A Novel Sulfonamide, 4-FS, Reduces Ethanol Drinking and Physical Withdrawal Associated With Ethanol Dependence

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
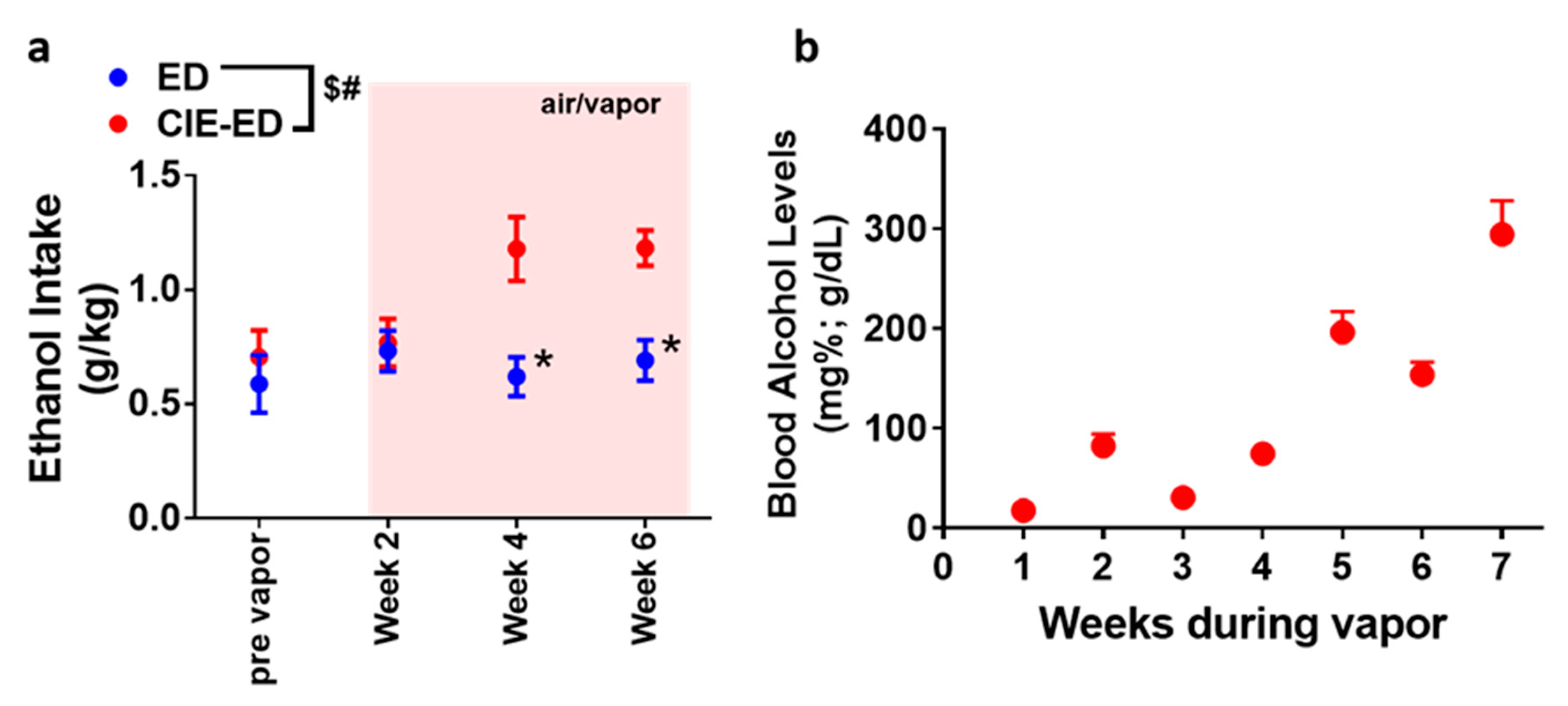
2.1. CIE-ED Rats Consume More Ethanol Than ED Rats
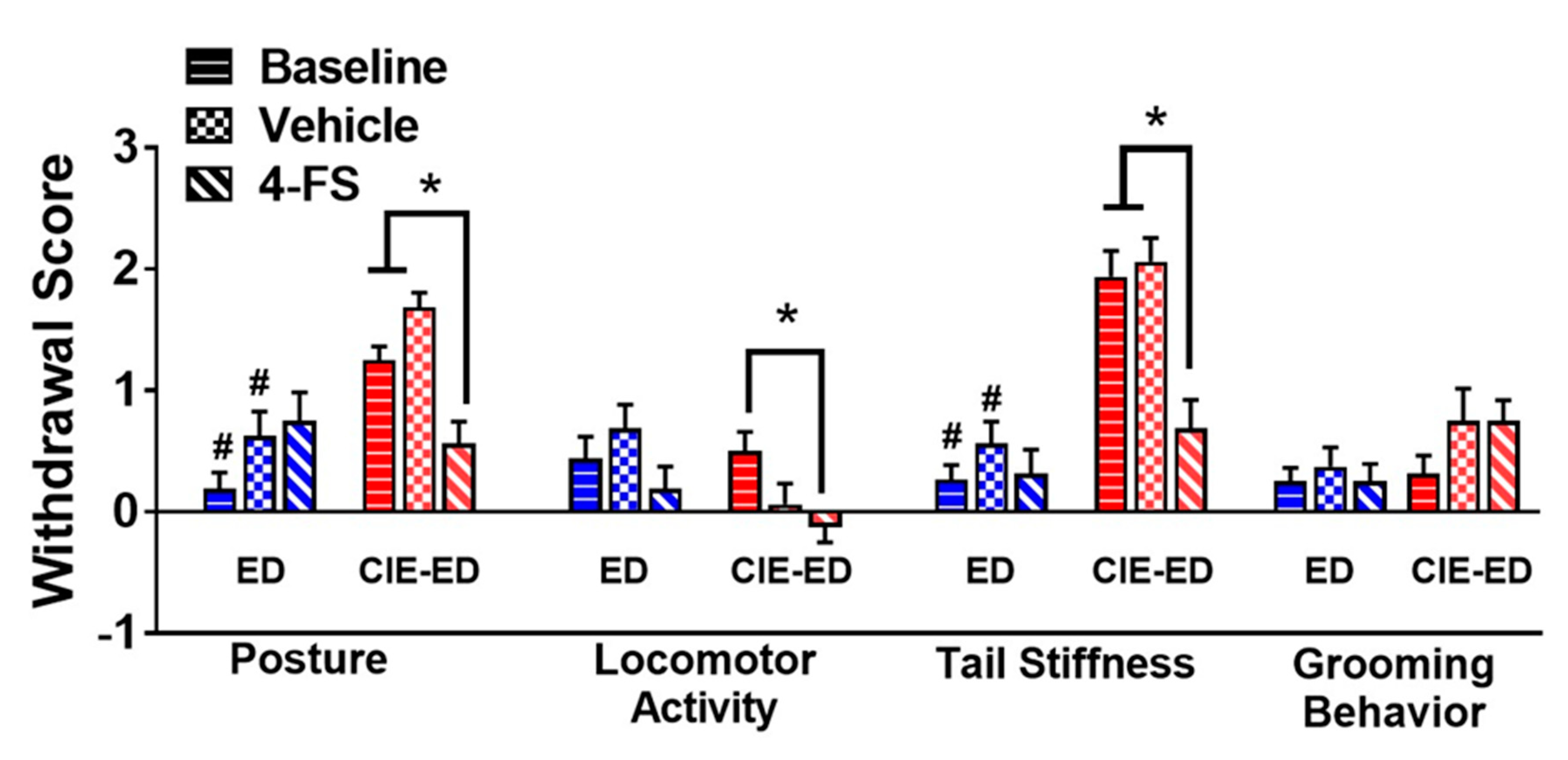
2.2. CIE-ED Rats Demonstrate Somatic Withdrawal Symptoms
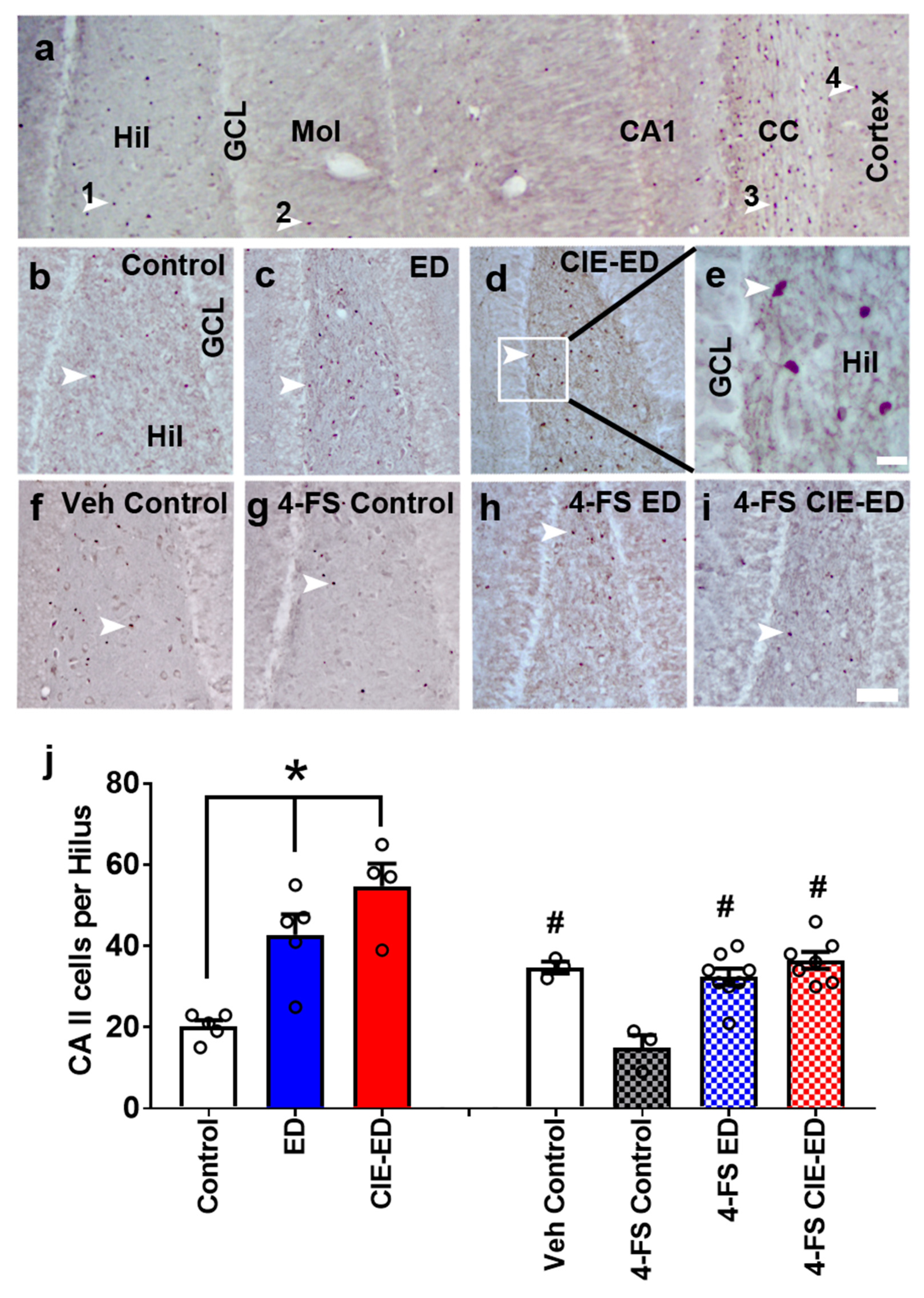
