
Influence of Dietary n-3 Long Chain Polyunsaturated Fatty Acid Intake on Oxylipins in Erythrocytes of Women with Rheumatoid Arthritis
 , , ,
, , , 
Abstract
:1. Introduction
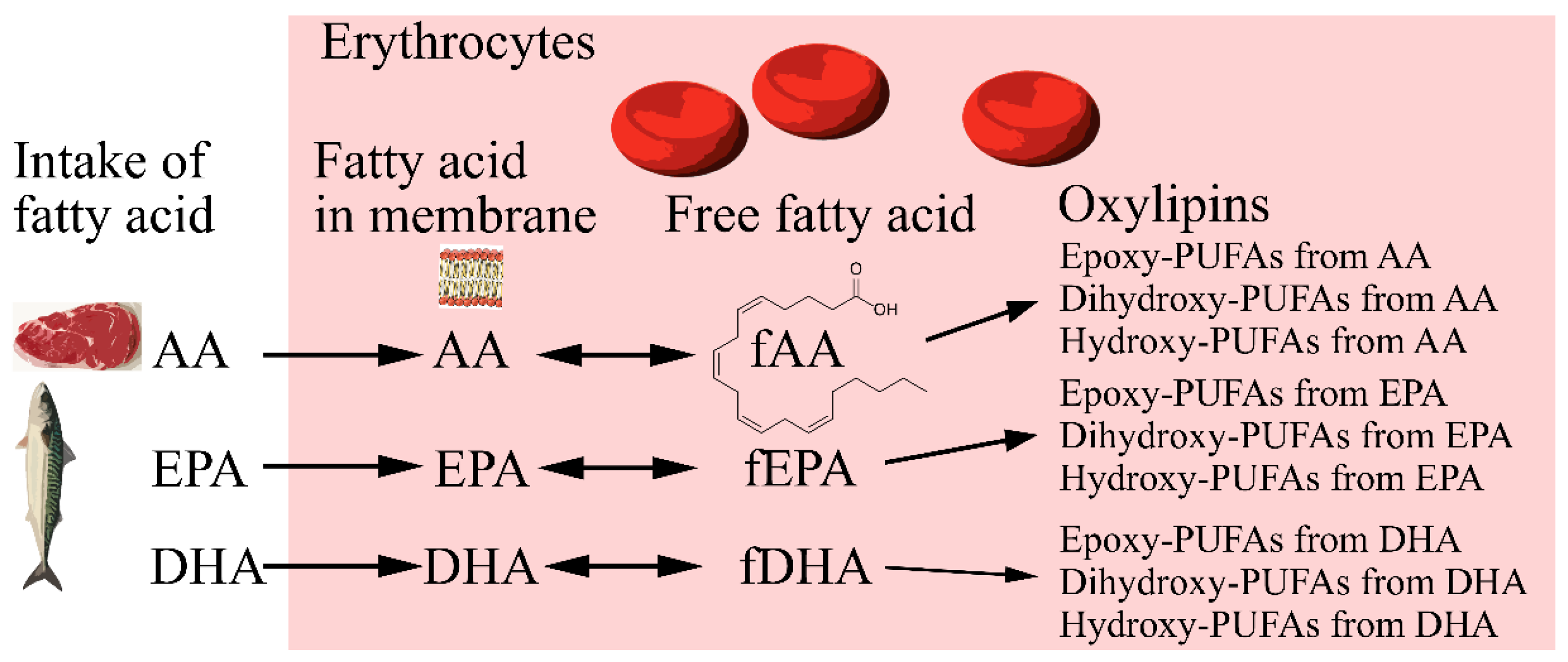
2. Results
2.1. Results from Analysis of Oxylipins and Free Fatty Acids in Erythrocytes
2.2. Correlations between Erythrocyte Oxylipins and Free Fatty Acids in Baseline Samples
2.3. Correlations between Precursor Fatty Acids and Oxylipins in Erythrocytes at Baseline
2.4. Influence of the Dietary Intervention on Free Fatty Acids in Erythrocytes
2.5. Effect of the Intervention and Control Diets on Oxylipin Patterns and Single Oxylipins
3. Discussion
4. Materials and Methods
4.1. Patients and Experimental Design
4.2. Analysis of Oxylipins
4.2.1. Sample Collection and Handling
4.2.2. Lipid Extraction and LC–MS Measurement of Oxylipins
4.3. Statistical Methods
4.3.1. Pre-Processing of Data
4.3.2. Multivariate Methods
4.3.3. Univariate Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Volker, D.; Fitzgerald, P.; Major, G.; Garg, M. Efficacy of fish oil concentrate in the treatment of rheumatoid arthritis. J. Rheumatol. 2000, 27, 2343–2346. [Google Scholar]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Very long-chain n-3 fatty acids and human health: Fact, fiction and the future. Proc. Nutr. Soc. 2018, 77, 52–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostermann, A.I.; West, A.L.; Schoenfeld, K.; Browning, L.M.; Walker, C.G.; Jebb, S.A.; Calder, P.C.; Schebb, N.H. Plasma oxylipins respond in a linear dose-response manner with increased intake of EPA and DHA: Results from a randomized controlled trial in healthy humans. Am. J. Clin. Nutr. 2019, 109, 1251–1263. [Google Scholar] [CrossRef]
- Lundstrom, S.L.; Yang, J.; Kallberg, H.J.; Thunberg, S.; Gafvelin, G.; Haeggstrom, J.Z.; Gronneberg, R.; Grunewald, J.; van Hage, M.; Hammock, B.D.; et al. Allergic asthmatics show divergent lipid mediator profiles from healthy controls both at baseline and following birch pollen provocation. PLoS ONE 2012, 7, e33780. [Google Scholar] [CrossRef]
- Keenan, A.H.; Pedersen, T.L.; Fillaus, K.; Larson, M.K.; Shearer, G.C.; Newman, J.W. Basal omega-3 fatty acid status affects fatty acid and oxylipin responses to high-dose n3-HUFA in healthy volunteers. J. Lipid Res. 2012, 53, 1662–1669. [Google Scholar] [CrossRef] [Green Version]
