
Effect of Sphingomyelinase-Treated LDLs on HUVECs
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Sphingomyelinase Treatment Induces Physico-Chemical Modification in LDL
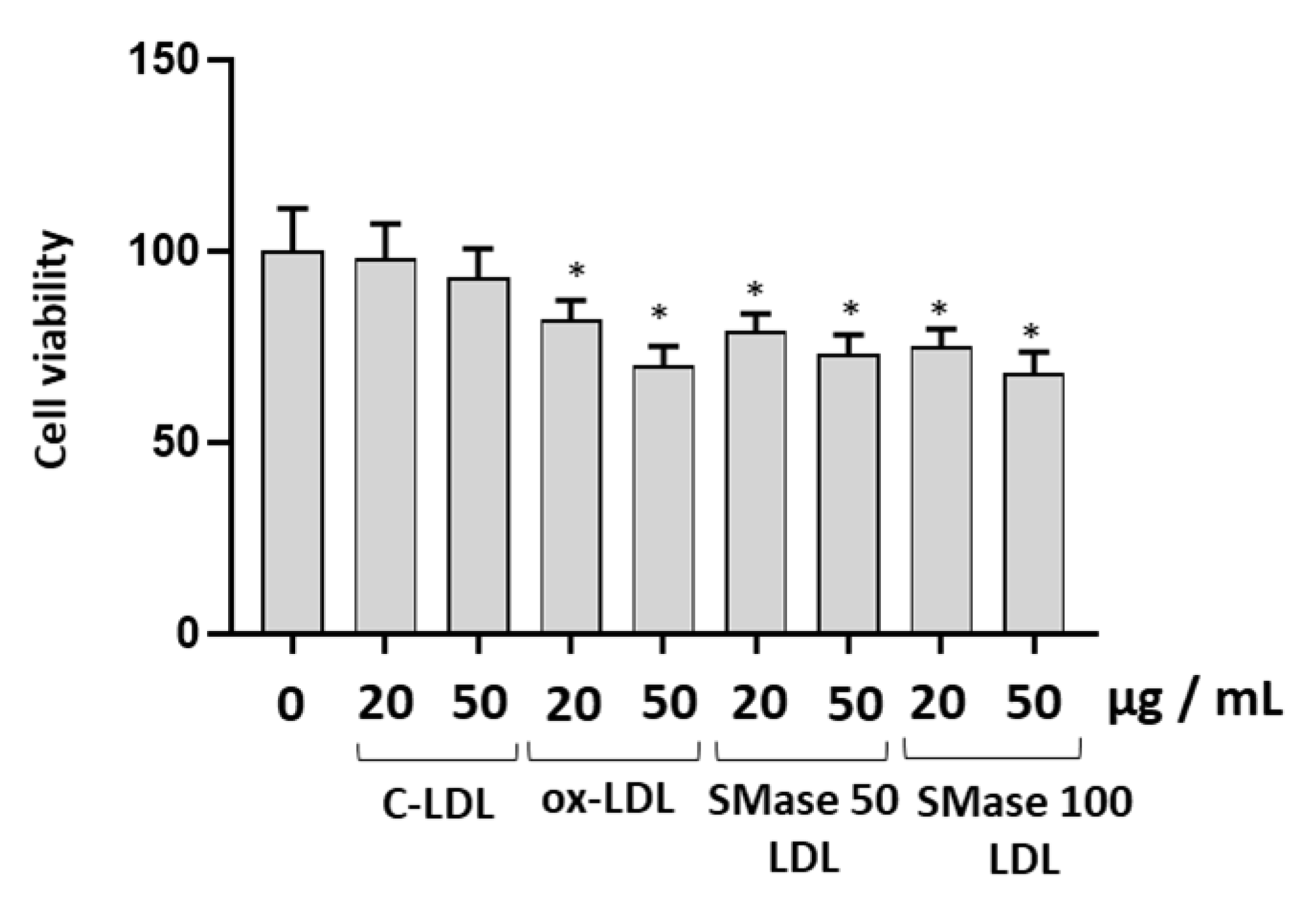
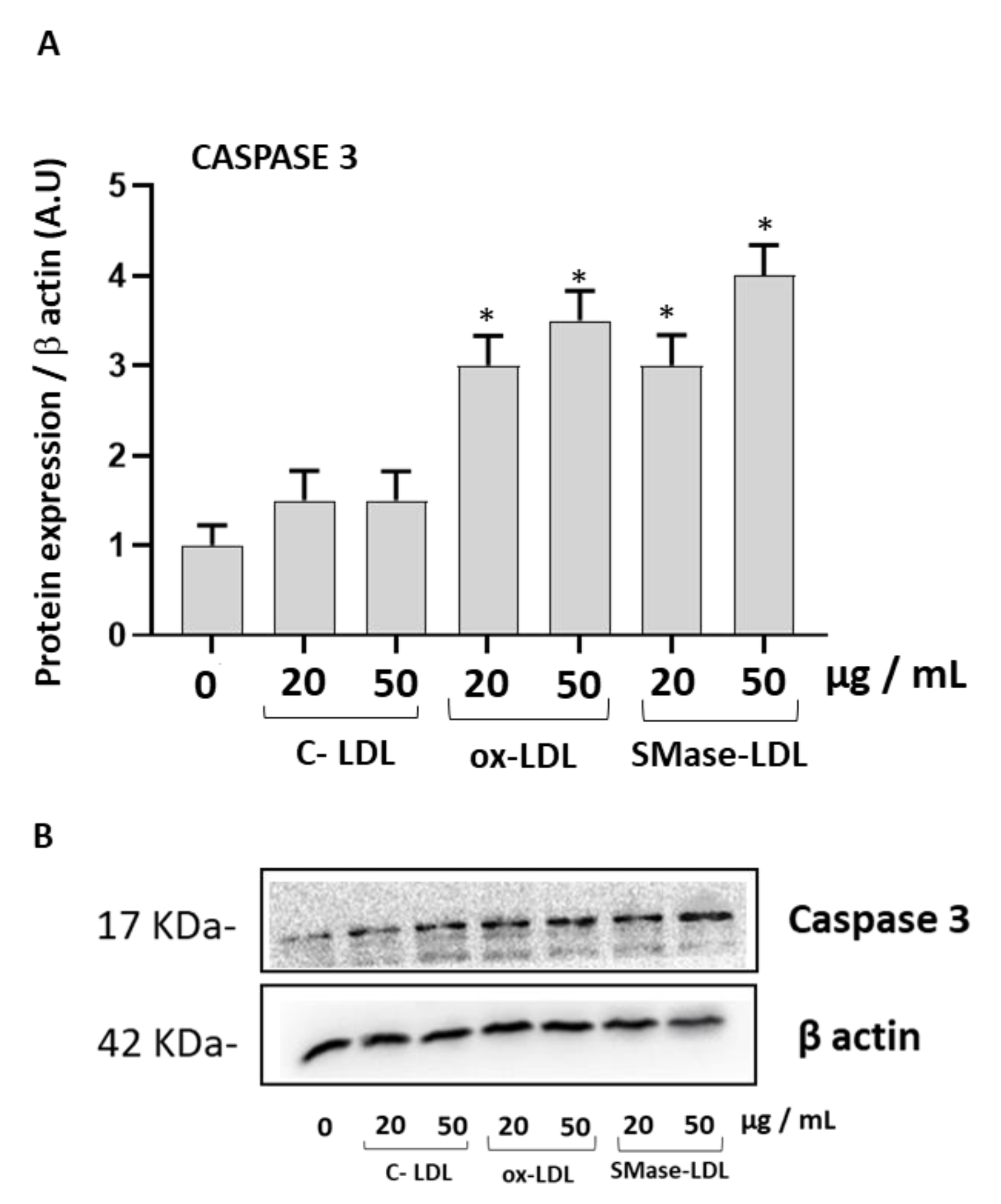
2.2. Modified LDLs Decreased Cell Viability and Increased Caspase-3 Expression in HUVECs
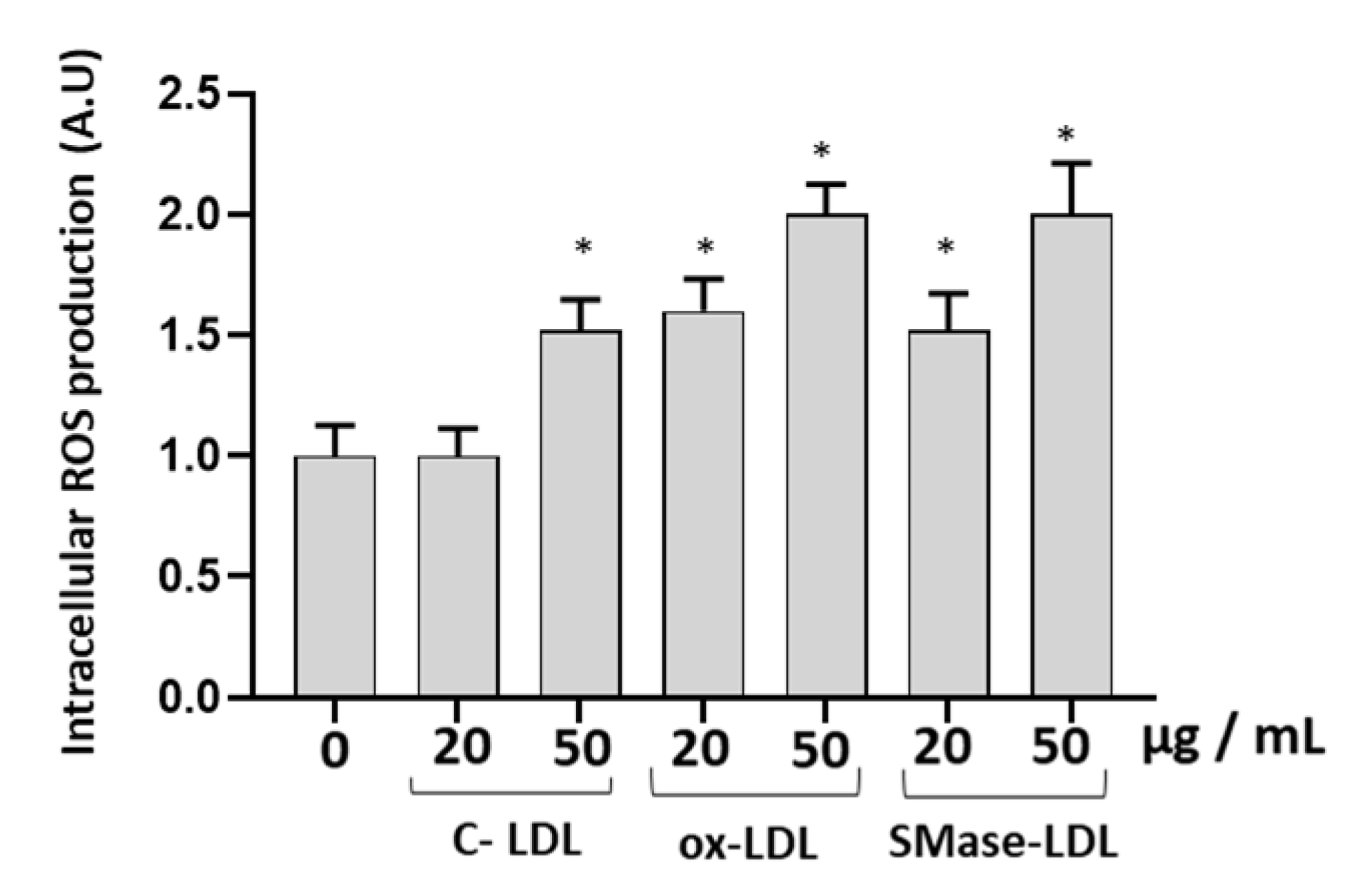
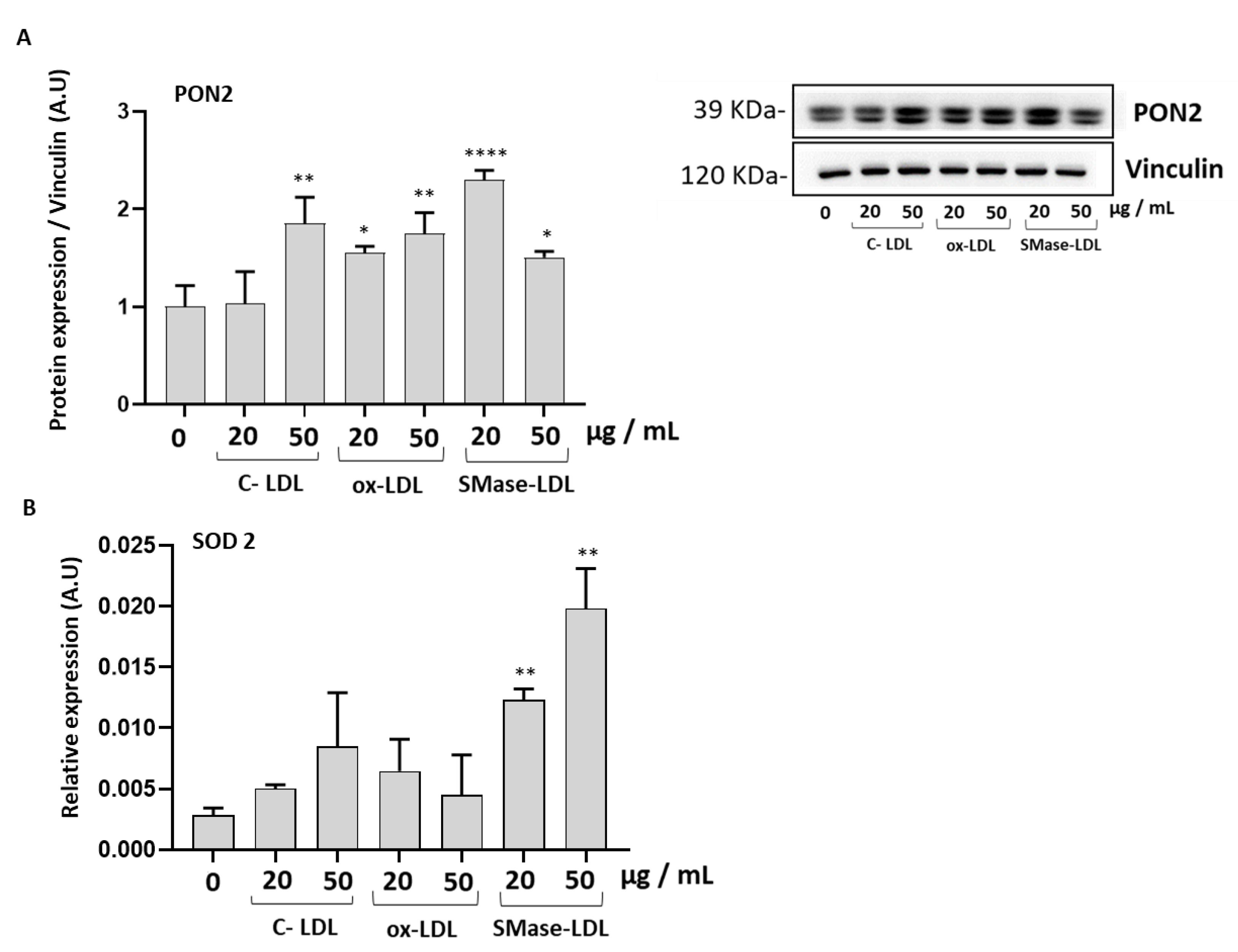
2.3. Modified LDLs Modulate Oxidative Status and Antioxidant Defense of HUVECs
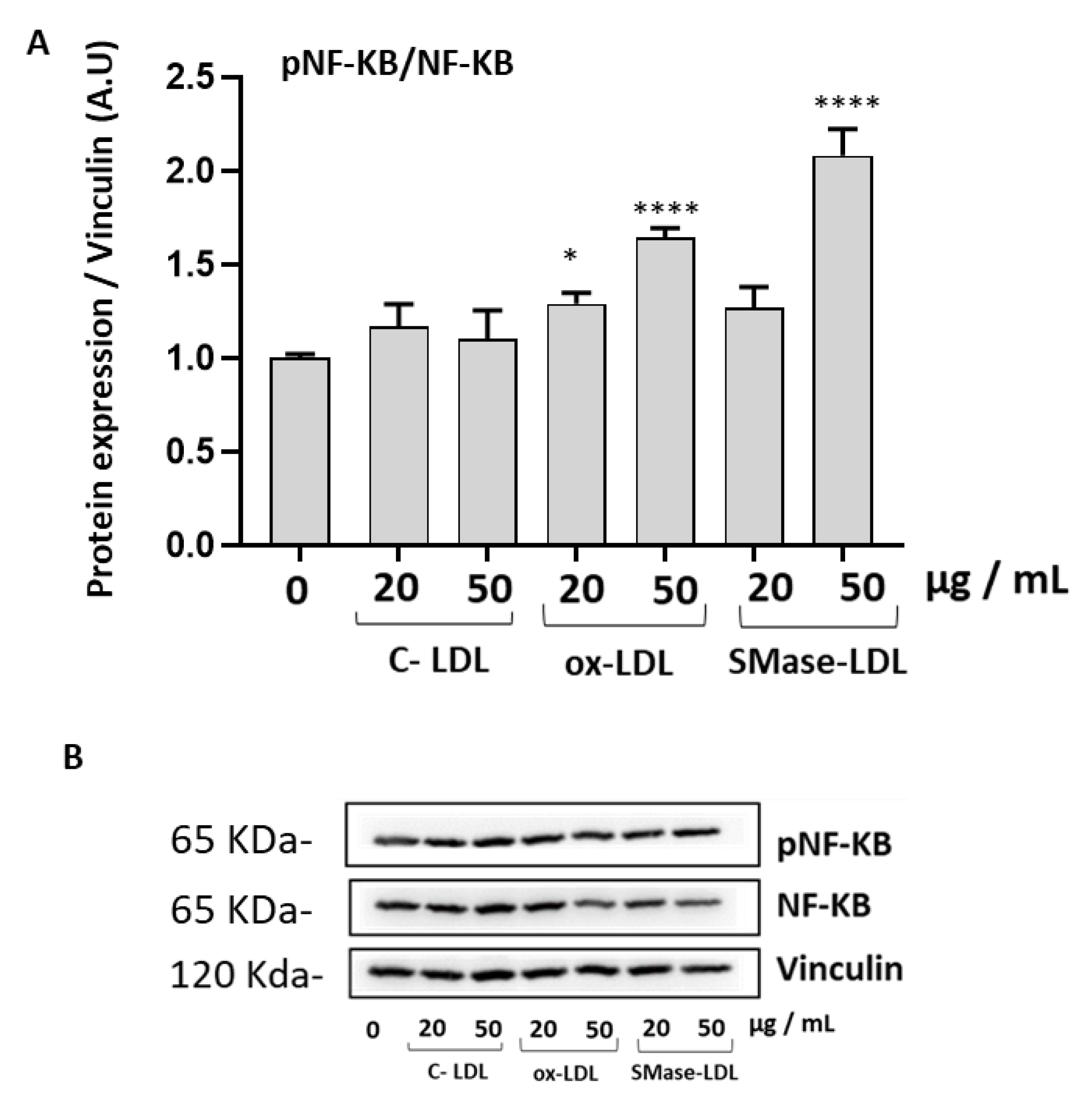
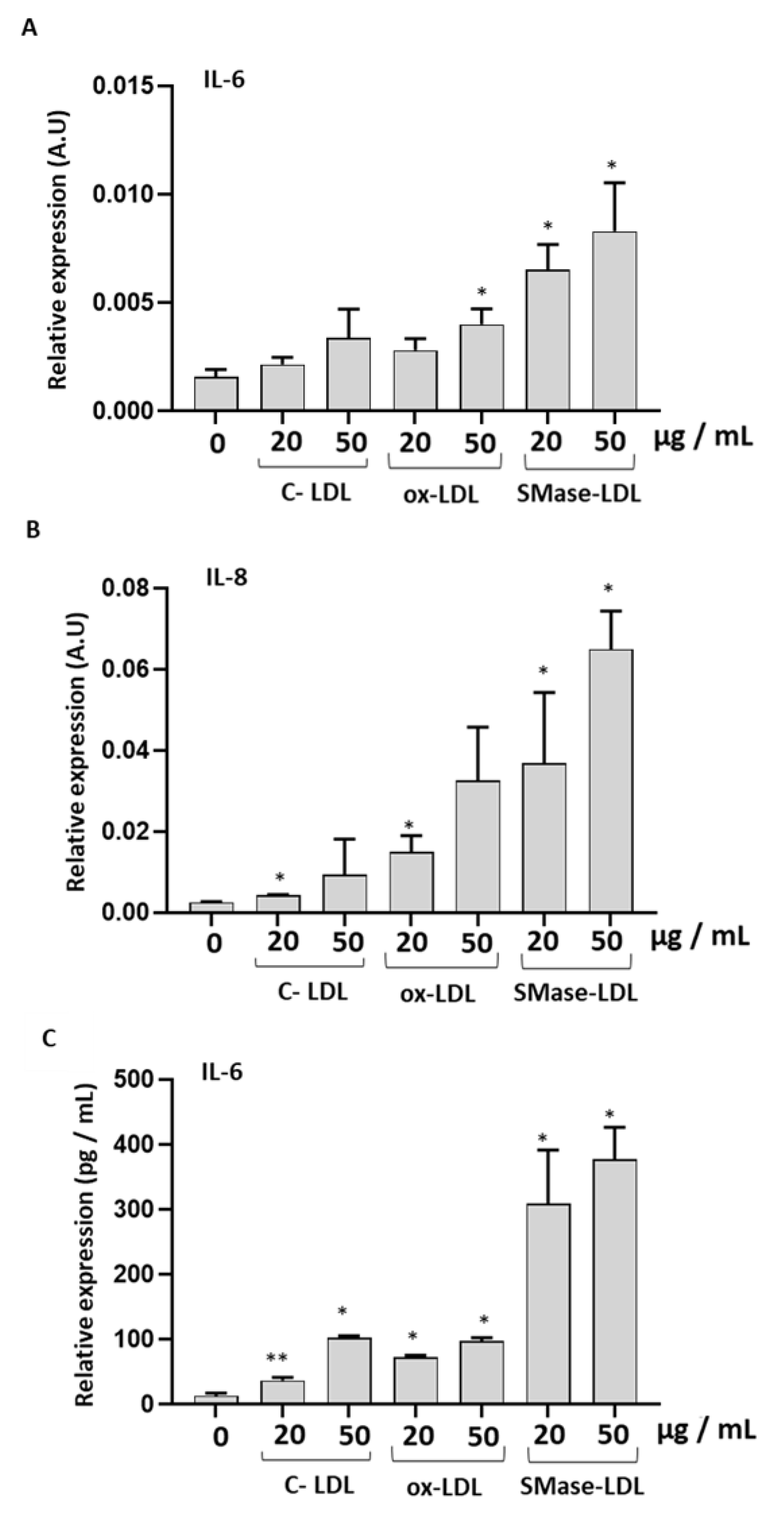
2.4. Ox-LDLs and SMase-LDLs Induce a Proinflammatory Response in HUVECs
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Modifications of LDLs
5.1.1. Oxidation of LDLs
5.1.2. Treatment of LDL Sphingomyelinase
5.2. Measurement of LDL Peroxidation
5.3. Measurement of LDL Aggregation
5.4. Measurement of LDL Hyperchromicity
5.5. Physico-Chemical Properties of Untreated and Modified LDLs
5.6. Intrinsic Fluorescence Spectroscopy
5.7. Cell Culture
5.8. Cell Viability Assay
5.9. Intracellular ROS Levels
5.10. Western Blot
5.11. ELISA Assay
5.12. RNA Isolation and MRNA Expression
5.13. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Ference, B.A.; Ginsberg, H.N.; Graham, I.; Ray, K.K.; Packard, C.J.; Bruckert, E.; Hegele, R.A.; Krauss, R.M.; Raal, F.J.; Schunkert, H.; et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease. 1. Evidence from genetic, epidemiologic, and clinical studies. A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2017, 38, 2459–2472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, R.M.; Pearson, T.A. LDL-cholesterol: A risk factor for coronary artery disease--from epidemiology to clinical trials. Can. J. Cardiol. 1998, 14 (Suppl. SB), 3B–10B. [Google Scholar] [PubMed]
- Hevonoja, T.; Pentikainen, M.O.; Hyvonen, M.T.; Kovanen, P.T.; Ala-Korpela, M. Structure of low density lipoprotein (LDL) particles: Basis for understanding molecular changes in modified LDL. Biochim. Biophys. Acta 2000, 1488, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, D.; Parthasarathy, S.; Carew, T.E.; Khoo, J.C.; Witztum, J.L. Beyond cholesterol. Modifications of low-density lipoprotein that increase its atherogenicity. N. Engl. J. Med. 1989, 320, 915–924. [Google Scholar] [PubMed]
