Carbonic Anhydrase Inhibitor Acetazolamide Enhances CHOP Treatment Response and Stimulates Effector T-Cell Infiltration in A20/BalbC Murine B-Cell Lymphoma

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
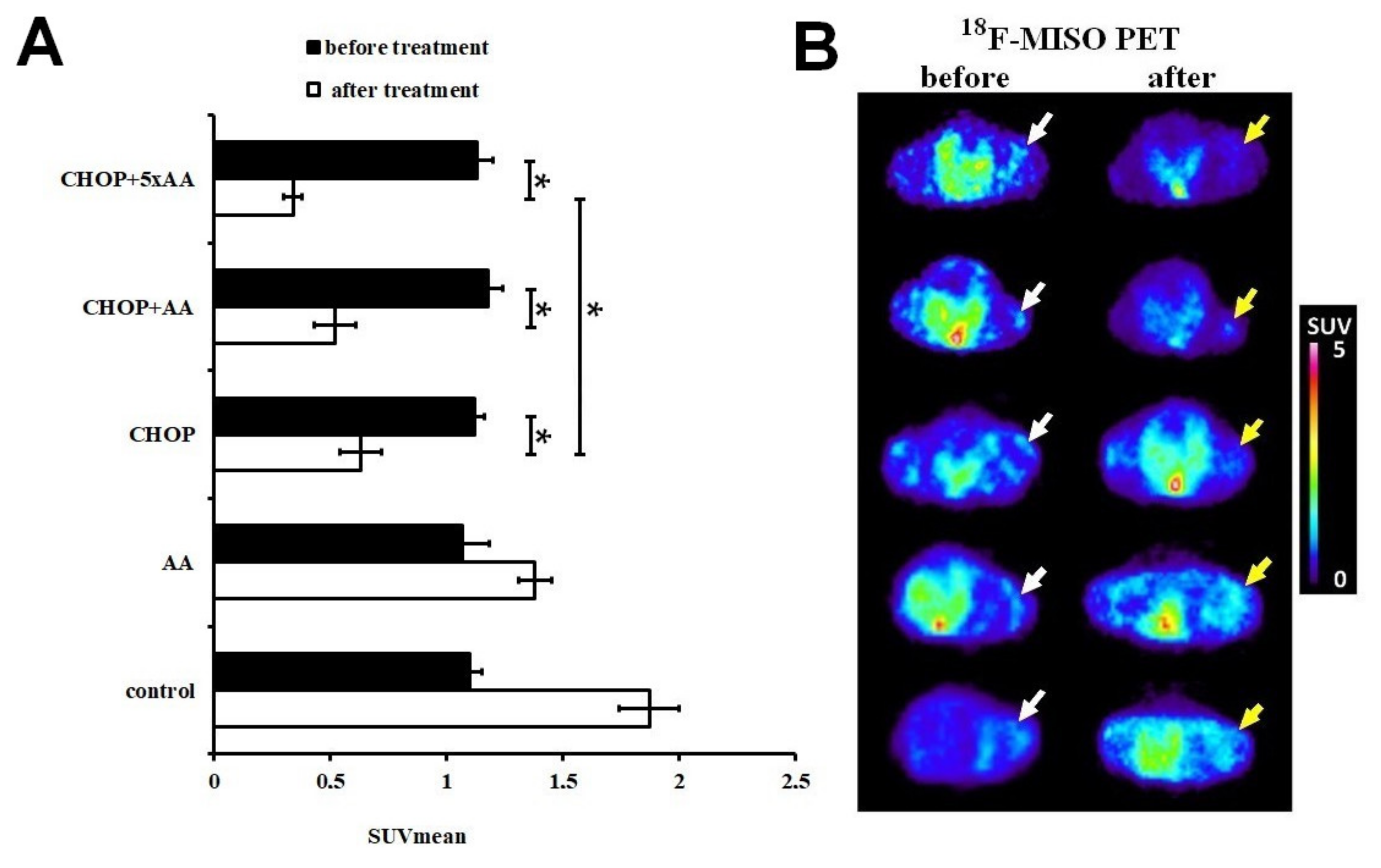
2.1. Effect of CHOP-AA Combination on In Vivo Tumor Metabolic Activity
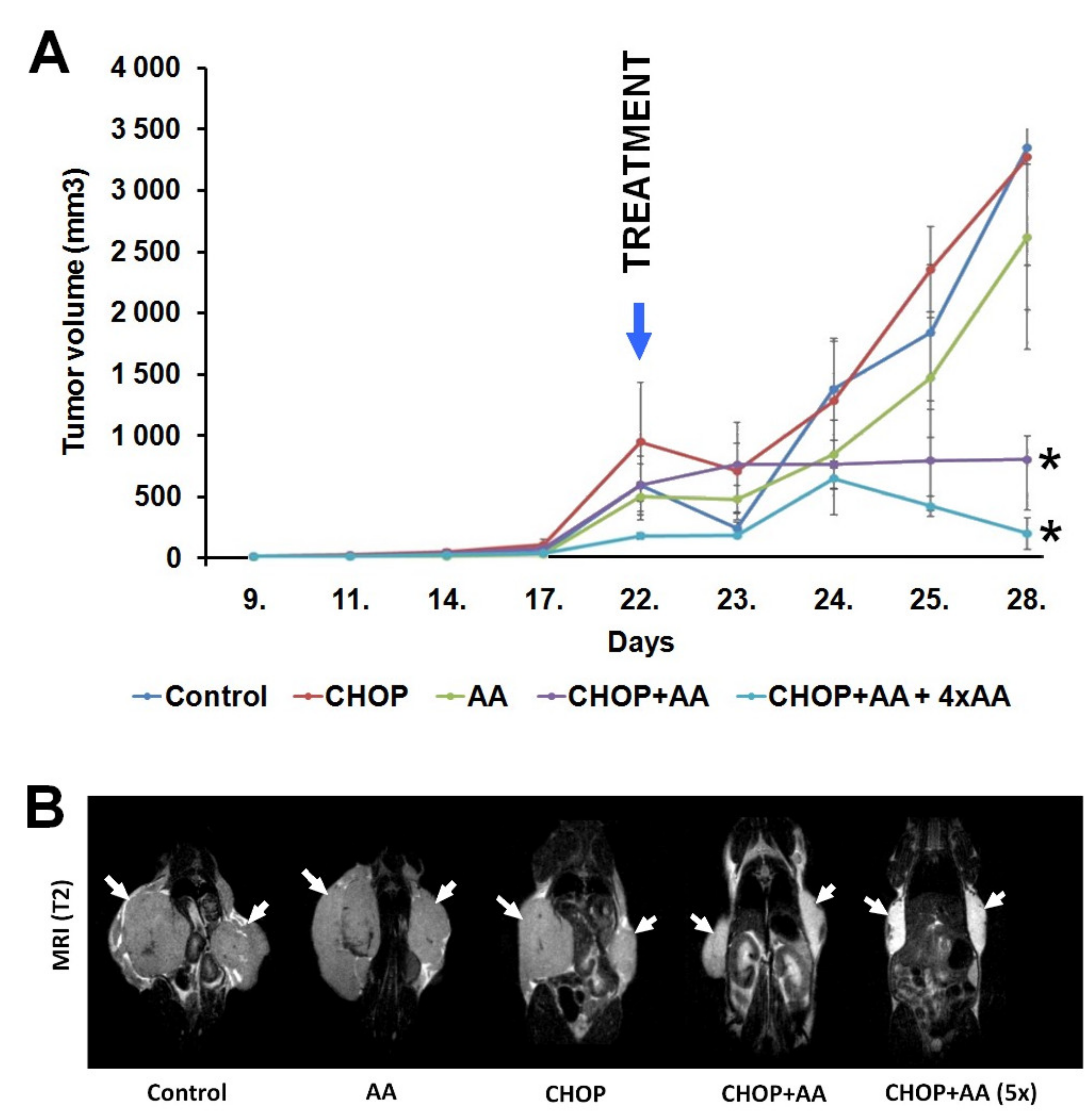
2.2. Tumor Volume
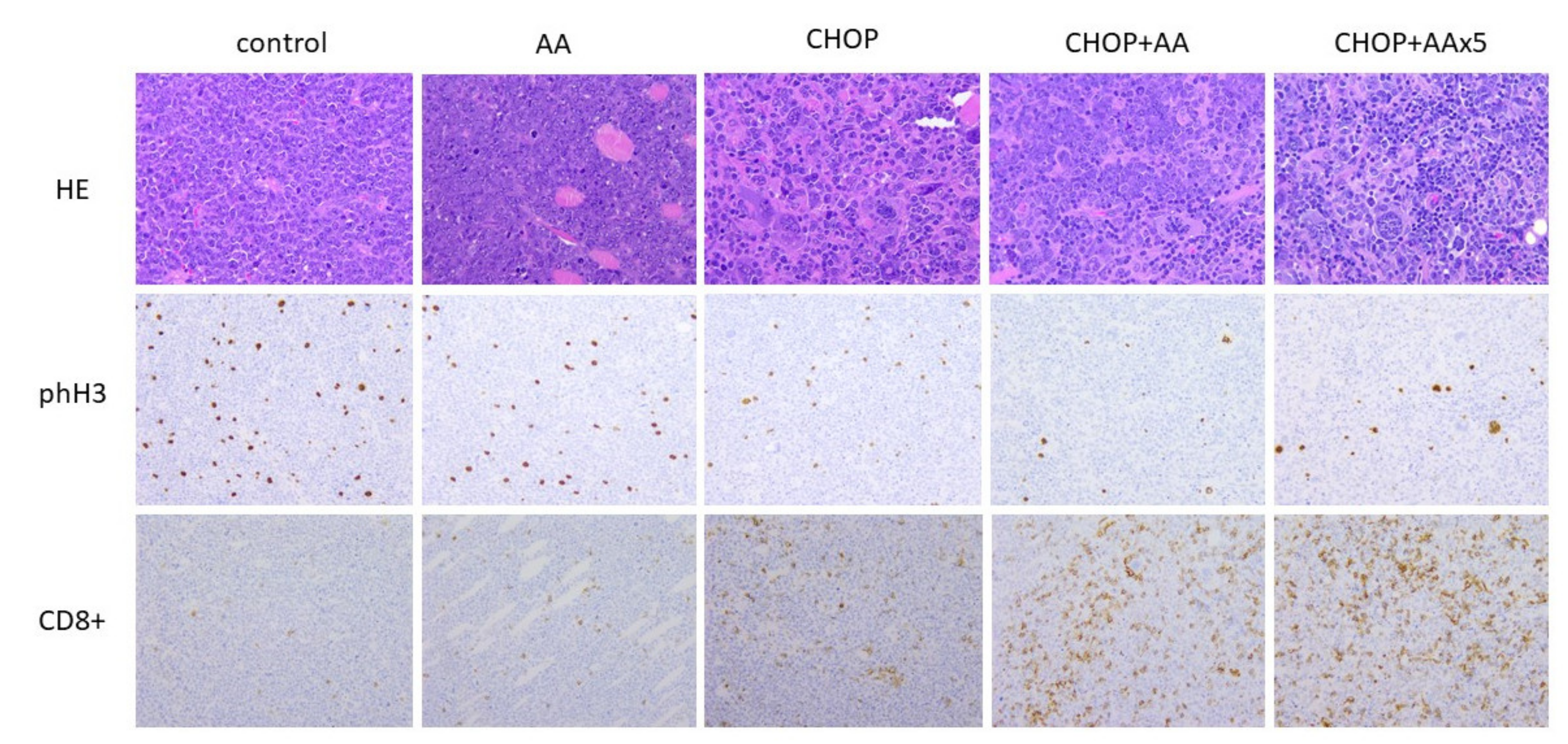
2.3. General Histomorphology
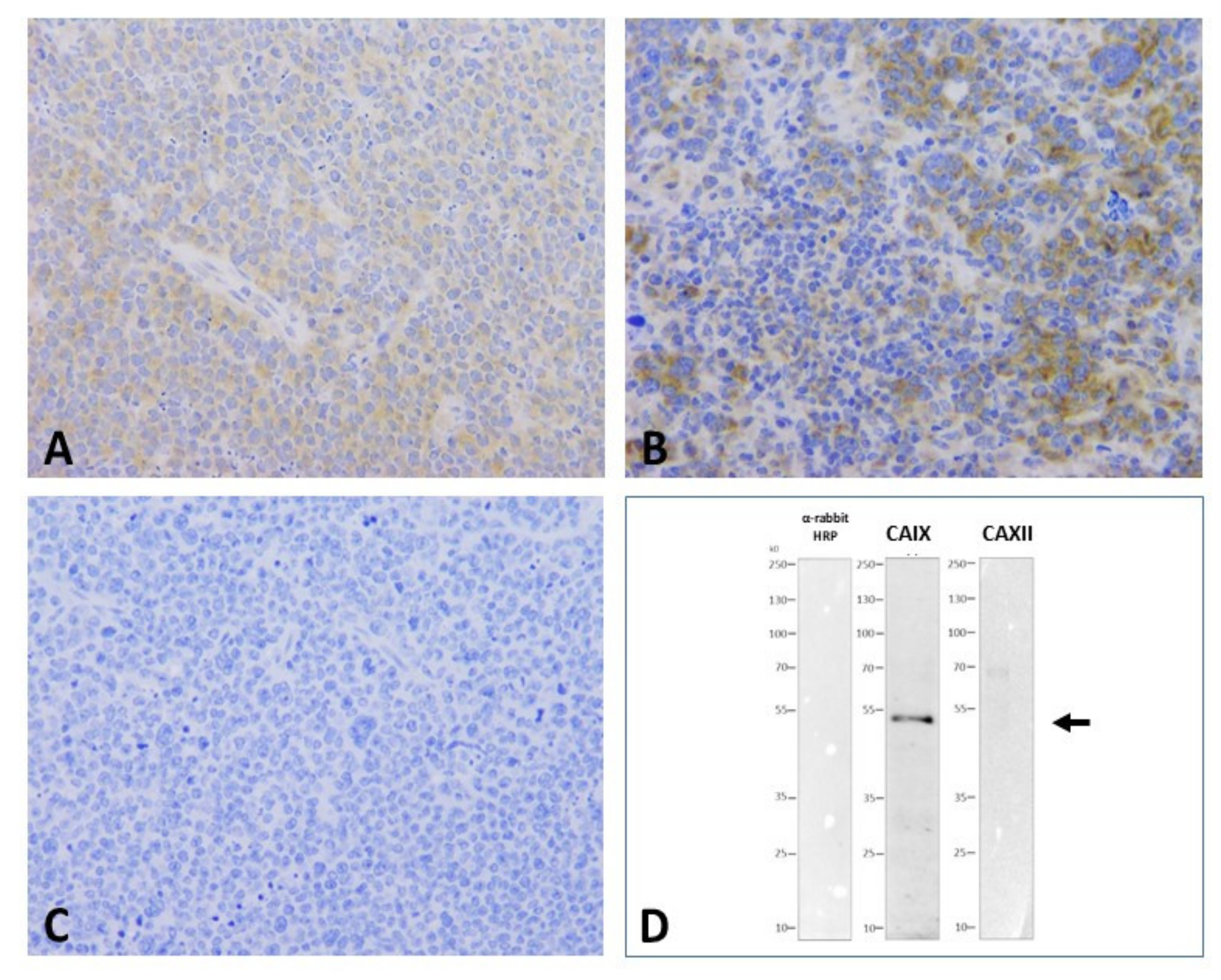
2.4. CAIX and CAXII Expression
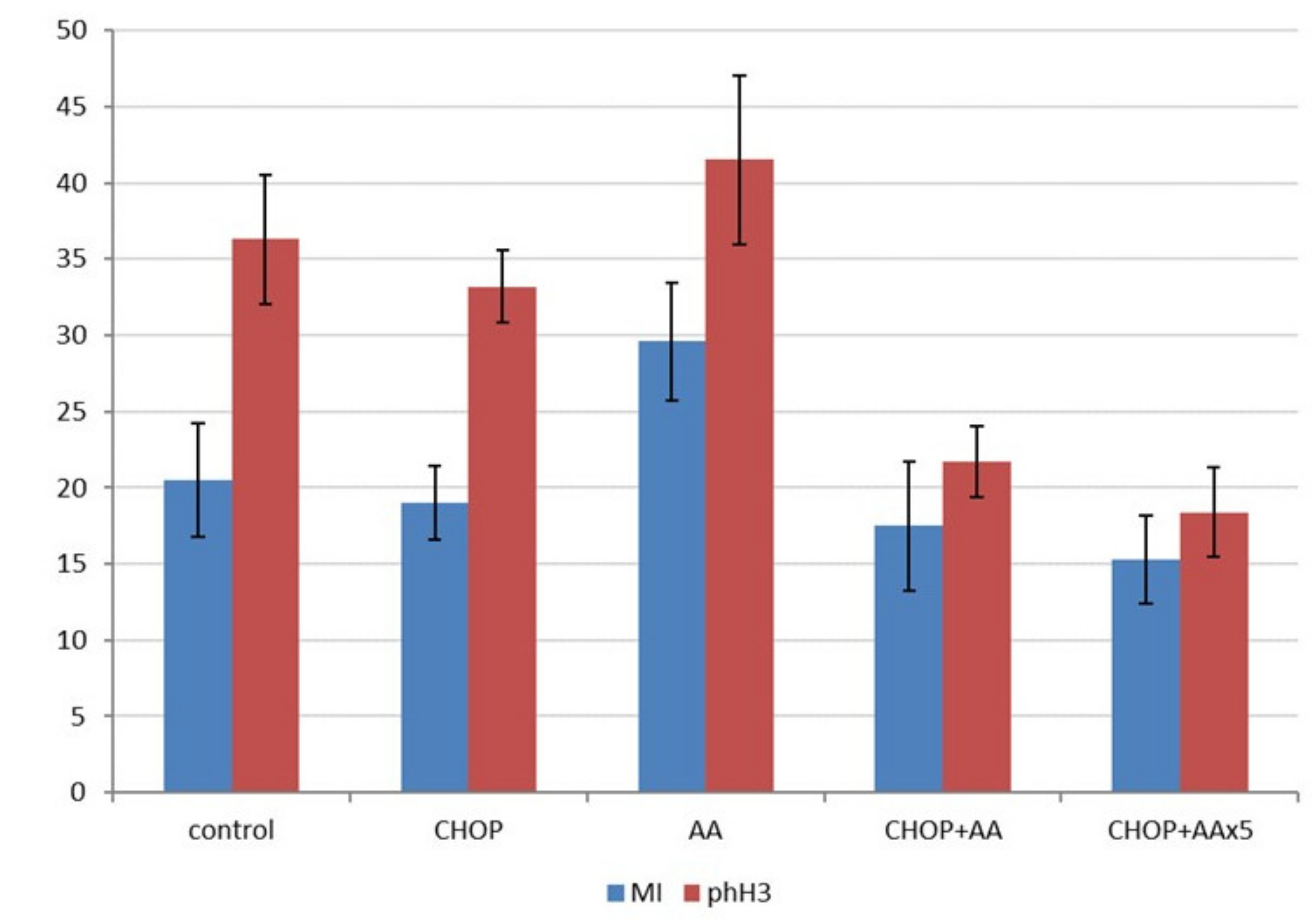
2.5. Proliferative/Mitotic Activity
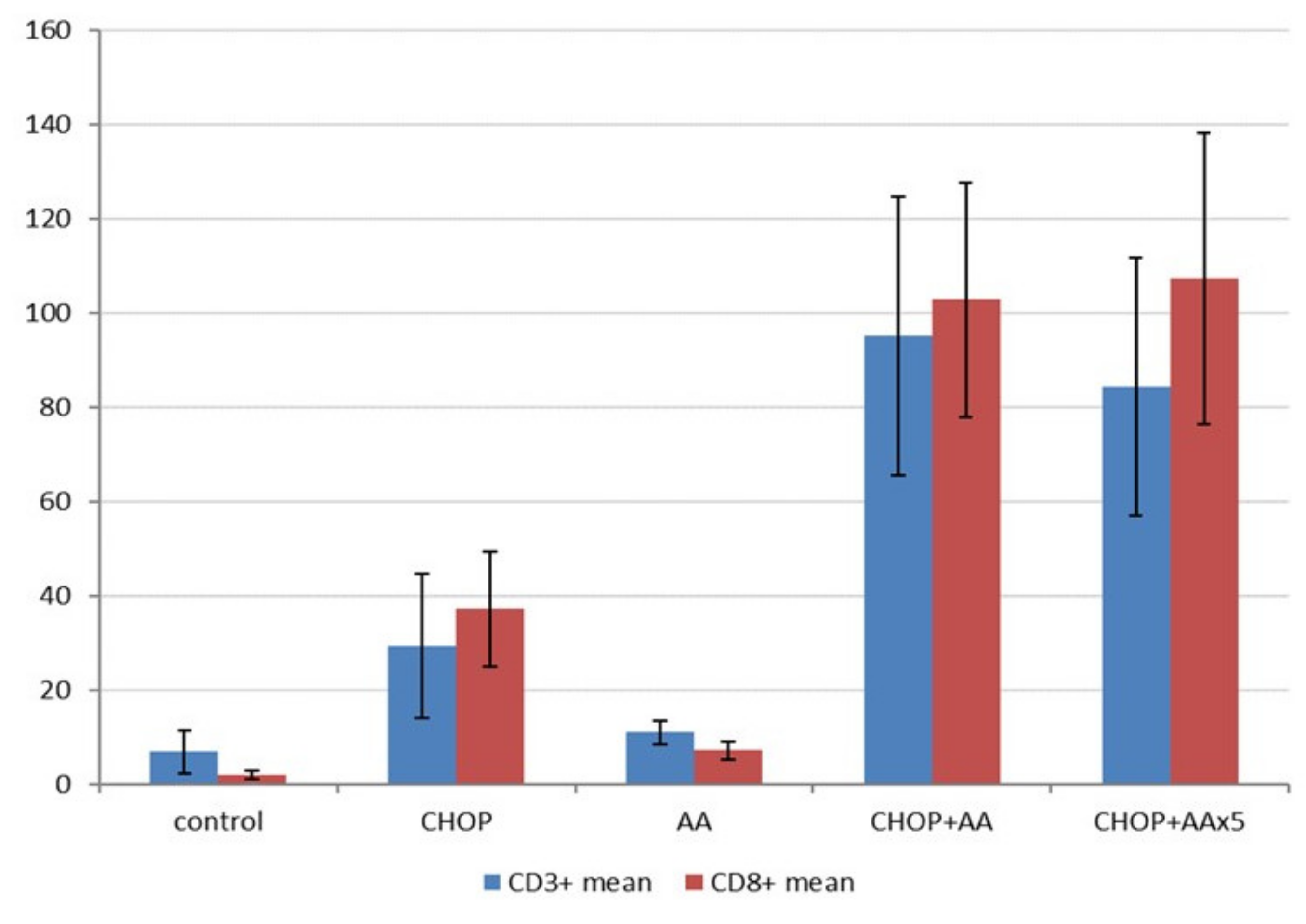
2.6. T-cell Immune Infiltrate
3. Discussion
4. Materials and Methods
4.1. In Vitro Culturing of A20 Murine Lymphoma Cells
4.2. A20/BalbC Syngenic Murine Lymphoma Model
4.3. Animal Treatment and Drugs
4.4. In Vivo MRI and PET Imaging of Tumor Hypoxia
4.5. Quantitative PET Data Analysis
4.6. Tissue-based Analysis
4.7. Western Blotting and Validation of CAIX and CAXII Specific Staining
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Semenza, G.L. The hypoxic tumor microenvironment: A driving force for breast cancer progression. Biochim. Biophys. Acta 2016, 1863, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Vadde, R.; Vemula, S.; Jinka, R.; Merchant, N.; Bramhachari, P.V.; Nagaraju, G.P. Role of Hypoxia-Inducible Factors (HIF) in the Maintenance of Stemness and Malignancy of Colorectal Cancer. Crit. Rev. Oncol. Hematol. 