2.3. ED and CIE-ED Rats Show Increased Number of CA II Immunoreactive Cells in the Hilus and 4-FS Reduces This Effect
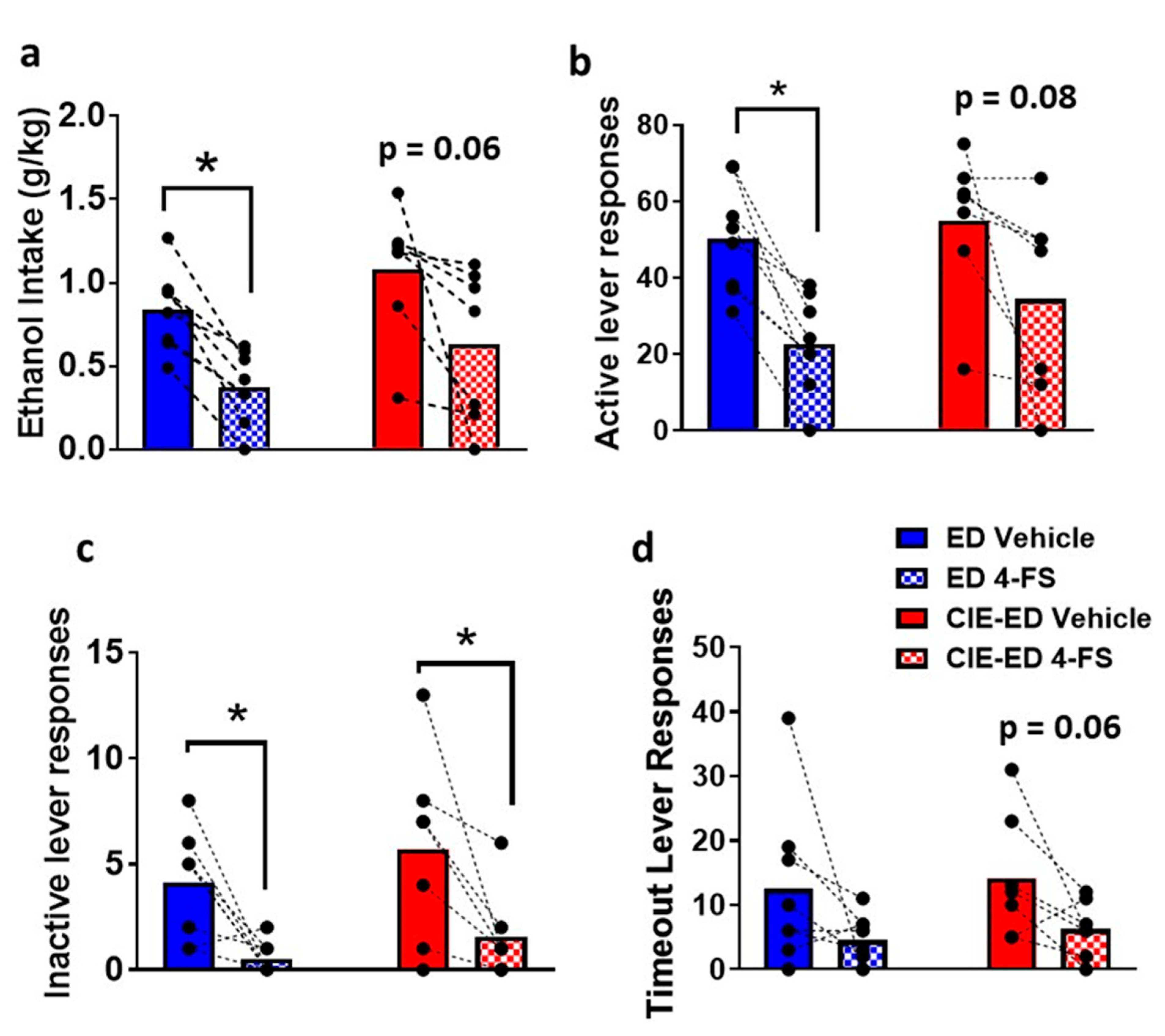
2.4. 4-FS reduces Withdrawal Behavior in CIE-ED Rats and Reduces Drinking in ED Rats
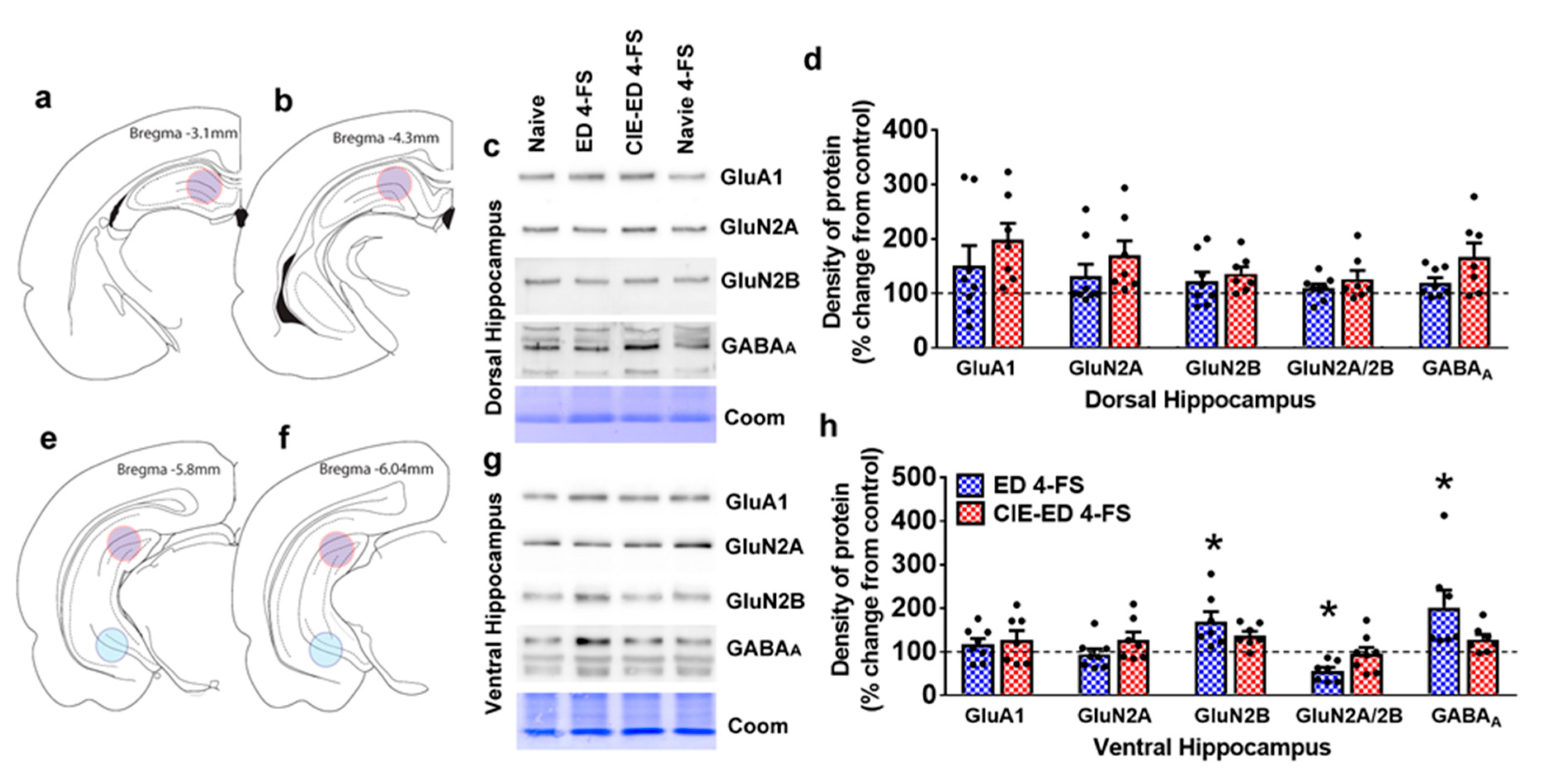
2.5. 4-FS Alters Expression of GluRs and GABAARs in the Ventral Hippocampus
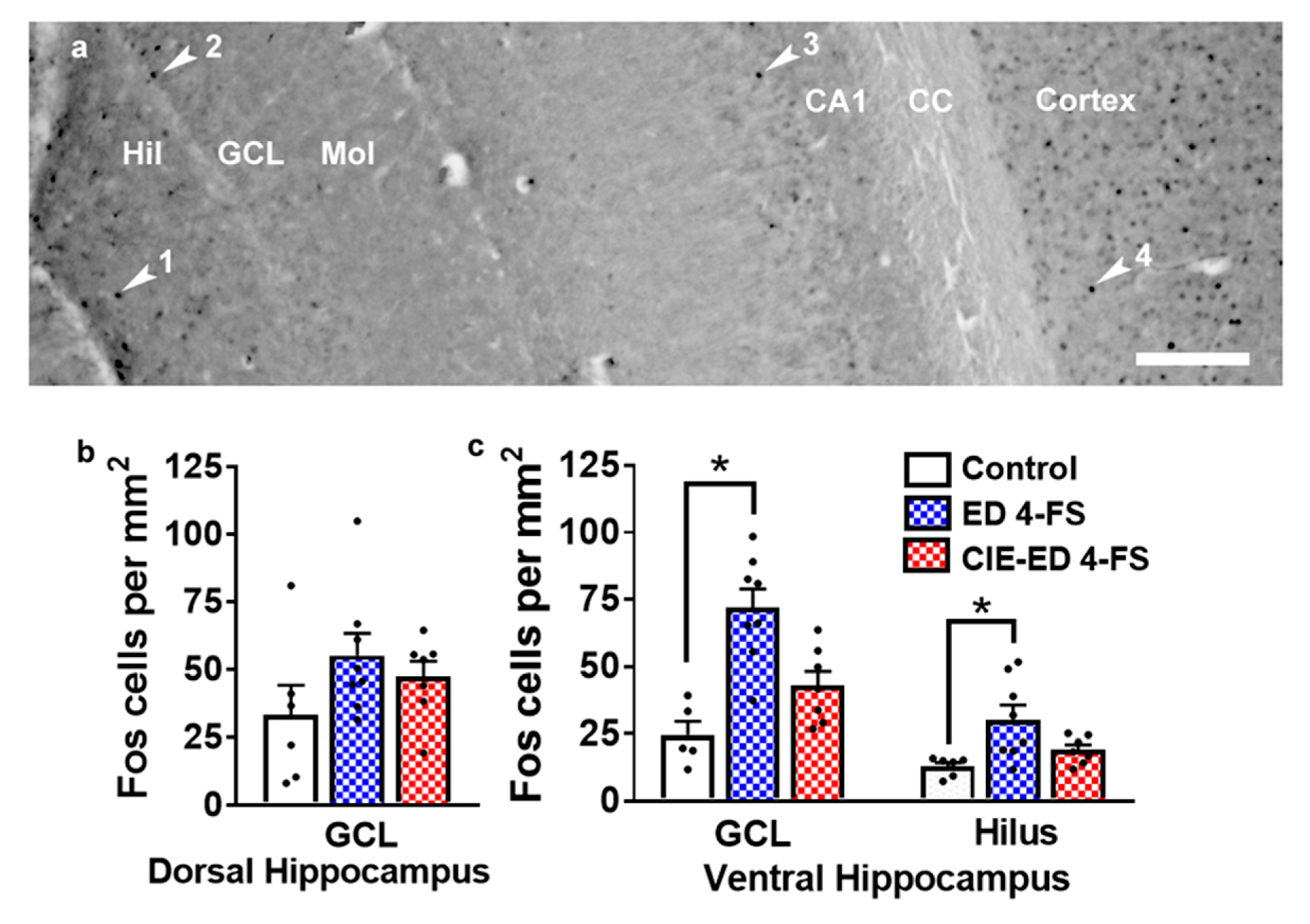
