

Quantitation of Tissue Amyloid via Fluorescence Spectroscopy Using Controlled Concentrations of Thioflavin-S

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
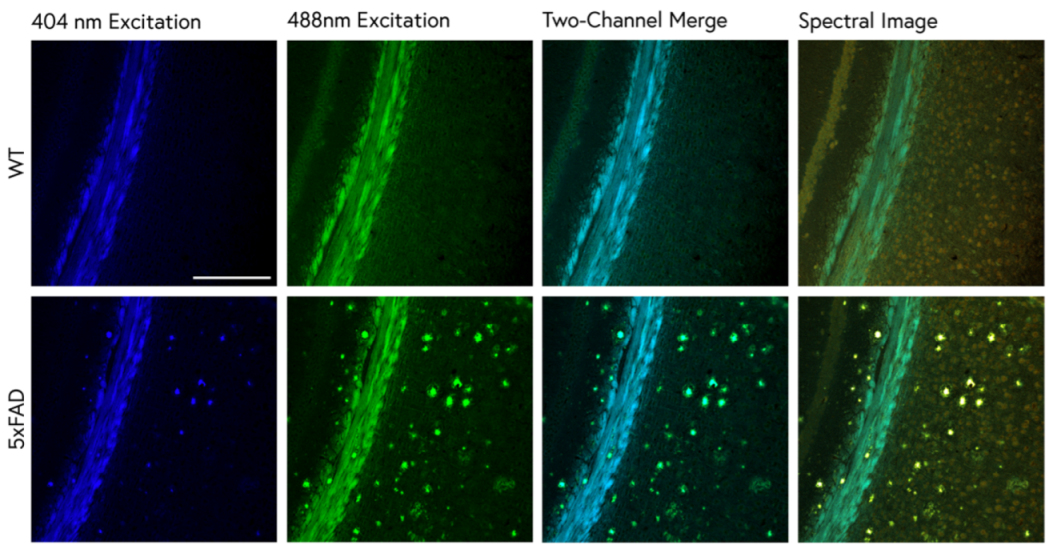
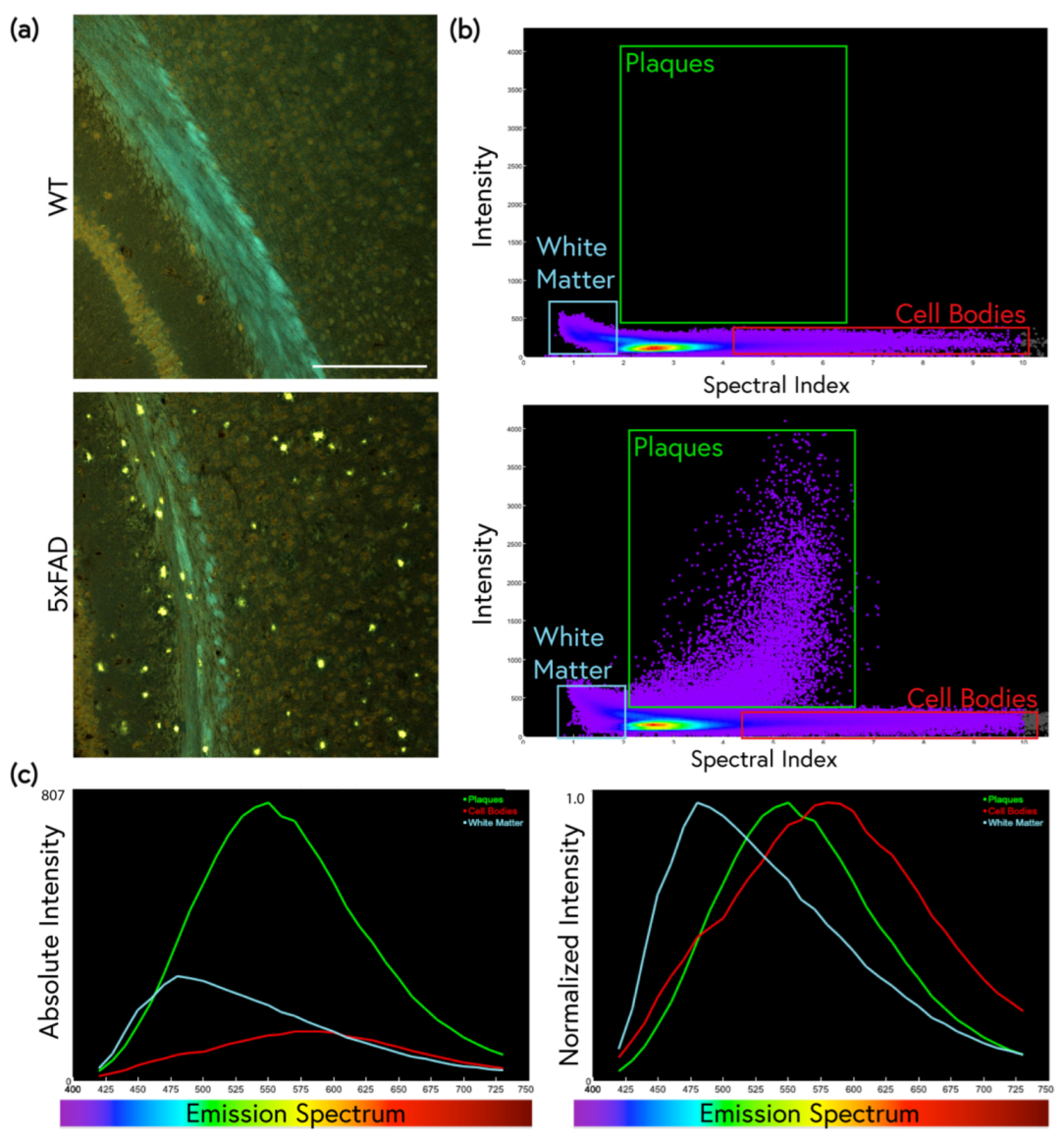
2.1. Traditional ThS Staining Protocols Revealed Conventional Features in 5xFAD Brain
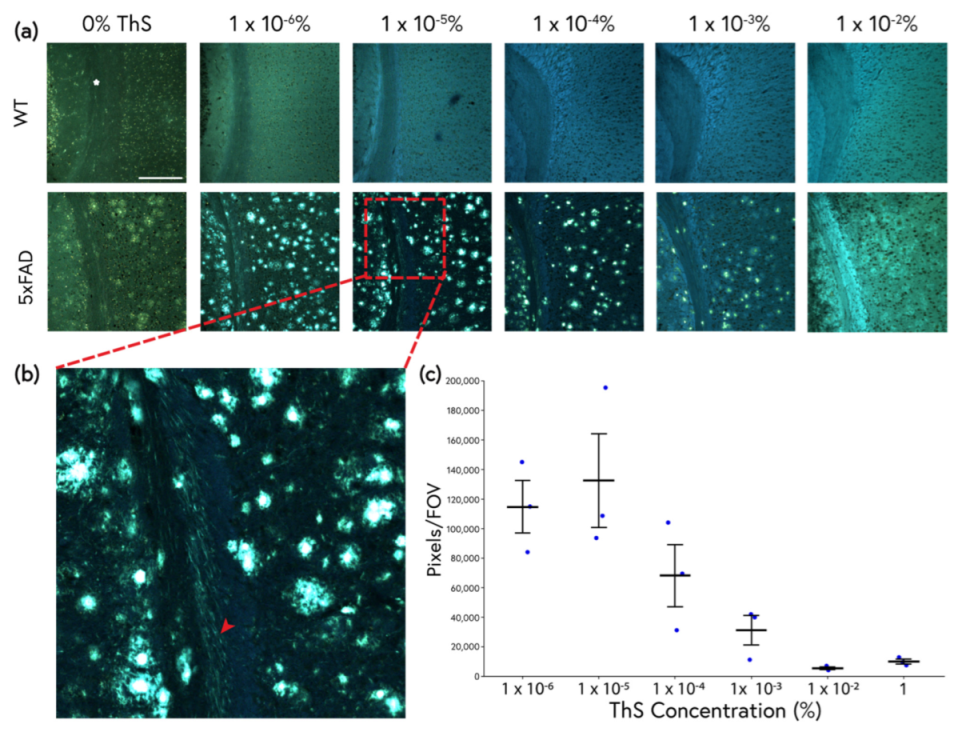
2.2. An Improved ThS Staining Protocol for Subtle Amyloid Detection
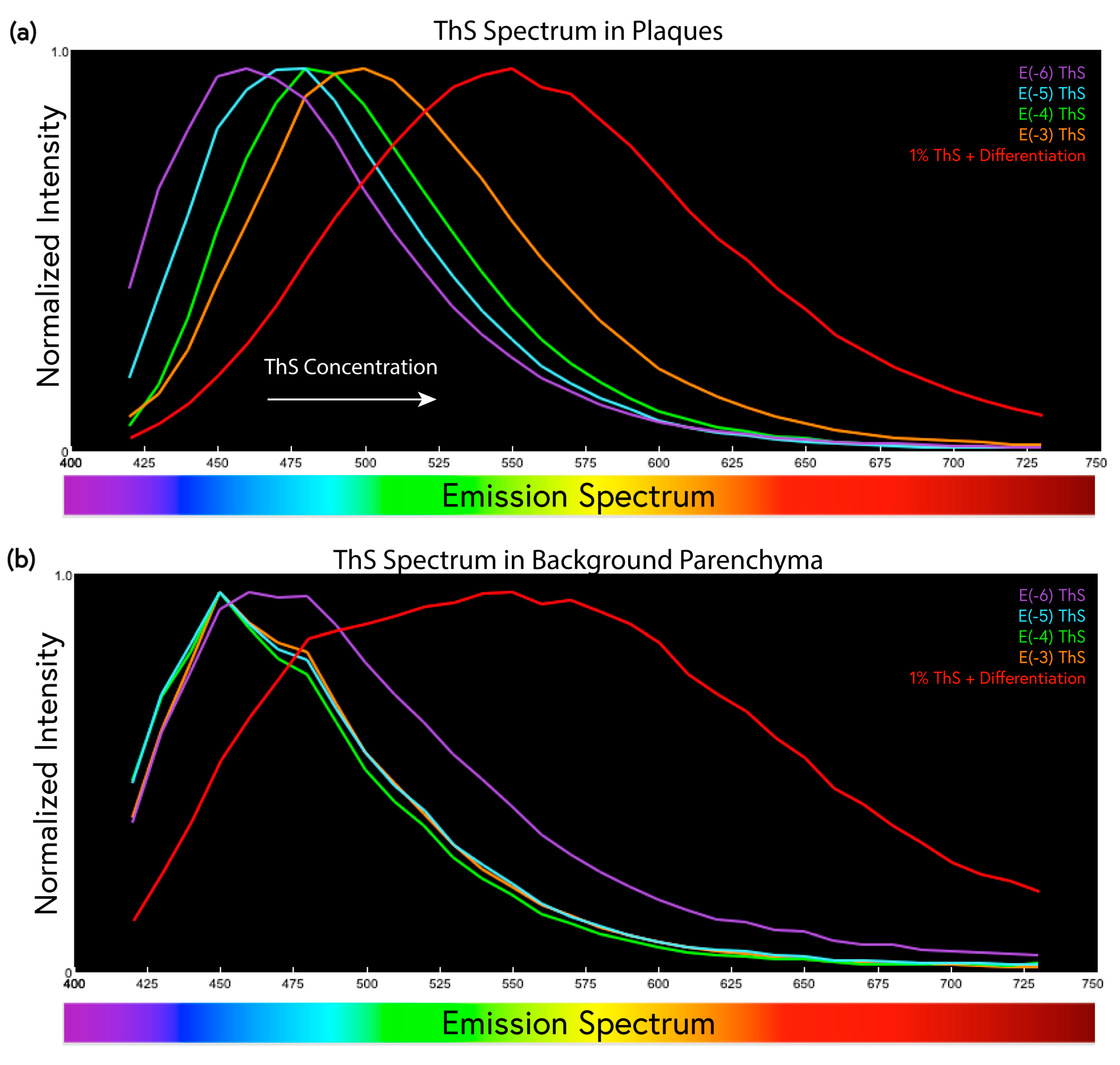
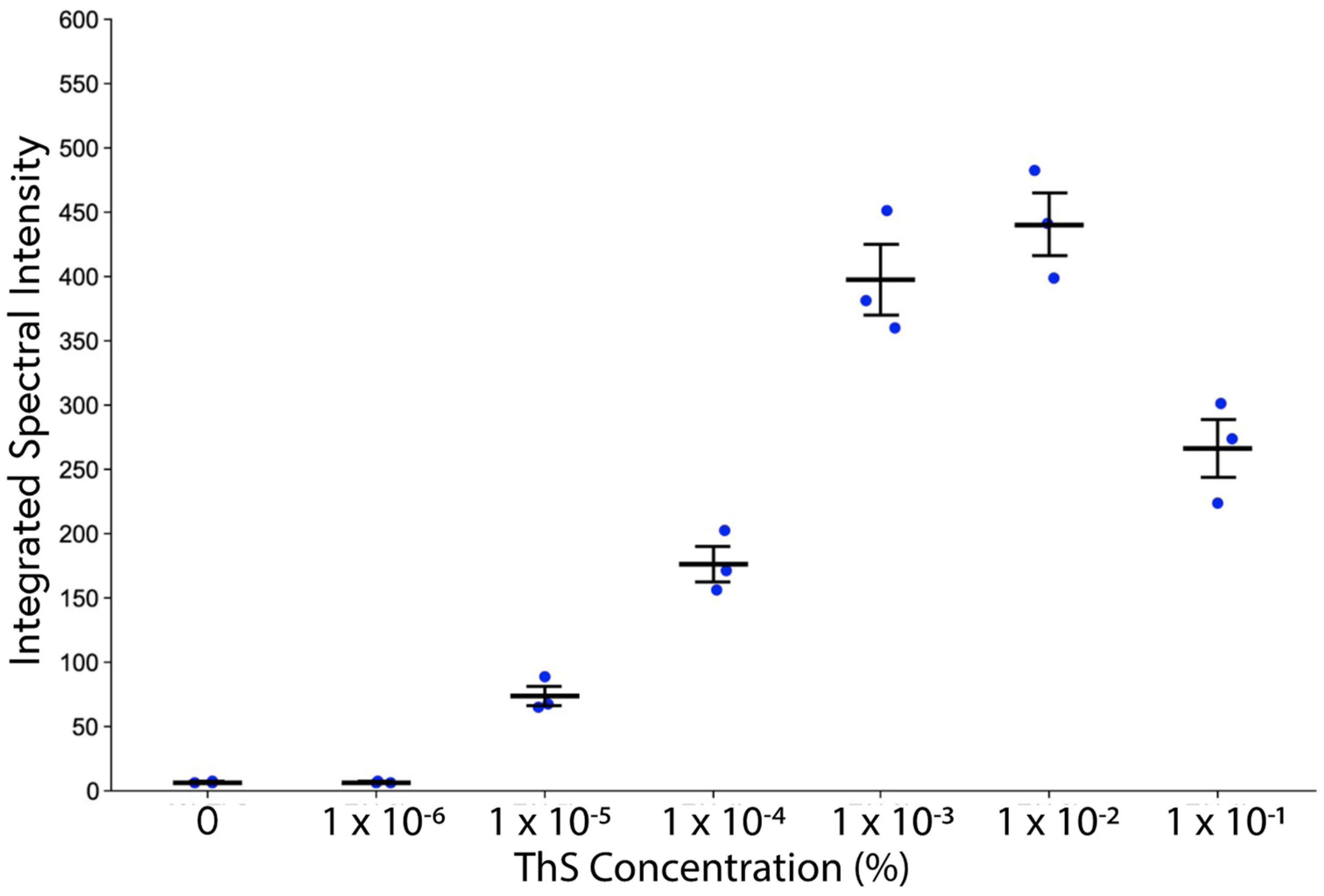
2.3. ThS Fluorescence Intensity as a Function of Concentration
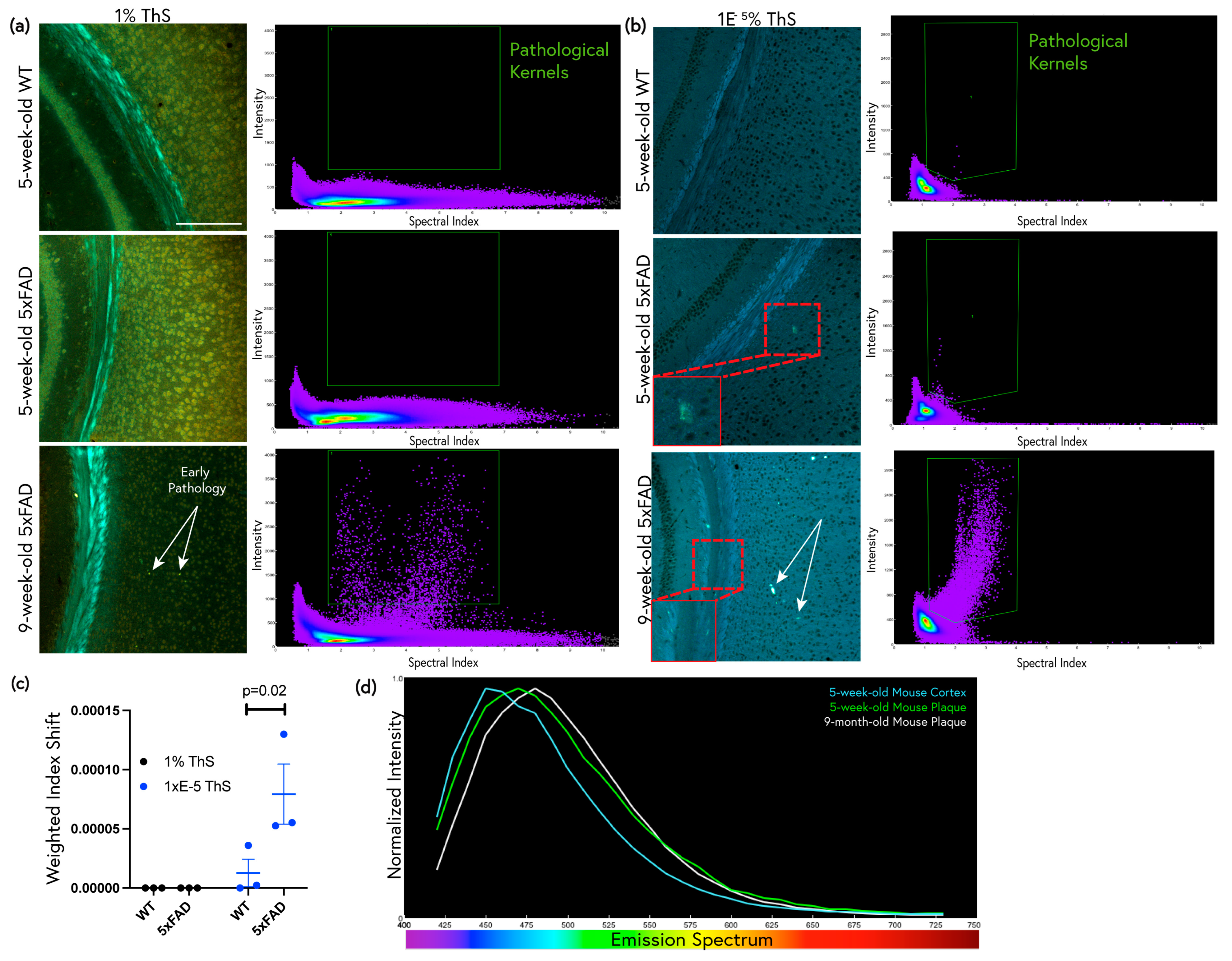
2.4. Low-Concentration ThS Staining Detects Early Amyloid Pathology in Young 5xFAD Mice
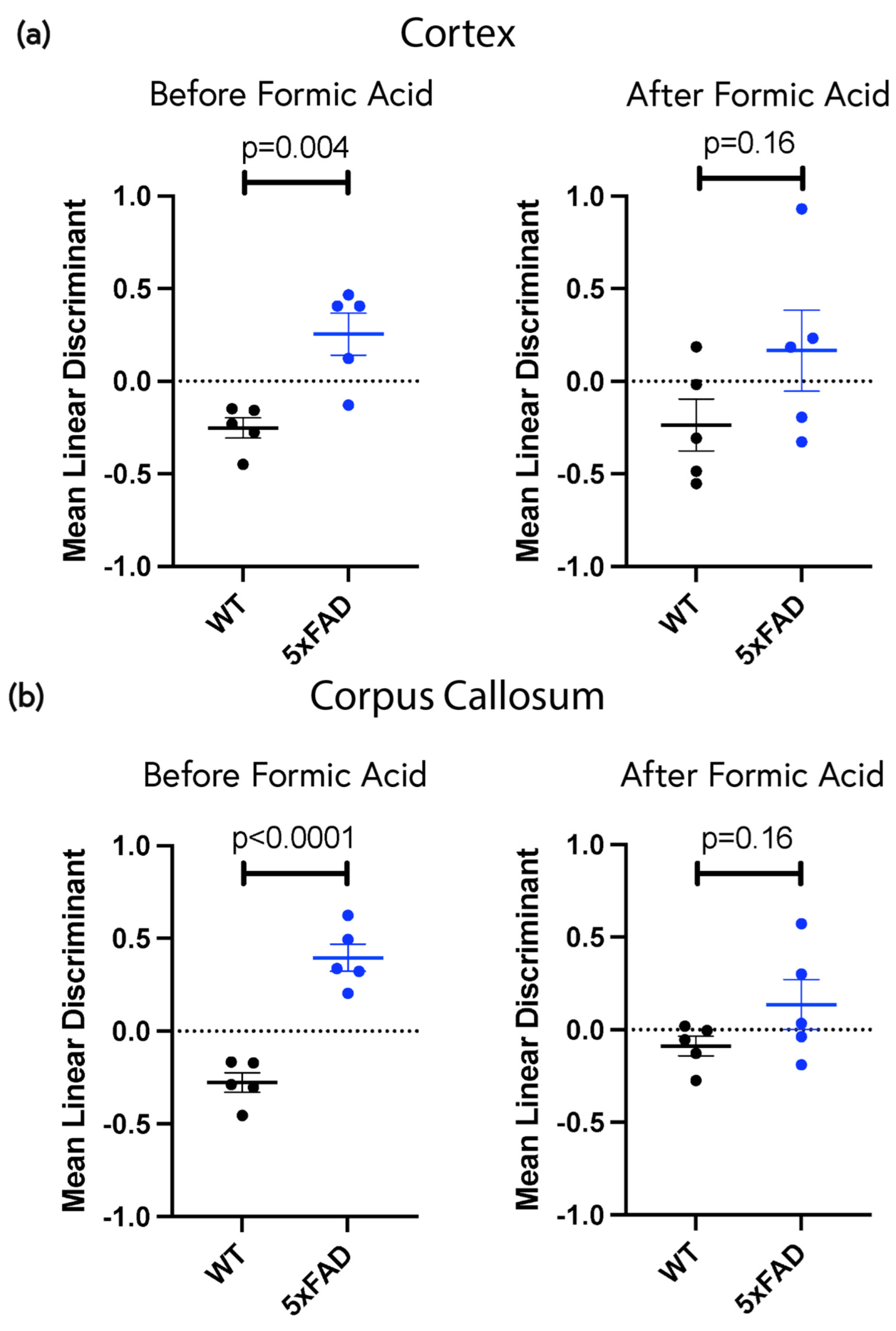
2.5. Low-Concentration ThS Staining Revealed Widespread Subtle Amyloid Deposition in the Non-Plaque Parenchyma
3. Discussion
4. Materials and Methods
4.1. Animals and Animal Care
4.2. Mouse Tissue Processing
4.3. Staining Procedure
4.4. Imaging and Analysis
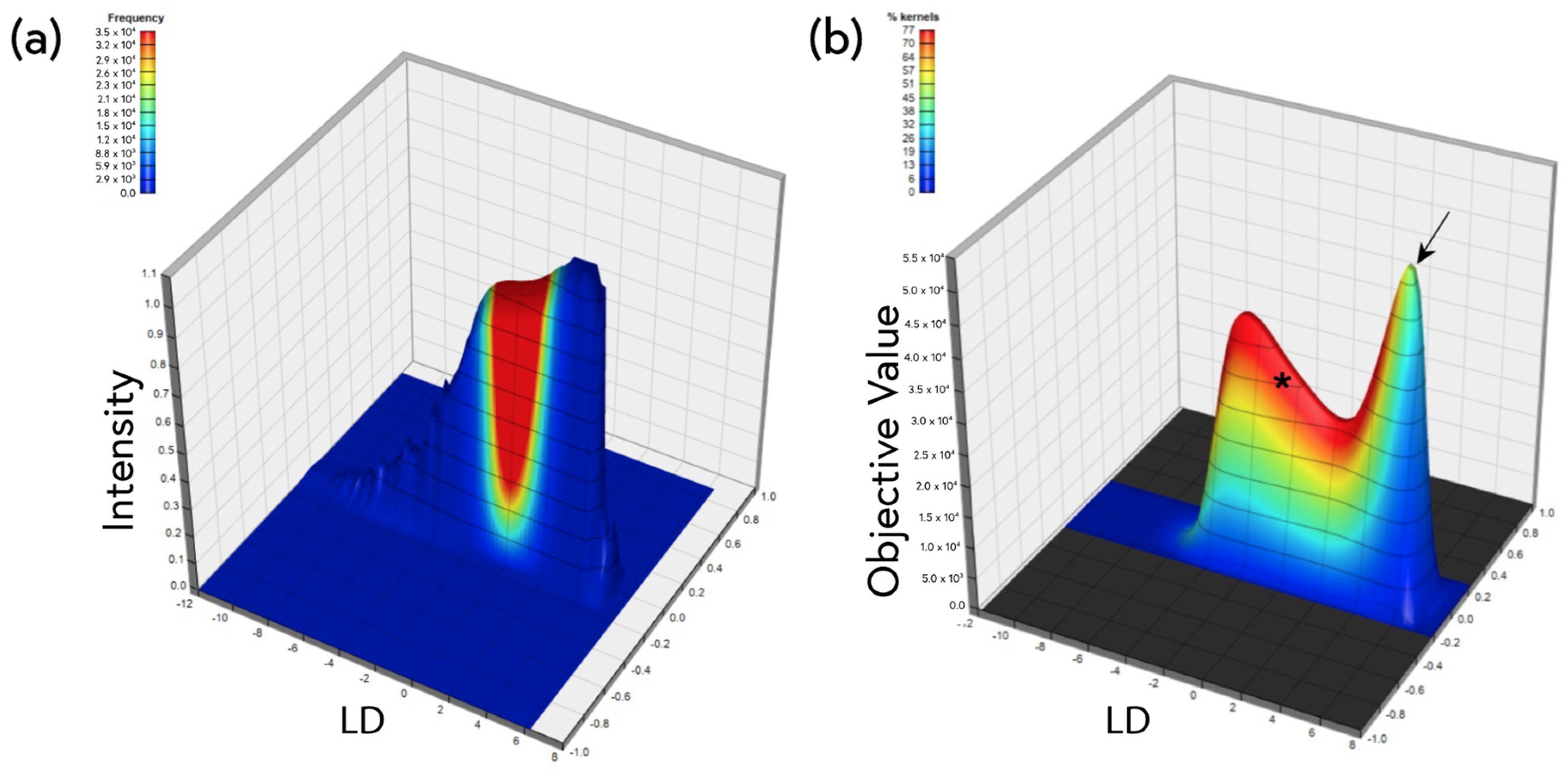
4.5. Machine Learning
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Naeem, A.; Fazili, N.A. Defective Protein Folding and Aggregation as the Basis of Neurodegenerative Diseases: The Darker Aspect of Proteins. Cell Biochem. Biophys. 2011, 61, 237–250. [Google Scholar] [CrossRef]
- Chuang, E.; Hori, A.M.; Hesketh, C.D.; Shorter, J. Amyloid Assembly and Disassembly. J. Cell Sci. 2018, 131, jcs189928. [Google Scholar] [CrossRef] [PubMed]
