

Pestalotiopsis Diversity: Species, Dispositions, Secondary Metabolites, and Bioactivities

Abstract
:1. Introduction
2. Habitat and Functional Diversity of Pestalotiopsis Species
3. New Secondary Metabolites in Pestalotiopsis Species
3.1. Pestalotiopsis diploclisia


3.2. Pestalotiopsis disseminate

3.3. Pestalotiopsis fici

3.4. Pestalotiopsis foedan

3.5. Pestalotiopsis guepinii

3.6. Pestalotiopsis heterocornis



3.7. Pestalotiopsis humus

3.8. Pestalotiopsis karstenii

3.9. Pestalotiopsis mangiferae
3.10. Pestalotiopsis microspora





3.11. Pestalotiopsis neglecta


3.12. Pestalotiopsis palmarum

3.13. Pestalotiopsis sydowiana

3.14. Pestalotiopsis theae


3.15. Pestalotiopsis uvicola


3.16. Pestalotiopsis vaccinii
3.17. Pestalotiopsis versicolor

3.18. Pestalotiopsis zonata
3.19. Pestalotiopsis sp.














4. Accurate Biosynthesis Pathways and Enhanced Accumulation of Secondary Metabolites in Pestalotiopsis
4.1. Transcription Factors Involved in Secondary Metabolite Biosynthesis in Pestalotiopsis
4.2. Histone Acetylation
4.3. Polyketide Synthases
4.4. Other Regulatory Proteins and Enzymes
5. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Subramanian, M.; Marudhamuthu, M. Hitherto unknown terpene synthase organization in taxol-producing endophytic bacteria isolated from marine macroalgae. Curr. Microbiol. 2020, 77, 918–923. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef]
- Gangadevi, V.; Muthumary, J. A Novel Endophytic taxol-producing fungus Chaetomella raphigera isolated from a medicinal plant, Terminalia arjuna. Appl. Biochem. Biotechnol. 2009, 158, 675–684. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, E.-S.R.; Zaki, A.G.; Ahmed, A.S.; Ismaiel, A.A. Production of the anticancer drug taxol by the endophytic fungus Epicoccum nigrum TXB502: Enhanced production by gamma irradiation mutagenesis and immobilization technique. Appl. Microbiol. Biotechnol. 2020, 104, 6991–7003. [Google Scholar] [CrossRef]
- Abdel-Fatah, S.S.; El-Batal, A.I.; El-Sherbiny, G.M.; Khalaf, M.A.; El-Sayed, A.S. Production, bioprocess optimization and γ-irradiation of Penicillium polonicum, as a new taxol producing endophyte from Ginko biloba. Biotechnol. Rep. 2021, 30, e00623. [Google Scholar] [CrossRef] [PubMed]
- Steyaert, R.L. Contributions à l’étude monographique de Pestalotia de Not. et Monochaetia Sacc. (Truncatella gen. nov. et Pestalotiopsis gen. nov.). Bull. Jard. Bot. Brux. 1949, 19, 285–354. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.; Guo, L.-D.; Chukeatirote, E.; Bahkali, A.; Hyde, K.D. Pestalotiopsis—Morphology, phylogeny, biochemistry and diversity. Fungal Divers. 2011, 50, 167–187. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.; Hyde, K.; Groenewald, J.; Xu, J.; Crous, P. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Ebada, S.S.; Proksch, P. Pestalotiopsis a highly creative genus: Chemistry and bioactivity of secondary metabolites. Fungal Divers. 2010, 44, 15–31. [Google Scholar] [CrossRef]
- Xu, J.; Yang, X.; Lin, Q. Chemistry and biology of Pestalotiopsis-derived natural products. Fungal Divers. 2014, 66, 37–68. [Google Scholar] [CrossRef]
- Strobel, G.; Yang, X.; Sears, J.; Kramer, R.; Sidhu, R.S.; Hess, W.M. Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology 1996, 142, 435–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, H.; Fujimoto, H.; Kimura, Y.; Hamasaki, T. Importance of the ketone function for the phytotoxicity of spiciferone A, a phytotoxin produced by the fungus Cochlioholus spicifer. Biosci. Biotechnol. Biochem. 1993, 57, 1938–1939. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Liu, H.; Niu, X.; Akhberdi, O.; Wei, D.; Wang, D.; Zhu, X. The Gα1-cAMP signaling pathway controls conidiation, development and secondary metabolism in the taxol-producing fungus Pestalotiopsis microspora. Microbiol. Res. 2017, 203, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Kathiravan, G.; Sureban, S.M. Effect of taxol from Pestalotiopsis mangiferae on A549 cells-in vitro study. J. Basic Clin. Pharm. 2010, 1, 1–9. [Google Scholar]
- Gangadevi, V.; Murugan, M.; Muthumary, J. Taxol determination from Pestalotiopsis pauciseta, a fungal endophyte of a medicinal plant. Chin. J. Biotechnol. 2008, 24, 1433–1438. [Google Scholar] [CrossRef]
- Kathiravan, G.; Raman, V.S. In vitro TAXOL production, by Pestalotiopsis breviseta—A first report. Fitoterapia 2010, 81, 557–564. [Google Scholar] [CrossRef]
- Kathiravan, G.; Sureban, S.M.; Sree, H.N.; Bhuvaneshwari, V.; Kramony, E. Isolation of anticancer drug TAXOL from Pestalotiopsis breviseta with apoptosis and B-Cell lymphoma protein docking studies. J. Basic Clin. Pharm. 2012, 4, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Gangadevi, V.; Muthumary, J. Taxol production by Pestalotiopsis terminaliae, an endophytic fungus of Terminalia arjuna (arjun tree). Biotechnol. Appl. Biochem. 2009, 52, 9–15. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, Y.; Ma, X.; Wang, C.; Yue, G.; Zhang, Y.; Zhang, Y.; Li, S.; Ling, S.; Liu, X.; et al. Greater taxol yield of fungus Pestalotiopsis hainanensis from dermatitic scurf of the giant panda (Ailuropoda melanoleuca). Appl. Biochem. Biotechnol. 2014, 175, 155–165. [Google Scholar] [CrossRef]
