evaluation of Cognitive Impairment, Oxidative Stress, Tissue Injury and Nature of Cell Death. Oxidative Med. Cell. Longev. 2018, 2018, 7878050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrêa, M.G.; Bittencourt, L.O.; Nascimento, P.C.; Ferreira, R.O.; Aragão, W.A.B.; Silva, M.C.F.; Gomes-Leal, W.; Fernandes, M.S.; Dionizio, A.; Buzalaf, M.R.; et al. Spinal cord neurodegeneration after inorganic mercury long-term exposure in adult rats: Ultrastructural, proteomic and biochemical damages associated with reduced neuronal density. Ecotoxicol. Environ. Saf. 2020, 191, 110159. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.B.; Fernandes, R.M.; Farias-Junior, P.M.; Costa, N.M.; Fernandes, L.M.; Santana, L.N.; Silva-Junior, A.F.; Silva, M.C.; Maia, C.S.; Lima, R.R. Evaluation of the effects of chronic intoxication with inorganic mercury on memory and motor control in rats. Int. J. Environ. Res. Public Health 2014, 11, 9171–9185. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, F.B.; de Oliveira, A.C.A.; Leão, L.K.R.; Fagundes, N.C.F.; Fernandes, R.M.; Fernandes, L.M.P.; da Silva, M.C.F.; Amado, L.L.; Sagica, F.E.S.; de Oliveira, E.H.C.; et al. Exposure to Inorganic Mercury Causes Oxidative Stress, Cell Death, and Functional Deficits in the Motor Cortex. Front. Mol. Neurosci. 2018, 11, 125. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, F.B.; Leão, L.K.R.; Bittencourt, L.O.; Aragão, W.A.B.; Nascimento, P.C.; Luz, D.A.; Braga, D.V.; Silva, M.; Oliveira, K.R.M.; Herculano, A.M.; et al. Neurochemical dysfunction in motor cortex and hippocampus impairs the behavioral performance of rats chronically exposed to inorganic mercury. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. 2019, 52, 143–150. [Google Scholar] [CrossRef]
- Bridges, C.C.; Zalups, R.K. Transport of inorganic mercury and methylmercury in target tissues and organs. J. Toxicol. Environ. Health. Part B Crit. Rev. 2010, 13, 385–410. [Google Scholar] [CrossRef]
- Aragão, W.A.B.; da Costa, N.M.M.; Fagundes, N.C.F.; Silva, M.C.F.; Alves-Junior, S.M.; Pinheiro, J.J.V.; Amado, L.L.; Crespo-López, M.E.; Maia, C.S.F.; Lima, R.R. Chronic exposure to inorganic mercury induces biochemical and morphological changes in the salivary glands of rats. Met. Integr. Biometal Sci. 2017, 9, 1271–1278. [Google Scholar] [CrossRef]
- Bonacker, D.; Stoiber, T.; Wang, M.; Böhm, K.J.; Prots, I.; Unger, E.; Thier, R.; Bolt, H.M.; Degen, G.H. Genotoxicity of inorganic mercury salts based on disturbed microtubule function. Arch. Toxicol. 2004, 78, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ozer, E.Y.; Rosenspire, A.J.; McCabe, M.J., Jr.; Worth, R.G.; Kindzelskii, A.L.; Warra, N.S.; Petty, H.R. Mercuric chloride damages cellular DNA by a non-apoptotic mechanism. Mutat. Res. 2000, 470, 19–27. [Google Scholar] [CrossRef]
- Silva-Pereira, L.C.; Cardoso, P.C.; Leite, D.S.; Bahia, M.O.; Bastos, W.R.; Smith, M.A.; Burbano, R.R. Cytotoxicity and genotoxicity of low doses of mercury chloride and methylmercury chloride on human lymphocytes in vitro. Braz. J. Med. Biol. Res. Rev. Bras. De Pesqui. Med. E Biol. 2005, 38, 901–907. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, J.M.; Harding, P.G.; Humes, H.D. Mitochondrial bioenergetics during the initiation of mercuric chloride-induced renal injury. I. Direct effects of in vitro mercuric chloride on renal mitochondrial function. J. Biol. Chem. 1982, 257, 60–67. [Google Scholar] [CrossRef]
- Araragi, S.; Kondoh, M.; Kawase, M.; Saito, S.; Higashimoto, M.; Sato, M. Mercuric chloride induces apoptosis via a mitochondrial-dependent pathway in human leukemia cells. Toxicology 2003, 184, 1–9. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Crespo-López, M.E.; Macêdo, G.L.; Pereira, S.I.; Arrifano, G.P.; Picanço-Diniz, D.L.; do Nascimento, J.L.; Herculano, A.M. Mercury and human genotoxicity: Critical considerations and possible molecular mechanisms. Pharmacol. Res. 2009, 60, 212–220. [Google Scholar] [CrossRef]
- Lim, P.; Wuenschell, G.E.; Holland, V.; Lee, D.H.; Pfeifer, G.P.; Rodriguez, H.; Termini, J. Peroxyl radical mediated oxidative DNA base damage: Implications for lipid peroxidation induced mutagenesis. Biochemistry 2004, 43, 15339–15348. [Google Scholar] [CrossRef] [PubMed]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. IJCB 2015, 30, 11–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative Stress and Advanced Lipoxidation and Glycation End Products (ALEs and AGEs) in Aging and Age-Related Diseases. Oxidative Med. Cell. Longev. 2019, 2019, 3085756. [Google Scholar] [CrossRef] [Green Version]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef] [Green Version]
- Perkins, A.; Nelson, K.J.; Parsonage, D.; Poole, L.B.; Karplus, P.A. Peroxiredoxins: Guardians against oxidative stress and modulators of peroxide signaling. Trends Biochem. Sci. 2015, 40, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef]
- Trostchansky, A.; Quijano, C.; Yadav, H.; Kelley, E.E.; Cassina, A.M. Interplay between Oxidative Stress and Metabolism in Signalling and Disease. Oxidative Med. Cell. Longev. 2016, 2016, 3274296. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Liu, D. Energetics and oxidative stress in synaptic plasticity and neurodegenerative disorders. Neuromolecular Med. 2002, 2, 215–231. [Google Scholar] [CrossRef] [PubMed]
- Korovila, I.; Hugo, M.; Castro, J.P.; Weber, D.; Höhn, A.; Grune, T.; Jung, T. Proteostasis, oxidative stress and aging. Redox Biol. 2017, 13, 550–567. [Google Scholar] [CrossRef]
- Dahl, J.U.; Gray, M.J.; Jakob, U. Protein quality control under oxidative stress conditions. J. Mol. Biol. 2015, 427, 1549–1563. [Google Scholar] [CrossRef] [Green Version]
- Reichmann, D.; Voth, W.; Jakob, U. Maintaining a Healthy Proteome during Oxidative Stress. Mol. Cell 2018, 69, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Shang, F.; Taylor, A. Ubiquitin-proteasome pathway and cellular responses to oxidative stress. Free Radic. Biol. Med. 2011, 51, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.J.; Fort, P.E. Heat Shock Proteins Regulatory Role in Neurodevelopment. Front. Neurosci. 2018, 12, 821. [Google Scholar] [CrossRef] [Green Version]
- Takei, Y.; Kikkawa, Y.S.; Atapour, N.; Hensch, T.K.; Hirokawa, N. Defects in Synaptic Plasticity, Reduced NMDA-Receptor Transport, and Instability of Postsynaptic Density Proteins in Mice Lacking Microtubule-Associated Protein 1A. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 15539–15554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, M.B.; do Nascimento, J.L.; Herculano, A.M.; Crespo-López, M.E. Molecular chaperones: Toward new therapeutic tools. Biomed. Pharmacother. Biomed. Pharmacother. 2011, 65, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Kannan, K.; Jain, S.K. Oxidative stress and apoptosis. Pathophysiology 2000, 7, 153–163. [Google Scholar] [CrossRef]
- Garrido, C.; Galluzzi, L.; Brunet, M.; Puig, P.E.; Didelot, C.; Kroemer, G. Mechanisms of cytochrome c release from mitochondria. Cell Death Differ. 2006, 13, 1423–1433. [Google Scholar] [CrossRef] [Green Version]