2017, 113, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Choudhry, H.; Harris, A.L. Advances in Hypoxia-Inducible Factor Biology. Cell Metab. 2018, 27, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Manoochehri Khoshinani, H.; Afshar, S.; Najafi, R. Hypoxia: A Double-Edged Sword in Cancer Therapy. Cancer Invest. 2016, 34, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Vaupel, P.; Mayer, A. Tumor Hypoxia: Causative Mechanisms, Microregional Heterogeneities, and the Role of Tissue-Based Hypoxia Markers. Adv. Exp. Med. Biol. 2016, 923, 77–86. [Google Scholar] [CrossRef]
- McDonald, P.C.; Chafe, S.C.; Dedhar, S. Overcoming Hypoxia-Mediated Tumor Progression: Combinatorial Approaches Targeting pH Regulation, Angiogenesis and Immune Dysfunction. Front. Cell Dev. Biol. 2016, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Mboge, M.Y.; Mahon, B.P.; McKenna, R.; Frost, S.C. Carbonic anhydrases: Role in pH control and cancer. Metabolites 2018, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T. Carbonic anhydrases and metabolism. Metabolites 2018, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Harguindey, S.; Reshkin, S.J. The new pH-centric anticancer paradigm in oncology and medicine. Semin Cancer Biol. 2017, 43, 1–4. [Google Scholar] [CrossRef]
- Lee, S.H.; McIntyre, D.; Honess, D.; Hulikova, A.; Pacheco-Torres, J.; Cerdan, S.; Swietach, P.; Harris, A.L.; Griffith, J.R. Carbonic anhydrase IX is a pH-stat that sets an acidic tumor extracellular pH in vivo. Br. J. Cancer 2018, 119, 622–630. [Google Scholar] [CrossRef] [Green Version]