- Li, J.-Y.; Strobel, G.; Sidhu, R.; Hess, W.M.; Ford, E.J. Endophytic taxol-producing fungi from bald cypress, Taxodium distichum. Microbiology 1996, 142, 2223–2226. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-L.; Zhang, J.-Z.; Luo, D.-Q. The taxonomy, biology and chemistry of the fungal Pestalotiopsis genus. Nat. Prod. Rep. 2012, 29, 622–641. [Google Scholar] [CrossRef] [PubMed]
- Helaly, S.E.; Thongbai, B.; Stadler, M. Diversity of biologically active secondary metabolites from endophytic and saprotrophic fungi of the ascomycete order Xylariales. Nat. Prod. Rep. 2018, 35, 992–1014. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Stadler, M. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J. Antibiot. 2021, 74, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Sati, S.; Belwal, M. Aquatic hyphomycetes as endophytes of riparian plant roots. Mycologia 2005, 97, 45–49. [Google Scholar] [CrossRef]
- Aydoğdu, H.; Asan, A. Airborne fungi in child day care centers in Edirne City, Turkey. Environ. Monit. Assess. 2008, 147, 423–444. [Google Scholar] [CrossRef]
- Wang, J.; Liang, Z.; Li, K.; Yang, B.; Liu, Y.; Fang, W.; Tang, L.; Zhou, X. Ene-yne hydroquinones from a marine-derived strain of the fungus Pestalotiopsis neglecta with effects on liver X receptor alpha. J. Nat. Prod. 2020, 83, 1258–1264. [Google Scholar] [CrossRef]
- Wang, J.; Peng, Q.; Yao, X.; Liu, Y.; Zhou, X. New pestallic acids and diphenylketone derivatives from the marine alga-derived endophytic fungus Pestalotiopsis neglecta SCSIO41403. J. Antibiot. 2020, 73, 585–588. [Google Scholar] [CrossRef]
- Lei, H.; Zhou, X.; Hu, M.; Niu, H.; Song, C.; Chen, S.; Liu, Y.; Zhang, D. Cytotoxic polyketides from the marine sponge-derived fungus Pestalotiopsis heterocornis XWS03F09. Molecules 2019, 24, 2655. [Google Scholar] [CrossRef] [Green Version]
- Lei, H.; Lin, X.; Han, L.; Ma, J.; Dong, K.; Wang, X.; Zhong, J.; Mu, Y.; Liu, Y.; Huang, X. Polyketide derivatives from a marine-sponge-associated fungus Pestalotiopsis heterocornis. Phytochemistry 2017, 142, 51–59. [Google Scholar] [CrossRef]
- Lei, H.; Lin, X.; Han, L.; Ma, J.; Ma, Q.; Zhong, J.; Liu, Y.; Sun, T.; Wang, J.; Huang, X. New metabolites and bioactive chlorinated benzophenone derivatives produced by a marine-derived fungus Pestalotiopsis heterocornis. Mar. Drugs 2017, 15, 69. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Park, C.; Oh, E.; Sung, Y.; Lee, J.; Park, K.-H.; Kang, H. Benzophenone compounds, from a marine-derived strain of the fungus Pestalotiopsis neglecta, inhibit proliferation of pancreatic cancer cells by targeting the MEK/ERK pathway. J. Nat. Prod. 2019, 82, 3357–3365. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-F.; Liang, R.; Liao, S.-R.; Yang, B.; Tu, Z.-C.; Lin, X.-P.; Wang, B.-G.; Liu, Y. Vaccinols J–S, ten new salicyloid derivatives from the marine mangrove-derived endophytic fungus Pestalotiopsis vaccinii. Fitoterapia 2017, 120, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Parasuraman, P.; Devadatha, B.; Sarma, V.V.; Ranganathan, S.; Ampasala, D.R.; Kim, I.-W.; Patel, S.K.S.; Kalia, V.C.; Lee, J.-K.; Siddhardha, B. Inhibition of microbial quorum sensing mediated virulence factors by Pestalotiopsis sydowiana. J. Microbiol. Biotechnol. 2020, 30, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-Y.; Xue, D.-S.; Feng, X.-Y.; Yao, S.-J. Screening and production of ligninolytic enzyme by a marine-derived fungal Pestalotiopsis sp. J63. Appl. Biochem. Biotechnol. 2011, 165, 1754–1769. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, A.L.; de Medeiros, J.V.F.; Grault, C.E.; Santos, M.J.S.; Souza, A.L.A.; de Carvalho, R.W. The fungus Pestalotiopsis sp., isolated from Perna perna (Bivalvia:Mytilidae) cultured on marine farms in Southeastern Brazil and destined for human consumption. Mar. Pollut. Bull. 2020, 153, 110976. [Google Scholar] [CrossRef]
- Koh, S.; Mizuno, M.; Izuoka, Y.; Fujino, N.; Hamada-Sato, N.; Amano, Y. Xylanase from marine filamentous fungus Pestalotiopsis sp. AN-7 was activated with diluted salt solution like brackish water. J. Appl. Glycosci. 2021, 68, 11–18. [Google Scholar] [CrossRef]
- Wikee, S.; Hatton, J.; Turbé-Doan, A.; Mathieu, Y.; Daou, M.; Lomascolo, A.; Kumar, A.; Lumyong, S.; Sciara, G.; Faulds, C.B.; et al. Characterization and dye decolorization potential of two laccases from the marine-derived fungus Pestalotiopsis sp. Int. J. Mol. Sci. 2019, 20, 1864. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L.; Ge, H.M.; Li, F.; Song, Y.C.; Tan, R.X. New phytotoxic metabolites from Pestalotiopsis sp. HC02, a fungus residing in Chondracris rosee Gut. Chem. Biodivers. 2008, 5, 2402–2407. [Google Scholar] [CrossRef]
- Lv, C.; Huang, B.; Qiao, M.; Wei, J.; Ding, B. Entomopathogenic fungi on Hemiberlesia pitysophila. PLoS ONE 2011, 6, e23649. [Google Scholar] [CrossRef]
- Monden, Y.; Yamamoto, S.; Yamakawa, R.; Sunada, A.; Asari, S.; Makimura, K.; Inoue, Y. First case of fungal keratitis caused by Pestalotiopsis clavispora. Clin. Ophthalmol. 2013, 7, 2261–2264. [Google Scholar] [CrossRef] [Green Version]