- Sommer, A.; Prenner, E.; Gorges, R.; Stutz, H.; Grillhofer, H.; Kostner, G.M.; Paltauf, F.; Hermetter, A. Organization of phosphatidylcholine and sphingomyelin in the surface monolayer of low density lipoprotein and lipoprotein(a) as determined by time-resolved fluorometry. J. Biol. Chem. 1992, 267, 24217–24222. [Google Scholar] [CrossRef] [PubMed]
- Steinbrecher, U.P.; Witztum, J.L.; Parthasarathy, S.; Steinberg, D. Decrease in reactive amino groups during oxidation or endothelial cell modification of LDL. Correlation with changes in receptor-mediated catabolism. Arteriosclerosis 1987, 7, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esterbauer, H.; Gebicki, J.; Puhl, H.; Jurgens, G. The role of lipid peroxidation and antioxidants in oxidative modification of LDL. Free Radic. Biol. Med. 1992, 13, 341–390. [Google Scholar] [CrossRef]
- Plihtari, R.; Hurt-Camejo, E.; Oorni, K.; Kovanen, P.T. Proteolysis sensitizes LDL particles to phospholipolysis by secretory phospholipase A2 group V and secretory sphingomyelinase. J. Lipid Res. 2010, 51, 1801–1809. [Google Scholar] [CrossRef] [Green Version]
- Marathe, S.; Schissel, S.L.; Yellin, M.J.; Beatini, N.; Mintzer, R.; Williams, K.J.; Tabas, I. Human vascular endothelial cells are a rich and regulatable source of secretory sphingomyelinase. Implications for early atherogenesis and ceramide-mediated cell signaling. J. Biol. Chem. 1998, 273, 4081–4088. [Google Scholar] [CrossRef] [Green Version]
- Schissel, S.L.; Jiang, X.; Tweedie-Hardman, J.; Jeong, T.; Camejo, E.H.; Najib, J.; Rapp, J.H.; Williams, K.J.; Tabas, I. Secretory sphingomyelinase, a product of the acid sphingomyelinase gene, can hydrolyze atherogenic lipoproteins at neutral pH. Implications for atherosclerotic lesion development. J. Biol. Chem. 1998, 273, 2738–2746. [Google Scholar] [CrossRef] [Green Version]
- Kornhuber, J.; Rhein, C.; Muller, C.P.; Muhle, C. Secretory sphingomyelinase in health and disease. Biol. Chem. 2015, 396, 707–736. [Google Scholar] [CrossRef] [PubMed]