2.6. 4-FS Alters the Number of Fos Cells in the Ventral Hippocampus
3. Discussion
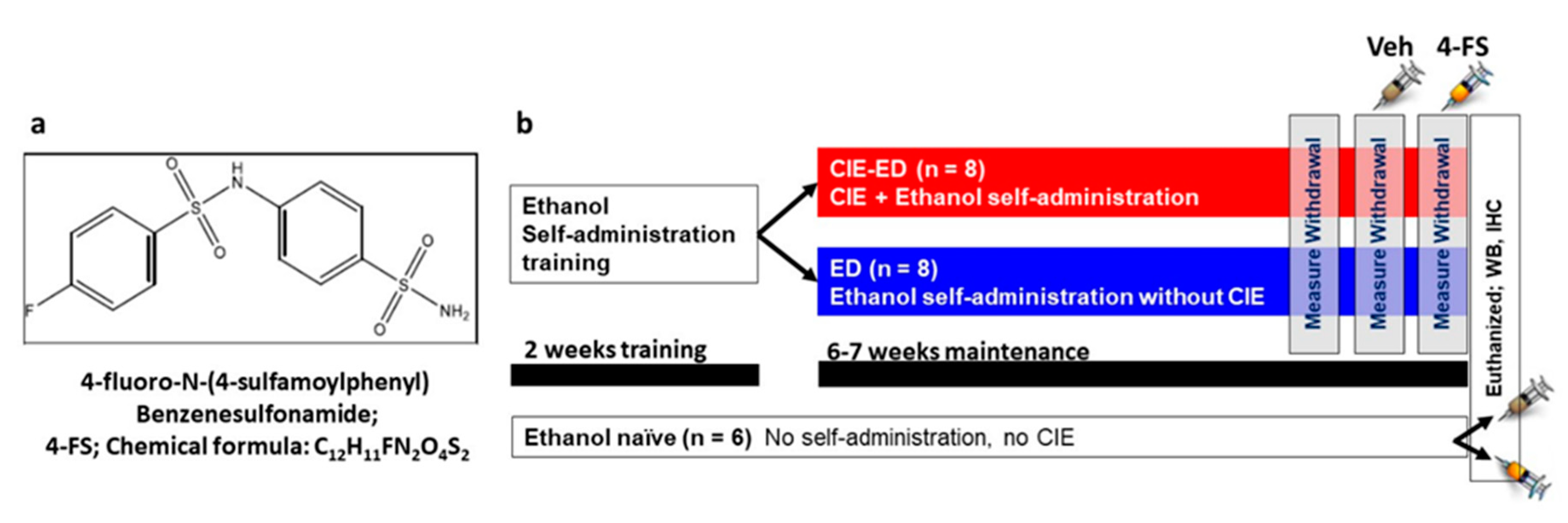
4. Materials and Methods
4.1. Animals
4.2. Ethanol Self-Administration
4.3. Tail Bleeding for Determination of Blood Alcohol Levels (BAL)
4.4. 4-FS Synthesis and Treatment
4.5. Scoring of Physical Withdrawal
4.6. Brain Tissue Collection
4.7. Western Blotting
4.8. Immunohistochemistry and Quantitative Analysis of Carbonic Anhydrase II (CA II) and Fos Labeled Cells
4.9. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- SAMHSA. Results from the 2012 National Survey on Drug Use and Health: Summary of National Findings. Available online: https://store.samhsa.gov/product/Results-from-the-2012-National-Survey-on-Drug-Use-and-Health-NSDUH-/SMA13-4795 (accessed on 20 June 2020).