- Magnan, R.F.; Rodrigues-Fo, E.; Daolio, C.; Ferreira, A.G.; de Souza, A.Q.L. Three highly oxygenated caryophyllene sesquiterpenes from Pestalotiopsis sp., a aungus Iiolated from bark of Pinus taeda. Zeitschrift für Naturforschung C 2003, 58, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.Q.; Zhang, L.; Shi, B.Z.; Song, X.M. Two new oxysporone derivatives from the fermentation broth of the endophytic plant fungus Pestalotiopsis karstenii isolated from stems of Camellia sasanqua. Molecules 2012, 17, 8554–8560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, H.; Wu, Y.; Ahmed, T.; Qi, X.; Li, B. Response of resistant and susceptible bayberry cultivars to infection of twig blight pathogen by histological observation and gibberellin related genes expression. Pathogens 2021, 10, 402. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-R.; Xing, Y.P.; Zhang, T.X.; Zheng, J.T.; Xu, J.Y.; Wang, Z.R.; Tong, Y.H. First report of Pestalotiopsis sydowiana causing leaf necrosis of Myrica rubra in China. Plant Dis. 2012, 96, 764. [Google Scholar] [CrossRef]
- Feng, Y.R.; Liu, B.S.; Sun, B.B. First Report of Leaf Blotch Caused by Pestalotiopsis clavispora on Rosa chinensis in China. Plant Dis. 2014, 98, 1009. [Google Scholar] [CrossRef]
- Jin, Y.-L.; Jiang, S.-L.; Jiang, X.-L. Disease-resistant identification and analysis to transcriptome differences of blueberry leaf spot induced by beta-aminobutyric acid. Arch. Microbiol. 2021, 203, 3623–3632. [Google Scholar] [CrossRef]
- Mahadevakumar, S.; Janardhana, G.R. First report of Pestalotiopsis species causing leaf spot of cowpea (Vigna unguiculata) in India. Plant Dis. 2014, 98, 686. [Google Scholar] [CrossRef]
- Nozawa, S.; Seto, Y.; Watanabe, K. First report of leaf blight caused by Pestalotiopsis chamaeropis and Neopestalotiopsis sp. in Japanese andromeda. J. Gen. Plant Pathol. 2019, 85, 449–452. [Google Scholar] [CrossRef]
- Akhberdi, O.; Zhang, Q.; Wang, H.; Li, Y.; Chen, L.; Wang, D.; Yu, X.; Wei, D.; Zhu, X. Roles of phospholipid methyltransferases in pycnidia development, stress tolerance, and secondary metabolism in the taxol-producing fungus Pestalotiopsis microspora. Microbiol. Res. 2018, 210, 33–42. [Google Scholar] [CrossRef]
- Prasannath, K.; Galea, V.J.; Akinsanmi, O.A. Influence of climatic factors on dry flower, grey and green mould diseases of macadamia flowers in Australia. J. Appl. Microbiol. 2021, 132, 1291–1306. [Google Scholar] [CrossRef]
- Keith, L.M. First report of Pestalotiopsis virgatula causing Pestalotiopsis Fruit Rot on rambutan in Hawaii. Plant Dis. 2008, 92, 835. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Zheng, Z.; Liu, S.; Zhang, H.; Guo, L.; Che, Y. Photinides A−F, cytotoxic benzofuranone-derived γ-lactones from the plant endophytic fungus Pestalotiopsis photiniae. J. Nat. Prod. 2009, 72, 942–945. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Xie, C.-X.; Chen, Q.-W.; Yu, Z.-D. Pestalotiopsis trachicarpicola, a novel pathogen causes twig blight of Pinus bungeana (Pinaceae: Pinoideae) in China. Antonie Van Leeuwenhoek 2021, 114, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sowndhararajan, K.; Marimuthu, S.; Manian, S. Biocontrol potential of phylloplane bacterium Ochrobactrum anthropi BMO-111 against blister blight disease of tea. J. Appl. Microbiol. 2013, 114, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiong, F.; Lu, Q.; Hao, X.; Zheng, M.; Wang, L.; Li, N.; Ding, C.; Wang, X.; Yang, Y. Diversity of Pestalotiopsis-Like species causing gray blight disease of tea plants (Camellia sinensis) in China, including two novel Pestalotiopsis species, and analysis of their pathogenicity. Plant Dis. 2019, 103, 2548–2558. [Google Scholar] [CrossRef] [PubMed]
- Akinsanmi, O.A.; Nisa, S.; Jeff-Ego, O.S.; Shivas, R.G.; Drenth, A. Dry flower disease of Macadamia in Australia caused by Neopestalotiopsis macadamiae sp. nov. and Pestalotiopsis macadamiae sp. nov. Plant Dis. 2017, 101, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Lu, L.; Wang, D.; Wang, Y.; Ni, H.; Du, Z. Biological characterization and genetic diversity analysis of two species of Pestalotiopsis causing twig dieback of Myrica rubra. Eur. J. Plant Pathol. 2013, 136, 737–747. [Google Scholar] [CrossRef]
- Espinoza, J.G.; Briceño, E.X.; Keith, L.M.; Latorre, B.A. Canker and twig dieback of blueberry caused by Pestalotiopsis spp. and a Truncatella sp. in Chile. Plant Dis. 2008, 92, 1407–1414. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Ceja, A.; Loeza-Lara, P.; Espinosa-García, F.; García-Rodríguez, Y.; Medina-Medrano, J.; Gutiérrez-Hernández, G.; Ceja-Torres, L. In Vitro antifungal activity of plant extracts on pathogenic fungi of blueberry (Vaccinium sp.). Plants 2021, 10, 852. [Google Scholar] [CrossRef]
- Singh, N.I.; Devi, R.K.T. First report of broad bean canker caused by Pestalotiopsis disseminata in India. Plant Dis. 2001, 85, 229. [Google Scholar] [CrossRef]
- Yun, Y.H.; Ahn, G.R.; Kim, S.H. First Report and Characterization of Pestalotiopsis ellipsospora Causing Canker on Acanthopanax divaricatus. Mycobiology 2015, 43, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Palou, L.; Montesinos-Herrero, C.; Tarazona, I.; Besada, C.; Taberner, V. Incidence and etiology of postharvest Fungal diseases of persimmon (Diospyros kaki Thunb. cv. Rojo Brillante) in Spain. Plant Dis. 2015, 99, 1416–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maharachchikumbura, S.S.; Guo, L.-D.; Chukeatirote, E.; McKenzie, E.H.; Hyde, K.D. A destructive new disease of Syzygium samarangense in Thailand caused by the new species Pestalotiopsis samarangensis. Trop. Plant Pathol. 2013, 38, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Tang, Q.; Huang, K.H. First report of leaf blight of Eriobotrya japonica caused by Pestalotiopsis microspora in Anhui Province, China. Plant Dis. 2016, 100, 1014. [Google Scholar] [CrossRef]