- Marathe, S.; Kuriakose, G.; Williams, K.J.; Tabas, I. Sphingomyelinase, an enzyme implicated in atherogenesis, is present in atherosclerotic lesions and binds to specific components of the subendothelial extracellular matrix. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 2648–2658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruuth, M.; Nguyen, S.D.; Vihervaara, T.; Hilvo, M.; Laajala, T.D.; Kondadi, P.K.; Gistera, A.; Lahteenmaki, H.; Kittila, T.; Huusko, J.; et al. Susceptibility of low-density lipoprotein particles to aggregate depends on particle lipidome, is modifiable, and associates with future cardiovascular deaths. Eur. Heart J. 2018, 39, 2562–2573. [Google Scholar] [CrossRef] [Green Version]
- Sneck, M.; Nguyen, S.D.; Pihlajamaa, T.; Yohannes, G.; Riekkola, M.L.; Milne, R.; Kovanen, P.T.; Oorni, K. Conformational changes of apoB-100 in SMase-modified LDL mediate formation of large aggregates at acidic pH. J. Lipid Res. 2012, 53, 1832–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schissel, S.L.; Tweedie-Hardman, J.; Rapp, J.H.; Graham, G.; Williams, K.J.; Tabas, I. Rabbit aorta and human atherosclerotic lesions hydrolyze the sphingomyelin of retained low-density lipoprotein. Proposed role for arterial-wall sphingomyelinase in subendothelial retention and aggregation of atherogenic lipoproteins. J. Clin. Investig. 1996, 98, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.X.; Tabas, I. Sphingomyelinase enhances low density lipoprotein uptake and ability to induce cholesteryl ester accumulation in macrophages. J. Biol. Chem. 1991, 266, 24849–24858. [Google Scholar] [CrossRef]
- Akita, K.; Isoda, K.; Sato-Okabayashi, Y.; Kadoguchi, T.; Kitamura, K.; Ohtomo, F.; Shimada, K.; Daida, H. An Interleukin-6 Receptor Antibody Suppresses Atherosclerosis in Atherogenic Mice. Front. Cardiovasc. Med. 2017, 4, 84. [Google Scholar] [CrossRef] [Green Version]
- Kong, P.; Cui, Z.Y.; Huang, X.F.; Zhang, D.D.; Guo, R.J.; Han, M. Inflammation and atherosclerosis: Signaling pathways and therapeutic intervention. Signal Transduct. Target. Ther. 2022, 7, 131. [Google Scholar] [CrossRef]
- Alouffi, S.; Faisal, M.; Alatar, A.A.; Ahmad, S. Oxidative Modification of LDL by Various Physicochemical Techniques: Its Probable Role in Diabetes Coupled with CVDs. Biomed Res. Int. 2018, 2018, 7390612. [Google Scholar] [CrossRef] [Green Version]