- Willbold, D.; Strodel, B.; Schröder, G.F.; Hoyer, W.; Heise, H. Amyloid-Type Protein Aggregation and Prion-like Properties of Amyloids. Chem. Rev. 2021, 121, 8285–8307. [Google Scholar] [CrossRef] [PubMed]
- Nelson, R.; Eisenberg, D. Recent Atomic Models of Amyloid Fibril Structure. Curr. Opin. Struct. Biol. 2006, 16, 260–265. [Google Scholar] [CrossRef]
- Chatani, E.; Yuzu, K.; Ohhashi, Y.; Goto, Y. Current Understanding of the Structure, Stability and Dynamic Properties of Amyloid Fibrils. Int. J. Mol. Sci. 2021, 22, 4349. [Google Scholar] [CrossRef] [PubMed]
- Dugger, B.N.; Dickson, D.W. Pathology of Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2017, 9, a028035. [Google Scholar] [CrossRef]
- Walker, L.C.; LeVine, H.; Mattson, M.P.; Jucker, M. Inducible Proteopathies. Trends Neurosci. 2006, 29, 438–443. [Google Scholar] [CrossRef]
- Sperling, R.A.; LaViolette, P.S.; O’Keefe, K.; O’Brien, J.; Rentz, D.M.; Pihlajamaki, M.; Marshall, G.; Hyman, B.T.; Selkoe, D.J.; Hedden, T.; et al. Amyloid Deposition Is Associated with Impaired Default Network Function in Older Persons without Dementia. Neuron 2009, 63, 178–188. [Google Scholar] [CrossRef]
- Strømland, Ø.; Jakubec, M.; Furse, S.; Halskau, Ø. Detection of Misfolded Protein Aggregates from a Clinical Perspective. J. Clin. Transl. Res. 2016, 2, 11–26. [Google Scholar]
- Sulatskaya, A.I.; Rodina, N.P.; Sulatsky, M.I.; Povarova, O.I.; Antifeeva, I.A.; Kuznetsova, I.M.; Turoverov, K.K. Investigation of α-Synuclein Amyloid Fibrils Using the Fluorescent Probe Thioflavin T. Int. J. Mol. Sci. 2018, 19, 2486. [Google Scholar] [CrossRef]