- Jeon, Y.H.; Cheon, W. First report of leaf blight of Japanese yew caused by Pestalotiopsis microspora in Korea. Plant Dis. 2014, 98, 691. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Wu, P.; Liu, Z.; Xue, J.; Li, H.; Wei, X. 2H-pyranone and isocoumarin derivatives from the endophytic fungus Pestalotiopsis microspora SC3082 derived from Scaevola taccada (Gaertn.) Roxb. Nat. Prod. Res. 2021, 35, 3644–3651. [Google Scholar] [CrossRef]
- Nalin Rathnayake, G.R.; Kumar, N.S.; Jayasinghe, L.; Araya, H.; Fujimoto, Y. Secondary metabolites produced by an endophytic fungus Pestalotiopsis microspora. Nat. Prod. Bioprospecting 2019, 9, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Riga, R.; Happyana, N.; Hakim, E.H. Sesquiterpenes produced by Pestalotiopsis microspora HF 12440 isolated from Artocarpus heterophyllus. Nat. Prod. Res. 2020, 34, 2229–2231. [Google Scholar] [CrossRef]
- Li, E.; Jiang, L.; Guo, L.; Zhang, H.; Che, Y. Pestalachlorides A–C, antifungal metabolites from the plant endophytic fungus Pestalotiopsis adusta. Bioorganic Med. Chem. 2008, 16, 7894–7899. [Google Scholar] [CrossRef]
- Xu, M.-F.; Jia, O.-Y.; Wang, S.-J.; Zhu, Q. A new bioactive diterpenoid from pestalotiopsis adusta, an endophytic fungus from Clerodendrum canescens. Nat. Prod. Res. 2016, 30, 2642–2647. [Google Scholar] [CrossRef]
- Liu, H.; Liu, S.; Guo, L.; Zhang, Y.; Cui, L.; Ding, G. New furanones from the plant endophytic fungus Pestalotiopsis besseyi. Molecules 2012, 17, 14015–14021. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-L.; Huang, L.; Ruan, X.-L. Epigenetic modifiers alter the secondary metabolite composition of a plant endophytic fungus, Pestalotiopsis crassiuscula obtained from the leaves of Fragaria chiloensis. J. Asian Nat. Prod. Res. 2014, 16, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Bunyapaiboonsri, T.; Yoiprommarat, S.; Nithithanasilp, S.; Choowong, W.; Preedanon, S.; Suetrong, S. Two new farnesyl hydroquinones from Pestalotiopsis diploclisia (BCC 35283), the fungus associated with algae. Nat. Prod. Res. 2021, 30, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Deyrup, S.T.; Swenson, D.C.; Gloer, J.B.; Wicklow, D.T. Caryophyllene sesquiterpenoids from a fungicolous isolate of Pestalotiopsis disseminata. J. Nat. Prod. 2006, 69, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.H.; Swenson, D.C.; Gloer, J.B.; Wicklow, D.T. Disseminins and spiciferone analogues: Polyketide-derived metabolites from a fungicolous isolate of Pestalotiopsis disseminata. J. Nat. Prod. 2016, 79, 523–530. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, L.; Guan, F.; Li, E.; Jin, J.; Li, J.; Che, Y.; Liu, G. Characterization of a prenyltransferase for Iso-A82775C biosynthesis and generation of new congeners of chloropestolides. ACS Chem. Biol. 2018, 13, 703–711. [Google Scholar] [CrossRef]
- Liu, L.; Tian, R.; Liu, S.; Chen, X.; Guo, L.; Che, Y. Pestaloficiols A–E, bioactive cyclopropane derivatives from the plant endophytic fungus Pestalotiopsis fici. Bioorganic Med. Chem. 2008, 16, 6021–6026. [Google Scholar] [CrossRef]
- Wang, X.; Wu, F.; Liu, L.; Liu, X.; Che, Y.; Keller, N.P.; Guo, L.; Yin, W.-B. The bZIP transcription factor PfZipA regulates secondary metabolism and oxidative stress response in the plant endophytic fungus Pestalotiopsis fici. Fungal Genet. Biol. 2015, 81, 221–228. [Google Scholar] [CrossRef]
- Liu, L.; Liu, S.; Chen, X.; Guo, L.; Che, Y. Pestalofones A–E, bioactive cyclohexanone derivatives from the plant endophytic fungus Pestalotiopsis fici. Bioorganic Med. Chem. 2009, 17, 606–613. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; Liu, S.; Zheng, Z.; Chen, X.; Zhang, H.; Guo, L.; Che, Y. Chloropestolide A, an antitumor metabolite with an unprecedented spiroketal skeleton from Pestalotiopsis fici. Org. Lett. 2009, 11, 2836–2839. [Google Scholar] [CrossRef]
- Liu, L.; Bruhn, T.; Guo, L.; Götz, D.C.G.; Brun, R.; Stich, A.; Che, Y.; Bringmann, G. Chloropupukeanolides C-E: Cytotoxic pupukeanane chlorides with a spiroketal skeleton from Pestalotiopsis fici. Chem. A Eur. J. 2011, 17, 2604–2613. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, L.; Zhang, F.; Wang, W.; Li, J.; Guo, L.; Che, Y.; Liu, G. Identification of the first diphenyl ether gene cluster for pestheic acid biosynthesis in plant endophyte Pestalotiopsis fici. Chem Bio Chem 2014, 15, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, S.; Jiang, L.; Chen, X.; Guo, L.; Che, Y. Chloropupukeananin, the first chlorinated pupukeanane derivative, and its precursors from Pestalotiopsis fici. Org. Lett. 2008, 10, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, X.; Fan, A.; Zheng, Y.; Liu, X.; Wang, S.; Zou, H.; Oakley, B.R.; Keller, N.P.; Yin, W.-B. A cryptic pigment biosynthetic pathway uncovered by heterologous expression is essential for conidial development in Pestalotiopsis fici. Mol. Microbiol. 2017, 105, 469–483. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Niu, S.; Lu, X.; Chen, X.; Zhang, H.; Guo, L.; Che, Y. Unique metabolites of Pestalotiopsis fici suggest a biosynthetic hypothesis involving a Diels–Alder reaction and then mechanistic diversification. Chem. Commun. 2010, 46, 460–462. [Google Scholar] [CrossRef]