- Sullivan, E.V.; Rosenbloom, M.J.; Lim, K.O.; Pfefferbaum, A. Longitudinal changes in cognition, gait, and balance in abstinent and relapsed alcoholic men: Relationships to changes in brain structure. Neuropsychology 2000, 14, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Bechara, A.; Dolan, S.; Denburg, N.; Hindes, A.; Anderson, S.W.; Nathan, P.E. Decision-making deficits, linked to a dysfunctional ventromedial prefrontal cortex, revealed in alcohol and stimulant abusers. Neuropsychologia 2001, 39, 376–389. [Google Scholar] [CrossRef]
- Oscar-Berman, M.; Marinkovic, K. Alcoholism and the brain: An overview. Alcohol. Res. Health 2003, 27, 125–133. [Google Scholar] [PubMed]
- Rosenbloom, M.; Sullivan, E.V.; Pfefferbaum, A. Using magnetic resonance imaging and diffusion tensor imaging to assess brain damage in alcoholics. Alcohol Res. Health 2003, 27, 146–152. [Google Scholar]
- Sullivan, E.V.; Rosenbloom, M.J.; Pfefferbaum, A. Pattern of motor and cognitive deficits in detoxified alcoholic men. Alcohol Clin. Exp. Res. 2000, 24, 611–621. [Google Scholar] [CrossRef]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [Green Version]
- Crews, F.T.; Boettiger, C.A. Impulsivity, frontal lobes and risk for addiction. Pharmacol. Biochem. Behav. 2009, 93, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Crews, F.T.; Nixon, K. Mechanisms of neurodegeneration and regeneration in alcoholism. Alcohol Alcohol 2009, 44, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Stephens, D.N.; Duka, T. Review. Cognitive and emotional consequences of binge drinking: Role of amygdala and prefrontal cortex. Philos. Trans. R. Soc. Lond. Biol. Sci. 2008, 363, 3169–3179. [Google Scholar] [CrossRef]
- O’Dell, L.E.; Roberts, A.J.; Smith, R.T.; Koob, G.F. Enhanced alcohol self-administration after intermittent versus continuous alcohol vapor exposure. Alcohol Clin. Exp. Res. 2004, 28, 1676–1682. [Google Scholar] [CrossRef]
- Valdez, G.R.; Roberts, A.J.; Chan, K.; Davis, H.; Brennan, M.; Zorrilla, E.P.; Koob, G.F. Increased ethanol self-administration and anxiety-like behavior during acute ethanol withdrawal and protracted abstinence: Regulation by corticotropin-releasing factor. Alcohol Clin. Exp. Res. 2002, 26, 1494–1501. [Google Scholar] [CrossRef] [PubMed]
- Hansson, A.C.; Nixon, K.; Rimondini, R.; Damadzic, R.; Sommer, W.H.; Eskay, R.; Crews, F.T.; Heilig, M. Long-term suppression of forebrain neurogenesis and loss of neuronal progenitor cells following prolonged alcohol dependence in rats. Int. J. Neuropsychopharmacol. 2010, 13, 583–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pian, J.P.; Criado, J.R.; Milner, R.; Ehlers, C.L. N-methyl-D-aspartate receptor subunit expression in adult and adolescent brain following chronic ethanol exposure. Neuroscience. 2010, 170, 645–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Criado, J.R.; Liu, T.; Ehlers, C.L.; Mathe, A.A. Prolonged chronic ethanol exposure alters neuropeptide Y and corticotropin-releasing factor levels in the brain of adult Wistar rats. Pharmacol. Biochem. Behav. 2011, 99, 104–111. [Google Scholar] [CrossRef] [Green Version]
- Vendruscolo, L.F.; Barbier, E.; Schlosburg, J.E.; Misra, K.K.; Whitfield, T.W.; Logrip, M.L., Jr.; Rivier, C.; Repunte-Canonigo, V.; Zorrilla, E.P.; Sanna, P.P.; et al. Corticosteroid-dependent plasticity mediates compulsive alcohol drinking in rats. J. Neurosci. 2012, 32, 7563–7571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, H.N.; Chan, S.H.; Crawford, E.F.; Lee, Y.K.; Funk, C.K.; Koob, G.F.; Mandyam, C.D. Permanent impairment of birth and survival of cortical and hippocampal proliferating cells following excessive drinking during alcohol dependence. Neurobiol. Dis. 2009, 36, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staples, M.C.; Kim, A.; Mandyam, C.D. Dendritic remodeling of hippocampal neurons is associated with altered NMDA receptor expression in alcohol dependent rats. Mol. Cell Neurosci. 2015, 65, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Roberto, M.; Varodayan, F.P. Synaptic targets: Chronic alcohol actions. Neuropharmacology 2017, 122, 85–99. [Google Scholar] [CrossRef]
- Somkuwar, S.S.; Fannon, M.J.; Staples, M.C.; Zamora-Martinez, E.R.; Navarro, A.I.; Kim, A.; Quigley, J.A.; Edwards, S.; Mandyam, C.D. Alcohol dependence-induced regulation of the proliferation and survival of adult brain progenitors is associated with altered BDNF-TrkB signaling. Brain Struct. Funct. 2016, 221, 4319–4335. [Google Scholar] [CrossRef] [Green Version]
- Mandyam, C.D. The Interplay between the Hippocampus and Amygdala in Regulating Aberrant Hippocampal Neurogenesis during Protracted Abstinence from Alcohol Dependence. Front. Psychiatry 2013, 4, 61. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, M.A.; Harris, B.R.; Mullholland, P.J.; Blanchard, J.A.; 2nd Gibson, D.A.; Holley, R.C.; Littleton, J.M. Hippocampal CA1 region neurodegeneration produced by ethanol withdrawal requires activation of intrinsic polysynaptic hippocampal pathways and function of N-methyl-D-aspartate receptors. Neuroscience 2004, 124, 869–877. [Google Scholar] [CrossRef]
- Mulholland, P.J.; Harris, B.R.; Wilkins, L.H.; Self, R.L.; Blanchard, J.A.; Holley, R.C.; Littleton, J.M.; Prendergast, M.A. Opposing effects of ethanol and nicotine on hippocampal calbindin-D28k expression. Alcohol 2003, 31, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, L.H.; Prendergast, M.A., Jr.; Blanchard, J.; Holley, R.C.; Chambers, E.R.; Littleton, J.M. Potential value of changes in cell markers in organotypic hippocampal cultures associated with chronic EtOH exposure and withdrawal: Comparison with NMDA-induced changes. Alcohol Clin. Exp. Res. 2006, 30, 1768–1880. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.A.; Harris, B.R.; Prendergast, M.A.; Hart, S.R.; Blanchard, J.A., 2nd; Holley, R.C.; Pedigo, N.W.; Littleton, J.M. Polyamines contribute to ethanol withdrawal-induced neurotoxicity in rat hippocampal slice cultures through interactions with the NMDA receptor. Alcohol Clin. Exp. Res. 2003, 27, 1099–1106. [Google Scholar] [CrossRef]
- Claus, D.; Kim, J.S.; Kornhuber, M.E.; Ahn, Y.S. [Effect of ethanol on the neurotransmitters glutamate and GABA]. Arch. Psychiatr. Nervenkr. 1982, 232, 183–189. [Google Scholar] [CrossRef]
- Keller, E.; Cummins, J.T.; von Hungen, K. Regional effects of ethanol on glutamate levels, uptake and release in slice and synaptosome preparations from rat brain. Subst. Alcohol Actions. Misuse 1983, 4, 383–392. [Google Scholar] [PubMed]