- Hodis, H.N.; Kramsch, D.M.; Avogaro, P.; Bittolo-Bon, G.; Cazzolato, G.; Hwang, J.; Peterson, H.; Sevanian, A. Biochemical and cytotoxic characteristics of an in vivo circulating oxidized low density lipoprotein (LDL-). J. Lipid Res. 1994, 35, 669–677. [Google Scholar] [CrossRef]
- Auge, N.; Garcia, V.; Maupas-Schwalm, F.; Levade, T.; Salvayre, R.; Negre-Salvayre, A. Oxidized LDL-induced smooth muscle cell proliferation involves the EGF receptor/PI-3 kinase/Akt and the sphingolipid signaling pathways. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1990–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, K.; Eisele, T.; Kreusel, U.; Page, M.; Page, S.; Haas, M.; Gerling, A.; Kaltschmidt, C.; Neumann, F.J.; Mackman, N.; et al. Dysregulation of monocytic nuclear factor-kappa B by oxidized low-density lipoprotein. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1901–1909. [Google Scholar] [CrossRef] [PubMed]
- Cominacini, L.; Pasini, A.F.; Garbin, U.; Davoli, A.; Tosetti, M.L.; Campagnola, M.; Rigoni, A.; Pastorino, A.M.; Lo Cascio, V.; Sawamura, T. Oxidized low density lipoprotein (ox-LDL) binding to ox-LDL receptor-1 in endothelial cells induces the activation of NF-kappaB through an increased production of intracellular reactive oxygen species. J. Biol. Chem. 2000, 275, 12633–12638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Zhou, Y.; Nabavi, S.M.; Sahebkar, A.; Little, P.J.; Xu, S.; Weng, J.; Ge, J. Mechanisms of Oxidized LDL-Mediated Endothelial Dysfunction and Its Consequences for the Development of Atherosclerosis. Front. Cardiovasc. Med. 2022, 9, 925923. [Google Scholar] [CrossRef] [PubMed]
- Hajri, T. Effects of oxidized lipids and lipoproteins on cardiac function. Front. Biosci. (Landmark Ed.) 2018, 23, 1822–1847. [Google Scholar] [CrossRef]
- Bismuth, J.; Lin, P.; Yao, Q.; Chen, C. Ceramide: A common pathway for atherosclerosis? Atherosclerosis 2008, 196, 497–504. [Google Scholar] [CrossRef] [Green Version]
- Guarino, A.J.; Lee, S.P.; Tulenko, T.N.; Wrenn, S.P. Aggregation kinetics of low density lipoproteins upon exposure to sphingomyelinase. J. Colloid Interface Sci. 2004, 279, 109–116. [Google Scholar] [CrossRef]