- Liu, L.; Liu, S.; Niu, S.; Guo, L.; Chen, X.; Che, Y. Isoprenylated chromone derivatives from the plant endophytic fungus Pestalotiopsis fici. J. Nat. Prod. 2009, 72, 1482–1486. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; Li, L.; Cao, Y.; Guo, L.; Liu, G.; Che, Y. Spiroketals of Pestalotiopsis fici provide evidence for a biosynthetic hypothesis involving diversified Diels–Alder reaction cascades. J. Org. Chem. 2013, 78, 2992–3000. [Google Scholar] [CrossRef]
- Zheng, Y.; Ma, K.; Lyu, H.; Huang, Y.; Liu, H.; Liu, L.; Che, Y.; Liu, X.; Zou, H.; Yin, W.-B. Genetic manipulation of the COP9 signalosome subunit PfCsnE leads to the discovery of pestaloficins in Pestalotiopsis fici. Org. Lett. 2017, 19, 4700–4703. [Google Scholar] [CrossRef]
- Liu, S.; Guo, L.; Che, Y.; Liu, L. Pestaloficiols Q–S from the plant endophytic fungus Pestalotiopsis fici. Fitoterapia 2013, 85, 114–118. [Google Scholar] [CrossRef]
- Zhang, P.; Zhou, S.; Wang, G.; An, Z.; Liu, X.; Li, K.; Yin, W. Two transcription factors cooperatively regulate DHN melanin biosynthesis and development in Pestalotiopsis fici. Mol. Microbiol. 2019, 112, 649–666. [Google Scholar] [CrossRef]
- Liu, S.; Liu, X.; Guo, L.; Che, Y.; Liu, L. 2H-pyran-2-one and 2H-furan-2-one derivatives from the plant endophytic fungus Pestalotiopsis fici. Chem. Biodivers. 2013, 10, 2007–2013. [Google Scholar] [CrossRef] [PubMed]
- Rao, L.; You, Y.-X.; Su, Y.; Liu, Y.; He, Q.; Fan, Y.; Hu, F.; Xu, Y.-K.; Zhang, C.-R. Two spiroketal derivatives with an unprecedented amino group and their cytotoxicity evaluation from the endophytic fungus Pestalotiopsis flavidula. Fitoterapia 2019, 135, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Liu, S.; Guo, L.; Zhou, Y.; Che, Y. Antifungal metabolites from the plant endophytic fungus Pestalotiopsis foedan. J. Nat. Prod. 2008, 71, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-L.; Li, Z.-Z. New spiral γ-lactone enantiomers from the plant endophytic fungus Pestalotiopsis foedan. Molecules 2013, 18, 2236–2242. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Zhang, B.-Y.; Yang, X.-L. Antifungal monoterpene derivatives from the plant endophytic fungus Pestalotiopsis foedan. Chem. Biodivers. 2016, 13, 1422–1425. [Google Scholar] [CrossRef] [PubMed]
- Parshikov, I.A.; Heinze, T.M.; Moody, J.D.; Freeman, J.P.; Williams, A.J.; Sutherland, J.B. The fungus Pestalotiopsis guepini as a model for biotransformation of ciprofloxacin and norfloxacin. Appl. Microbiol. Biotechnol. 2001, 56, 474–477. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, K.F.; Hesse, M.; Werner, C. Antimicrobial activities of secondary metabolites produced by endophytic fungi from Spondias mombin. J. Basic Microbiol. 2000, 40, 261–267. [Google Scholar] [CrossRef]
- Sousa, J.; Matos, L.; Alcântara, D.; Ribeiro, H.; Santos, L.; Oliveira, M.; Brito-Junior, L.; Khayat, A.; Guimarães, A.; Cunha, L.; et al. Cellular responses induced in vitro by pestheic acid, a fungal metabolite, in a gastric adenocarcinoma cell line (PG100). Genet. Mol. Res. 2013, 12, 4106–4115. [Google Scholar] [CrossRef]
- Evidente, A.; Zonno, M.C.; Andolfi, A.; Troise, C.; Cimmino, A.; Vurro, M. Phytotoxic α-pyrones produced by Pestalotiopsis guepinii, the causal agent of hazelnut twig blight. J. Antibiot. 2012, 65, 203–206. [Google Scholar] [CrossRef]
- Zhang, Y.; Bai, J.; Yan, D.; Liu, B.; Zhang, L.; Zhang, C.; Chen, M.; Mou, Y.; Hu, Y. Highly oxygenated caryophyllene-type Sesquiterpenes from a plant-associated fungus, Pestalotiopsis hainanensis, and their biosynthetic gene cluster. J. Nat. Prod. 2020, 83, 3262–3269. [Google Scholar] [CrossRef]
- Xing, J.-G.; Deng, H.-Y.; Luo, D.-Q. Two new compounds from an endophytic fungus Pestalotiopsis heterocornis. J. Asian Nat. Prod. Res. 2011, 13, 1069–1073. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Hirose, T.; Iwatsuki, M.; Chinen, T.; Kimura, T.; Suga, T.; Nonaka, K.; Nakashima, T.; Sunazuka, T.; Usui, T.; et al. Pestynol, an antifungal compound discovered using a Saccharomyces cerevisiae 12 gene Δ0HSR-iERG6-based assay. J. Nat. Prod. 2018, 81, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Suga, T.; Iwatsuki, M.; Chinen, T.; Nonaka, K.; Usui, T.; Asami, Y.; Ōmura, S.; Shiomi, K. Pestiocandin, a new papulacandin class antibiotic isolated from Pestalotiopsis humus. J. Antibiot. 2018, 71, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Strobel, G.A. Jesterone and hydroxy-jesterone antioomycete cyclohexenone epoxides from the endophytic fungus Pestalotiopsis jesteri. Phytochemistry 2001, 57, 261–265. [Google Scholar] [CrossRef]
- Kumar, D.S.S.; Lau, C.S.; Wan, J.M.; Yang, D.; Hyde, K.D. Immunomodulatory compounds from Pestalotiopsis leucothës, an endophytic fungus from Tripterygium wilfordii. Life Sci. 2005, 78, 147–156. [Google Scholar] [CrossRef]