- Sedlakova, O.; Svastova, E.; Takacova, M.; Kopacek, J.; Pastorek, J.; Pastorekova, S. Carbonic anhydrase IX, a hypoxia induced catalytic component of the pH regulating machinery in tumors. Front. Physiol. 2014, 4, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastorek, J.; Pastorekova, S. Hypoxia induced carbonic anhydrase IX as a target for cancer therapy: From biology to clinical use. Semin Cancer Biol. 2015, 31, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Becker, H.M. Carbonic anhydrase IX and acid transport in cancer. Br. J. Cancer 2020, 122, 157–167. [Google Scholar] [CrossRef] [PubMed]
- van Kuijk, S.J.; Yaromina, A.; Houben, R.; Niemans, R.; Lambin, P.; Dubois, L.J. Prognostic significance of carbonic anhydrase IX in cancer patients: A meta analysis. Front. Oncol. 2016, 29, 69. [Google Scholar]
- Teppema, L.J.; Balanos, G.M.; Steinback, C.D.; Brown, A.D.; Foster, G.E.; Duff, H.J.; Leigh, R.; Poulin, M.J. Effects of Acetazolamide on Ventilatory, Cerebrovascular, and Pulmonary Vascular Responses to Hypoxia. Am. J. Respir. Crit. Care Med. 2007, 175, 277–281. [Google Scholar] [CrossRef]
- Van Berkel, M.A.; Elefritz, J.L. Evaluating Off-Label Uses of Acetazolamide. Am. J. Health Syst. Pharm. 2018, 75, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Carbonic Anhydrase Inhibitors as Emerging Agents for the Treatment and Imaging of Hypoxic Tumors. Expert Opin Investig Drugs 2018, 27, 963–970. [Google Scholar] [CrossRef]