- Rivas-Urbina, A.; Rull, A.; Montoliu-Gaya, L.; Perez-Cuellar, M.; Ordonez-Llanos, J.; Villegas, S.; Sanchez-Quesada, J.L. Low-density lipoprotein aggregation is inhibited by apolipoprotein J-derived mimetic peptide D-[113-122]apoJ. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158541. [Google Scholar] [CrossRef]
- Maziere, C.; Meignotte, A.; Dantin, F.; Conte, M.A.; Maziere, J.C. Oxidized LDL induces an oxidative stress and activates the tumor suppressor p53 in MRC5 human fibroblasts. Biochem. Biophys. Res. Commun. 2000, 276, 718–723. [Google Scholar] [CrossRef]
- Maziere, C.; Salle, V.; Gomila, C.; Maziere, J.C. Oxidized low density lipoprotein increases RANKL level in human vascular cells. Involvement of oxidative stress. Biochem. Biophys. Res. Commun. 2013, 440, 295–299. [Google Scholar] [CrossRef]
- Gliozzi, M.; Scicchitano, M.; Bosco, F.; Musolino, V.; Carresi, C.; Scarano, F.; Maiuolo, J.; Nucera, S.; Maretta, A.; Paone, S.; et al. Modulation of Nitric Oxide Synthases by Oxidized LDLs: Role in Vascular Inflammation and Atherosclerosis Development. Int. J. Mol. Sci. 2019, 20, 3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.K.; Rudney, H. Sphingomyelinase treatment of low density lipoprotein and cultured cells results in enhanced processing of LDL which can be modulated by sphingomyelin. J. Lipid Res. 1992, 33, 1741–1752. [Google Scholar] [CrossRef]
- Boyanovsky, B.; Karakashian, A.; King, K.; Giltiay, N.; Nikolova-Karakashian, M. Uptake and metabolism of low density lipoproteins with elevated ceramide content by human microvascular endothelial cells: Implications for the regulation of apoptosis. J. Biol. Chem. 2003, 278, 26992–26999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uciechowski, P.; Dempke, W.C.M. Interleukin-6: A Masterplayer in the Cytokine Network. Oncology 2020, 98, 131–137. [Google Scholar] [CrossRef]
- Ridker, P.M.; Rane, M. Interleukin-6 Signaling and Anti-Interleukin-6 Therapeutics in Cardiovascular Disease. Circ. Res. 2021, 128, 1728–1746. [Google Scholar] [CrossRef]
- Demuth, K.; Myara, I.; Chappey, B.; Vedie, B.; Pech-Amsellem, M.A.; Haberland, M.E.; Moatti, N. A cytotoxic electronegative LDL subfraction is present in human plasma. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 773–783. [Google Scholar] [CrossRef]