- Santa-María, I.; Pérez, M.; Hernández, F.; Avila, J.; Moreno, F.J. Characteristics of the Binding of Thioflavin S to Tau Paired Helical Filaments. J. Alzheimer’s Dis. 2006, 9, 279–285. [Google Scholar] [CrossRef]
- Stiller, D.; Katenkamp, D.; Thoss, K. [Fluorescence Histochemistry Demonstration of Amyloid with Thioflavin S and Acridine Orange]. Acta Histochem. 1970, 38, 18–30. [Google Scholar] [PubMed]
- Kelényi, G. Thioflavin S Fluorescent and Congo Red Anisotropic Stainings in the Histologic Demonstration of Amyloid. Acta Neuropathol. 1967, 7, 336–348. [Google Scholar] [CrossRef]
- Åslund, A.; Sigurdson, C.J.; Klingstedt, T.; Grathwohl, S.; Bolmont, T.; Dickstein, D.L.; Glimsdal, E.; Prokop, S.; Lindgren, M.; Konradsson, P.; et al. Novel Pentameric Thiophene Derivatives for in Vitro and in Vivo Optical Imaging of a Plethora of Protein Aggregates in Cerebral Amyloidoses. ACS Chem. Biol. 2009, 4, 673–684. [Google Scholar] [CrossRef]
- Biancalana, M.; Koide, S. Molecular Mechanism of Thioflavin-T Binding to Amyloid Fibrils. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2010, 1804, 1405–1412. [Google Scholar] [CrossRef]
- Mukherjee, A.; Al-Lahham, R.; Corkins, M.E.; Samanta, S.; Schmeichel, A.M.; Singer, W.; Low, P.A.; Govindaraju, T.; Soto, C. Identification of Multicolor Fluorescent Probes for Heterogeneous Aβ Deposits in Alzheimer’s Disease. Front. Aging Neurosci. 2022, 13, 802614. [Google Scholar] [CrossRef] [PubMed]