- Nocetini, A.; Supuran, C.T. Carbonic Anhydrase Inhibitors as antitumor/antimetastatic Agents: A Patent Review (2008-2018). Expert Opin Ther Pat. 2018, 28, 729–740. [Google Scholar] [CrossRef]
- Koyuncu, I.; Gonel, A.; Durgun, M.; Kocyigit, A.; Yuksekdag, O.; Supuran, C.T. Assessment of the Antiproliferative and Apoptotic Roles of Sulfonamide Carbonic Anhydrase IX Inhibitors in HeLa Cancer Cell Line. J. Enzyme Inhib. Med. Chem. 2019, 34, 75–86. [Google Scholar] [CrossRef] [Green Version]
- McDonald, P.C.; Chia, S.; Bedard, P.L.; Chu, Q.; Lyle, M.; Tang, L.; Singh, M.; Zhang, Z.; Supuran, C.T.; Renouf, D.J.; et al. A phase 1 study of SLC-0111, a novel inhibitor of carbonic anhydrase IX, in patients with advanced solid tumors. Am. J. Clin. Oncol 2020, 43, 484–490. [Google Scholar] [CrossRef]
- Ahlskog, J.K.; Dumelin, C.E.; Trüssel, S.; Mårlind, J.; Neri, D. In Vivo Targeting of Tumor-Associated Carbonic Anhydrases Using Acetazolamide Derivatives. Bioorg. Med. Chem. Lett. 2009, 19, 4851–4856. [Google Scholar] [CrossRef]
- Dubois, L.; Peeters, S.; Lieuwes, N.G.; Geusens, N.; Thiry, A.; Wigfield, S.; Carta, F.; McIntyre, A.; Scozzafava, A.; Dogné, J.M.; et al. Specific Inhibition of Carbonic Anhydrase IX Activity Enhances the in Vivo Therapeutic Effect of Tumor Irradiation. Radiother. Oncol. 