- Wilce, P.A.; Le, F.; Matsumoto, I.; Shanley, B.C. Ethanol inhibits NMDA-receptor mediated regulation of immediate early gene expression. Alcohol Alcohol Suppl. 1993, 2, 359–363. [Google Scholar]
- Snell, L.D.; Nunley, K.R.; Lickteig, R.L.; Browning, M.D.; Tabakoff, B.; Hoffman, P.L. Regional and subunit specific changes in NMDA receptor mRNA and immunoreactivity in mouse brain following chronic ethanol ingestion. Brain Res. Mol. Brain Res. 1996, 40, 71–78. [Google Scholar] [CrossRef]
- Wirkner, K.; Poelchen, W.; Koles, L.; Muhlberg, K.; Scheibler, P.; Allgaier, C.; Illes, P. Ethanol-induced inhibition of NMDA receptor channels. Neurochem. Int. 1999, 35, 153–162. [Google Scholar] [CrossRef]
- Davidson, M.; Shanley, B.; Wilce, P. Increased NMDA-induced excitability during ethanol withdrawal: A behavioural and histological study. Brain Res. 1995, 674, 91–96. [Google Scholar] [CrossRef]
- Davidson, M.D.; Wilce, P.; Shanley, B.C. Increased sensitivity of the hippocampus in ethanol-dependent rats to toxic effect of N-methyl-D-aspartic acid in vivo. Brain Res. 1993, 606, 5–9. [Google Scholar] [CrossRef]
- Hoffman, P.L. NMDA receptors in alcoholism. Int. Rev. Neurobiol. 2003, 56, 35–82. [Google Scholar] [PubMed]
- Chesler, M. Regulation and modulation of pH in the brain. Physiol. Rev. 2003, 83, 1183–1221. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Chesler, M. pH transients evoked by excitatory synaptic transmission are increased by inhibition of extracellular carbonic anhydrase. Proc. Natl. Acad. Sci. USA 1992, 89, 7786–7790. [Google Scholar] [CrossRef] [Green Version]
- Hamidi, S.; Avoli, M. Carbonic anhydrase inhibition by acetazolamide reduces in vitro epileptiform synchronization. Neuropharmacology 2015, 95, 377–387. [Google Scholar] [CrossRef] [Green Version]
- De Simone, G.; Scozzafava, A.; Supuran, C.T. Which carbonic anhydrases are targeted by the antiepileptic sulfonamides and sulfamates? Chem. Biol. Drug Des. 2009, 74, 317–321. [Google Scholar] [CrossRef]
- Lombroso, C.T.; Forxythe, I. A long-term follow-up of acetazolamide (diamox) in the treatment of epilepsy. Epilepsia 1960, 1, 493–500. [Google Scholar] [CrossRef]
- Van Berkel, M.A.; Elefritz, J.L. Evaluating off-label uses of acetazolamide. Off. J. Am. Soc. Health-Syst. Pharm. 2018, 75, 524–531. [Google Scholar] [CrossRef]
- Hamada, K.; Song, H.K.; Ishida, S.; Yagi, K.; Seino, M. Contrasting effects of zonisamide and acetazolamide on amygdaloid kindling in rats. Epilepsia 2001, 42, 1379–1386. [Google Scholar] [CrossRef] [Green Version]
- Guglielmo, R.; Martinotti, G.; Quatrale, M.; Ioime, L.; Kadilli, I.; Di Nicola, M.; Janiri, L. Topiramate in Alcohol Use Disorders: Review and Update. CNS Drugs 2015, 29, 383–395. [Google Scholar] [CrossRef]
- Santos, L.E.C.; Rodrigues, A.M.; Lopes, M.R.; Costa, V.D.C.; Scorza, C.A.; Scorza, F.A.; Cavalheiro, E.A.; Almeida, A.G. Long-term alcohol exposure elicits hippocampal nonsynaptic epileptiform activity changes associated with expression and functional changes in NKCC1, KCC2 co-transporters and Na(+)/K(+)-ATPase. Neuroscience 2017, 340, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Al-Rashida, M.; Ejaz, S.A.; Ali, S.; Shaukat, A.; Hamayoun, M.; Ahmed, M.; Iqbal, J. Diarylsulfonamides and their bioisosteres as dual inhibitors of alkaline phosphatase and carbonic anhydrase: Structure activity relationship and molecular modelling studies. Bioorg. Med. Chem. 2015, 23, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Ozsoy, H.Z. Carbonic anhydrase enzymes: Likely targets for inhalational anesthetics. Med. Hypotheses 2019, 123, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Carbonic anhydrase activators. Future Med. Chem. 2018, 10, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; An, L.; Duan, Q.; Ferraris Araneta, M.; Johnson, C.S.; Shen, J. Determining the Rate of Carbonic Anhydrase Reaction in the Human Brain. Sci. Rep. 2018, 8, 2328. [Google Scholar] [CrossRef] [Green Version]
- Tong, C.K.; Cammer, W.; Chesler, M. Activity-dependent pH shifts in hippocampal slices from normal and carbonic anhydrase II-deficient mice. Glia 2000, 31, 125–130. [Google Scholar] [CrossRef]
- Giacobini, E. A cytochemical study of the localization of carbonic anhydrase in the nervous system. J. Neurochem. 1962, 9, 169–177. [Google Scholar] [CrossRef]
- Nogradi, A.; Mihaly, A. Distribution of carbonic anhydrase activity in the rat central nervous system, as revealed by a new semipermeable technique. Acta Histochem. 1988, 84, 153–162. [Google Scholar] [CrossRef]
- Tong, C.K.; Chen, K.; Chesler, M. Kinetics of activity-evoked pH transients and extracellular pH buffering in rat hippocampal slices. J. Neurophysiol. 2006, 95, 3686–3697. [Google Scholar] [CrossRef]
- Walz, W. pH shifts evoked by neuronal stimulation in slices of rat hippocampus. Can. J. Physiol. Pharmacol. 1989, 67, 577–581. [Google Scholar] [CrossRef]
- Esbaugh, A.J.; Tufts, B.L. The structure and function of carbonic anhydrase isozymes in the respiratory system of vertebrates. Respir. Physiol. Neurobiol. 2006, 154, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Chesler, M.; Kaila, K. Modulation of pH by neuronal activity. Trends Neurosci. 1992, 15, 396–402. [Google Scholar] [CrossRef]
- Chesler, M.; Chen, J.C.; Kraig, R.P. Determination of extracellular bicarbonate and carbon dioxide concentrations in brain slices using carbonate and pH-selective microelectrodes. J. Neurosci. Methods 1994, 53, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Tong, C.K.; Chesler, M. Endogenous pH shifts facilitate spreading depression by effect on NMDA receptors. J. Neurophysiol. 1999, 81, 1988–1991. [Google Scholar] [CrossRef]
- Wagner, N.; Franz, N.; Dieteren, S.; Perl, M.; Mors, K.; Marzi, I.; Relja, B. Acute Alcohol Binge Deteriorates Metabolic and Respiratory Compensation Capability After Blunt Chest Trauma Followed by Hemorrhagic Shock-A New Research Model. Alcohol Clin. Exp. Res. 2017, 41, 1559–1567. [Google Scholar] [CrossRef]
- Gilliam, D.M.; Collins, A.C. Acute ethanol effects on blood pH, PCO2, and PO2 in LS and SS mice. Physiol. Behav. 1982, 28, 879–883. [Google Scholar] [CrossRef]
- Wang, T.; Eskandari, D.; Zou, D.; Grote, L.; Hedner, J. Increased Carbonic Anhydrase Activity is Associated with Sleep Apnea Severity and Related Hypoxemia. Sleep 2015, 38, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.T.; Lee, P.; Hui, D.; Michaelis, E.K.; Choi, I.Y. Effects of Ethanol Exposure on the Neurochemical Profile of a Transgenic Mouse Model with Enhanced Glutamate Release Using In Vivo (1)H MRS. Neurochem. Res. 2019, 44, 133–146. [Google Scholar] [CrossRef]
