
Metabolomic Profiling, Antioxidant and Enzyme Inhibition Properties and Molecular Docking Analysis of Antarctic Lichens

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Metabolomics Profiles and Chemical Fingerprints of Lichen Extracts
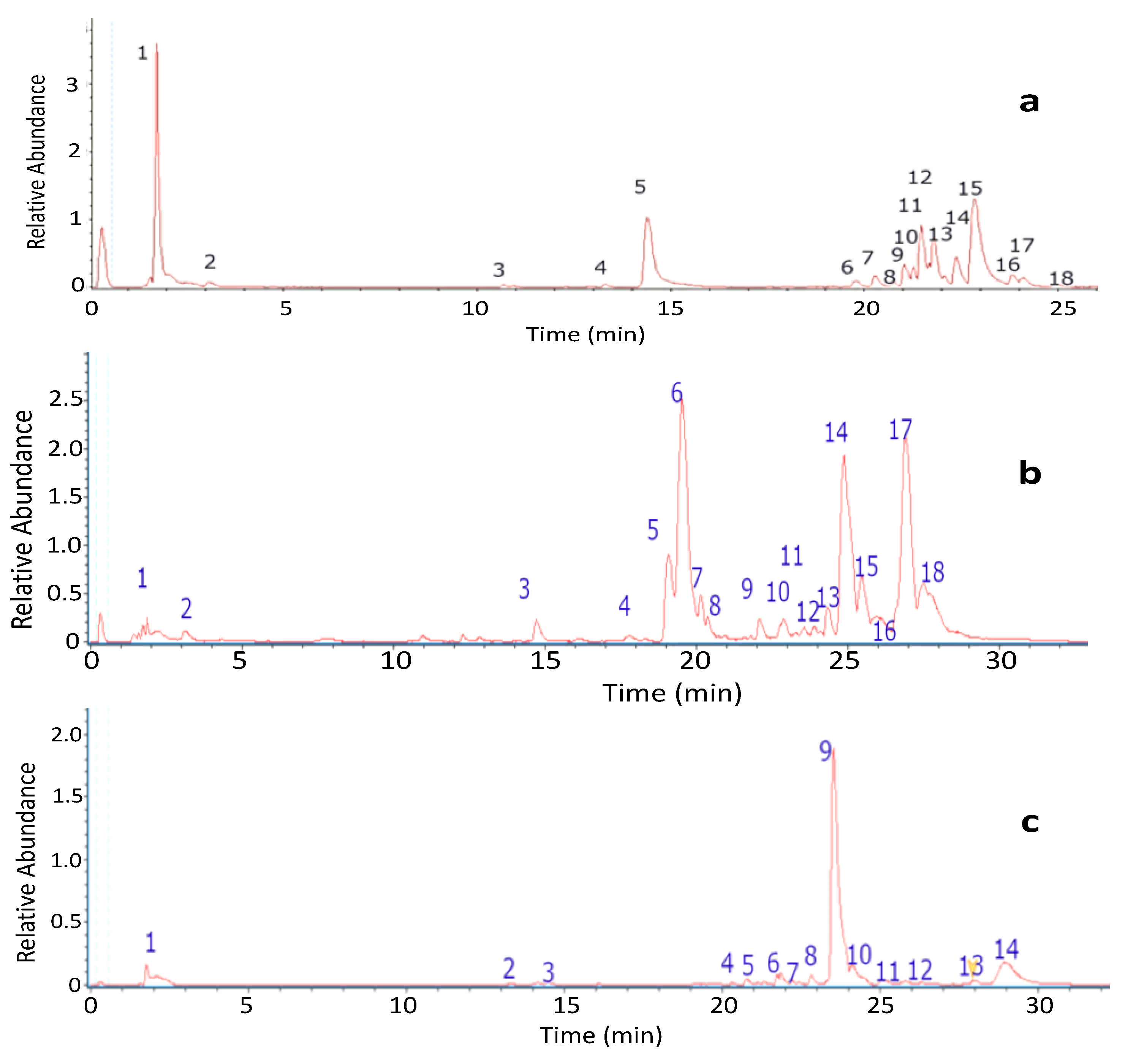
2.1.1. UHPLC Chromatographic Analysis of Lecania brialmontii
Carbohydrates
Fatty Acids
Anthraquinones
Aromatic Derivatives
Lipids
Depsides
2.1.2. UHPLC Chromatographic Analysis of Pseudephebe pubescens
Carbohydrates
Fatty Acids
Lipids
Depsides
Aromatic Derivatives
Dibenzofurans
2.1.3. UHPLC Chromatographic Analysis of Sphaerophorus globosus
Carbohydrates
Aromatic Derivatives
Fatty Acids
Depsides
Dibenzofurans
2.2. Total Phenolic Contents and Antioxidant Activity
2.3. Enzymatic Inhibitory Activity
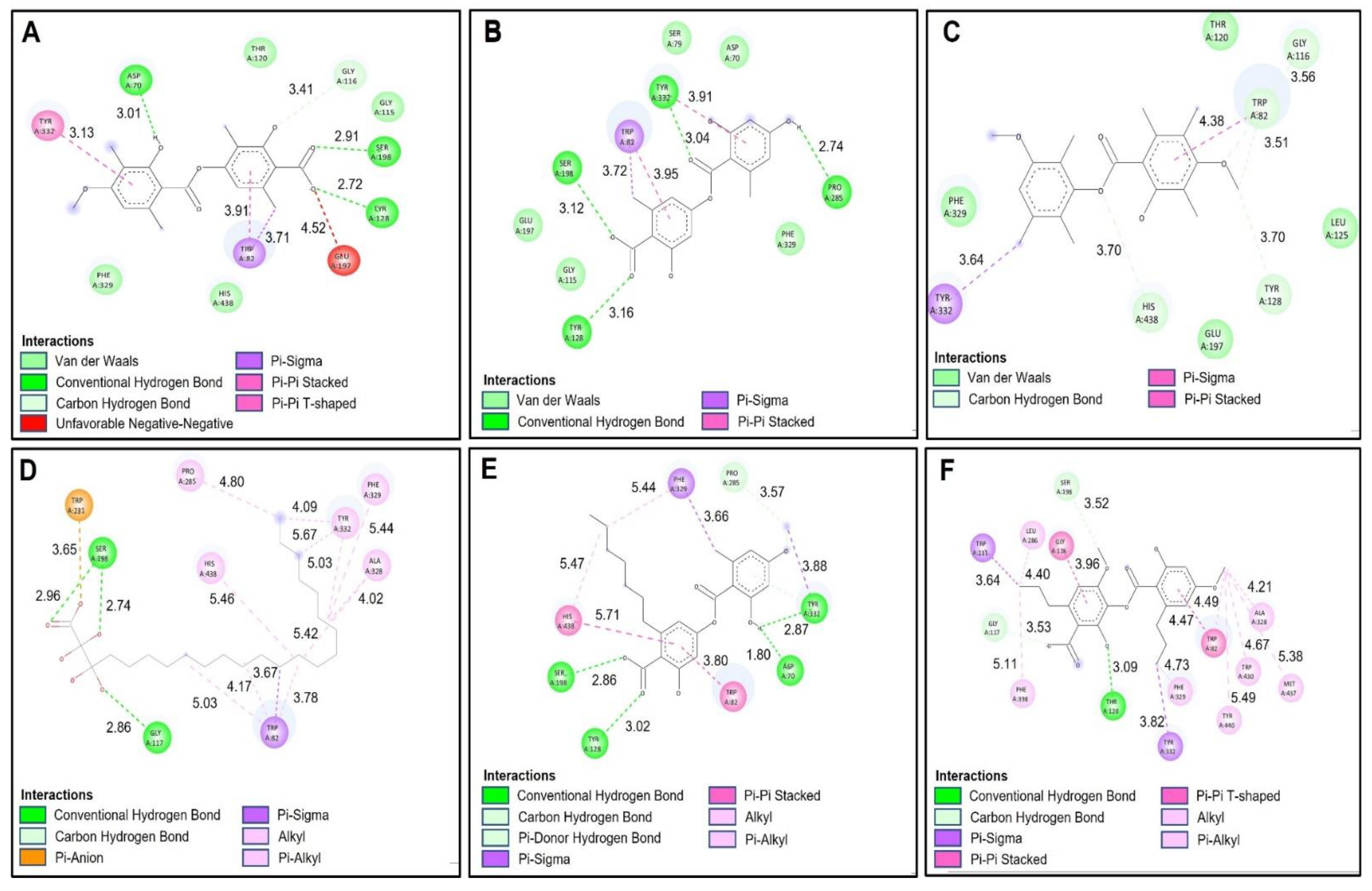
2.4. Docking Studies
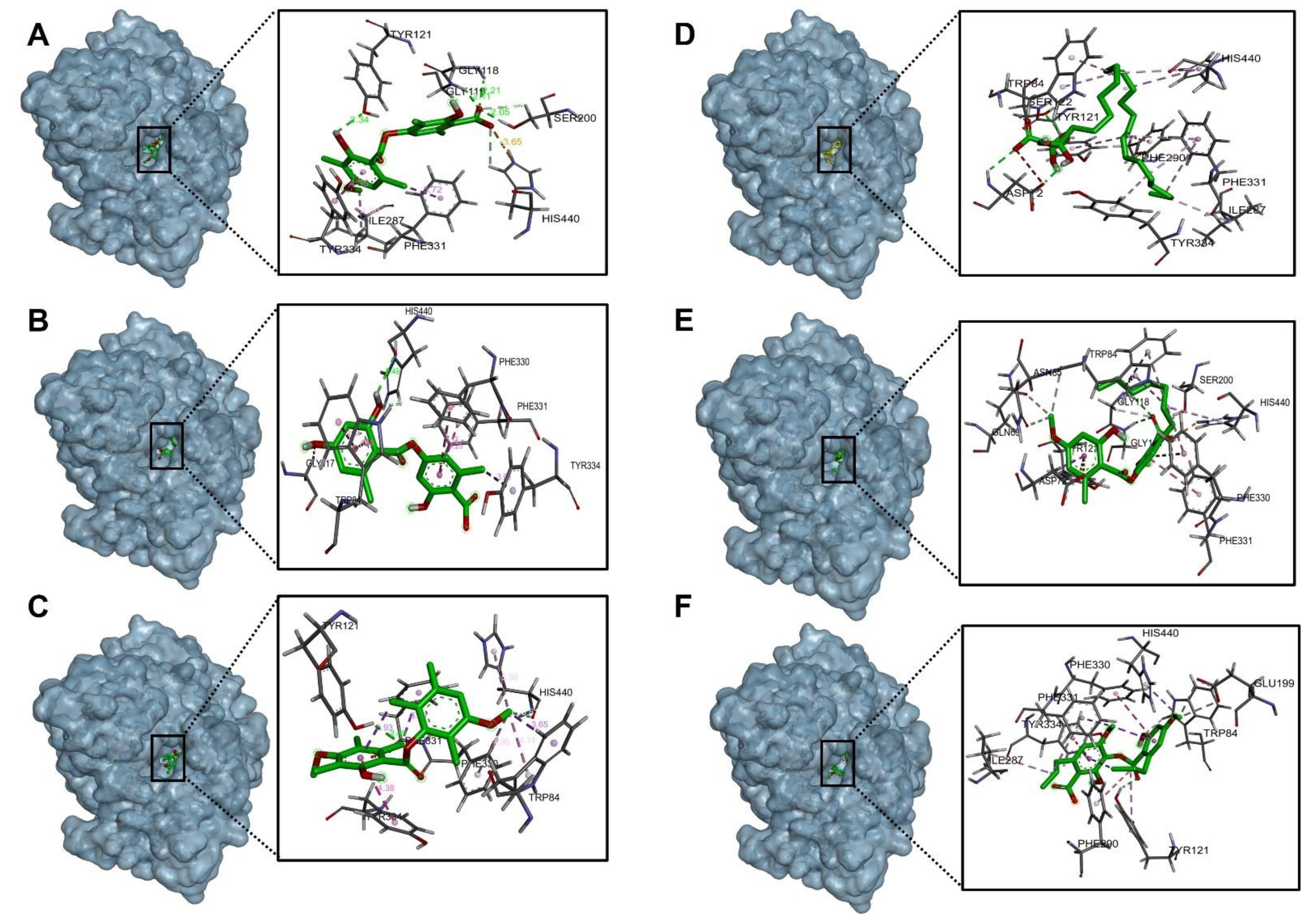
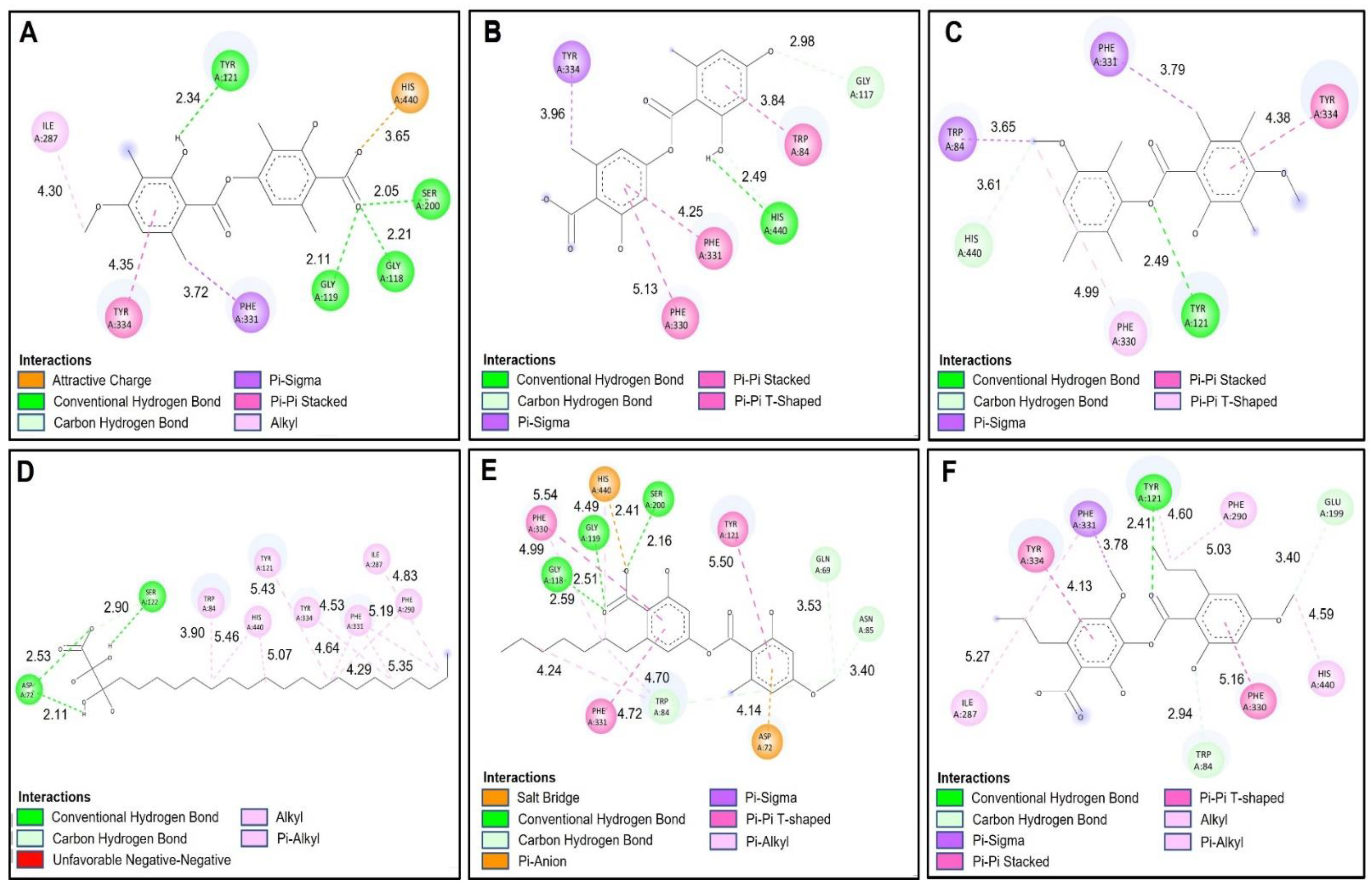
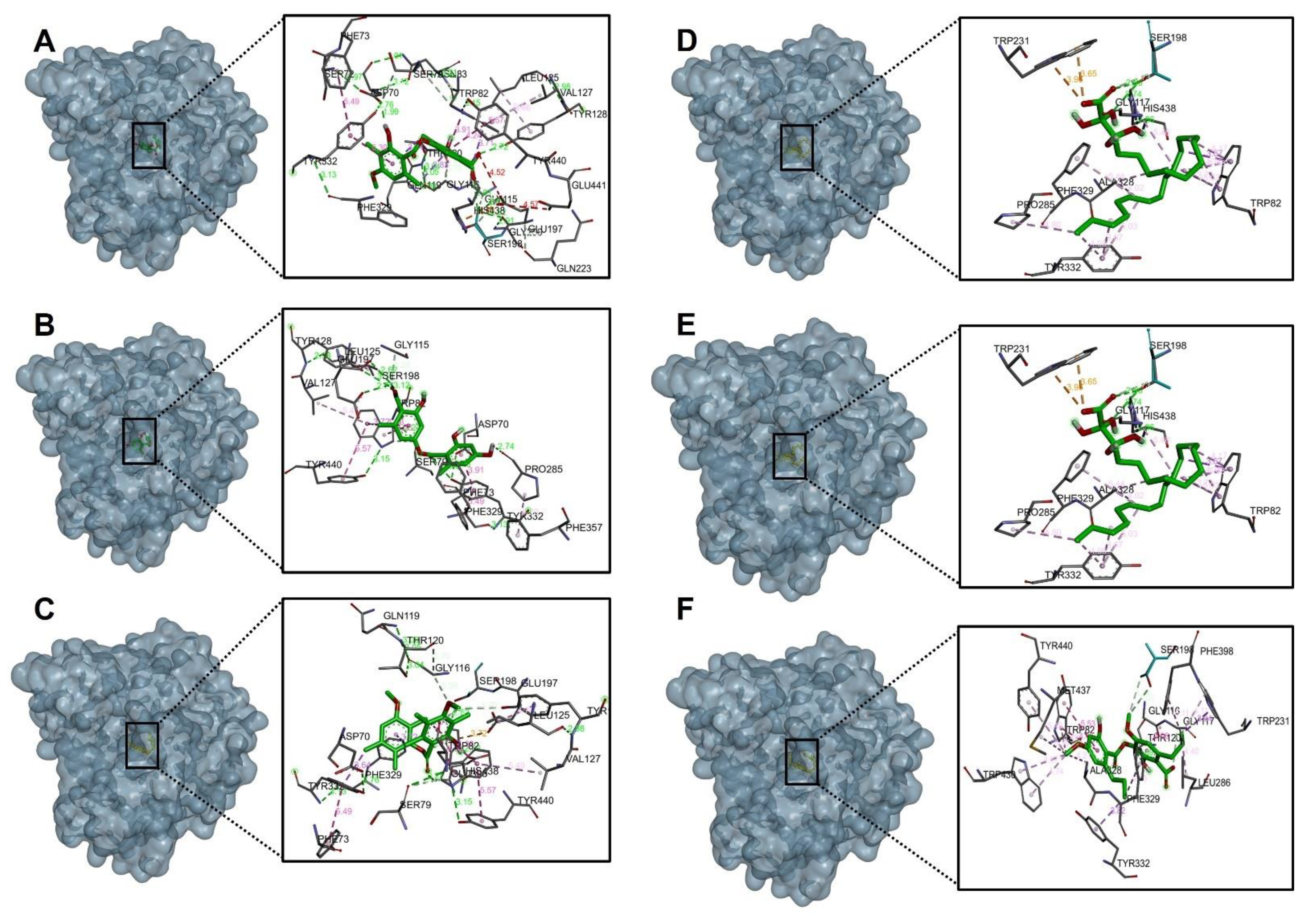