2011, 99, 424–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Dong, H.; Li, G.; Jin, H. Combined Treatment With Acetazolamide and Cisplatin Enhances Chemosensitivity in Laryngeal Carcinoma Hep-2. Cells Oncol. Lett. 2018, 15, 9299–9306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazzamalli, S.; Figueras, E.; Pethő, L.; Borbély, A.; Steinkühler, C.; Neri, D.; Sewald, N. In Vivo Antitumor Activity of a Novel Acetazolamide-Cryptophycin Conjugate for the Treatment of Renal Cell Carcinomas. ACS Omega 2018, 3, 14726–14731. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.; Talks, K.; Leek, R.; Turley, H.; Pezzella, F.; Harris, A.; Gatter, K. Expression of angiogenic factors and hypoxia inducible factors HIF1, HIF2 and CAIX in non-Hodgkin’s lymphoma. Histopathology 2002, 40, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Matolay, O.; Méhes, G. Sustain, adapt and overcome: Hypoxia associated changes in the progression of lymphatic neoplasia. Front. Oncol. 2019, 9, 1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.Q.; Howison, C.M.; Spier, C.; Stopeck, A.T.; Malm, S.W.; Pagel, M.D.; Baker, A.F. Assessment of carbonic anhydrase IX expression and extracellular pH in B-cell lymphoma cell line models. Leuk. Lymphoma 2015, 56, 1432–1439. [Google Scholar] [CrossRef] [Green Version]
- Méhes, G.; Matolay, O.; Beke, L.; Czenke, M.; Jóna, Á.; Miltényi, Z.; Illés, Á.; Bedekovics, J. Hypoxia-related carbonic anhydrase IX expression is associated with unfavourable response to first-line therapy in classical Hodgkin’s lymphoma. Histopathology 2019, 74, 699–708. [Google Scholar] [CrossRef]