- Groenning, M. Binding Mode of Thioflavin T and Other Molecular Probes in the Context of Amyloid Fibrils—Current Status. J. Chem. Biol. 2010, 3, 1–18. [Google Scholar] [CrossRef]
- Nilsson, K.P.R. Small Organic Probes as Amyloid Specific Ligands—Past and Recent Molecular Scaffolds. FEBS Lett. 2009, 583, 2593–2599. [Google Scholar] [CrossRef] [PubMed]
- Naiki, H.; Higuchi, K.; Hosokawa, M.; Takeda, T. Fluorometric Determination of Amyloid Fibrils in Vitro Using the Fluorescent Dye, Thioflavine T. Anal. Biochem. 1989, 177, 244–249. [Google Scholar] [CrossRef]
- Levine, H. Thioflavine T Interaction with Synthetic Alzheimer’s Disease Β-amyloid Peptides: Detection of Amyloid Aggregation in Solution. Protein Sci. 1993, 2, 404–410. [Google Scholar] [CrossRef]
- Espargaró, A.; Sabate, R.; Ventura, S. Thioflavin-S Staining Coupled to Flow Cytometry. A Screening Tool to Detect in Vivo Protein Aggregation. Mol. BioSyst. 2012, 8, 2839–2844. [Google Scholar] [CrossRef] [PubMed]
- Sun, A.; Nguyen, X.V.; Bing, G. Comparative Analysis of an Improved Thioflavin-S Stain, Gallyas Silver Stain, and Immunohistochemistry for Neurofibrillary Tangle Demonstration on the Same Sections. J. Histochem. Cytochem. 2002, 50, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Hefti, M.M.; Kim, S.H.; Bell, A.J.; Betters, R.K.; Fiock, K.L.; Iida, M.A.; Smalley, M.E.; Farrell, K.; Fowkes, M.E.; Crary, J.F. Tau Phosphorylation and Aggregation in the Developing Human Brain. J. Neuropathol. Exp. Neurol. 