
Protective Effect of Polysaccharides Extracted from Cudrania tricuspidata Fruit against Cisplatin-Induced Cytotoxicity in Macrophages and a Mouse Model

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of CTPS
2.2. Effects of CTPS on Cisp-Stimulated Lung Cancer Cells and Macrophage Cytotoxicity
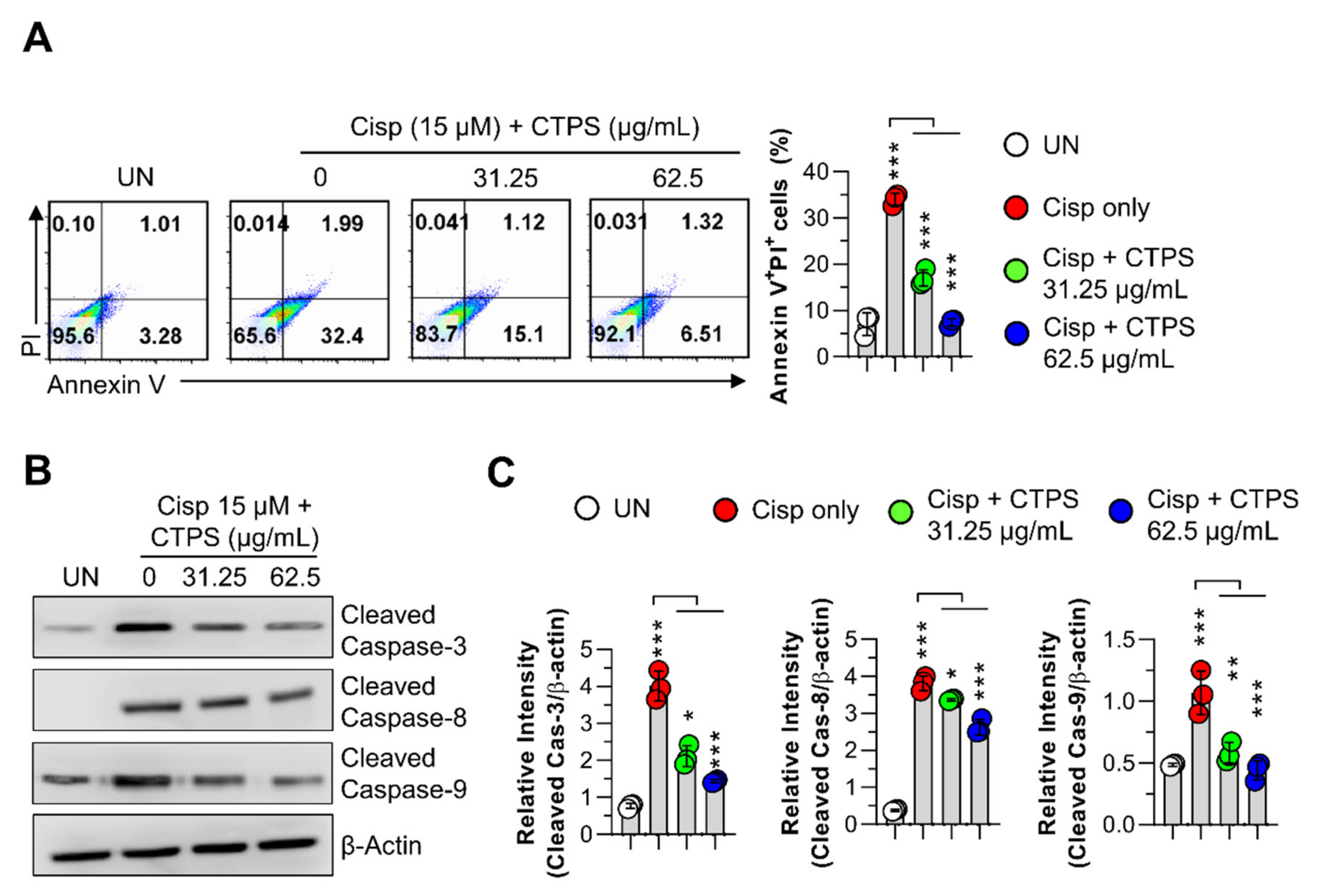
2.3. Inhibitory Effects of CTPS on Cisp-Stimulated Macrophage Apoptosis
2.4. Effects of CTPS on the Mitochondrial Apoptotic Pathway in Cisp-Treated Macrophages
2.5. Effects of CTPS on ROS Production and MTP Loss in Cisp-Treated Macrophages
2.6. Effects of CTPS on Cisp-Stimulated BMDM Cytotoxicity
2.7. Protective Effects of CTPS on Cisp-Induced Immunotoxicity in Mice with Lung Metastasis
2.8. Effects of CTPS on Serum Biochemical Markers in Mice with Lung Metastasis
3. Discussion
4. Materials and Methods
4.1. Preparation of CTPS
4.2. Analysis of Monosaccharide Composition and Acidic Polysaccharides of CTPS
4.3. Fourier Transform Infrared (FTIR) Spectroscopy
4.4. Reagents and Antibodies
4.5. Cell Culture
4.6. Cytotoxicity Analysis
4.7. Cell Migration Assay
4.8. Immunoblotting
4.9. Measurement of ROS Levels
4.10. Assessment of MTP
4.11. TUNEL Staining
4.12. Animals
4.13. Development of Lung Metastasis via B16BL6 Treatment
4.14. Analysis of Immunotoxicity in Splenic T Cells
4.15. Analysis of Biochemical Marker in Serum
4.16. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Akazawa, H. Cardiotoxicity of Cancer Chemotherapy—Mechanisms and Therapeutic Approach. Gan Kagaku Ryoho Cancer Chemother. 2017, 44, 2058–2063. [Google Scholar] [CrossRef]
- Vineis, P.; Wild, C.P. Global cancer patterns: Causes and prevention. Lancet 2014, 383, 549–557. [Google Scholar] [CrossRef]
- Ames, B.N.; Gold, L.S.; Willett, W.C. The Causes and Prevention of Cancer. Proc. Natl. Acad. Sci. USA 1995, 92, 5258–5265. [Google Scholar] [CrossRef] [Green Version]
- Varmus, H. The New Era in Cancer Research. Science 2006, 312, 1162–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Luo, H.; Zhang, W.; Shen, Z.; Hu, X.; Zhu, X. Molecular Mechanisms of Cisplatin Resistance in Cervical Cancer. Drug Des. Dev. Ther. 2016, 10, 1885–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufmann, S.H.; Earnshaw, W.C. Induction of Apoptosis by Cancer Chemotherapy. Exp. Cell Res. 2000, 256, 42–49. [Google Scholar] [CrossRef]
- DeVita, V.T., Jr.; Chu, E. A History of Cancer Chemotherapy. Cancer Res. 2008, 68, 8643–8653. [Google Scholar] [CrossRef] [Green Version]
- Galmarini, D.; Galmarini, C.M.; Galmarini, F.C. Cancer Chemotherapy: A Critical Analysis of Its 60 Years of History. Crit. Rev. Oncol./Hematol. 2012, 84, 181–199. [Google Scholar] [CrossRef]