- Schuchardt, J.P.; Schmidt, S.; Kressel, G.; Willenberg, I.; Hammock, B.D.; Hahn, A.; Schebb, N.H. Modulation of blood oxylipin levels by long-chain omega-3 fatty acid supplementation in hyper- and normolipidemic men. Prostaglandins Leukot Essent Fat. Acids 2014, 90, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Schebb, N.H.; Ostermann, A.I.; Yang, J.; Hammock, B.D.; Hahn, A.; Schuchardt, J.P. Comparison of the effects of long-chain omega-3 fatty acid supplementation on plasma levels of free and esterified oxylipins. Prostaglandins Other Lipid Mediat. 2014, 113–115, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, B.A.; Kim, J.; Kenny, A.; Pedersen, T.L.; Pappan, K.L.; Newman, J.W. Circulating levels of endocannabinoids and oxylipins altered by dietary lipids in older women are likely associated with previously identified gene targets. Biochim. Biophys. Acta 2016, 1861, 1693–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zivkovic, A.M.; Yang, J.; Georgi, K.; Hegedus, C.; Nording, M.L.; O’Sullivan, A.; German, J.B.; Hogg, R.J.; Weiss, R.H.; Bay, C.; et al. Serum oxylipin profiles in IgA nephropathy patients reflect kidney functional alterations. Metabolomics 2012, 8, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Demler, O.V.; Liu, Y.; Luttmann-Gibson, H.; Watrous, J.D.; Lagerborg, K.A.; Dashti, H.; Giulianini, F.; Heath, M.; Camargo, C.A., Jr.; Harris, W.S.; et al. One-Year Effects of Omega-3 Treatment on Fatty Acids, Oxylipins, and Related Bioactive Lipids and Their Associations with Clinical Lipid and Inflammatory Biomarkers: Findings from a Substudy of the Vitamin D and Omega-3 Trial (VITAL). Metabolites 2020, 10, 431. [Google Scholar] [CrossRef] [PubMed]
- Shearer, G.C.; Harris, W.S.; Pedersen, T.L.; Newman, J.W. Detection of omega-3 oxylipins in human plasma and response to treatment with omega-3 acid ethyl esters. J. Lipid Res. 2010, 51, 2074–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, R.; Konkel, A.; Mehling, H.; Blossey, K.; Gapelyuk, A.; Wessel, N.; von Schacky, C.; Dechend, R.; Muller, D.N.; Rothe, M.; et al. Dietary omega-3 fatty acids modulate the eicosanoid profile in man primarily via the CYP-epoxygenase pathway. J. Lipid Res. 2014, 55, 1150–1164. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Dogan, I.; Rothe, M.; Kunz, J.V.; Knauf, F.; Gollasch, M.; Luft, F.C.; Gollasch, B. Hemodialysis and biotransformation of erythrocyte epoxy fatty acids in peripheral tissue. Prostaglandins Leukot Essent Fat. Acids 2022, 181, 102453. [Google Scholar] [CrossRef]