2.4.1. Acetylcholinesterase (TcAChE) Docking Results
2.4.2. Butyrylcholinesterase (hBChE) Docking Results
2.5. Prediction of Pharmacokinetic Properties—ADME
2.6. Toxicity Prediction
3. Materials and Methods
3.1. Chemicals
3.2. Lichen Material
3.3. Preparation of the Ethanolic Extracts
3.4. LC Parameters and MS Parameters
3.5. Total Phenolic Content
3.6. Antioxidant Activity
3.6.1. Ferric-Reducing Antioxidant Power (FRAP) Assay
3.6.2. Oxygen Radical Absorbance Capacity (ORAC) Assay
3.7. Determination of Cholinesterase Inhibition
3.8. Docking Simulations
3.9. Calculation of ADME Parameters
3.10. Calculation of Risk Toxicity
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Spribille, T.; Tuovinen, V.; Resl, P.; Vanderpool, D.; Wolinski, H.; Aime, M.C.; Schneider, K.; Stabentheiner, E.; Toome-Heller, M.; Thor, G.; et al. Basidiomycete yeasts in the cortex of ascomycete macrolichens. Science 2016, 353, 488–492. [Google Scholar] [CrossRef] [Green Version]
- Hawksworth, D.; Grube, M. Lichens redefined as complex ecosystems. New Phytol. 2020, 227, 1281–1283. [Google Scholar] [CrossRef]
- Nash, T. Lichen Biology, 2nd ed.; Cambridge University Publishing House: Cambridge, UK, 2008; 486p. [Google Scholar]
- Chaparro, M.; Aguirre, J. Lichenized Fungi; National University of Colombia: Bogotá, Colombia, 2002; 220p. [Google Scholar]
- Jaramillo, M.M.; Botero, L.R. Lichenic communities as bioindicators of air quality in the Aburra Valley. Gestión Ambiente 2010, 13, 97–110. [Google Scholar]
- Redón, J. Antarctic Lichens; Instituto Antártico Chileno: Santiago de Chile, Chile, 1985; 123p. [Google Scholar]
- Ovstedal, D.O.; Lewis, R.I. Four additional lichens from the Antarctic and South Georgia, including a new Leciophysma species. Folia Cryptogam. Est. 2011, 48, 65–68. [Google Scholar]
- Index Fungorum. Available online: http://www.indexfungorum.org/ (accessed on 26 October 2022).