- Khalaf, A.A.; Hussein, S.; Tohamy, A.F.; Marouf, S.; Yassa, H.D.; Zaki, A.R.; Bishayee, A. Protective effect of Echinacea Purpurea (Immulant) against Cisplatin-Induced Immunotoxicity in Rats. DARU J. Pharm. Sci. 2019, 27, 233–241. [Google Scholar] [CrossRef]
- Chen, R.L.; Wang, Z.; Huang, P.; Sun, C.H.; Yu, W.Y.; Zhang, H.H.; Yu, C.H.; He, J.Q. Isovitexin Potentiated the Antitumor Activity of Cisplatin by Inhibiting the Glucose Metabolism of Lung Cancer Cells and Reduced Cisplatin-Induced Immunotoxicity in Mice. Int. Immunopharmacol. 2021, 94, 107357. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.; Arvin, A. Chemotherapy-Induced Immunosuppression. Environ. Health Perspect. 1982, 43, 21–25. [Google Scholar] [CrossRef]
- Deng, X.; Luo, S.; Luo, X.; Hu, M.; Ma, F.; Wang, Y.; Zhou, L.; Huang, R. Fraction From Lycium barbarum Polysaccharides Reduces Immunotoxicity and Enhances Antitumor Activity of Doxorubicin in Mice. Integr. Cancer Ther. 2018, 17, 860–866. [Google Scholar] [CrossRef] [Green Version]
- Fulda, S. Modulation of apoptosis by natural products for cancer therapy. Planta Med. 2010, 76, 1075–1079. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Q.; Wang, X.L.; He, D.H.; Cheng, Y.X. Protection against Chemotherapy- and Radiotherapy-Induced Side Effects: A Review Based on the Mechanisms and Therapeutic Opportunities of Phytochemicals. Phytomedicine 2021, 80, 153402. [Google Scholar] [CrossRef]
- Fu, B.; Wang, N.; Tan, H.Y.; Li, S.; Cheung, F.; Feng, Y. Multi-Component Herbal Products in the Prevention and Treatment of Chemotherapy-Associated Toxicity and Side Effects: A Review on Experimental and Clinical Evidences. Front. Pharmacol. 2018, 9, 1394. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.Y.; Wang, F.X.; Jia, K.K.; Kong, L.D. Natural Product Interventions for Chemotherapy and Radiotherapy-Induced Side Effects. Front. Pharmacol. 2018, 9, 1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Y.S.; Yu, M.S.; Yang, X.F.; So, K.F.; Yuen, W.H.; Chang, R.C. Neuroprotective effects of polysaccharides from wolfberry, the fruits of Lycium barbarum, against homocysteine-induced toxicity in rat cortical neurons. J. Alzheimer’s Dis. 2010, 19, 813–827. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.X.; Wang, H.; Xu, H.S.; Ni, Y. Novel Polysaccharide Adjuvant from the Roots of Actinidia Eriantha with Dual Th1 and Th2 Potentiating Activity. Vaccine 2009, 27, 3984–3991. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.F.; Wan, L.L.; Peng, J.L.; Guo, C. Alleviation of the Acute Doxorubicin-Induced Cardiotoxicity by Lycium Barbarum Polysaccharides through the Suppression of Oxidative Stress. Food Chem. Toxicol. 