- Matolay, O.; Beke, L.; Gyurkovics, A.; Francz, M.; Varjasi, G.; Rejtő, L.; Illés, Á.; Bedekovics, J.; Méhes, G. Quantitative Analysis of Carbonic Anhydrase IX Uncovers Hypoxia-Related Functional Differences in Classical Hodgkin Lymphoma Subtypes. Int J. Mol. Sci. 2019, 20, 3463. [Google Scholar] [CrossRef] [Green Version]
- Bascuas, T.; Moreno, M.; Mónaco, A.; Reyes, L.; Paolino, A. A novel non-Hodgkin lymphoma murine model closer to the clinical scenario. J. Transl. Med. 2016, 14, 323–339. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T. An update on drug interaction considerations in the therapeutic use of carbonic anhydrase inhibitors. Expert Opin. Drug Metab. Toxicol. 2020, 16, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Donnou, S.; Galand, C.; Touitou, V.; Sautès-Fridman, C.; Fabry, Z.; Fisson, S. Murine models of B-cell lymphomas: Promising tools for designing cancer therapies. Adv Hematol. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagal, A.S.; Leach, J.L.; Fernandez-Ulloa, M.; Zuccarello, M. The acetazolamide challenge: Techniques and applications in the evaluation of chronic cerebral ischemia. AJNR Am. J. Neuroradiol. 2009, 30, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Suefuji, H.; Hirose, Y.; Kaida, H.; Suzuki, G.; Uozumi, J. 18F-FDG uptake in primary gastric malignant lymphoma correlates with glucose transporter 1 expression and histologic malignant potential. Intl. J. Hematol. 2013, 97, 43–49. [Google Scholar] [CrossRef]