- MycoBank Database. Available online: https://www.mycobank.org/ (accessed on 26 October 2022).
- Consortium of North American Lichen Herbaria. Available online: https://www.lichenportal.org/cnalh/ (accessed on 26 October 2022).
- Rankovic, B. Lichen Secondary Metabolites; Springer International Publishing: Cham, Switzerland, 2015; 202p. [Google Scholar]
- Calcott, M.J.; Ackerley, D.F.; Knight, A.; Keyzers, R.A.; Owen, J.G. Secondary metabolism in the lichen symbiosis. Chem. Soc. Rev. 2018, 47, 1730. [Google Scholar] [CrossRef]
- Olivier-Jimenez, D.; Chollet-Krugler, M.; Rondeau, D.; Beniddir, M.A.; Ferron, S.; Delhaye, T.; Allard, P.M.; Wolfender, J.L.; Sipman, H.J.M.; Lucking, R.; et al. A database of high-resolution MS/MS spectra for lichen metabolites. Sci. Data 2019, 6, 294. [Google Scholar] [CrossRef] [Green Version]
- Molnar, K.; Farkas, E. Current results on biological activities of lichen secondary metabolites: A review. Z. Naturforsch. 2010, 65, 157–173. [Google Scholar] [CrossRef]
- Mitrović, T.; Stamenković, S.; Cvetković, V.; Tošić, S.; Stanković, M.; Radojević, I.; Stefanović, O.; Čomić, L.; Đačić, D.; Ćurčić, M.; et al. Antioxidant, antimicrobial and antiproliferative activities of five lichens species. Int. J. Mol. Sci. 2011, 12, 5428–5448. [Google Scholar]
- Cardile, V.; Graziano, A.; Avola, R.; Piovano, M.; Russo, A. Potencial anticâncer activity of lichen secondary metabolite physodic acid. Chem.-Biol. Interact. 2017, 263, 36–45. [Google Scholar]
- White, P.A.S.; Oliveira, R.C.M.; Oliveira, A.P.; Serafini, M.R.; Araújo, A.A.S.; Gelain, D.P.; Moreira, J.C.F.; Almeida, J.R.G.S.; Quintans, J.S.S.; Quintans-Junior, L.J.; et al. Antioxidant Activity and Mechanisms of Action of Natural Compounds Isolated from Lichens: A Systematic Review. Molecules 2014, 19, 14496–14527. [Google Scholar] [CrossRef]
- Elečko, J.; Vilková, M.; Frenák, R.; Routray, D.; Ručová, D.; Bačkor, M.; Goga, M. A Comparative Study of Isolated Secondary Metabolites from Lichens and Their Antioxidative Properties. Plants 2022, 11, 1077. [Google Scholar] [CrossRef]
- Hawrył, A.; Hawrył, M.; Hajnos-Stolarz, A.; Abramek, J.; Bogucka-Kocka, A.; Komsta, Ł. HPLC Fingerprint Analysis with the Antioxidant and Cytotoxic Activities of Selected Lichens Combined with the Chemometric Calculations. Molecules 2020, 25, 4301. [Google Scholar] [CrossRef]
- Fernández-Moriano, C.; Gómez-Serranillos, M.P.; Crespo, A. Antioxidant potential of lichen species and their secondary metabolites. A systematic review. Pharm. Biol. 2016, 54, 1–17. [Google Scholar]
- Plaza, C.; Díaz de Torres, L.; Lücking, R.; Vizcaya, M.; Medina, G. Antioxidant activity, total phenols and flavonoids of lichens from Venezuelan Andes. J. Pharm. Pharmacogn. Res. 2014, 2, 138–147. [Google Scholar]
- Tas, I.; Yildirim, A.B.; Ozkan, E.; Ozyigitoglu, G.C.; Yavuz, M.Z.; Turker, A.U. Biological evaluation and phytochemical profiling of some lichens species. Acta Aliment. 2019, 48, 457–465. [Google Scholar] [CrossRef]
- Ureña-Vacas, I.; González-Burgos, E.; De Vita, S.; Divakar, P.K.; Bifulco, G.; Gómez-Serranillos, M.P. Phytochemical Characte rization and Pharmacological Properties of Lichen Extracts from Cetrarioid Clade by Multivariate Analysis and Molecular Docking. Evid.-Based Complement. Altern. 2022, 2022, 5218248. [Google Scholar]
- Torres-Benítez, A.; Rivera-Montalvo, M.; Sepúlveda, B.; Castro, O.N.; Nagles, E.; Simirgiotis, M.J.; Garciá-Beltrán, O.; Areche, C. Metabolomic analysis of two Parmotrema lichens: P. robustum (Degel.) Hale and P. andinum (Mull. rg.) hale using UHPLC-ESI-OT-MS-MS. Molecules 2017, 22, 1861. [Google Scholar]
- Salgado, F.; Albornoz, L.; Cortéz, C.; Stashenko, E.; Urrea-Vallejo, K.; Nagles, E.; Galicia-Virviescas, C.; Cornejo, A.; Ardiles, A.; Simirgiotis, M.; et al. Secondary metabolite profiling of species of the genus Usnea by UHPLC-ESI-OT-MS-MS. Molecules 2018, 23, 54. [Google Scholar] [CrossRef]
- Albornoz, L.; Torres-Benítez, A.; Moreno-Palacios, M.; Simirgiotis, M.J.; Montoya-Serrano, S.A.; Sepulveda, B.; Stashenko, E.; García-Beltrán, O.; Areche, C. Phylogenetic Studies and Metabolite Analysis of Sticta Species from Colombia and Chile by Ultra-High Performance Liquid Chromatography-High Resolution-Q-Orbitrap-Mass Spectrometry. Metabolites 2022, 12, 156. [Google Scholar] [CrossRef]
- Sepúlveda, B.; Cornejo, A.; Bárcenas-Pérez, D.; Cheel, J.; Areche, C. Two New Fumarprotocetraric Acid Lactones Identified and Characterized by UHPLC-PDA/ESI/ORBITRAP/MS/MS from the Antarctic Lichen Cladonia metacorallifera. Separations 2022, 9, 41. [Google Scholar] [CrossRef]
- Areche, C.; Parra, J.R.; Sepulveda, B.; García-Beltrán, O.; Simirgiotis, M.J. UHPLC-MS Metabolomic Fingerprinting, Antioxidant, and Enzyme Inhibition Activities of Himantormia lugubris from Antarctica. Metabolites 2022, 12, 560. [Google Scholar] [CrossRef]
- Guedes, I.A.; Pereira, F.; Dardenne, L. Empirical scoring functions for structure-based virtual screening: Applications, critical aspects, and challenges. Front. Pharmacol. 2018, 9, 1089. [Google Scholar] [CrossRef] [PubMed]