- Chen, J.C.; Chesler, M. Modulation of extracellular pH by glutamate and GABA in rat hippocampal slices. J. Neurophysiol. 1992, 67, 29–36. [Google Scholar] [CrossRef]
- Mendelson, J.H.; Mello, N.K. Basic mechanisms underlying physical dependence upon alcohol. Ann. N. Y. Acad. Sci. 1978, 311, 69–79. [Google Scholar] [CrossRef]
- Roelofs, S.M.; Dikkenberg, G.M. Hyperventilation and anxiety: Alcohol withdrawal symptoms decreasing with prolonged abstinence. Alcohol 1987, 4, 215–220. [Google Scholar] [CrossRef]
- Teng, Y.H.; Tsai, H.T.; Hsieh, Y.S.; Chen, Y.C.; Lin, C.W.; Lee, M.C.; Lin, L.Y.; Yang, S.F. Elevated erythrocyte carbonic anhydrase activity is a novel clinical marker in hyperventilation syndrome. Clin. Chem. Lab. Med 2009, 47, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.W.; Liang, J. Role of GABAA receptors in alcohol use disorders suggested by chronic intermittent ethanol (CIE) rodent model. Mol. Brain. 2017, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Nelson, T.E.; Ur, C.L.; Gruol, D.L. Chronic intermittent ethanol exposure enhances NMDA-receptor-mediated synaptic responses and NMDA receptor expression in hippocampal CA1 region. Brain Res. 2005, 1048, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Frasca, A.; Aalbers, M.; Frigerio, F.; Fiordaliso, F.; Salio, M.; Gobbi, M.; Cagnotto, A.; Gardoni, F.; Battaglia, G.S.; Hoogland, G.; et al. Misplaced NMDA receptors in epileptogenesis contribute to excitotoxicity. Neurobiol. Dis. 2011, 43, 507–515. [Google Scholar] [CrossRef]
- Smith, M.A.; Weiss, S.R.; Berry, R.L.; Zhang, L.X.; Clark, M.; Massenburg, G.; Post, R.M. Amygdala-kindled seizures increase the expression of corticotropin-releasing factor (CRF) and CRF-binding protein in GABAergic interneurons of the dentate hilus. Brain Res. 1997, 745, 248–256. [Google Scholar] [CrossRef]
- Macey, D.J.; Schulteis, G.; Heinrichs, S.C.; Koob, G.F. Time-dependent quantifiable withdrawal from ethanol in the rat: Effect of method of dependence induction. Alcohol 1996, 13, 163–170. [Google Scholar] [CrossRef]
- Prior, P.L.; Galduroz, J.C. Glutamatergic hyperfunctioning during alcohol withdrawal syndrome: Therapeutic perspective with zinc and magnesium. Med. Hypotheses 2011, 77, 368–370. [Google Scholar] [CrossRef] [Green Version]
- McCool, B.A.; Christian, D.T.; Diaz, M.R.; Lack, A.K. Glutamate plasticity in the drunken amygdala: The making of an anxious synapse. Int. Rev. Neurobiol. 2010, 91, 205–233. [Google Scholar]
- Hoffman, P.L.; Tabakoff, B. The role of the NMDA receptor in ethanol withdrawal. Exs 1994, 71, 61–70. [Google Scholar]
- Tsai, G.; Gastfriend, D.R.; Coyle, J.T. The glutamatergic basis of human alcoholism. Am. J. Psychiatry 1995, 152, 332–340. [Google Scholar] [PubMed]
- Baram, T.Z.; Hirsch, E.; Snead, O.C., III; Schultz, L. Corticotropin-releasing hormone-induced seizures in infant rats originate in the amygdala. Ann. Neurol. 1992, 31, 488–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapp, C.M.; Mercado, M.; Markley, T.L.; Crosby, S.; Ciraulo, D.A.; Kornetsky, C. Zonisamide decreases ethanol intake in rats and mice. Pharmacol. Biochem. Behav. 2007, 87, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalewska-Kaszubska, J.; Bajer, B.; Gorska, D.; Andrzejczak, D.; Dyr, W.; Bieńkowski, P. Effect of repeated treatment with topiramate on voluntary alcohol intake and beta-endorphin plasma level in Warsaw alcohol high-preferring rats. Psychopharmacology 2013, 225, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Knapp, C.M.; Sarid-Segal, O.; Richardson, M.A.; Colaneri, L.S.; Afshar, M.; Devine, E.; Streeter, C.C.; Piechniczek-Buczek, J.; Ciraulo, D.A. Open label trial of the tolerability and efficacy of zonisamide in the treatment of alcohol dependence. Am. J. Drug Alcohol Abus. 2010, 36, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Perucca, E. Clinical pharmacology and therapeutic use of the new antiepileptic drugs. Fundam. Clin. Pharmacol. 2001, 15, 405–417. [Google Scholar] [CrossRef]
- Rubio, G.; López-Muñoz, F.; Ferre, F.; Martínez-Gras, I.; Ponce, G.; Pascual, J.M.; Jiménez-Arriero, M.A.; Alamo, C. Effects of zonisamide in the treatment of alcohol dependence. Clin. Neuropharmacol. 2010, 33, 250–253. [Google Scholar] [CrossRef]
- Sarid-Segal, O.; Knapp, C.M.; Burch, W.; Richardson, M.A.; Bahtia, S.; DeQuattro, K.; Afshar, M.; Richambault, C.; Sickels, L.; Devine, E. The anticonvulsant zonisamide reduces ethanol self-administration by risky drinkers. Am. J. Drug Alcohol Abus. 2009, 35, 316–319. [Google Scholar] [CrossRef]
- Sripathirathan, K.; Brown, J., III; Neafsey, E.J.; Collins, M.A. Linking binge alcohol-induced neurodamage to brain edema and potential aquaporin-4 upregulation: Evidence in rat organotypic brain slice cultures and in vivo. J. Neurotrauma 2009, 26, 261–273. [Google Scholar] [CrossRef]
- Swenson, E.R. Safety of carbonic anhydrase inhibitors. Expert. Opin. Drug Saf. 2014, 13, 459–472. [Google Scholar] [CrossRef]
- De Simone, G.; Di Fiore, A.; Menchise, V.; Pedone, C.; Antel, J.; Casini, A.; Scozzafava, A.; Wurl, M.; Supuran, C.T. Carbonic anhydrase inhibitors. Zonisamide is an effective inhibitor of the cytosolic isozyme II and mitochondrial isozyme V: Solution and X-ray crystallographic studies. Bioorg. Med. Chem. Lett. 2005, 15, 2315–2320. [Google Scholar] [CrossRef] [PubMed]
- White, H.S. Mechanism of action of newer anticonvulsants. J. Clin. Psychiatry 2003, 64, 5–8. [Google Scholar] [PubMed]
- Yamamura, S.; Saito, H.; Suzuki, N.; Kashimoto, S.; Hamaguchi, T.; Ohoyama, K.; Suzuki, D.; Kanehara, S.; Nakagawa, M.; Shiroyama, T. Effects of zonisamide on neurotransmitter release associated with inositol triphosphate receptors. Neurosci. Lett. 2009, 454, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Okada, M.; Zhu, G.; Kaneko, S. Effects of zonisamide on neurotransmitter exocytosis associated with ryanodine receptors. Epilepsy Res. 2005, 67, 153–162. [Google Scholar] [CrossRef]
- Amaral, D.G.; Witter, M.P. The three-dimensional organization of the hippocampal formation: A review of anatomical data. Neuroscience 1989, 31, 571–591. [Google Scholar] [CrossRef]