- Liu, Y.M.; Zhai, X.M.; Wu, Y.W. Biological correlation between glucose transporters, Ki-67 and 2-deoxy-2-[18F]-fluoro-D-glucose uptake in diffuse large B-cell lymphoma and natural killer/T-cell lymphoma. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Fischer, K.; Hoffmann, P.; Voelkl, S.; Meidenbauer, N.; Ammer, J.; Edinger, M. Inhibitory effect of tumor cell derived lactic acid on human T-cells. Blood 2007, 109, 3812–3819. [Google Scholar] [CrossRef]
- Wang, J.; Liu, G.; Wang, R. The intercellular metabolic interplay between tumor and immune cells. Front. Immunol. 2014, 5, 358. [Google Scholar] [CrossRef] [Green Version]
- Huber, V.; Camisashi, C.; Berzi, A.; Ferro, S.; Lugini, L.; Triulzi, T.; Tuccitto, A.; Tagliabue, E.; Castelli, C.; Rivoltini, L. Cancer acidity: An ultimate frontier of tumor immune escape and novel target of immunomodulation. Semin Cancer Biol. 2017, 43, 74–89. [Google Scholar] [CrossRef]
- Pilon-Thomas, S.; Kodumudi, K.N.; El-Kenawi, A.E.; Russel, S.; Weber, A.M.; Luddy, K.; Damaghi, M.; Wojtkowiak, J.W.; Mule, J.J.; Ibrahim-Hasim, A. Neutralization of tumor acidity improves antitumor responses to immunotherapy. Cancer Res. 2016, 76, 1381–1390. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, R.; Rozeman, E.A.; Kreutz, M.; Renner, K.; Blank, C.U. Targeting tumor-associated acidity in cancer immunotherapy. Cancer Immunol. Immunother. 2018, 67, 1331–1348. [Google Scholar] [CrossRef]