- Cerón-Carrasco, J. When virtual screening yields inactive drugs: Dealing with false theoretical friends. ChemMedChem 2022, 17, e202200278. [Google Scholar] [CrossRef] [PubMed]
- Zev, S.; Raz, K.; Schwartz, R.; Tarabeh, R.; Kumar, P.; Major, D. Benchmarking the Ability of Common Docking Programs to Correctly Reproduce and Score Binding Modes in SARS-CoV-2 Protease Mpro. J. Chem. Inf. Model. 2021, 61, 2957–2966. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Suh, Y.J.; Yang, S.; Hong, D.G.; Ishigami, A.; Kim, H.; Hur, J.-S.; Chang, S.-C.; Lee, J. Neuroprotective and Anti-Inflammatory Effects of Evernic Acid in an MPTP-Induced Parkinson’s Disease Model. Int. J. Mol. Sci. 2021, 22, 2098. [Google Scholar] [CrossRef] [PubMed]
- Vinet, C.; Quilhot, W. Studies on chilean lichens, XIII. Polysubstituted depsides from Lecania brialmontii. J. Nat. Prod. 1990, 53, 500–502. [Google Scholar]
- Reese, R.; Ekman, S.; Tibell, L. Molecular phylogeny of the genus Lecania (Ramalinaceae, lichenized Ascomycota). Mycol. Res. 2007, 111, 581–591. [Google Scholar] [CrossRef]
- Da Silva, M.; Da Silva, A.; Fernandez, P.; Montone, R.; Alves, R.; De Queiroz, A.; De Oliveira, V.; Dos Santos, V.; Putzke, J.; Rosa, L.; et al. Extracellular hydrolytic enzymes produced by yeasts from Antarctic lichens. An. Acad. Bras. Cienc. 2022, 94, e20210540. [Google Scholar] [CrossRef]
- Reddy, S.D.; Siva, B.; Kumar, K.; Babu, V.S.P.; Sravanthi, V.; Boustie, J.; Nayak, V.L.; Tiwari, A.K.; Rao, C.V.; Sridhar, B.; et al. Comprehensive Analysis of Secondary Metabolites in Usnea longissima (Lichenized Ascomycetes, Parmeliaceae) Using UPLC-ESI-QTOF-MS/MS and Pro-Apoptotic Activity of Barbatic Acid. Molecules 2019, 24, 2270. [Google Scholar] [CrossRef]
- Garrido-Benavent, I.; Pérez-Ortega, S.; de Los Ríos, A.; Mayrhofer, H.; Fernández-Mendoza, F. Neogene speciation and Pleistocene expansion of the genus Pseudephebe (Parmeliaceae, lichenized fungi) involving multiple colonizations of Antarctica. Mol. Phylogenetic. Evol. 2021, 155, 107020. [Google Scholar] [CrossRef]
- Russo, A.; Piovano, M.; Lombardo, L.; Garbarino, J.; Cardile, V. Lichen metabolites prevent UV light and nitric oxide-mediated plasmid DNA damage and induce apoptosis in human melanoma cell. Life Sci. 2008, 83, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Ingólfsdóttir, K.; Kook Lee, S.; Bhat, K.P.; Lee, K.; Chai, H.B.; Kristinsson, H.; Song, L.L.; Gills, J.; Gudmundsdóttir, J.T.; Mata-Greenwood, E.; et al. Evaluation of selected lichens from iceland for cancer chemopreventive and cytotoxic activity. Phar. Biol. 2000, 38, 313–317. [Google Scholar] [CrossRef]
- Putzke, J.; Schaefer, C.E.; Villa, P.; Pereira, A.; Schunemann, A.; Putzke, M. The diversity and structure of plant communities in the maritime Antarctic is shaped by southern giant petrel’s (Macronectes giganteus) breeding activities. An. Acad. Bras. Cienc. 2022, 94, e20210597. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, P.; Ravindra, R. Screening of antioxidant potential of Artic lichen. Polar Biol. 2011, 34, 1775–1782. [Google Scholar] [CrossRef]
- Luo, H.; Yamamoto, Y.; Kim, J.; Jung, J.; Koh, Y.; Hur, J. Lecanoric acid, a secondary lichen substance with antioxidant properties from Umbilicaria antarctica in maritime Antarctica (King George Island). Polar Biol. 2009, 32, 1033–1040. [Google Scholar] [CrossRef]
- Ranković, B.; Kosanić, M.; Stanojković, T.; Vasiljević, P.; Manojlović, N. Biological activities of Tonidia candida and Usnea barbata together with their nostictic acid and usnic acid constituents. Int. J. Mol. Sci. 2012, 13, 14707–14722. [Google Scholar] [CrossRef] [Green Version]
- Aslan, A.; Güllüce, M.; Sökmen, M.; Adgüzel, A.; Sahin, F.; Özkan, H. Antioxidant and Antimicrobial Properties of the Lichens Cladonia foliacea, Dermatocarponminiatum., Everiniadivaricata., Everniaprunastri., and Neofuscella pulla. Pharm. Biol. 2006, 44, 247–252. [Google Scholar] [CrossRef]
- Nascimento, A.; Braz-Filho, R.; Mussi-Dias, V.; Curvino, I. Chemistry and biological activity of Ramalina lichenized fungi. Molecules 2015, 20, 8952–8987. [Google Scholar]
- Studzińska-Sroka, E.; Majchrzak-Celińska, A.; Zalewski, P.; Szwajgier, D.; Baranowska-Wójcik, E.; Kaproń, B.; Plech, T.; Żarowski, M.; Cielecka-Piontek, J. Lichen-Derived Compounds and Extracts as Biologically Active Substances with Anticancer and Neuroprotective Properties. Pharmaceuticals 2021, 14, 1293. [Google Scholar] [CrossRef]
- Jeong, G.S.; Hillman, P.F.; Kang, M.-G.; Hwang, S.; Park, J.-E.; Nam, S.-J.; Park, D.; Kim, H. Potent and Selective Inhibitors of Human Monoamine Oxidase A from an Endogenous Lichen Fungus Diaporthe mahothocarpus. J. Fungi 2021, 7, 876. [Google Scholar] [CrossRef]
- Pajouhesh, H.; Lenz, G. Medicinal chemistry properties of successful central nervous system drugs. NeuroRx 2005, 2, 541–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shultz, M. Two decades under the influence of the rule of five and the changing properties of approved oral drugs. J. Med. Chem. 2019, 62, 1701–1714. [Google Scholar] [CrossRef] [PubMed]