- Snyder, J.S.; Radik, R.; Wojtowicz, J.M.; Cameron, H.A. Anatomical gradients of adult neurogenesis and activity: Young neurons in the ventral dentate gyrus are activated by water maze training. Hippocampus 2009, 19, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Wells, A.M.; Lasseter, H.C.; Xie, X.; Cowhey, K.E.; Reittinger, A.M.; Fuchs, R.A. Interaction between the basolateral amygdala and dorsal hippocampus is critical for cocaine memory reconsolidation and subsequent drug context-induced cocaine-seeking behavior in rats. Learn. Mem. 2011, 18, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Rogers, J.L.; See, R.E. Selective inactivation of the ventral hippocampus attenuates cue-induced and cocaine-primed reinstatement of drug-seeking in rats. Neurobiol. Learn. Mem. 2007, 87, 688–692. [Google Scholar] [CrossRef] [Green Version]
- Lasseter, H.C.; Xie, X.; Ramirez, D.R.; Fuchs, R.A. Sub-region specific contribution of the ventral hippocampus to drug context-induced reinstatement of cocaine-seeking behavior in rats. Neuroscience 2010, 171, 830–839. [Google Scholar] [CrossRef] [Green Version]
- Lau, J.Y.; Goldman, D.; Buzas, B.; Hodgkinson, C.; Leibenluft, E.; Nelson, E.; Sankin, L.; Pine, D.S.; Ernst, M. BDNF gene polymorphism (Val66Met) predicts amygdala and anterior hippocampus responses to emotional faces in anxious and depressed adolescents. Neuroimage 2010, 53, 952–961. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.; Feng, R.; Jacobs, S.; Duan, Y.; Wang, H.; Cao, X.; Tsien, J.Z. Increased NR2A:NR2B ratio compresses long-term depression range and constrains long-term memory. Sci. Rep. 2013, 3, 1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.; Elberger, A.J.; Matthews, D.B.; Hamre, K.M. Heterozygous deletion of NR1 subunit of the NMDA receptor alters ethanol-related behaviors and regional expression of NR2 subunits in the brain. Neurotoxicol. Teratol. 2012, 34, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, H.S.; Mehta, A.K.; Ticku, M.K. Up-regulation of NMDA receptor subunits in rat brain following chronic ethanol treatment. Brain Res. 1998, 58, 221–224. [Google Scholar] [CrossRef]
- Prendergast, M.A.; Mulholland, P.J. Glucocorticoid and polyamine interactions in the plasticity of glutamatergic synapses that contribute to ethanol-associated dependence and neuronal injury. Addict. Biol. 2012, 17, 209–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milenkovic, I.; Vasiljevic, M.; Maurer, D.; Hoger, H.; Klausberger, T.; Sieghart, W. The parvalbumin-positive interneurons in the mouse dentate gyrus express GABAA receptor subunits alpha1, beta2, and delta along their extrasynaptic cell membrane. Neuroscience 2013, 254, 80–96. [Google Scholar] [CrossRef]
- Tong, X.; Peng, Z.; Zhang, N.; Cetina, Y.; Huang, C.S.; Wallner, M.; Otis, T.S.; Houser, C.R. Ectopic Expression of alpha6 and delta GABAA Receptor Subunits in Hilar Somatostatin Neurons Increases Tonic Inhibition and Alters Network Activity in the Dentate Gyrus. J. Neurosci. 2015, 35, 16142–16158. [Google Scholar] [CrossRef] [Green Version]
- O’Dell, L.E.; Chen, S.A.; Smith, R.T.; Specio, S.E.; Balster, R.L.; Paterson, N.E.; Markou, A.; Zorrilla, E.P.; Koob, G.F. Extended access to nicotine self-administration leads to dependence: Circadian measures, withdrawal measures, and extinction behavior in rats. J. Pharmacol. Exp. Ther. 2007, 320, 180–193. [Google Scholar] [CrossRef]
- Gilpin, N.W.; Richardson, H.N.; Cole, M.; Koob, G.F. Vapor inhalation of alcohol in rats. Curr. Protoc. Neurosci. 2008, 44, 1–19. [Google Scholar] [CrossRef]
- Richardson, H.N.; Lee, S.Y.; O’Dell, L.E.; Koob, G.F.; Rivier, C.L. Alcohol self-administration acutely stimulates the hypothalamic-pituitary-adrenal axis, but alcohol dependence leads to a dampened neuroendocrine state. Eur. J. Neurosci. 2008, 28, 1641–1653. [Google Scholar] [CrossRef] [Green Version]
- Majchrowicz, E. Induction of physical dependence upon ethanol and the associated behavioral changes in rats. Psychopharmacologia 1975, 43, 245–254. [Google Scholar] [CrossRef]
- Cohen, A.; Soleiman, M.T.; Talia, R.; Koob, G.F.; George, O.; Mandyam, C.D. Extended access nicotine self-administration with periodic deprivation increases immature neurons in the hippocampus. Psychopharmacology 2015, 232, 453–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welinder, C.; Ekblad, L. Coomassie staining as loading control in Western blot analysis. J. Proteome Res. 2011, 10, 1416–1419. [Google Scholar] [CrossRef] [PubMed]
- Thacker, J.S.; Yeung, D.H.; Staines, W.R.; Mielke, J.G. Total protein or high-abundance protein: Which offers the best loading control for Western blotting? Anal. Biochem. 2016, 496, 76–78. [Google Scholar] [CrossRef]
- Recinto, P.; Samant, A.R.; Chavez, G.; Kim, A.; Yuan, C.J.; Soleiman, M.; Grant, Y.; Edwards, S.; Wee, S.; Koob, G.F.; et al. Levels of neural progenitors in the hippocampus predict memory impairment and relapse to drug seeking as a function of excessive methamphetamine self-administration. Neuropsychopharmacology 2012, 37, 1275–1287. [Google Scholar] [CrossRef] [PubMed]
- Mandyam, C.D.; Norris, R.D.; Eisch, A.J. Chronic morphine induces premature mitosis of proliferating cells in the adult mouse subgranular zone. J. Neurosci. Res. 2004, 76, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Somkuwar, S.S.; Fannon-Pavlich, M.J.; Ghofranian, A.; Quigley, J.A.; Dutta, R.R.; Galinato, M.H.; Mandyam, C.D. Wheel running reduces ethanol seeking by increasing neuronal activation and reducing oligodendroglial/neuroinflammatory factors in the medial prefrontal cortex. Brain Behav. Immun. 2016, 58, 357–368. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sona Khan, M.; Trenet, W.; Xing, N.; Sibley, B.; Abbas, M.; al-Rashida, M.; Rauf, K.; Mandyam, C.D. A Novel Sulfonamide, 4-FS, Reduces Ethanol Drinking and Physical Withdrawal Associated With Ethanol Dependence. Int. J. Mol. Sci. 2020, 21, 4411. https://doi.org/10.3390/ijms21124411
Sona Khan M, Trenet W, Xing N, Sibley B, Abbas M, al-Rashida M, Rauf K, Mandyam CD. A Novel Sulfonamide, 4-FS, Reduces Ethanol Drinking and Physical Withdrawal Associated With Ethanol Dependence. International Journal of Molecular Sciences. 2020; 21(12):4411. https://doi.org/10.3390/ijms21124411
Chicago/Turabian StyleSona Khan, Muhammad, Wulfran Trenet, Nancy Xing, Britta Sibley, Muzaffar Abbas, Mariya al-Rashida, Khalid Rauf, and Chitra D. Mandyam. 2020. "A Novel Sulfonamide, 4-FS, Reduces Ethanol Drinking and Physical Withdrawal Associated With Ethanol Dependence" International Journal of Molecular Sciences 21, no. 12: 4411. https://doi.org/10.3390/ijms21124411