2019, 78, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Bussière, T.; Bard, F.; Barbour, R.; Grajeda, H.; Guido, T.; Khan, K.; Schenk, D.; Games, D.; Seubert, P.; Buttini, M. Morphological Characterization of Thioflavin-S-Positive Amyloid Plaques in Transgenic Alzheimer Mice and Effect of Passive Aβ Immunotherapy on Their Clearance. Am. J. Pathol. 2004, 165, 987–995. [Google Scholar] [CrossRef]
- Schmidt, M.L.; Robinson, K.A.; Lee, V.M.Y.; Trojanowski, J.Q. Chemical and Immunological Heterogeneity of Fibrillar Amyloid in Plaques of Alzheimer’s Disease and Down’s Syndrome Brains Revealed by Confocal Microscopy. Am. J. Pathol. 1995, 147, 503–515. [Google Scholar]
- Klymchenko, A.S. Solvatochromic and Fluorogenic Dyes as Environment-Sensitive Probes: Design and Biological Applications. Acc. Chem. Res. 2017, 50, 366–375. [Google Scholar] [CrossRef]
- Maskevich, A.A.; Stsiapura, V.I.; Kuzmitsky, V.A.; Kuznetsova, I.M.; Povarova, O.I.; Uversky, V.N.; Turoverov, K.K. Spectral Properties of Thioflavin T in Solvents with Different Dielectric Properties and in a Fibril-Incorporated Form. J. Proteome Res. 2007, 6, 1392–1401. [Google Scholar] [CrossRef]
- Stsiapura, V.I.; Maskevich, A.A.; Kuzmitsky, V.A.; Turoverov, K.K.; Kuznetsova, I.M. Computational Study of Thioflavin T Torsional Relaxation in the Excited State. J. Phys. Chem. A 2007, 111, 4829–4835. [Google Scholar] [CrossRef]
- Stsiapura, V.I.; Maskevich, A.A.; Kuzmitsky, V.A.; Uversky, V.N.; Kuznetsova, I.M.; Turoverov, K.K. Thioflavin T as a Molecular Rotor: Fluorescent Properties of Thioflavin T in Solvents with Different Viscosity. J. Phys. Chem. B 2008, 112, 15893–15902. [Google Scholar] [CrossRef]