- Ley-Martínez, J.S.; Ortega-Valencia, J.E.; García-Barradas, O.; Jiménez-Fernández, M.; Uribe-Lam, E.; Vencedor-Meraz, C.I.; Oliva-Ramírez, J. Active Compounds in Zingiber officinale as Possible Redox Inhibitors of 5-Lipoxygenase Using an In Silico Approach. Int. J. Mol. Sci. 2022, 23, 6093. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin–Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990–5999. [Google Scholar] [CrossRef]
- Karadag, A.; Ozcelik, B.; Saner, S. Review of methods to determine antioxidant capacities. Food Anal. Methods 2009, 2, 41–60. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]
- Larrazábal-Fuentes, M.J.; Fernández-Galleguillos, C.; Palma-Ramírez, J.; Romero-Parra, J.; Sepúlveda, K.; Galetovic, A.; González, J.; Paredes, A.; Bórquez, J.; Simirgiotis, M.J.; et al. Chemical Profiling, Antioxidant, Anticholinesterase, and Antiprotozoal Potentials of Artemisia copa Phil. (Asteraceae). Front. Pharmacol. 2020, 11, 594174. [Google Scholar] [CrossRef]
- Greenblatt, H.M.; Kryger, G.; Lewis, T.; Silman, I.; Sussman, J.L. Structure of acetylcholinesterase complexed with (3)-galanthamine at 23 A resolution. FEBS Lett. 1999, 463, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Nachon, F.; Carletti, E.; Ronco, C.; Trovaslet, M.; Nicolet, Y.; Jean, L.; Renard, P. Crystal structure of human cholinesterases in complex with huprine W and tacrine: Elements of specificity for anti-Alzheimer’s drugs targeting acetyl- and butyryl-cholinesterase. Biochem. J. 2013, 453, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Ismaya, W.; Rozeboom, H.; Weijn, A.; Mes, J.; Fusetti, F.; Wichers, H.; Dijkstra, B. Crystal structure of Agaricus bisporus mushroom tyrosinase: Identity of the tetramer subunits and interaction with tropolone. Biochemistry 2011, 50, 5477–5486. [Google Scholar] [CrossRef]
- Berman, H.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.; Bourne, P. The Protein Data Bank. Nucl. Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, Q.; Triana, J.; Eiroa, J.; Calcul, L.; Rivera, E.; Wojtas, L.; Padrón, J.; Boberieth, L.; Keramane, M.; Abel-Santos, E.; et al. ent-Labdane diterpenoids from the aerial parts of Eupatorium obtusissmum. J. Nat. Prod. 2016, 79, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Pal, D.; Sur, S.; Mandal, S.; Saha, P.; Kumar, C. Pongapin and Karanjin, furanoflavanoids of Pongamia pinnata, induce G2/M arrest and apoptosis in cervical cancer cells by differential reactive oxygen species modulation, DNA damage, and nuclear factor kappa-light-chain-enhancer of activated B cell signaling. Phytother. Res. 2019, 33, 1084–1094. [Google Scholar] [PubMed]
- Amat-ur-Rasool, H.; Ahmed, M.; Hasnain, S.; Ahmed, A.; Carter, W.G. In Silico Design of Dual-Binding Site Anti-Cholinesterase Phytochemical Heterodimers as Treatment Options for Alzheimer’s Disease. Curr. Issues Mol. Biol. 2022, 44, 152–175. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hefni, M.E.; Witthöft, C.M. Characterization of Flavonoid Compounds in Common Swedish Berry Species. Foods 2020, 9, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Duan, W.; Zhang, J.; Huang, Y.; Zhang, Y.; Sun, B. Characterization and molecular docking study of taste peptides from chicken soup by sensory analysis combined with nano-LC-Q-TOF-MS/MS. Food Chem. 2022, 383, 132455. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Zhou, C.; Zhang, J.; Huang., Y.; Zhao, J.; Sun, B.; Zhang, Y. Characteristics of umami peptides identified from porcine bone soup and molecular docking to the taste receptor T1R1/T1R3. Food Chem. 2022, 387, 132870. [Google Scholar] [CrossRef]
- Mekar, N.; Yanti, E.; Levita, J. Structure-based in silico study of 6-gingerol, 6-ghogaol, and 6-paradol, active compounds of ginger (Zingiber officinale) as COX-2 inhibitors. Int. J. Chem. 2013, 5, 12–18. [Google Scholar]
- Zadorozhnii, P.V.; Kiselev, V.V.; Kharchenko, A.V. In Silico ADME Profiling of Salubrinal and Its Analogues. Future Pharmacol. 2022, 2, 160–197. [Google Scholar] [CrossRef]
- Lohning, A.; Marx, W.; Isenring, L. In silico investigation into the interactions between murine 5-HT3 receptor and the principle active compounds of ginger (Zingiber officinale). J. Mol. Graph. Model. 2016, 70, 315–327. [Google Scholar] [CrossRef]
- Volkamer, A.; Kuhn, D.; Grombacher, T.; Rippmann, F.; Rarey, M. Combining global and local measures for structure-based druggability preditions. J. Chem. Inf. Model. 2012, 52, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, M.R.; Bhuiyan, M.I.; Saha, A.; Mosleh, I.M.; Mondol, S.; Ahmed, C.M. Identification and analysis of potential targets in Streptococcus sanguinis using computer aided protein data analysis. Adv. Appl. Bioinform. Chem. 2014, 25, 45–54. [Google Scholar]
- BIOVIA. Available online: https://www.3ds.com/products-services/biovia/products/molecular-modeling-simulation/biovia-discovery-studio/ (accessed on 24 October 2022).