- Oestvang, J.; Bonnefont-Rousselot, D.; Ninio, E.; Hakala, J.K.; Johansen, B.; Anthonsen, M.W. Modification of LDL with human secretory phospholipase A2 or sphingomyelinase promotes its arachidonic acid-releasing propensity. J. Lipid Res. 2004, 45, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Mehta, J.L.; Haider, N.; Zhang, X.; Narula, J.; Li, D. Role of caspases in Ox-LDL-induced apoptotic cascade in human coronary artery endothelial cells. Circ. Res. 2004, 94, 370–376. [Google Scholar] [CrossRef] [Green Version]
- Pettus, B.J.; Chalfant, C.E.; Hannun, Y.A. Ceramide in apoptosis: An overview and current perspectives. Biochim. Biophys. Acta 2002, 1585, 114–125. [Google Scholar] [CrossRef]
- Matsunaga, T.; Kotamraju, S.; Kalivendi, S.V.; Dhanasekaran, A.; Joseph, J.; Kalyanaraman, B. Ceramide-induced intracellular oxidant formation, iron signaling, and apoptosis in endothelial cells: Protective role of endogenous nitric oxide. J. Biol. Chem. 2004, 279, 28614–28624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlong, C.E.; Marsillach, J.; Jarvik, G.P.; Costa, L.G. Paraoxonases-1, -2 and -3: What are their functions? Chem. Biol. Interact. 2016, 259 Pt B, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Bacchetti, T.; Sartini, D.; Pozzi, V.; Cacciamani, T.; Ferretti, G.; Emanuelli, M. Exploring the role of paraoxonase-2 in bladder cancer: Analyses performed on tissue samples, urines and cell cultures. Oncotarget 2017, 8, 28785–28795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morresi, C.; Cianfruglia, L.; Sartini, D.; Cecati, M.; Fumarola, S.; Emanuelli, M.; Armeni, T.; Ferretti, G.; Bacchetti, T. Effect of High Glucose-Induced Oxidative Stress on Paraoxonase 2 Expression and Activity in Caco-2 Cells. Cells 2019, 8, 1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravi, R.; Ragavachetty Nagaraj, N.; Subramaniam Rajesh, B. Effect of advanced glycation end product on paraoxonase 2 expression: Its impact on endoplasmic reticulum stress and inflammation in HUVECs. Life Sci. 2020, 246, 117397. [Google Scholar] [CrossRef] [PubMed]