- Nemkov, T.; Reisz, J.A.; Xia, Y.; Zimring, J.C.; D’Alessandro, A. Red blood cells as an organ? How deep omics characterization of the most abundant cell in the human body highlights other systemic metabolic functions beyond oxygen transport. Expert Rev. Proteom. 2018, 15, 855–864. [Google Scholar] [CrossRef]
- Fu, X.; Felcyn, J.R.; Odem-Davis, K.; Zimring, J.C. Bioactive lipids accumulate in stored red blood cells despite leukoreduction: A targeted metabolomics study. Transfusion 2016, 56, 2560–2570. [Google Scholar] [CrossRef]
- Tedeschi, S.K.; Frits, M.; Cui, J.; Zhang, Z.Z.; Mahmoud, T.; Iannaccone, C.; Lin, T.C.; Yoshida, K.; Weinblatt, M.E.; Shadick, N.A.; et al. Diet and Rheumatoid Arthritis Symptoms: Survey Results From a Rheumatoid Arthritis Registry. Arthritis Care Res. 2017, 19, 23225. [Google Scholar] [CrossRef] [Green Version]
- Vadell, A.K.E.; Barebring, L.; Hulander, E.; Gjertsson, I.; Lindqvist, H.M.; Winkvist, A. Anti-inflammatory Diet In Rheumatoid Arthritis (ADIRA)-a randomized, controlled crossover trial indicating effects on disease activity. Am. J. Clin. Nutr. 2020, 111, 1203–1213. [Google Scholar] [CrossRef] [Green Version]
- Lindqvist, H.M.; Gjertsson, I.; Eneljung, T.; Winkvist, A. Influence of Blue Mussel (Mytilus edulis) Intake on Disease Activity in Female Patients with Rheumatoid Arthritis: The MIRA Randomized Cross-Over Dietary Intervention. Nutrients 2018, 10, 481. [Google Scholar] [CrossRef] [Green Version]
- Lindqvist, H.M.; Gjertsson, I.; Andersson, S.; Calder, P.C.; Barebring, L. Influence of blue mussel (Mytilus edulis) intake on fatty acid composition in erythrocytes and plasma phospholipids and serum metabolites in women with rheumatoid arthritis. Prostaglandins Leukot Essent Fat. Acids 2019, 150, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Hulander, E.; Bärebring, L.; Turesson Wadell, A.; Gjertsson, I.; Calder, P.C.; Winkvist, A.; Lindqvist, H.M. Proposed Anti-Inflammatory Diet Reduces Inflammation in Compliant, Weight-Stable Patients with Rheumatoid Arthritis in a Randomized Controlled Crossover Trial. J. Nutr. 2021, 151, 3856–3864. [Google Scholar] [CrossRef] [PubMed]
- Coras, R.; Murillo-Saich, J.D.; Guma, M. Circulating Pro- and Anti-Inflammatory Metabolites and Its Potential Role in Rheumatoid Arthritis Pathogenesis. Cells 2020, 9, 827. [Google Scholar] [CrossRef] [Green Version]
- Markworth, J.F.; Vella, L.; Lingard, B.S.; Tull, D.L.; Rupasinghe, T.W.; Sinclair, A.J.; Maddipati, K.R.; Cameron-Smith, D. Human inflammatory and resolving lipid mediator responses to resistance exercise and ibuprofen treatment. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1281–R1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heitmann, J.; Iversen, L.; Kragballe, K.; Ziboh, V.A. Incorporation of 15-hydroxyeicosatrienoic acid in specific phospholipids of cultured human keratinocytes and psoriatic plaques. Exp. Dermatol. 1995, 4, 74–78. [Google Scholar] [CrossRef]
- Xi, S.; Pham, H.; Ziboh, W.A. 15-hydroxyeicosatrienoic acid (15-HETrE) suppresses epidermal hyperproliferation via the modulation of nuclear transcription factor (AP-1) and apoptosis. Arch. Dermatol. Res. 2000, 292, 397–403. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Q.; Li, J.; Zhong, Y.; Zhang, L.; Huang, Q.; Chen, B.; Mo, M.; Shen, S.; Zhong, Q.; et al. Distinct differences in serum eicosanoids in healthy, enteritis and colorectal cancer individuals. Metabolomics 2018, 14, 4. [Google Scholar] [CrossRef]
- Wang, Y.; Armando, A.M.; Quehenberger, O.; Yan, C.; Dennis, E.A. Comprehensive ultra-performance liquid chromatographic separation and mass spectrometric analysis of eicosanoid metabolites in human samples. J. Chromatogr. A 2014, 1359, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Quehenberger, O.; Dahlberg-Wright, S.; Jiang, J.; Armando, A.M.; Dennis, E.A. Quantitative determination of esterified eicosanoids and related oxygenated metabolites after base hydrolysis. J. Lipid Res. 2018, 59, 2436–2445. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Control | Blue Mussel | ||||||
|---|---|---|---|---|---|---|---|
| Free Fatty Acids/Oxylipin | Pre | Post | P (Pre vs. Post) | Pre | Post | P (Pre vs. Post) | P (Post BM vs. Post C) |
| α-linolenic acid (18:3 n-3) a | 1.26 (0.98,1.74) | 1.35 (0.95,1.61) | 0.738 | 1.33 (1.19,1.6) | 1.37 (1.08,1.73) | 0.563 | 0.761 |
| Free α-linolenic acid (18:3 n-3) b | ND | ND | ND | ND | |||
| 9-HOTrE b | 0.72 (0.48,1.39) | 0.49 (0.28,0.72) | 0.029 | 0.46 (0.30,0.77) | 0.42 (0.31,0.72) | 0.465 | 0.758 |
| Dihomo-ɣ-linolenic acid (20:3 n-6) b | 12.6 (10.2,14.6) | 13.5 (10.6,15.9) | 0.036 | 13.2 (11.5,15.4) | 12.2 (10.1,14.8) | 0.002 | 0.073 |
| Free dihomo-ɣ-linolenic acid (20:3 n-6) b | ND | ND | ND | ND | |||
| 15-HETrE b | 2.26 (1.40,3.15) | 2.26 (1.04,3.29) | 0.670 | 2.11 (1.34,3.14) | 1.87 (1.24,2.75) | 0.024 | 0.171 |
| Arachidonic acid (20:4 n-6) a | 108.6 (91.0,125.1) | 112.8 (98.2,125.3) | 0.128 | 114.8 (105.9,128.6) | 111.0 (100.2,116.5) | 0.039 | 0.465 |
| Free arachidonic acid (20:4 n-6) b | 9820 (1650,13770) | 2890 (1660,7500) | 0.144 | 3940 (1550,7840) | 4280 (1300,7650) | 0.627 | 0.627 |
| 5,6-diHETrE b | 1.12 (0.68,1.87) | 0.57 (0.08,1.62) | 0.201 | 0.90 (0.09,1.90) | 0.36 (0.09,0.87) | 0.012 | 0.108 |
| Adrenic acid (22:4 n-6) | |||||||
| Free adrenic acid (22:4 n-6) b | 3390 (1870,6650) | 1730 (1060,3380) | 0.171 | 2080 (1250,4900) | 2200 (770,3210) | 0.465 | 0.761 |
| Eicosapentaenoic acid (20:5 n-3) a | 10.49 (8.09,12.53) | 8.78 (6.86,10.82) | 0.212 | 10.51 (7.54,12.22) | 12.24 (10.27,14.46) | 0.001 | <0.001 |
| Free eicosapentaenoic acid (20:5 n-3) b | 321 (99,424) | 125 (87,194) | 0.048 | 179 (97,270) | 207 (113,330) | 0.967 | 0.048 |
| Docosahexaenoic acid (22:6 n-3) a | 47.6 (39.3,53.4) | 45.0 (39.6,48.9) | 0.627 | 47.6 (40.4,56.4) | 47.0 (42.9,53.1) | 0.584 | 0.007 |
| Free docosahexaenoic acid (22:6 n-3) b | 2500 (481,4160) | 628 (370,1880) | 0.048 | 1140 (298,2210) | 1120 (350,2110) | 0.761 | 0.236 |
| Model | Scaling | Nr of Lv a | N | R2X [cum] b | R2Y [cum] c | Q2 [cum] d | CV-ANOVA e (p-Value) | Permutation Test (Q2) f | Correct Classified (%C/%M) |
|---|---|---|---|---|---|---|---|---|---|
| PCA | UV | 4 | 23 | 0.694 | 0.063 | ||||
| OPLS-EP mussels vs. control | UVN | 1 + 1 + 0 | 23 | 0.629 | 0.974 | −0.0268 | |||
| OPLS-DA Δmussels vs. Δcontrol g | UV | 1 + 1 + 0 | 46 | 0.588 | 0.208 | −0.142 | 1 | −0.132 h | 48/96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindqvist, H.M.; Winkvist, A.; Gjertsson, I.; Calder, P.C.; Armando, A.M.; Quehenberger, O.; Coras, R.; Guma, M. Influence of Dietary n-3 Long Chain Polyunsaturated Fatty Acid Intake on Oxylipins in Erythrocytes of Women with Rheumatoid Arthritis. Molecules 2023, 28, 717. https://doi.org/10.3390/molecules28020717
Lindqvist HM, Winkvist A, Gjertsson I, Calder PC, Armando AM, Quehenberger O, Coras R, Guma M. Influence of Dietary n-3 Long Chain Polyunsaturated Fatty Acid Intake on Oxylipins in Erythrocytes of Women with Rheumatoid Arthritis. Molecules. 2023; 28(2):717. https://doi.org/10.3390/molecules28020717
Chicago/Turabian StyleLindqvist, Helen M., Anna Winkvist, Inger Gjertsson, Philip C. Calder, Aaron M. Armando, Oswald Quehenberger, Roxana Coras, and Monica Guma. 2023. "Influence of Dietary n-3 Long Chain Polyunsaturated Fatty Acid Intake on Oxylipins in Erythrocytes of Women with Rheumatoid Arthritis" Molecules 28, no. 2: 717. https://doi.org/10.3390/molecules28020717