- Zhao, Y.; Abraham, M.; Le, J.; Hersey, A.; Luscombe, C.; Beck, G.; Sherborne, B.; Cooper, I. Rate limited steps of human oral absorption and QSAR studies. Pharm. Res. 2002, 19, 1446–1457. [Google Scholar] [CrossRef] [PubMed]
- Amatur-Rasool, H.; Ahmed, M. Designing Second Generation Anti-Alzheimer Compounds as Inhibitors of Human Acetylcholinesterase: Computational Screening of Synthetic Molecules and Dietary Phytochemicals. PLoS ONE 2015, 10, e0136509. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Tentative Identification | [M−H]− | Retention Time (min) | Theoretical Mass (m/z) | Measured Mass (m/z) | Accuracy (ppm) | Metabolite Type | MS Ions (ppm) |
|---|---|---|---|---|---|---|---|---|
| 1 | Mannitol | C6H13O6 | 1.34 | 181.0712 | 181.0723 | 3.9 | C | 151.0598 |
| 2 | Citric acid | C6H7O7 | 3.21 | 191.0192 | 191.0184 | 4.2 | Ac | 111.0074 |
| 3 | Swertianin | C14H9O6 | 10.67 | 273.0352 | 273.0349 | 10.2 | Ant | - |
| 4 | 2,4-Diformyl-3,5-dihydroxytoluene o 2,6-Diformyl-3,5-dihydroxytoluene | C9H7O4 | 13.28 | 179.0344 | 179.0353 | 4.7 | A | 107.0488; 135.0437; 151.0386 |
| 5 | 5,7-Dihydroxy-6-methylphtalide | C9H7O4 | 14.32 | 179.0344 | 179.0349 | 3.9 | A | 135.0438; 107.0488 |
| 6 | Atranol * | C8H7O3 | 19.82 | 151.0395 | 151.0401 | 8.3 | A | 135.0438; 123.0438; 107.0488 |
| 7 | Rhein | C15H7O6 | 20.33 | 283.02441 | 283.0221 | 2.5 | Ant | 273.011; 242.1745 |
| 8 | 9,10,12,13-Tetrahydroxyheneicosanoic acid | C21H41O6 | 20.73 | 389.2903 | 389.2897 | 2.5 | L | 371.2784 |
| 9 | Tetrahydroxypentacosanedioic acid | C25H47O8 | 21.2 | 475.3246 | 475.3252 | 2.2 | L | - |
| 10 | 9,10,12,13-Tetrahydroxydocosanoic acid | C22H43O6 | 21.30 | 403.3060 | 403.3028 | 3.9 | L | 385.2939; 215.1273 |
| 11 | Pentahydroxyoxohexacosanoic acid | C26H49O8 | 21.51 | 489.3432 | 489.3403 | −5.9 | L | 403. 3001; 979.6848 (2M-H) |
| 12 | Evernic acid * | C17H15O7 | 21.72 | 331.0818 | 331.0809 | 2.7 | d | 135.0438; 123.0439; 181.0494; 151.0386; 167.0336; 313.0703 |
| 13 | Brialmontin 2 | C21H25O5 | 22.31 | 343.1551 | 343.1567 | 2.8 | d | 123.0432; 313.0721 |
| 14 | 9,10,12,13-Tetrahidroxytricosanoic acid | C23H45O6 | 22.12 | 417.3236 | 417.3189 | 7.7 | L | 399.3095 |
| 15 | 9,10,12,13,14,15-Hexahydroxyheptacosenoic | C27H51O8 | 22.40 | 503.3584 | 503.3558 | 5.0 | L | 475.3615; 443.3355; 371.0377 |
| 16 | Methyl-9,10,11,12,13-pentahydroxy-14-oxoheptacosanoate | C28H53O8 | 22.72 | 517.3740 | 517.3685 | 6.3 | L | 457.3510; 431.3352 |
| 17 | Lecanoric acid | C16H13O7 | 23.81 | 317.0666 | 317.0624 | −10.92 | d | 167.034 |
| 18 | Barbatic acid * | C19H19O7 | 24.28 | 359.1131 | 359.1120 | 3.1 | d | 181.0493; 163.0387; 137.0594 |
| Peak | Tentative Identification | [M−H]− | Retention Time (min) | Theoretical Mass (m/z) | Measured Mass (m/z) | Accuracy (ppm) | Metabolite Type | MS Ions (ppm) |
|---|---|---|---|---|---|---|---|---|
| 1 | Mannitol | C6H13O6 | 1.34 | 181.0712 | 181.0723 | 3.9 | C | 151.0598 |
| 2 | Citric acid | C6H7O7 | 3.21 | 191.0192 | 191.0184 | 4.2 | Ac | 111.0074 |
| 3 | Azelaic acid | C9H15O4 | 14.70 | 187.0775 | 187.0969 | −3.63 | L | - |
| 4 | Visnagin | C13H9O5 | 17.30 | 245.0484 | 245.0431 | −22.2 | C | 165.0923 |
| 5 | Khellinol | C13H9O5 | 19.08 | 245.0489 | 245.0431 | −23.2 | C | 165.0914 |
| 6 | 9-Octadecenedioic acid | C18H31O4 | 19.56 | 311.2227 | 311.2228 | 0.2 | L | - |
| 7 | Pinellic acid | C18H33O5 | 20.16 | 329.2333 | 329.2345 | 3.6 | L | - |
| 8 | Olivetolic acid (2,4-Dihydroxy-6-pentylbenzoate) | C12H15O4 | 20.40 | 223.0983 | 223.0981 | 0.93 | L | 165.0923 |
| 9 | Pentahydroxyoxohexacosanoic acid | C26H49O8 | 22.17 | 489.3432 | 489.3561 | −7.8 | L | 403. 3001; 979.6848 (2M-H) |
| 10 | Lecanoric acid | C16H13O7 | 22.91 | 317.0666 | 317.0653 | −3.2 | d | 167.034 |
| 11 | Tetrahydroxytricosanoic acid | C23H46O6 | 23.14 | 417.3221 | 417.3230 | 2.0 | d | 245.0456 |
| 12 | 3,5-Diethoxybenzoic acid | C11H13O4 | 23.50 | 209.0822 | 209.0823 | 0.47 | A | 163.0360 |
| 13 | Sekikaic acid | C22H25O8 | 24.87 | 417.1553 | 417.3171 | −4.90 | A | 247.16944 |
| 14 | 17-Hydroxylinolenic acid | C18H29O3 | 25.19 | 293.2122 | 293.2136 | 4.9 | L | 243.19740 |
| 15 | Porrigenic acid | C18H29O4 | 25.31 | 309.2070 | 309.2091 | 6.51 | L | 291.19653 |
| 16 | Usnic acid * | C18H15O7 | 26.13 | 343.0823 | 343.0822 | −0.38 | DBF | 295.2291; 231.0647; 328.0570 |
| 17 | 18-Hydroxylinoleic acid | C18H31O3 | 26.87 | 295.22787 | 295.22878 | 2.8 | L | - |
| 18 | 18-Hydroxylinolenic acid | C18H29O3 | 27.89 | 293.2122 | 293.2136 | 4.7 | L | 243.19740 |
| Peak | Tentative Identification | [M−H]− | Retention Time (min) | Theoretical Mass (m/z) | Measured Mass (m/z) | Accuracy (ppm) | Metabolite Type | MS Ions (ppm) |
|---|---|---|---|---|---|---|---|---|