- Kumar, S.S.D.; Lau, C.S.; Chan, W.K.; Yang, D.; Cheung, H.Y.; Chen, F.; Hyde, K.D. Immunomodulatory activity of an endophytic fungus from Tripterygium wilfordii. In Proceedings of the 2nd International Conference on Medicinal Mushroom and the International Conference on Biodiversity and Bioactive Compounds, Pattaya, Thailand, 17–19 July 2003; pp. 367–373. [Google Scholar]
- Subban, K.; Subramani, R.; Johnpaul, M. A novel antibacterial and antifungal phenolic compound from the endophytic fungus Pestalotiopsis mangiferae. Nat. Prod. Res. 2013, 27, 1445–1449. [Google Scholar] [CrossRef]
- Metz, A.M.; Haddad, A.; Worapong, J.; Long, D.M.; Ford, E.J.; Hess, W.M.; Strobel, G.A. Induction of the sexual stage of Pestalotiopsis microspora, a taxol-producing fungus The GenBank accession numbers for the sequences determined in this work are: Pestalotiopsis microspora NE-32 18S rDNA, AF104356; Pestalosphaeria hansenii ATCC 48245 18S rDNA, AF242846. Microbiology 2000, 146, 2079–2089. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Guo, Z.; Deng, Z.; Yang, J.; Zou, K. A new ɑ-pyrone derivative from endophytic fungus Pestalotiopsis microspora. Recf. Nat. Prod. 2015, 9, 503–508. [Google Scholar]
- Strobel, G.; Ford, E.; Worapong, J.; Harper, J.K.; Arif, A.M.; Grant, D.M.; Fung, P.C.; Chau, R.M.W. Isopestacin, an isobenzofuranone from Pestalotiopsis microspora, possessing antifungal and antioxidant activities. Phytochemistry 2002, 60, 179–183. [Google Scholar] [CrossRef]
- Chen, L.; Li, Y.; Zhang, Q.; Wang, D.; Akhberdi, O.; Wei, D.; Pan, J.; Zhu, X. Improved pestalotiollide B production by deleting competing polyketide synthase genes in Pestalotiopsis microspora. J. Ind. Microbiol. Biotechnol. 2017, 44, 237–246. [Google Scholar] [CrossRef]
- Niu, X.; Hao, X.; Hong, Z.; Chen, L.; Yu, X.; Zhu, X. A Putative histone deacetylase modulates the biosynthesis of pestalotiollide B and conidiation in Pestalotiopsis microspora. J. Microbiol. Biotechnol. 2015, 25, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, L.; Yu, X.; Liu, H.; Akhberdi, O.; Pan, J.; Zhu, X. A B-type histone acetyltransferase Hat1 regulates secondary metabolism, conidiation, and cell wall integrity in the taxol-producing fungus Pestalotiopsis microspora. J. Basic Microbiol. 2016, 56, 1380–1391. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Akhberdi, O.; Hao, X.; Yu, X.; Chen, L.; Liu, Y.; Zhu, X. Amino acid Sensor kinase Gcn2 is required for conidiation, secondary metabolism, and cell wall integrity in the taxol-producer Pestalotiopsis microspora. Front. Microbiol. 2017, 8, 1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subban, K.; Singh, S.; Subramani, R.; Johnpaul, M.; Chelliah, J. Fungal 7-epi-10-deacetyltaxol produced by an endophytic Pestalotiopsis microspora induces apoptosis in human hepatocellular carcinoma cell line (HepG2). BMC Complement. Altern. Med. 2017, 17, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Huo, L.; Liu, H.; Chen, L.; Wang, Y.; Zhu, X. Melanin is required for the formation of the multi-cellular conidia in the endophytic fungus Pestalotiopsis microspora. Microbiol. Res. 2015, 179, 1–11. [Google Scholar] [CrossRef]
- Fu, S.-B.; Yang, J.-S.; Cui, J.-L.; Meng, Q.-F.; Feng, X.; Sun, D.-A. Multihydroxylation of ursolic acid by Pestalotiopsis microspora isolated from the medicinal plant Huperzia serrata. Fitoterapia 2011, 82, 1057–1061. [Google Scholar] [CrossRef]
- Liu, S.; Dai, H.; Makhloufi, G.; Heering, C.; Janiak, C.; Hartmann, R.; Mándi, A.; Kurtán, T.; Müller, W.E.G.; Kassack, M.U.; et al. Cytotoxic 14-membered macrolides from a mangrove-derived endophytic fungus, Pestalotiopsis microspora. J. Nat. Prod. 2016, 79, 2332–2340. [Google Scholar] [CrossRef]
- Brückner, D.; Hafner, F.-T.; Li, V.; Schmeck, C.; Telser, J.; Vakalopoulos, A.; Wirtz, G. Dibenzodioxocinones—A new class of CETP inhibitors. Bioorganic Med. Chem. Lett. 2005, 15, 3611–3614. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, L.; Xie, Q.; Yu, X.; Duan, A.; Lin, Y.; Xiang, B.; Hao, X.; Chen, W.; Zhu, X. A gene cluster for the biosynthesis of dibenzodioxocinons in the endophyte Pestalotiopsis microspora, a taxol producer. J. Microbiol. Biotechnol. 2019, 29, 1570–1579. [Google Scholar] [CrossRef]
- Kanno, K.; Tsurukawa, Y.; Kamisuki, S.; Shibasaki, H.; Iguchi, K.; Murakami, H.; Uchiyama, J.; Kuramochi, K. Novel neuroprotective hydroquinones with a vinyl alkyne from the fungus, Pestalotiopsis microspora. J. Antibiot. 2019, 72, 793–799. [Google Scholar] [CrossRef]
- Wu, X.; Wang, Y.; Liu, S.; Liu, X.; Guo, L. Microsporols A-C from the plant endophytic fungus Pestalotiopsis microspora. Nat. Prod. Commun. 2015, 10, 1643–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Q.-Y.; Li, E.-W.; Han, J.-J.; Pei, Y.-F.; Ma, K.; Bao, L.; Huang, Y.; Zhao, F.; Liu, H.-W. New ambuic acid derivatives from the solid culture of Pestalotiopsis neglecta and their nitric oxide inhibitory activity. Sci. Rep. 2015, 5, srep09958. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Pramanik, A.; Agrawal, P.K. Evaluation of bioactive secondary metabolites from endophytic fungus Pestalotiopsis neglecta BAB-5510 isolated from leaves of Cupressus torulosa D.Don. 3 Biotech 2016, 6, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.; Gu, T.; Wang, J.; She, J.; Ye, Y.; Cao, W.; Luo, X.; Xiao, J.; Liu, Y.; Tang, L.; et al. Chromene and chromone derivatives as liver X receptors modulators from a marine-derived Pestalotiopsis neglecta fungus. Bioorganic Chem. 2021, 112, 104927. [Google Scholar] [CrossRef]