- Amdursky, N.; Erez, Y.; Huppert, D. Molecular Rotors: What Lies behind the High Sensitivity of the Thioflavin-T Fluorescent Marker. Acc. Chem. Res. 2012, 45, 1548–1557. [Google Scholar] [CrossRef]
- Oakley, H.; Cole, S.L.; Logan, S.; Maus, E.; Shao, P.; Craft, J.; Guillozet-Bongaarts, A.; Ohno, M.; Disterhoft, J.; Eldik, L.V.; et al. Intraneuronal β-Amyloid Aggregates, Neurodegeneration, and Neuron Loss in Transgenic Mice with Five Familial Alzheimer’s Disease Mutations: Potential Factors in Amyloid Plaque Formation. J. Neurosci. 2006, 26, 10129–10140. [Google Scholar] [CrossRef]
- Forner, S.; Kawauchi, S.; Balderrama-Gutierrez, G.; Kramár, E.A.; Matheos, D.P.; Phan, J.; Javonillo, D.I.; Tran, K.M.; Hingco, E.; da Cunha, C.; et al. Systematic Phenotyping and Characterization of the 5xFAD Mouse Model of Alzheimer’s Disease. Sci. Data 2021, 8, 270. [Google Scholar] [CrossRef]
- Tikhonova, T.N.; Rovnyagina, N.R.; Zherebker, A.Y.; Sluchanko, N.N.; Rubekina, A.A.; Orekhov, A.S.; Nikolaev, E.N.; Fadeev, V.V.; Uversky, V.N.; Shirshin, E.A. Dissection of the Deep-Blue Autofluorescence Changes Accompanying Amyloid Fibrillation. Arch. Biochem. Biophys. 2018, 651, 13–20. [Google Scholar] [CrossRef]
- Yoshiyama, Y.; Higuchi, M.; Zhang, B.; Huang, S.M.; Iwata, N.; Saido, T.C.C.; Maeda, J.; Suhara, T.; Trojanowski, J.Q.; Lee, V.M.Y. Synapse Loss and Microglial Activation Precede Tangles in a P301S Tauopathy Mouse Model. Neuron 2007, 53, 337–351. [Google Scholar] [CrossRef]