| 1 | Mannitol | C6H13O6 | 1.34 | 181.0712 | 181.0705 | 3.9 | C | 151.0598 |
| 2 | Vanillic acid | C8H8O4 | 13.25 | 167.0749 | 167.0754 | 3.0 | A | 123.0448 |
| 3 | Vanilloloside | C14H19O8 | 14.23 | 315.1085 | 315.1059 | −8.32 | A | 162.9945 |
| 4 | Methyl orsellinate | C9H9O4 | 20.23 | 181.0501 | 181.0507 | 0.5 | A | 151.0387; 123,0439; 135.0438 |
| 5 | 2,6-Diformyl-3,5-dihydroxytoluene | C9H7O4 | 20.81 | 179.0344 | 179.0338 | 3.4 | A | 151.0386; 107.0488; 135.0437 |
| 6 | Protolichesterinic acid | C19H31O4 | 21.80 | 323.2222 | 323.2213 | 2.8 | Ac | 279.2315; 267.2314 |
| 7 | 4′-O-methyl norhomosekikaic acid | C23H27O8 | 22.24 | 431.1657 | 431.1681 | 6.6 | d | 417.15290; 401.08231; 267.1228 |
| 8 | Sphaerophorin | C23H27O7 | 22.80 | 415.1757 | 415.1744 | 3.1 | d | 233.1166; 207.1376; 251.1275 |
| 9 | 6-Heptylresorcylic acid | C14H19O4 | 23.56 | 251.1288 | 251.1320 | 12.6 | A | 207.1403 |
| 10 | Lecanoric acid | C16H13O7 | 24.72 | 317.0666 | 317.0668 | 0.45 | d | 251.13175; 213.7944 |
| 11 | Sekikaic acid | C22H25O8 | 25.09 | 417.1554 | 417.15290 | −6.2 | d | 267.1228; 251.1289; 285.09033 |
| 12 | Usnic acid * | C18H15O7 | 26.13 | 343.0823 | 343.0822 | −0.38 | DBF | 295.2291; 231.0647; 328.0570 |
| 13 | Acetoxyisovalerylalkannin | C23H25O8 | 27.99 | 429.1514 | 429.1545 | −2.18 | A | 167.0360; 251.1298 |
| 14 | 2′-O-methyldivaricatic acid | C22H25O7 | 29.09 | 401.1605 | 401.1606 | 0.1 | A | 251.1321; 167.0358 |
| Assay | TPC (mg AG/g) | FRAP (µmol Trolox/g) | ORAC (µmol Trolox/g) |
|---|---|---|---|
| L. brialmontii | 0.279 ± 0.005 * | 45.089 ± 0.002 | 219.334 ± 0.75 * |
| P. pubescens | 0.579 ± 0.01 * | 46.422 ± 0.004 | 146.359 ± 0.56 * |
| S. globosus | 2.821 ± 0.08 * | 16.662 ± 0.004 * | 254.118 ± 0.82 * |
| Assay | AChE IC50 (µg/mL) | BChE IC50 (µg/mL) |
|---|---|---|
| L. brialmontii | 3.949 ± 0.04 * | 4.476 ± 0.06 * |
| P. pubescens | 2.805 ± 0.07 * | 8.828 ± 0.08 * |
| S. globosus | 10.422 ± 0.08 * | 6.785 ± 0.04 * |
| Galanthamine * | 0.26 ± 0.02 * | 3.82 ± 0.02 * |
| Compound. | Binding Energy (Kcal/mol) Acetylcholinesterase (TcAChE) | Binding Energy (Kcal/mol) Butyrylcholinestarase (hBChE) |
|---|---|---|
| Barbatic Acid | −10.30 | −8.80 |
| Lecanoric Acid | −9.90 | −9.40 |
| Brialmontin 2 | −9.80 | −9.10 |
| Tetrahydroxytricosanoic Acid | −7.90 | −6.60 |
| Sphaerophorin | −9.50 | −8.70 |
| Sekikaic Acid | −9.30 | −8.30 |
| Galanthamine | −10.80 | −8.80 |
| Compound | %ABS a | TPSA (Å2) b | MW c | cLogP d | HBD e | HBA f | n-ROTB g | Violation of Lipinski’s Rule |
|---|---|---|---|---|---|---|---|---|
| Rule | - | - | <500 | ≤5 | ≤5 | ≤10 | ≤10 | ≤1 |
| Barbatic Acid | 69.91 | 113.29 | 360.36 | 3.19 | 3 | 7 | 5 | 0 |
| Lecanoric Acid | 66.12 | 124.29 | 318.28 | 2.23 | 4 | 7 | 4 | 0 |
| Brialmontin 2 | 86.58 | 64.99 | 358.43 | 4.67 | 1 | 5 | 5 | 0 |
| Tetrahydroxytricosanoic Acid | 68.21 | 118.22 | 418.61 | 6.95 | 5 | 6 | 21 | 2 |
| Sphaerophorin | 69.91 | 113.29 | 416.47 | 5.19 | 3 | 7 | 11 | 1 |
| Sekikaic Acid | 66.73 | 122.52 | 418.44 | 4.17 | 3 | 8 | 10 | 0 |
| Galanthamine * | 94.53 | 41.93 | 287.35 | 1.19 | 1 | 4 | 3 | 0 |
| Compound | Mutagenic | Tumorigenic | Irritant | Reproductive Effect |
|---|---|---|---|---|
| Galanthamine | None | None | None | None |
| Barbatic Acid | None | None | None | None |
| Lecanoric Acid | None | None | None | None |
| Brialmontin 2 | None | High | High | None |
| Tetrahydroxytricosanoic acid | None | None | Low | Low |
| Sphaerophorin | None | None | None | None |
| Sekikaic Acid | None | None | None | None |
| Compound | Fraction of Molecule | Risk of Toxicity |
|---|---|---|
| Brialmontin 2 |  | High-risk fragment indicating Tumorigenicity |
 | High-risk fragment indicating Irritating |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Benítez, A.; Ortega-Valencia, J.E.; Sanchez, M.; Divakar, P.K.; Simirgiotis, M.J.; Gómez-Serranillos, M.P. Metabolomic Profiling, Antioxidant and Enzyme Inhibition Properties and Molecular Docking Analysis of Antarctic Lichens. Molecules 2022, 27, 8086. https://doi.org/10.3390/molecules27228086
Torres-Benítez A, Ortega-Valencia JE, Sanchez M, Divakar PK, Simirgiotis MJ, Gómez-Serranillos MP. Metabolomic Profiling, Antioxidant and Enzyme Inhibition Properties and Molecular Docking Analysis of Antarctic Lichens. Molecules. 2022; 27(22):8086. https://doi.org/10.3390/molecules27228086
Chicago/Turabian StyleTorres-Benítez, Alfredo, José Erick Ortega-Valencia, Marta Sanchez, Pradeep Kumar Divakar, Mario J. Simirgiotis, and María Pilar Gómez-Serranillos. 2022. "Metabolomic Profiling, Antioxidant and Enzyme Inhibition Properties and Molecular Docking Analysis of Antarctic Lichens" Molecules 27, no. 22: 8086. https://doi.org/10.3390/molecules27228086