- Precourt, L.P.; Seidman, E.; Delvin, E.; Amre, D.; Deslandres, C.; Dominguez, M.; Sinnett, D.; Levy, E. Comparative expression analysis reveals differences in the regulation of intestinal paraoxonase family members. Int. J. Biochem. Cell Biol. 2009, 41, 1628–1637. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Yu, J.; Shi, R.; Yan, L.; Yang, T.; Li, Y.; Zhang, Z.; Yu, G.; Bai, Y.; Schuchman, E.H.; et al. Elevation of ceramide and activation of secretory acid sphingomyelinase in patients with acute coronary syndromes. Coron. Artery Dis. 2014, 25, 230–235. [Google Scholar] [CrossRef]
- Gorska, M.; Baranczuk, E.; Dobrzyn, A. Secretory Zn2+-dependent sphingomyelinase activity in the serum of patients with type 2 diabetes is elevated. Horm. Metab. Res. 2003, 35, 506–507. [Google Scholar]
- Wong, M.L.; Xie, B.; Beatini, N.; Phu, P.; Marathe, S.; Johns, A.; Gold, P.W.; Hirsch, E.; Williams, K.J.; Licinio, J.; et al. Acute systemic inflammation up-regulates secretory sphingomyelinase in vivo: A possible link between inflammatory cytokines and atherogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 8681–8686. [Google Scholar] [CrossRef] [Green Version]
- Ke, L.Y.; Chan, H.C.; Chen, C.C.; Lu, J.; Marathe, G.K.; Chu, C.S.; Chan, H.C.; Wang, C.Y.; Tung, Y.C.; McIntyre, T.M.; et al. Enhanced Sphingomyelinase Activity Contributes to the Apoptotic Capacity of Electronegative Low-Density Lipoprotein. J. Med. Chem. 2016, 59, 1032–1040. [Google Scholar] [CrossRef]
- Ferretti, G.; Bacchetti, T.; Masciangelo, S.; Bicchiega, V. Effect of phytosterols on copper lipid peroxidation of human low-density lipoproteins. Nutrition 2010, 26, 296–304. [Google Scholar] [CrossRef]
- Walters, M.J.; Wrenn, S.P. Effect of sphingomyelinase-mediated generation of ceramide on aggregation of low-density lipoprotein. Langmuir 2008, 24, 9642–9647. [Google Scholar] [CrossRef]
- Ferretti, G.; Rabini, R.A.; Bacchetti, T.; Vignini, A.; Salvolini, E.; Ravaglia, F.; Curatola, G.; Mazzanti, L. Glycated low density lipoproteins modify platelet properties: A compositional and functional study. J. Clin. Endocrinol. Metab. 2002, 87, 2180–2184. [Google Scholar] [CrossRef] [PubMed]