- Conway, K.A.; Harper, J.D.; Lansbury, P.T. Fibrils Formed in Vitro from α-Synuclein and Two Mutant Forms Linked to Parkinson’s Disease Are Typical Amyloid. Biochemistry 2000, 39, 2552–2563. [Google Scholar] [CrossRef]
- Nilsson, M.R. Insulin Amyloid at Injection Sites of Patients with Diabetes. Amyloid 2016, 23, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Barton, S.M.; To, E.; Rogers, B.P.; Whitmore, C.; Uppal, M.; Matsubara, J.A.; Pham, W. Inhalable Thioflavin s for the Detection of Amyloid Beta Deposits in the Retina. Molecules 2021, 26, 835. [Google Scholar] [CrossRef] [PubMed]
- Vadukul, D.M.; Al-Hilaly, Y.K.; Serpell, L.C. Methods for Structural Analysis of Amyloid Fibrils in Misfolding Diseases. In Protein Misfolding Diseases: Methods and Protocols; Springer: Humana, NY, USA, 2019; Volume 1873. [Google Scholar]
- Lindberg, D.J.; Wenger, A.; Sundin, E.; Wesén, E.; Westerlund, F.; Esbjörner, E.K. Binding of Thioflavin-T to Amyloid Fibrils Leads to Fluorescence Self-Quenching and Fibril Compaction. Biochemistry 2017, 56, 2170–2174. [Google Scholar] [CrossRef]
- Sachdev, P.S.; Zhuang, L.; Braidy, N.; Wen, W. Is Alzheimer’s a Disease of the White Matter? Curr. Opin. Psychiatry 2013, 26, 244–251. [Google Scholar] [CrossRef]
- Roher, A.E.; Palmer, K.C.; Yurewicz, E.C.; Ball, M.J.; Greenberg, B.D. Morphological and Biochemical Analyses of Amyloid Plaque Core Proteins Purified from Alzheimer Disease Brain Tissue. J. Neurochem. 1993, 61, 1916–1926. [Google Scholar] [CrossRef] [PubMed]
- McLean, C.A.; Cherny, R.A.; Fraser, F.W.; Fuller, S.J.; Smith, M.J.; Vbeyreuther, K.; Bush, A.I.; Masters, C.L. Soluble Pool of Aβ Amyloid as a Determinant of Severity of Neurodegeneration in Alzheimer’s Disease. Ann. Neurol. 1999, 46, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Fiock, K.L.; Betters, R.K.; Hefti, M.M. Thioflavin S Staining and Amyloid Formation Are Unique to Mixed Tauopathies. J. Histochem. Cytochem. 2023, 71, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A. The Use of Multiple Measurements in Taxonomic Problems. Ann. Eugen. 1936, 7, 179–188. [Google Scholar] [CrossRef]
- Xanthopoulos, P.; Pardalos, P.M.; Trafais, T.B. Linear Discriminant Analysis. In Robust Data Mining; Springer: New York, NY, USA, 2013. [Google Scholar]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-Learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MacKeigan, T.P.; Morgan, M.L.; Stys, P.K. Quantitation of Tissue Amyloid via Fluorescence Spectroscopy Using Controlled Concentrations of Thioflavin-S. Molecules 2023, 28, 4483. https://doi.org/10.3390/molecules28114483
MacKeigan TP, Morgan ML, Stys PK. Quantitation of Tissue Amyloid via Fluorescence Spectroscopy Using Controlled Concentrations of Thioflavin-S. Molecules. 2023; 28(11):4483. https://doi.org/10.3390/molecules28114483
Chicago/Turabian StyleMacKeigan, Tatiana P., Megan L. Morgan, and Peter K. Stys. 2023. "Quantitation of Tissue Amyloid via Fluorescence Spectroscopy Using Controlled Concentrations of Thioflavin-S" Molecules 28, no. 11: 4483. https://doi.org/10.3390/molecules28114483