- Zhang, Q.; Luan, R.; Li, H.; Liu, Y.; Liu, P.; Wang, L.; Li, D.; Wang, M.; Zou, Q.; Liu, H.; et al. Anti-inflammatory action of ambuic acid, a natural product isolated from the solid culture of Pestalotiopsis neglecta, through blocking ERK/JNK mitogen-activated protein kinase signaling pathway. Exp. Ther. Med. 2018, 16, 1538–1546. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Han, J.; Wang, J.; Zhang, A.-X.; Miao, Y.-Y.; Tan, N.-H.; Wang, Z. Pestalopyrones A–D, four tricyclic pyrone derivatives from the endophytic fungus Pestalotiopsis neglecta S3. Phytochemistry 2020, 179, 112505. [Google Scholar] [CrossRef]
- Xiao, J.; Hu, J.-Y.; Sun, H.-D.; Zhao, X.; Zhong, W.-T.; Duan, D.-Z.; Wang, L.; Wang, X.-L. Sinopestalotiollides A–D, cytotoxic diphenyl ether derivatives from plant endophytic fungus Pestalotiopsis palmarum. Bioorganic Med. Chem. Lett. 2018, 28, 515–518. [Google Scholar] [CrossRef]
- Yang, X.-L.; Zhang, S.; Hu, Q.-B.; Luo, D.-Q.; Zhang, Y. Phthalide derivatives with antifungal activities against the plant pathogens isolated from the liquid culture of Pestalotiopsis photiniae. J. Antibiot. 2011, 64, 723–727. [Google Scholar] [CrossRef]
- Ding, G.; Qi, Y.; Liu, S.; Guo, L.; Chen, X. Photipyrones A and B, new pyrone derivatives from the plant endophytic fungus Pestalotiopsis photiniae. J. Antibiot. 2012, 65, 271–273. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Yang, R. A phthalide derivative isolated from endophytic fungi Pestalotiopsis photiniae induces G1 cell cycle arrest and apoptosis in human HeLa cells. Braz. J. Med. Biol. Res. 2013, 46, 643–649. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Hu, S.-Y.; Luo, D.-Q.; Zhu, S.-Y.; Zhou, C.-Q. Potential antitumor agent from the endophytic fungus Pestalotiopsis photiniae induces apoptosis via the mitochondrial pathway in HeLa cells. Oncol. Rep. 2013, 30, 1773–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.-L.; Zhang, S.; Zhu, H.-J.; Luo, D.-Q. Dihydroberkleasmin A: A New eremophilane sesquiterpenoid from the fermentation broth of the plant endophytic fungus Pestalotiopsis photiniae. Molecules 2011, 16, 1910–1916. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Ando, K.; Aotani, Y.; Shinoda, K.; Tanaka, T.; Tsukuda, E.; Yoshida, M.; Matsuda, Y. RES-1214-1 and -2, Novel non-peptidic endothelin type A receptor antagonists produced by Pestalotiopsis sp. J. Antibiot. 1995, 48, 1401–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Lin, Q.; Wang, B.; Wray, V.; Lin, W.-H.; Proksch, P. Pestalotiopamide E, a new amide from the endophytic fungus Pestalotiopsis sp. J. Asian Nat. Prod. Res. 2011, 13, 373–376. [Google Scholar] [CrossRef]
- Hemberger, Y.; Xu, J.; Wray, V.; Proksch, P.; Wu, J.; Bringmann, G. Pestalotiopens A and B: Stereochemically challenging flexible sesquiterpene-cyclopaldic acid hybrids from Pestalotiopsis sp. Chem. A Eur. J. 2013, 19, 15556–15564. [Google Scholar] [CrossRef]
- Davis, R.A.; Carroll, A.R.; Andrews, K.T.; Boyle, G.M.; Tran, T.L.; Healy, P.C.; Kalaitzis, J.A.; Shivas, R.G. Pestalactams A–C: Novel caprolactams from the endophytic fungus Pestalotiopsis sp. Org. Biomol. Chem. 2010, 8, 1785–1790. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.-Y.; Sun, W.-G.; Liu, J.-J.; Wang, J.-P.; Hu, Z.-X.; Zhang, Y.-H. A new pair of cytotoxic enantiomeric isoprenylated chromone derivatives from Pestalotiopsis sp. J. Asian Nat. Prod. Res. 2021, 8, 1–7. [Google Scholar] [CrossRef]
- Ding, G.; Zhang, F.; Chen, H.; Guo, L.; Zou, Z.; Che, Y. Pestaloquinols A and B, Isoprenylated epoxyquinols from Pestalotiopsis sp. J. Nat. Prod. 2011, 74, 286–291. [Google Scholar] [CrossRef]
- Zhou, J.; Li, G.; Deng, Q.; Zheng, D.; Yang, X.; Xu, J. Cytotoxic constituents from the mangrove endophytic Pestalotiopsis sp. induce G0/G1 cell cycle arrest and apoptosis in human cancer cells. Nat. Prod. Res. 2018, 32, 2968–2972. [Google Scholar] [CrossRef]
- Hwang, I.H.; Swenson, D.C.; Gloer, J.B.; Wicklow, D.T. Pestaloporonins: Caryophyllene-derived sesquiterpenoids from a fungicolous isolate of Pestalotiopsis sp. Org. Lett. 2015, 17, 4284–4287. [Google Scholar] [CrossRef]
- Yuan, C.; Ding, G.; Wang, H.-Y.; Guo, Y.-H.; Shang, H.; Ma, X.-J.; Zou, Z.-M. Polyketide-terpene hybrid metabolites from an endolichenic fungus Pestalotiopsis sp. BioMed Res. Int. 2017, 2017, 6961928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Yang, M.-H.; Wang, X.-B.; Li, T.-X.; Kong, L.-Y. Caryophyllene sesquiterpenoids from the endophytic fungus, Pestalotiopsis sp. Fitoterapia 2016, 109, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Song, R.-Y.; Wang, X.-B.; Yin, G.-P.; Liu, R.-H.; Kong, L.-Y.; Yang, M.-H. Isocoumarin derivatives from the endophytic fungus, Pestalotiopsis sp. Fitoterapia 2017, 122, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.-Y.; Li, D.; Shao, C.-L.; Deng, D.-S.; Wang, C.-Y. (±)-Pestalachloride D, an antibacterial racemate of Chlorinated benzophenone derivative from a soft coral-derived fungus Pestalotiopsis sp. Mar. Drugs 2013, 11, 1050–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Kjer, J.; Sendker, J.; Wray, V.; Guan, H.; Edrada, R.; Müller, W.E.; Bayer, M.; Lin, W.; Wu, J.; et al. Cytosporones, coumarins, and an alkaloid from the endophytic fungus Pestalotiopsis sp. isolated from the Chinese mangrove plant Rhizophora mucronata. Bioorganic Med. Chem. 2009, 17, 7362–7367. [Google Scholar] [CrossRef]