2011, 49, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Fu, Y. Isolation, Purification, and Immunomodulatory Activity In Vitro of Three Polysaccharides from Roots of Cudrania Tricuspidata. Acta Biochim. Biophys. Sin. 2011, 43, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.; Hiep, N.T.; Kim, D.W.; Hong, S.; Guo, Y.; Hwang, B.Y.; Lee, H.J.; Mar, W.; Lee, D. Chemical Constituents Isolated from the Root Bark of Cudrania Tricuspidata and Their Potential Neuroprotective Effects. J. Nat. Prod. 2016, 79, 1938–1951. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.; Yoon, C.S.; Kim, K.W.; Lee, H.; Kim, N.; Woo, E.R.; Kim, Y.C.; Kang, D.G.; Lee, H.S.; Oh, H.; et al. Neuroprotective and Anti-Inflammatory Effects of Kuwanon C from Cudrania tricuspidata Are Mediated by Heme Oxygenase-1 in HT22 Hippocampal Cells, RAW264.7 Macrophage, and BV2 Microglia. Int. J. Mol. Sci. 2020, 21, 4839. [Google Scholar] [CrossRef]
- Dasari, S.; Tchounwou, P.B. Cisplatin in Cancer Therapy: Molecular Mechanisms of Action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on Mechanisms of In Vitro Antioxidant Activity of Polysaccharides. Oxid. Med. Cell. Longev. 2016, 2016, 5692852. [Google Scholar] [CrossRef] [Green Version]
- Arends, J.; Bachmann, P.; Baracos, V.; Barthelemy, N.; Bertz, H.; Bozzetti, F.; Fearon, K.; Hutterer, E.; Isenring, E.; Kaasa, S.; et al. ESPEN Guidelines on Nutrition in Cancer Patients. Clin. Nutr. 2017, 36, 11–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allemani, C.; Coleman, M.P. Public Health Surveillance of Cancer Survival in the United States and Worldwide: The Contribution of the CONCORD Programme. Cancer 2017, 123, 4977–4981. [Google Scholar] [CrossRef] [Green Version]
- Early Breast Cancer Trialists’ Collaborative Group. Effects of Chemotherapy and Hormonal Therapy for Early Breast Cancer on Recurrence and 15-Year Survival: An Overview of the Randomised Trials. Lancet 2005, 365, 1687–1717. [Google Scholar] [CrossRef]
- Patel, F.; Spassieva, S.D. Side Effects in Cancer Therapy: Are Sphingolipids to Blame? Adv. Cancer Res. 2018, 140, 367–388. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S. Cisplatin: The First Metal Based Anticancer Drug. Bioorg. Chem. 2019, 88, 102925. [Google Scholar] [CrossRef]
- Qi, L.; Luo, Q.; Zhang, Y.; Jia, F.; Zhao, Y.; Wang, F. Advances in Toxicological Research of the Anticancer Drug Cisplatin. Chem. Res. Toxicol. 2019, 32, 1469–1486. [Google Scholar] [CrossRef]