- Smeyne, R.J.; Chu, T.; Lewin, A.; Bian, F.; Sanlioglu, S.; Kunsch, C.; Lira, S.A.; Oberdick, J. Local control of granule cell generation by cerebellar Purkinje cells. Mol. Cell. Neurosci. 1995, 6, 230–251. [Google Scholar] [CrossRef]
- Medina, J.F. The multiple roles of Purkinje cells in sensori-motor calibration: To predict, teach and command. Curr. Opin. Neurobiol. 2011, 21, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Lamarão-Vieira, K.; Pamplona-Santos, D.; Nascimento, P.C.; Corrêa, M.G.; Bittencourt, L.O.; Dos Santos, S.M.; Cartágenes, S.C.; Fernandes, L.M.P.; Monteiro, M.C.; Maia, C.S.F.; et al. Physical Exercise Attenuates Oxidative Stress and Morphofunctional Cerebellar Damages Induced by the Ethanol Binge Drinking Paradigm from Adolescence to Adulthood in Rats. Oxidative Med. Cell. Longev. 2019, 2019, 6802424. [Google Scholar] [CrossRef]
- Jockusch, B.M.; Rothkegel, M.; Schwarz, G. Linking the synapse to the cytoskeleton: A breath-taking role for microfilaments. Neuroreport 2004, 15, 1535–1538. [Google Scholar] [CrossRef] [PubMed]
- Südhof, T.C. Calcium control of neurotransmitter release. Cold Spring Harb. Perspect. Biol. 2012, 4, a011353. [Google Scholar] [CrossRef]
- Yamauchi, T. Neuronal Ca2+/calmodulin-dependent protein kinase II--discovery, progress in a quarter of a century, and perspective: Implication for learning and memory. Biol. Pharm. Bull. 2005, 28, 1342–1354. [Google Scholar] [CrossRef] [Green Version]
- Picconi, B.; Gardoni, F.; Centonze, D.; Mauceri, D.; Cenci, M.A.; Bernardi, G.; Calabresi, P.; Di Luca, M. Abnormal Ca2+-calmodulin-dependent protein kinase II function mediates synaptic and motor deficits in experimental parkinsonism. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 5283–5291. [Google Scholar] [CrossRef]
- Chung, C.; Barylko, B.; Leitz, J.; Liu, X.; Kavalali, E.T. Acute dynamin inhibition dissects synaptic vesicle recycling pathways that drive spontaneous and evoked neurotransmission. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 1363–1376. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.E.; Chapman, E.R. Synaptophysin regulates the kinetics of synaptic vesicle endocytosis in central neurons. Neuron 2011, 70, 847–854. [Google Scholar] [CrossRef] [Green Version]
- Béïque, J.C.; Andrade, R. PSD-95 regulates synaptic transmission and plasticity in rat cerebral cortex. J. Physiol. 2003, 546, 859–867. [Google Scholar] [CrossRef]
- Boggs, J.M. Myelin basic protein: A multifunctional protein. Cell. Mol. Life Sci. CMLS 2006, 63, 1945–1961. [Google Scholar] [CrossRef]
- Pronker, M.F.; Lemstra, S.; Snijder, J.; Heck, A.J.; Thies-Weesie, D.M.; Pasterkamp, R.J.; Janssen, B.J. Structural basis of myelin-associated glycoprotein adhesion and signalling. Nat. Commun. 2016, 7, 13584. [Google Scholar] [CrossRef]
- Quarles, R.H. Myelin-associated glycoprotein (MAG): Past, present and beyond. J. Neurochem. 2007, 100, 1431–1448. [Google Scholar] [CrossRef] [PubMed]
- Koziol, L.F.; Budding, D.; Andreasen, N.; D’Arrigo, S.; Bulgheroni, S.; Imamizu, H.; Ito, M.; Manto, M.; Marvel, C.; Parker, K.; et al. Consensus paper: The cerebellum’s role in movement and cognition. Cerebellum 2014, 13, 151–177. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.B.; Fontes Ede, A., Jr.; de Carvalho, S.; da Silva, J.B.; Fernandes, L.M.; Oliveira, M.C.; Prediger, R.D.; Gomes-Leal, W.; Lima, R.R.; Maia, C.S. Minocycline mitigates motor impairments and cortical neuronal loss induced by focal ischemia in rats chronically exposed to ethanol during adolescence. Brain Res. 2014, 1561, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Lopes, G.O.; Martins Ferreira, M.K.; Davis, L.; Bittencourt, L.O.; Bragança Aragão, W.A.; Dionizio, A.; Rabelo Buzalaf, M.A.; Crespo-Lopez, M.E.; Maia, C.S.F.; Lima, R.R. Effects of Fluoride Long-Term Exposure over the Cerebellum: Global Proteomic Profile, Oxidative Biochemistry, Cell Density, and Motor Behavior Evaluation. Int. J. Mol. Sci. 2020, 21, 7297. [Google Scholar] [CrossRef] [PubMed]