- Jia, Y.-L.; Wei, M.-Y.; Chen, H.-Y.; Guan, F.-F.; Wang, C.-Y.; Shao, C.-L. (+)- and (−)-Pestaloxazine A, a aair of antiviral enantiomeric alkaloid dimers with a symmetric spiro[oxazinane-piperazinedione] skeleton from Pestalotiopsis sp. Org. Lett. 2015, 17, 4216–4219. [Google Scholar] [CrossRef]
- Pulici, M.; Sugawara, F.; Koshino, H.; Uzawa, J.; Yoshida, S. Pestalotiopsins A and B: New caryophyllenes from an endophytic fungus of Taxus brevifolia. Phytochemistry 1996, 61, 2122–2124. [Google Scholar] [CrossRef]
- Ding, G.; Li, Y.; Fu, S.; Liu, S.; Wei, J.; Che, Y. Ambuic acid and torreyanic acid derivatives from the endolichenic fungus Pestalotiopsis sp. J. Nat. Prod. 2009, 72, 182–186. [Google Scholar] [CrossRef]
- Xu, J.; Kjer, J.; Sendker, J.; Wray, V.; Guan, H.; Edrada, R.; Lin, W.; Wu, J.; Proksch, P. Chromones from the Endophytic Fungus Pestalotiopsis sp. Isolated from the Chinese Mangrove Plant Rhizophora mucronata. J. Nat. Prod. 2009, 72, 662–665. [Google Scholar] [CrossRef]
- Sun, J.-F.; Lin, X.; Zhou, X.-F.; Wan, J.; Zhang, T.; Yang, B.; Yang, X.-W.; Tu, Z.; Liu, Y. Pestalols A–E, new alkenyl phenol and benzaldehyde derivatives from endophytic fungus Pestalotiopsis sp. AcBC2 isolated from the Chinese mangrove plant Aegiceras corniculatum. J. Antibiot. 2014, 67, 451–457. [Google Scholar] [CrossRef]
- Mahapatra, S.; Banerjee, D. Production and structural elucidation of exopolysaccharide from endophytic Pestalotiopsis sp. BC55. Int. J. Biol. Macromol. 2016, 82, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xie, J.; Yang, Y.-H.; Li, X.-L.; Zeng, Y.; Zhao, P.-J. Pestalpolyols A–D, cytotoxic polyketides from Pestalotiopsis sp. cr013. Planta Medica 2015, 81, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xie, J.; Yu, F.-X.; Chen, Y.-H.; Zhao, P.-J. Pestalotic acids A-I, antibacterial ambuic acid analogues, from a mycoparasite (Pestalotipsis sp. cr014) of Cronartium ribicola. Arch. Pharmacal Res. 2016, 1–9. [Google Scholar] [CrossRef]
- Xie, J.; Li, J.; Yang, Y.-H.; Li, X.-N.; Chen, Y.-H.; Zhao, P.-J. Four pestalpolyols from a mycoparasite Pestalotipsis sp. PG52. Arch. Pharmacal Res. 2015, 1–6. [Google Scholar] [CrossRef]
- Li, C.-S.; Yang, B.-J.; Turkson, J.; Cao, S. Anti-proliferative ambuic acid derivatives from Hawaiian endophytic fungus Pestalotiopsis sp. FT172. Phytochemistry 2017, 140, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wu, X.; Li, G.; Feng, Z.; Xu, J. Pestalotiopisorin B, a new isocoumarin derivative from the mangrove endophytic fungus Pestalotiopsis sp. HHL101. Nat. Prod. Res. 2020, 34, 1002–1007. [Google Scholar] [CrossRef]
- Zhou, J.; Zheng, D.-Y.; Xu, J. Two new polyketides from endophytic fungus Pestalotiopsis sp. HQD-6 isolated from the Chinese mangrove plant Rhizophora mucronata. J. Asian Nat. Prod. Res. 2021, 24, 52–58. [Google Scholar] [CrossRef]
- Rivera-Chávez, J.; Zacatenco-Abarca, J.; Morales-Jiménez, J.; Martínez-Aviña, B.; Hernández-Ortega, S.; Aguilar-Ramírez, E. Cuautepestalorin, a 7,8-dihydrochromene–oxoisochromane adduct bearing a hexacyclic scaffold from Pestalotiopsis sp. IQ-011. Org. Lett. 2019, 21, 3558–3562. [Google Scholar] [CrossRef]
- Kuang, C.; Jing, S.-X.; Liu, Y.; Luo, S.-H.; Li, S.-H. Drimane sesquiterpenoids and isochromone derivative from the endophytic fungus Pestalotiopsis sp. M-23. Nat. Prod. Bioprospecting 2016, 6, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Klaiklay, S.; Rukachaisirikul, V.; Tadpetch, K.; Sukpondma, Y.; Phongpaichit, S.; Buatong, J.; Sakayaroj, J. Chlorinated chromone and diphenyl ether derivatives from the mangrove-derived fungus Pestalotiopsis sp. PSU-MA69. Tetrahedron 2012, 68, 2299–2305. [Google Scholar] [CrossRef]
- Wu, B.; Wu, X.; Sun, M.; Li, M. Two novel tyrosinase inhibitory sesquiterpenes induced by CuCl2 from a marine-derived fungus Pestalotiopsis sp. Z233. Mar. Drugs 2013, 11, 2713–2721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.Y.; Harper, J.K.; Grant, D.M.; Tombe, B.O.; Bashyal, B.; Hess, W.; Strobel, G.A. Ambuic acid, a highly functionalized cyclohexenone with antifungal activity from Pestalotiopsis spp. and Monochaetia sp. Phytochemistry 2001, 56, 463–468. [Google Scholar] [CrossRef]
- Wang, K.; Lei, J.; Wei, J.; Yao, N. Bioactive Natural compounds from the plant endophytic fungi Pestalotiopsis spp. Mini-Rev. Med. Chem. 2012, 12, 1382–1393. [Google Scholar] [CrossRef]
- Xia, X.; Kim, S.; Liu, C.; Shim, S.H. Secondary metabolites produced by an endophytic fungus Pestalotiopsis sydowiana and their 20S proteasome inhibitory activities. Molecules 2016, 21, 944. [Google Scholar] [CrossRef] [Green Version]
- Uzor, P.F.; Odimegwu, D.C.; Ebrahim, W.; Osadebe, P.O.; Nwodo, N.J.; Okoye, F.B.C.; Liu, Z.; Proksch, P. Anti-respiratory syncytial virus compounds from two endophytic fungi isolated from Nigerian medicinal plants. Drug Res. 2016, 66, 527–531. [Google Scholar] [CrossRef]
- Kimura, Y.; Kouge, A.; Nakamura, K.; