

Comparative Analysis of Secondary Metabolites in Diplodia corticola Strains with Different Virulence Degrees Associated with Canker and Dieback of Quercus spp.
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
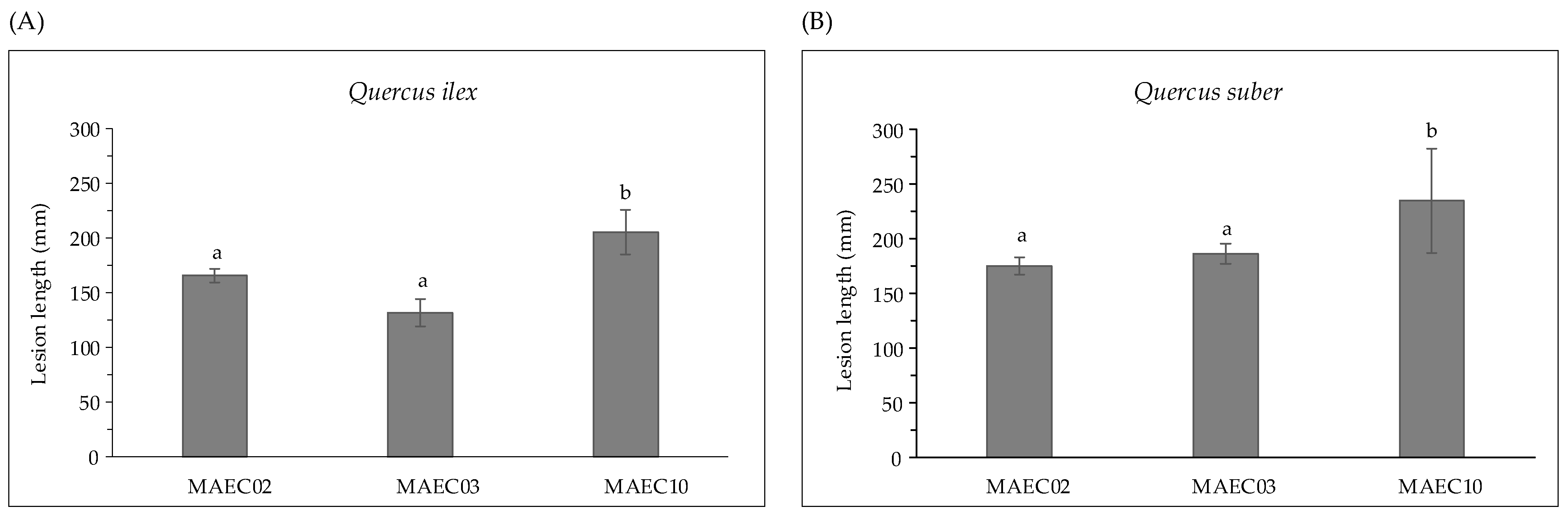
2.1. Pathogenicity Trials
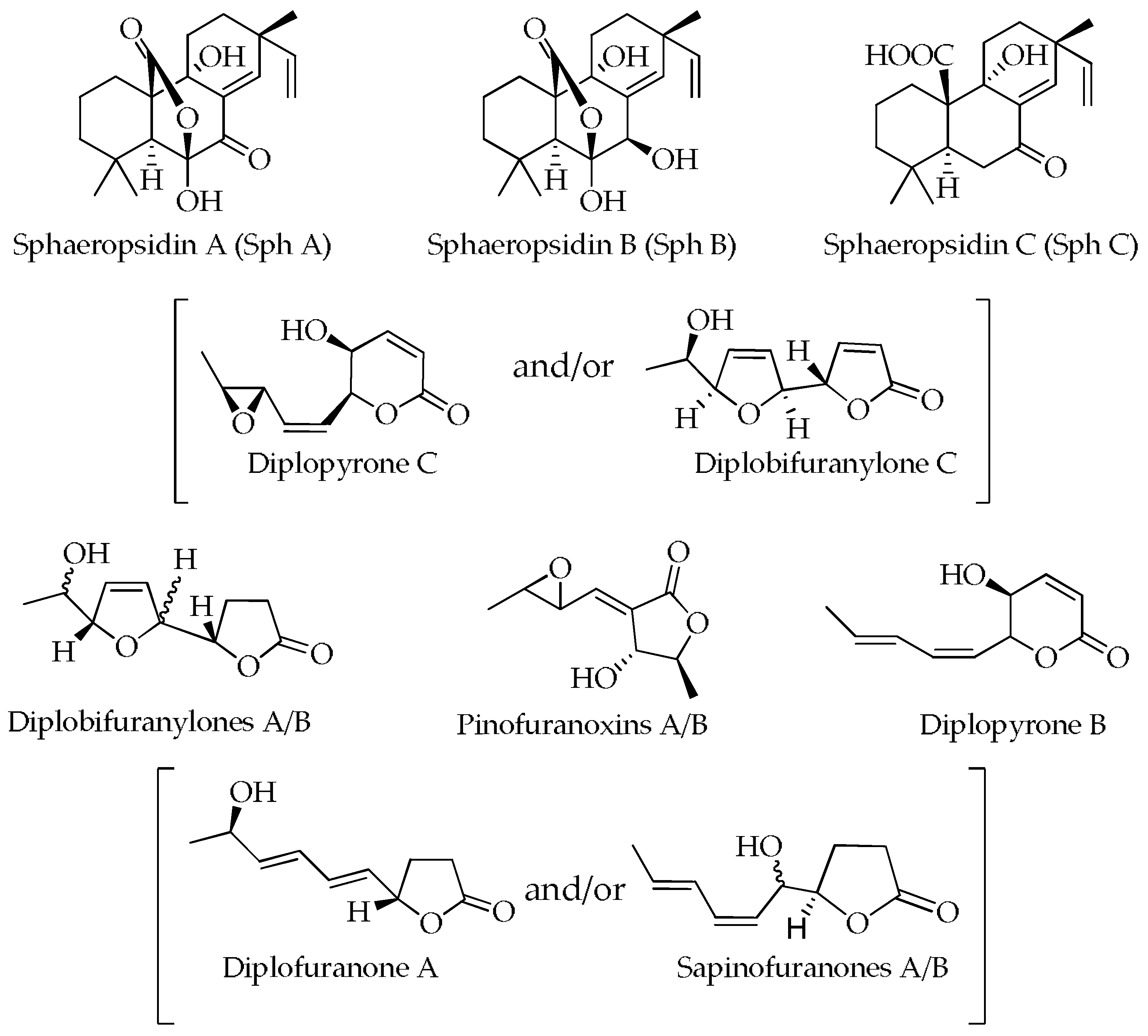
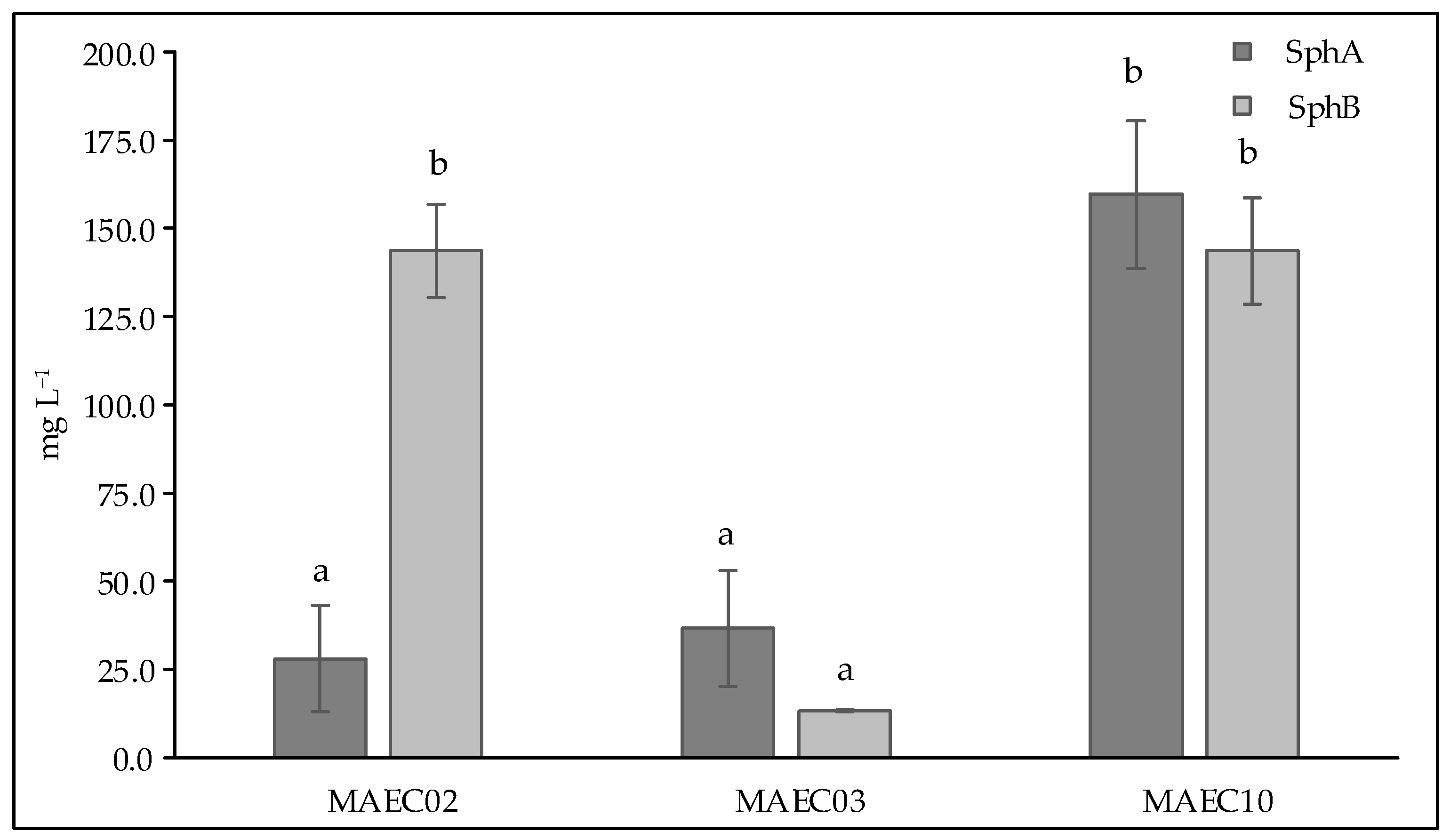
2.2. Qualitative and Quantitative Analyses of Secondary Metabolites
3. Discussion
4. Materials and Methods
4.1. Fungal Strains and Fermentation Conditions
4.2. Pathogenicity Trials
4.3. Extracts Procedures
4.4. Qualitative Analysis
4.5. Quantitative Analysis
4.6. Statisical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kremer, A.; Hipp, A.L. Oaks: An evolutionary success story. N. Phytol. 2020, 226, 987–1011. [Google Scholar] [CrossRef] [PubMed]
- Moradi-Amirabad, Y.; Rahimian, H.; Babaeizad, V.; Denman, S. Brenneria spp. and Rahnella victoriana associated with acute oak decline symptoms on oak and hornbeam in Iran. For. Pathol. 2019, 49, e12535. [Google Scholar] [CrossRef]
- Touhami, I.; Chirino, E.; Aouinti, H.; El Khorchani, A.; Elaieb, M.T.; Khaldi, A.; Nasr, Z. Decline and dieback of cork oak (Quercus suber L.) forests in the Mediterranean basin: A case study of Kroumirie, Northwest Tunisia. J. For. Res. 2020, 31, 1461–1477. [Google Scholar] [CrossRef]
- Brown, N.; Inward, D.J.G.; Jeger, M.; Denman, S. A review of Agrilus biguttatus in UK forests and its relationship with acute oak decline. Forestry 2015, 88, 53–63. [Google Scholar] [CrossRef]
- McConnell, M.E.; Balci, Y. Phytophthora cinnamomi as a contributor to white oak decline in mid-atlantic United States forests. Plant Dis. 2014, 98, 319–327. [Google Scholar] [CrossRef]
- Fan, Z.; Kabrick, J.M.; Spetich, M.A.; Shifley, S.R.; Jensen, R.G. Oak mortality associated with crown dieback and oak borer attack in the Ozark Highlands. For. Ecol. Manag. 2008, 255, 2297–2305. [Google Scholar] [CrossRef]
- Choi, W.I.; Lee, D.H.; Jung, J.B.; Park, Y.S. Oak decline syndrome in Korean forests: History, biology, and prospects for Korean oak wilt. Forests 2022, 13, 964. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Scanu, B.; Maddau, L.; Franceschini, A. Diplodia corticola and Phytophthora cinnamomi: The main pathogens involved in holm oak decline on Caprera Island (Italy). For. Pathol. 2014, 44, 191–200. [Google Scholar] [CrossRef]
- Gallego, F.J.; Perez De Algaba, A.; Fernandez-Escobar, R. Etiology of oak decline in Spain. Eur. J. For. Pathol. 1999, 29, 17–27. [Google Scholar] [CrossRef]
- Thomas, F.M.; Blank, R.; Hartmann, G. Abiotic and biotic factors and their interactions as causes of oak decline in Central Europe. For. Pathol. 2002, 32, 277–307. [Google Scholar] [CrossRef]
- Keča, N.; Koufakis, I.; Dietershagen, J.; Nowakowska, J.A.; Oszako, T. European oak decline phenomenon in relation to climatic changes. Folia For. Pol. Ser. A 2016, 58, 170–177. [Google Scholar] [CrossRef]
- Haavik, L.J.; Billings, S.A.; Guldin, J.M.; Stephen, F.M. Emergent insects, pathogens and drought shape changing patterns in oak decline in North America and Europe. For. Ecol. Manag. 2015, 354, 190–205. [Google Scholar] [CrossRef]
- Denman, S.; Brown, N.; Kirk, S.; Jeger, M.; Webber, J. A description of the symptoms of acute oak decline in Britain and a comparative review on causes of similar disorders on oak in Europe. Forestry 2014, 87, 535–551. [Google Scholar] [CrossRef]
- Lynch, S.C.; Eskalen, A.; Zambino, P.J.; Mayorquin, J.S.; Wang, D.H. Identification and pathogenicity of Botryosphaeriaceae species associated with coast live oak (Quercus agrifolia) decline in southern California. Mycologia 2013, 105, 125–140. [Google Scholar] [CrossRef]
- Mahamedi, A.E.; Phillips, A.J.L.; Lopes, A.; Djellid, Y.; Arkam, M.; Eichmeier, A.; Zitouni, A.; Alves, A.; Berraf-Tebbal, A. Diversity, distribution and host association of Botryosphaeriaceae species causing oak decline across different forest ecosystems in Algeria. Eur. J. Plant Pathol. 2020, 158, 745–765. [Google Scholar] [CrossRef]
- Panzavolta, T.; Panichi, A.; Bracalini, M.; Croci, F.; Ginetti, B.; Ragazzi, A.; Tiberi, R.; Moricca, S. Dispersal and propagule pressure of Botryosphaeriaceae species in a declining oak stand is affected by insect vectors. Forests 2017, 8, 228. [Google Scholar] [CrossRef]
- Ferreira, S.L.; Stauder, C.M.; Martin, D.K.H.; Kasson, M.T. Morphological and phylogenetic resolution of Diplodia corticola and D. quercivora, emerging canker pathogens of oak (Quercus spp.), in the United States. Plant Dis. 2021, 105, 1298–1307. [Google Scholar] [CrossRef]
- Macon, D.; Schohr, T.; Schmidt, D.; Garbelotto, M. Recent blue oak mortality on Sierra Nevada foothill rangelands may be linked to drought, climate change. Calif. Agric. 2020, 74, 71–72. [Google Scholar] [CrossRef]
- Jiranek, J.; Miller, I.F.; An, R.; Bruns, E.; Metcalf, C.J.E. Mechanistic models to meet the challenge of climate change in plant-pathogen systems. Philos. Trans. R. Soc. B Biol. Sci. 2023, 378, 20220017. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef]
- Moricca, S.; Disease, P. Endemic and emerging pathogens threatening cork oak trees: Management options for conserving a unique forest ecosystem. Plant Dis. 2011, 100, 2184–2193. [Google Scholar] [CrossRef] [PubMed]
- Linaldeddu, B.T.; Franceschini, A.; Alves, A.; Phillips, A.J.J.L. Diplodia quercivora sp. nov.: A new species of Diplodia found on declining Quercus canariensis trees in Tunisia. Mycologia 2013, 105, 1266–1274. [Google Scholar] [CrossRef]
- Smahi, H.; Belhoucine-Guezouli, L.; Berraf-Tebbal, A.; Chouih, S.; Arkam, M.; Franceschini, A.; Linaldeddu, B.T.; Phillips, A.J.L. Molecular characterization and pathogenicity of Diplodia corticola and other Botryosphaeriaceae species associated with canker and dieback of Quercus suber in Algeria. Mycosphere 2017, 8, 1261–1272. [Google Scholar] [CrossRef]
- Masi, M.; Evidente, A. Sphaeropsidin A: A pimarane diterpene with interesting biological activities and promising practical applications. ChemBioChem 2021, 22, 3263–3269. [Google Scholar] [CrossRef]
- Salvatore, M.M.; Di Lelio, I.; DellaGreca, M.; Nicoletti, R.; Salvatore, F.; Russo, E.; Volpe, G.; Becchimanzi, A.; Mahamedi, A.E.; Berraf-Tebbal, A.; et al. Secondary metabolites, including a new 5,6-dihydropyran-2-one, produced by the fungus Diplodia corticola. Aphicidal activity of the main metabolite, sphaeropsidin A. Molecules 2022, 27, 2327. [Google Scholar] [CrossRef] [PubMed]
- Möbius, N.; Hertweck, C. Fungal phytotoxins as mediators of virulence. Curr. Opin. Plant Biol. 2009, 12, 390–398. [Google Scholar] [CrossRef]
- Masi, M.; Maddau, L.; Linaldeddu, B.T.; Cimmino, A.; D’Amico, W.; Scanu, B.; Evidente, M.; Tuzi, A.; Evidente, A. Bioactive secondary metabolites produced by the oak pathogen Diplodia corticola. J. Agric. Food Chem. 2016, 64, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Félix, C.; Pinto, G.; Amaral, J.; Fernandes, I.; Alves, A.; Esteves, A.C. Strain-related pathogenicity in Diplodia corticola. For. Pathol. 2017, 47, e12366. [Google Scholar] [CrossRef]
- Ellestad, G.A.A.; Kunstmann, M.P.P.; Mirando, P.; Morton, G.O.O. Structures of fungal diterpene antibiotics LL-S491β and -γ. J. Am. Chem. Soc. 1972, 94, 6206–6208. [Google Scholar] [CrossRef]
- Evidente, A.; Sparapano, L.; Fierro, O.; Bruno, G.; Giordano, F.; Motta, A. Sphaeropsidins B and C, phytotoxic pimarane diterpenes from Sphaeropsis sapinea f. sp. cupressi and Diplodia mutila. Phytochemistry 1997, 4, 705–713. [Google Scholar] [CrossRef]
- Kawahara, K.; Iida, H.; Yokota, A. Detection of 2-hydroxy-fatty acids and 2-hydroxy-fatty acid-containing ceramides in a gliding marine bacterium Aureispira marina. J. Gen. Appl. Microbiol. 2021, 67, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Tannous, J.; Labbe, J.; Keller, N.P. Identifying fungal secondary metabolites and their role in plant pathogenesis. In Plant-Pathogen Interactions; Springer Nature: Berlin/Heidelberg, Germany, 2023; Volume 2659, pp. 193–218. ISBN 9781071631591. [Google Scholar]
- Pontes, J.G.D.M.; Fernandes, L.S.; Dos Santos, R.V.; Tasic, L.; Fill, T.P. Virulence factors in the phytopathogen-host interactions: An overview. J. Agric. Food Chem. 2020, 68, 7555–7570. [Google Scholar] [CrossRef] [PubMed]
- Macheleidt, J.; Mattern, D.J.; Fischer, J.; Netzker, T.; Weber, J.; Schroeckh, V.; Valiante, V.; Brakhage, A.A. Regulation and role of fungal secondary metabolites. Annu. Rev. Genet. 2016, 50, 371–392. [Google Scholar] [CrossRef]
- Fox, E.M.; Howlett, B.J. Secondary metabolism: Regulation and role in fungal biology. Curr. Opin. Microbiol. 2008, 11, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Orozco, R.; Kithsiri Wijeratne, E.M.; Espinosa-Artiles, P.; Leslie Gunatilaka, A.A.; Patricia Stock, S.; Molnár, I. Biosynthesis of the cyclooligomer depsipeptide bassianolide, an insecticidal virulence factor of Beauveria bassiana. Fungal Genet. Biol. 2009, 46, 353–364. [Google Scholar] [CrossRef]
- Pusztahelyi, T.; Holb, I.J.; Pócsi, I. Secondary metabolites in fungus-plant interactions. Front. Plant Sci. 2015, 6, 573. [Google Scholar] [CrossRef]
- Ribera, A.E.; Zuñiga, G. Induced plant secondary metabolites for phytopatogenic fungi control: A review. J. Soil Sci. Plant Nutr. 2012, 12, 893–911. [Google Scholar] [CrossRef]
- Evidente, A.; Sparapano, L.; Motta, A.; Giordano, F.; Fierro, O.; Frisullo, S. A phytotoxic pimarane diterpene of Sphaeropsis sapinea f. sp. cupressi, the pathogen of a canker disease of cypress. Phytochemistry 1996, 42, 1541–1546. [Google Scholar]
- Weber, R.W.S.; Kappe, R.; Paululat, T.; Mösker, E.; Anke, H. Anti-Candida metabolites from endophytic fungi. Phytochemistry 2007, 68, 886–892. [Google Scholar] [CrossRef]
- Roscetto, E.; Masi, M.; Esposito, M.; Di Lecce, R.; Delicato, A.; Maddau, L.; Calabro, V.; Evidente, A.; Catania, M.R. Anti-biofilm activity of the fungal phytotoxin sphaeropsidin A against clinical isolates of antibiotic-resistant bacteria. Toxins 2020, 12, 444. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Fiore, M.; Spanu, E.; Franceschini, A.; Maddau, L. Diplofuranones A and B, two further new 4-monosubstituted 2(3H)-dihydrofuranones produced by Diplodia corticola, a fungus pathogen of cork oak. Arkivoc 2007, 7, 318–328. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Fiore, M.; Spanu, E.; Franceschini, A.; Maddau, L. Diplobifuranylones A and B, 5′-monosubstituted tetrahydro-2H-bifuranyl-5-ones produced by Diplodia corticola, a fungus pathogen of cork oak. J. Nat. Prod. 2006, 69, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Vestal, J.R.; White, D.C. Lipid analysis in microbial ecology: Quantitative approaches to the study of microbial communities. Bioscience 1989, 39, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Miller, J. Statistics and Chemometrics for Analytical Chemistry, 4th ed.; Pearson Education: Harlow, UK, 2018; ISBN 0-131-29192-0. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Rt (Min) | Formula | Exact Mass (U) 1 | Diagnostic Ions m/z (Calcd. [M + H]+ or [M + Na]+) |
|---|---|---|---|---|
| Unknown1 | 3.16 | - | 216 | 455.1843 [2M + Na]+, 255.0631 [M + K]+, 239.0879 [M + Na]+, 217.1052 [M + H]+ |
| Unknown2 | 3.26 | - | 214 | 451.1557 [2M + Na]+, 429.1734 [2M + H]+, 253.0468 [M + K]+, 237.0735 [M + Na]+, 215.0915 [M + H]+, 197.0808 [M − OH]+, 179.0704 [M − OH − H2O]+ |
| Diplobifuranylones A/B 2 | 4.24 | C10H14O4 | 198.0892 | 435.1580 [2M + K]+, 419.1661 [2M + Na]+, 397.1838 [2M + H]+, 237.0535 [M + K]+, 221.0784 [M + Na]+, 199.0965 [M + H]+ (199.0970), 181.0855 [M − OH]+, 155.0706 [M + H − CO2]+ |
| Diplopyrone C and/or diplobifuranylone C | 4.85 | C10H12O4 | 196.0736 | 415.1359 [2M + Na]+, 235.0392 [M + K]+, 219.0627 [M + Na]+, 197.0790 [M + H]+ (197.0814), 179.0704 [M − OH]+ |
| Pinofuranoxins A/B 2 | 6.39 | C9H12O4 | 184.0736 | 391.1367 [2M + Na]+, 207.0628 [M + Na]+, 185.0810 [M + H]+ (185.0814) |
| Unknown3 | 7.61 | - | 168 | 359.1488 [2M + Na]+, 191.0679 [M + Na]+, 169.0860 [M + H]+ |
| Diplofuranone A and/or sapinofuranones A/B | 9.05 | C10H14O3 | 182.0943 | 387.1752 [2M + Na]+, 221.0560 [M + K]+, 205.0831 [M + Na]+, 183.1026 [M + H]+ (183.1021), 165.0908 [M − OH]+ |
| Unknown4 | 9.86 | - | 350 | 723.4011 [2M + Na]+, 389.1783 [M + K]+, 373.1975 [M + Na]+, 333.2036 [M − OH]+ |
| Diplopyrone B | 13.01 | C10H12O3 | 180.0786 | 383.1443 [2M + Na]+, 219.0405 [M + K]+, 203.0678 [M + Na]+, 181.0857 [M + H]+ (181.0865), 163.0756 [M − OH]+ |
| Unknown5 | 15.63 | - | 310 | 643.4107 [2M + Na]+, 349.1797 [M + K]+, 333.2010 [M + Na]+, 293.2083 [M − OH]+ |
| Unknown6 | 17.65 | - | 314 | 651.2913 [2M + Na]+, 357.2020 [M + K]+, 337.1404 [M + Na]+, 297.1480 [M − OH]+ |
| Unknown7 | 20.87 | - | 320 | 663.4526 [2M + Na]+, 359.2082 [M + K]+, 343.2220 [M + Na]+, 303.2299 [M − OH]+, 285.2197 [M – OH − H2O]+ |
| Sph C | 22.25 | C20H28O4 | 332.1988 | 1019.5783 [3M + Na]+, 687.3826 [2M + Na]+, 665.4014 [2M + H]+, 355.1871 [M + Na]+, 333.2056 [M + H]+ (333.2066), 315.1947 [M − OH]+ |
| Sph B | 24.01 | C20H28O5 | 348.1937 | 735.3452 [2M + K]+, 719.3727 [2M + Na]+, 387.1557 [M + K]+, 371.1819 [M + Na]+, 349.2001 [M + H]+ (349.2015), 331.1897 [M − OH]+, 313.1794 [M – OH − H2O]+ |
| Unknown8 | 25.80 | - | 312 | 663.3997 [2M + K]+, 647.4413 [2M + Na]+, 625.4603 [2M + H]+, 351.1787 [M + K]+, 335.2167 [M + Na]+, 295.2249 [M − OH]+, 277.2143 [M – OH − H2O]+ |
| Unknown9 | 26.16 | - | 328 | 695.3066 [2M + K]+, 679.3191 [2M + Na]+, 367.1393 [M + K]+, 351.1557 [M + Na]+, 311.1631 [M − OH]+ |
| Unknown10 | 29.96 | - | 320 | 679.4442 [2M + K]+, 663.4514 [2M + Na]+, 359.2104 [M + K]+, 343.2217 [M + Na]+, 303.2289 [M − OH]+, 285.2192 [M – OH − H2O]+ |
| Sph A | 32.30 | C20H26O5 | 346.1780 | 715.3422 [2M + Na]+, 385.1400 [M + K]+, 369.1672 [M + Na]+, 347.1852 [M + H]+ (347.1858), 329.1745 [M − OH]+ |
| Unknown11 | 34.79 | - | 300 | 623.3314 [2M + Na]+, 601.3183 [2M + H]+, 323.1614 [M + Na]+, 301.1483 [M + H]+, 283.1690 [M − OH]+ |
| Hydroxylinolenic acid | 35.96 | C18H30O3 | 294.2195 | 627.3961 [2M + K]+, 611.4226 [2M + Na]+, 589.4403 [2M + H]+, 333.1801 [M + K]+, 317.2070 [M + Na]+ (317.2093), 277.2147 [M − OH]+ |
| Unknown12 | 37.94 | - | 312 | 647.4414 [2M + Na]+, 351.1883 [M + K]+, 335.2171 [M + Na]+, 313.2352 [M + H]+, 295.2244 [M − OH]+, 277.2139 [M − OH − H2O]+ |
| Hydroxylinoleic acid | 40.41 | C18H32O3 | 296.2326 | 615.4529 [2M + Na]+, 335.1944 [M + K]+, 319.2223 [M + Na] + (319.2249), 279.2306 [M − OH]+ |
| Unknown13 | 42.21 | - | 565 | 1153.8128 [2M + Na]+, 604.3767 [M + K]+, 588.4040 [M + Na]+, 566.4222 [M + H]+ |
| Unknown14 | 43.41 | - | 434 | 473.2463 [M + K]+, 457.2792 [M + Na]+, 435.2510 [M + H]+ |
| Compound | Rt (Min) | MW | CE02 | CE03 | CE10 | ME02 | ME03 | ME10 |
|---|---|---|---|---|---|---|---|---|
| Unknown1 | 3.16 | 216 | - | - | - | + | + | - |
| Unknown2 | 3.26 | 214 | + | + | + | + | - | + |
| Diplobifuranylones A/B 1 | 4.24 | 198 | + | + | - | + | + | - |
| Diplopyrone C and/or diplobifuranylone C | 4.85 | 196 | + | + | - | + | - | - |
| Pinofuranoxins A/B 1 | 6.39 | 184 | + | - | - | - | - | - |
| Unknown3 | 7.61 | 168 | + | - | - | - | - | - |
| Diplofuranone A and/or Sapinofuranones A/B | 9.05 | 182 | + | + | - | + | - | - |
| Unknown4 | 9.86 | 350 | - | - | + | - | - | + |
| Diplopyrone B | 13.01 | 180 | - | - | + | - | - | + |
| Unknown5 | 15.63 | 310 | - | - | - | + | + | + |
| Unknown6 | 17.65 | 314 | - | - | + | - | - | + |
| Unknown7 | 20.87 | 320 | - | - | - | - | - | + |
| Sph C | 22.25 | 332 | + | + | + | + | + | + |
| Sph B | 24.01 | 348 | + | + | + | + | + | + |
| Unknown8 | 25.80 | 312 | - | - | - | + | + | + |
| Unknown9 | 26.16 | 328 | - | - | + | - | - | - |
| Unknown10 | 29.96 | 320 | - | - | - | - | - | + |
| Sph A | 32.30 | 346 | + | + | + | + | + | + |
| Unknown11 | 34.79 | 300 | - | - | + | - | - | - |
| Hydroxylinolenic acid | 35.96 | 294 | - | - | - | + | + | + |
| Unknown12 | 37.94 | 312 | - | - | - | - | + | + |
| Hydroxylinoleic acid | 40.41 | 296 | - | - | - | + | + | + |
| Unknown13 | 42.21 | 565 | - | - | - | + | + | + |
| Unknown14 | 43.41 | 434 | - | - | - | + | + | - |
| Metabolite | Wavelength (nm) | Rt (Min) | Intercept | Slope | R2 | LOD (mg L−1) | LOQ (mg L−1) |
|---|---|---|---|---|---|---|---|
| Sph A | 240 | 32.30 | 0.999 | 13.05 | 39.5 | ||
| Sph B | 220 | 24.01 | 0.999 | 44.23 | 134.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvatore, M.M.; Russo, M.T.; Nicoletti, R.; Mahamedi, A.E.; Berraf-Tebbal, A.; Cimmino, A.; Masi, M.; Andolfi, A. Comparative Analysis of Secondary Metabolites in Diplodia corticola Strains with Different Virulence Degrees Associated with Canker and Dieback of Quercus spp. Molecules 2023, 28, 6302. https://doi.org/10.3390/molecules28176302
Salvatore MM, Russo MT, Nicoletti R, Mahamedi AE, Berraf-Tebbal A, Cimmino A, Masi M, Andolfi A. Comparative Analysis of Secondary Metabolites in Diplodia corticola Strains with Different Virulence Degrees Associated with Canker and Dieback of Quercus spp. Molecules. 2023; 28(17):6302. https://doi.org/10.3390/molecules28176302
Chicago/Turabian StyleSalvatore, Maria Michela, Maria Teresa Russo, Rosario Nicoletti, Alla Eddine Mahamedi, Akila Berraf-Tebbal, Alessio Cimmino, Marco Masi, and Anna Andolfi. 2023. "Comparative Analysis of Secondary Metabolites in Diplodia corticola Strains with Different Virulence Degrees Associated with Canker and Dieback of Quercus spp." Molecules 28, no. 17: 6302. https://doi.org/10.3390/molecules28176302