- Leão, L.K.R.; Bittencourt, L.O.; Oliveira, A.C.; Nascimento, P.C.; Miranda, G.H.N.; Ferreira, R.O.; Nabiça, M.; Dantas, K.; Dionizio, A.; Cartágenes, S.; et al. Long-Term Lead Exposure Since Adolescence Causes Proteomic and Morphological Alterations in the Cerebellum Associated with Motor Deficits in Adult Rats. Int. J. Mol. Sci. 2020, 21, 3571. [Google Scholar] [CrossRef] [PubMed]
- Santana, L.; Bittencourt, L.O.; Nascimento, P.C.; Fernandes, R.M.; Teixeira, F.B.; Fernandes, L.M.P.; Freitas Silva, M.C.; Nogueira, L.S.; Amado, L.L.; Crespo-Lopez, M.E.; et al. Low doses of methylmercury exposure during adulthood in rats display oxidative stress, neurodegeneration in the motor cortex and lead to impairment of motor skills. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. 2019, 51, 19–27. [Google Scholar] [CrossRef]
- Freire, M.A.M.; Santana, L.N.S.; Bittencourt, L.O.; Nascimento, P.C.; Fernandes, R.M.; Leão, L.K.R.; Fernandes, L.M.P.; Silva, M.C.F.; Amado, L.L.; Gomes-Leal, W.; et al. Methylmercury intoxication and cortical ischemia: Pre-clinical study of their comorbidity. Ecotoxicol. Environ. Saf. 2019, 174, 557–565. [Google Scholar] [CrossRef]
- Nascimento, P.C.; Ferreira, M.K.M.; Balbinot, K.M.; Alves-Júnior, S.M.; Viana Pinheiro, J.J.; Silveira, F.M.; Martins, M.D.; Crespo-Lopez, M.E.; Lima, R.R. Methylmercury-Induced Toxicopathologic Findings in Salivary Glands of Offspring Rats After Gestational and Lactational Exposure. Biol. Trace Elem. Res. 2021, 199, 2983–2991. [Google Scholar] [CrossRef]
- Amado, L.L.; Garcia, M.L.; Ramos, P.B.; Freitas, R.F.; Zafalon, B.; Ferreira, J.L.R.; Yunes, J.S.; Monserrat, J.M. A method to measure total antioxidant capacity against peroxyl radicals in aquatic organisms: Application to evaluate microcystins toxicity. Sci. Total Environ. 2009, 407, 2115–2123. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Esterbauer, H.; Cheeseman, K.H. Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4-hydroxynonenal. Methods Enzymol. 1990, 186, 407–421. [Google Scholar] [CrossRef]
- Bittencourt, L.O.; Dionizio, A.; Nascimento, P.C.; Puty, B.; Leão, L.K.R.; Luz, D.A.; Silva, M.C.F.; Amado, L.L.; Leite, A.; Buzalaf, M.R.; et al. Proteomic approach underlying the hippocampal neurodegeneration caused by low doses of methylmercury after long-term exposure in adult rats. Met. Integr. Biometal Sci. 2019, 11, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Eiró, L.G.; Ferreira, M.K.M.; Bittencourt, L.O.; Aragão, W.A.B.; Souza, M.P.C.; Silva, M.C.F.; Dionizio, A.; Buzalaf, M.A.R.; Crespo-López, M.E.; Lima, R.R. Chronic methylmercury exposure causes spinal cord impairment: Proteomic modulation and oxidative stress. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2020, 146, 111772. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a Accession ID | Protein Name Description | PLGS Score | Fold Change |
|---|---|---|---|
| P34926 | Microtubule-associated protein 1A | 71.4 | 4.35 |
| P10888 | Cytochrome c oxidase subunit 4 isoform 1, mitochondrial | 146.14 | 1.46 |
| P0DP29 | Calmodulin-1 | 1825.33 | 1.39 |
| P0DP30 | Calmodulin-2 | 1825.33 | 1.39 |
| P0DP31 | Calmodulin-3 | 1867.79 | 1.38 |
| Q63345 | Myelin-oligodendrocyte glycoprotein | 290.08 | 1.30 |
| P21707 | Synaptotagmin-1 | 113.9 | 1.20 |
| P07825 | Synaptophysin | 286.54 | 1.16 |
| P68035 | Actin, alpha cardiac muscle 1 | 10,468.68 | −0.94 |
| P39052 | Dynamin-2 | 127.72 | −0.92 |
| P63039 | 60 kDa heat shock protein, mitochondrial | 870.92 | −0.91 |
| P08413 | Calcium/calmodulin-dependent protein kinase type II subunit beta | 450.44 | −0.90 |
| P11730 | Calcium/calmodulin-dependent protein kinase type II subunit gamma | 323.39 | −0.90 |
| P00406 | Cytochrome c oxidase subunit 2 | 786.54 | −0.89 |
| P0DMW0 | Heat shock 70 kDa protein 1A | 485.48 | −0.87 |
| Q06647 | ATP synthase subunit O, mitochondrial | 905.81 | −0.86 |
| P11275 | Calcium/calmodulin-dependent protein kinase type II subunit alpha | 220.44 | −0.85 |
| P12075 | Cytochrome c oxidase subunit 5B, mitochondrial | 942.79 | −0.84 |
| P15791 | Calcium/calmodulin-dependent protein kinase type II subunit delta | 215.3 | −0.84 |
| P60711 | Actin, cytoplasmic 1 | 18,625.63 | −0.83 |
| P35704 | Peroxiredoxin-2 | 2627.63 | −0.82 |
| P09951 | Synapsin-1 | 1081.01 | −0.82 |
| P02688 | Myelin basic protein | 13,414.88 | −0.76 |
| P82995 | Heat shock protein HSP 90-alpha | 1727.77 | −0.73 |
| P07895 | Superoxide dismutase [Mn], mitochondrial | 514.65 | −0.70 |
| Q5U300 | Ubiquitin-like modifier-activating enzyme 1 | 207.98 | −0.69 |
| P21575 | Dynamin-1 | 876.32 | −0.69 |
| Q08877 | Dynamin-3 | 202.61 | −0.69 |
| P31399 | ATP synthase subunit d, mitochondrial | 589.98 | −0.66 |
| P10719 | ATP synthase subunit beta, mitochondrial | 11,814.81 | −0.65 |
| O35244 | Peroxiredoxin-6 | 2447.06 | −0.64 |
| P31016 | Disks large homolog 4 | 86.82 | - |
| Q62671 | E3 ubiquitin-protein ligase UBR5 | 48.7 | - |
| O88600 | Heat shock 70 kDa protein 4 | 62.06 | - |
| Q66HA8 | Heat shock protein 105 kDa | 45.92 | - |
| Q63560 | Microtubule-associated protein 6 | 72.62 | - |
| P07722 | Myelin-associated glycoprotein | 55.31 | - |
| P34064 | Proteasome subunit alpha type-5 | 83.39 | - |
| P61959 | Small ubiquitin-related modifier 2 | 152.37 | - |
| Q5XIF4 | Small ubiquitin-related modifier 3 | 152.37 | - |
| B2RYG6 | Ubiquitin thioesterase OTUB1 | 121.2 | - |
| Q7M767 | Ubiquitin-conjugating enzyme E2 variant 2 | 735.2 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bittencourt, L.O.; Chemelo, V.S.; Aragão, W.A.B.; Puty, B.; Dionizio, A.; Teixeira, F.B.; Fernandes, M.S.; Silva, M.C.F.; Fernandes, L.M.P.; de Oliveira, E.H.C.; et al. From Molecules to Behavior in Long-Term Inorganic Mercury Intoxication: Unraveling Proteomic Features in Cerebellar Neurodegeneration of Rats. Int. J. Mol. Sci. 2022, 23, 111. https://doi.org/10.3390/ijms23010111
Bittencourt LO, Chemelo VS, Aragão WAB, Puty B, Dionizio A, Teixeira FB, Fernandes MS, Silva MCF, Fernandes LMP, de Oliveira EHC, et al. From Molecules to Behavior in Long-Term Inorganic Mercury Intoxication: Unraveling Proteomic Features in Cerebellar Neurodegeneration of Rats. International Journal of Molecular Sciences. 2022; 23(1):111. https://doi.org/10.3390/ijms23010111
Chicago/Turabian StyleBittencourt, Leonardo Oliveira, Victória Santos Chemelo, Walessa Alana Bragança Aragão, Bruna Puty, Aline Dionizio, Francisco Bruno Teixeira, Mileni Silva Fernandes, Márcia Cristina Freitas Silva, Luanna Melo Pereira Fernandes, Edivaldo Herculano Corrêa de Oliveira, and et al. 2022. "From Molecules to Behavior in Long-Term Inorganic Mercury Intoxication: Unraveling Proteomic Features in Cerebellar Neurodegeneration of Rats" International Journal of Molecular Sciences 23, no. 1: 111. https://doi.org/10.3390/ijms23010111