- Parasassi, T.; De Stasio, G.; d’Ubaldo, A.; Gratton, E. Phase fluctuation in phospholipid membranes revealed by Laurdan fluorescence. Biophys. J. 1990, 57, 1179–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferretti, G.; Bacchetti, T.; Moroni, C.; Vignini, A.; Nanetti, L.; Curatola, G. Effect of homocysteinylation of low density lipoproteins on lipid peroxidation of human endothelial cells. J. Cell. Biochem. 2004, 92, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Sladowski, D.; Steer, S.J.; Clothier, R.H.; Balls, M. An improved MTT assay. J. Immunol. Methods 1993, 157, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Bacchetti, T.; Campagna, R.; Sartini, D.; Cecati, M.; Morresi, C.; Bellachioma, L.; Martinelli, E.; Rocchetti, G.; Lucini, L.; Ferretti, G.; et al. C. spinosa L. subsp. rupestris Phytochemical Profile and Effect on Oxidative Stress in Normal and Cancer Cells. Molecules 2022, 27, 6488. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C-LDL | ox-LDL | SMase-LDL 50 mU/mL | SMase-LDL 100 mU/mL | |
|---|---|---|---|---|
| Turbidity (absorbance a 450 nm) | 0.291 ± 0.012 | 0.272 ± 0.017 | 0.545 ± 0.021 | 0.797 ± 0.025 ** |
| Ceramide level (µM) | 0.205 ± 0.015 | 8.476 ± 0.352 | 12.1± 0.02 * | 23.843 ± 0.9 ** |
| Fluorescenceintensity (AU) | 461 ± 21 | 128 ± 5 * | 400 ± 15 | 392 ± 13 * |
| Hyperchromicity(AU) | 0.080 ± 0.005 | nd | 0.085± 0.010 | 0.100 ± 0.004 * |
| GP value (Laurdan) | 0.52 ± 0.01 | 0.54 ± 0.03 | 0.48 ± 0.04 | 0.42 ± 0.02 |
| TBARS (nmol MDA/mg protein) | 0.10 ± 0.03 | 26.2 ± 1.3 ** | nd | 0.13 ± 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giuliani, A.; Morresi, C.; Mazzuferi, G.; Bellachioma, L.; Ramini, D.; Sabbatinelli, J.; Olivieri, F.; Bacchetti, T.; Ferretti, G. Effect of Sphingomyelinase-Treated LDLs on HUVECs. Molecules 2023, 28, 2100. https://doi.org/10.3390/molecules28052100
Giuliani A, Morresi C, Mazzuferi G, Bellachioma L, Ramini D, Sabbatinelli J, Olivieri F, Bacchetti T, Ferretti G. Effect of Sphingomyelinase-Treated LDLs on HUVECs. Molecules. 2023; 28(5):2100. https://doi.org/10.3390/molecules28052100
Chicago/Turabian StyleGiuliani, Angelica, Camilla Morresi, Gabriele Mazzuferi, Luisa Bellachioma, Deborah Ramini, Jacopo Sabbatinelli, Fabiola Olivieri, Tiziana Bacchetti, and Gianna Ferretti. 2023. "Effect of Sphingomyelinase-Treated LDLs on HUVECs" Molecules 28, no. 5: 2100. https://doi.org/10.3390/molecules28052100