- Kim, H.M.; Song, Y.; Hyun, G.H.; Long, N.P.; Park, J.H.; Hsieh, Y.S.Y.; Kwon, S.W. Characterization and Antioxidant Activity Determination of Neutral and Acidic Polysaccharides from Panax Ginseng C. A. Meyer. Molecules 2020, 25, 791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Xue, X.; Zhang, Z. Structural, Physicochemical, Antioxidant and Antitumor Property of an Acidic Polysaccharide from Polygonum Multiflorum. Int. J. Biol. Macromol. 2017, 96, 494–500. [Google Scholar] [CrossRef]
- Wong, R.S. Apoptosis in Cancer: From Pathogenesis to Treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Goldar, S.; Khaniani, M.S.; Derakhshan, S.M.; Baradaran, B. Molecular Mechanisms of Apoptosis and Roles in Cancer Development and Treatment. Asian Pac. J. Cancer Prev. 2015, 16, 2129–2144. [Google Scholar] [CrossRef] [Green Version]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as Anticancer Mechanism: Function and Dysfunction of Its Modulators and Targeted Therapeutic Strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Ren, Y.; Li, X.; Zhang, X.; Guo, H.; Han, Y.; Hu, J. A Polysaccharide from Green Tea (Camellia Sinensis L.) Protects Human Retinal Endothelial Cells against Hydrogen Peroxide-Induced Oxidative Injury and Apoptosis. Int. J. Biol. Macromol. 2018, 115, 600–607. [Google Scholar] [CrossRef]
- Feki, A.; Ben Saad, H.; Bkhairia, I.; Ktari, N.; Naifar, M.; Boudawara, O.; Droguet, M.; Magne, C.; Nasri, M.; Ben Amara, I. Cardiotoxicity and Myocardial Infarction-Associated DNA Damage Induced by Thiamethoxam In Vitro and In Vivo: Protective Role of Trigonella Foenum-Graecum Seed-Derived Polysaccharide. Environ. Toxicol. 2019, 34, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Byun, E.B.; Cho, E.J.; Kim, Y.E.; Kim, W.S.; Byun, E.H. Neuroprotective Effect of Polysaccharide Separated from Perilla Frutescens Britton Var. Acuta Kudo against H2O2-Induced Oxidative Stress in HT22 Hippocampus Cells. Biosci. Biotechnol. Biochem. 2018, 82, 1344–1358. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Li, X.B.; Hou, S.G.; Sun, Y.; Shi, Y.R.; Lin, S.S. Cedrol Induces Autophagy and Apoptotic Cell Death in A549 Non-Small Cell Lung Carcinoma Cells through the P13K/Akt Signaling Pathway, the Loss of Mitochondrial Transmembrane Potential and the Generation of ROS. Int. J. Mol. Med. 2016, 38, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Park, K.R.; Nam, D.; Yun, H.M.; Lee, S.G.; Jang, H.J.; Sethi, G.; Cho, S.K.; Ahn, K.S. Beta-Caryophyllene Oxide Inhibits Growth and Induces Apoptosis through the Suppression of PI3K/AKT/mTOR/S6K1 Pathways and ROS-Mediated MAPKs Activation. Cancer Lett. 2011, 312, 178–188. [Google Scholar] [CrossRef]
- Byun, E.B.; Kim, H.M.; Song, H.Y.; Kim, W.S. Hesperidin Structurally Modified by Gamma Irradiation Induces Apoptosis in Murine Melanoma B16BL6 Cells and Inhibits both Subcutaneous Tumor Growth and Metastasis in C57BL/6 Mice. Food Chem. Toxicol. 2019, 127, 19–30. [Google Scholar] [CrossRef]
- Nie, Z.; Deng, S.; Zhang, L.; Chen, S.; Lu, Q.; Peng, H. Crocin Protects Against Dexamethasoneinduced Osteoblast Apoptosis by Inhibiting the ROS/Ca2+Mediated Mitochondrial Pathway. Mol. Med. Rep. 2019, 20, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Joy, J.; Nair, C.K. Amelioration of Cisplatin Induced Nephrotoxicity in Swiss Albino Mice by Rubia Cordifolia Extract. J. Cancer Res. Ther. 2008, 4, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jung, S.Y.; Yang, K.J.; Kim, Y.; Lee, D.; Lee, M.H.; Kim, D.K. Alpha-Lipoic Acid Prevents against Cisplatin Cytotoxicity via Activation of the NRF2/HO-1 Antioxidant Pathway. PLoS ONE 2019, 14, e0226769. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Miao, J.; Liu, W.; Peng, L.; Chen, Y.; Zhong, Q. Formononetin Protects against Cisplatininduced Acute Kidney Injury through Activation of the PPARalpha/Nrf2/HO1/NQO1 Pathway. Int. J. Mol. Med. 2021, 47, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Bitter, T.; Muir, H.M. A Modified Uronic acid Carbazole reaction. Anal. Biochem. 1962, 4, 330–334. [Google Scholar] [CrossRef]
- Jungblut, M.; Oeltze, K.; Zehnter, I.; Hasselmann, D.; Bosio, A. Preparation of Single-Cell Suspensions from Mouse Spleen with the gentlemacs Dissociator. J. Vis. Exp. 2008, 10, e1029. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Contents (g/100 g) |
|---|---|
| Fucose | 0.004 ± 0.0048 |
| Glucosamine | 0.081 ± 0.0015 |
| Glucose | 17.454 ± 0.1550 |
| Mannose | 0.240 ± 0.0005 |
| Acidic polysaccharides | 42.325 ± 0.7889 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byun, E.-B.; Song, H.-Y.; Kim, W.S.; Han, J.M.; Seo, H.S.; Park, S.-H.; Kim, K.; Byun, E.-H. Protective Effect of Polysaccharides Extracted from Cudrania tricuspidata Fruit against Cisplatin-Induced Cytotoxicity in Macrophages and a Mouse Model. Int. J. Mol. Sci. 2021, 22, 7512. https://doi.org/10.3390/ijms22147512
Byun E-B, Song H-Y, Kim WS, Han JM, Seo HS, Park S-H, Kim K, Byun E-H. Protective Effect of Polysaccharides Extracted from Cudrania tricuspidata Fruit against Cisplatin-Induced Cytotoxicity in Macrophages and a Mouse Model. International Journal of Molecular Sciences. 2021; 22(14):7512. https://doi.org/10.3390/ijms22147512
Chicago/Turabian StyleByun, Eui-Baek, Ha-Yeon Song, Woo Sik Kim, Jeong Moo Han, Ho Seong Seo, Sang-Hyun Park, Kwangwook Kim, and Eui-Hong Byun. 2021. "Protective Effect of Polysaccharides Extracted from Cudrania tricuspidata Fruit against Cisplatin-Induced Cytotoxicity in Macrophages and a Mouse Model" International Journal of Molecular Sciences 22, no. 14: 7512. https://doi.org/10.3390/ijms22147512