- Nagy, L.; Marton, J.; Vida, A.; Kis, G.; Bokor, E.; Kun, S.; Gonczi, M.; Docsa, T.; Toth, A.; Antal, M.; et al. Glycogen phosphorylase inhibition improves beta cell function. Br. J. Pharmacol. 2018, 175, 301–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méhes, G.; Matolay, O.; Beke, L.; Czenke, M.; Pórszász, R.; Mikó, E.; Bai, P.; Berényi, E.; Trencsényi, G. Carbonic Anhydrase Inhibitor Acetazolamide Enhances CHOP Treatment Response and Stimulates Effector T-Cell Infiltration in A20/BalbC Murine B-Cell Lymphoma. Int. J. Mol. Sci. 2020, 21, 5001. https://doi.org/10.3390/ijms21145001
Méhes G, Matolay O, Beke L, Czenke M, Pórszász R, Mikó E, Bai P, Berényi E, Trencsényi G. Carbonic Anhydrase Inhibitor Acetazolamide Enhances CHOP Treatment Response and Stimulates Effector T-Cell Infiltration in A20/BalbC Murine B-Cell Lymphoma. International Journal of Molecular Sciences. 2020; 21(14):5001. https://doi.org/10.3390/ijms21145001
Chicago/Turabian StyleMéhes, Gábor, Orsolya Matolay, Lívia Beke, Marianna Czenke, Róbert Pórszász, Edit Mikó, Péter Bai, Ervin Berényi, and György Trencsényi. 2020. "Carbonic Anhydrase Inhibitor Acetazolamide Enhances CHOP Treatment Response and Stimulates Effector T-Cell Infiltration in A20/BalbC Murine B-Cell Lymphoma" International Journal of Molecular Sciences 21, no. 14: 5001. https://doi.org/10.3390/ijms21145001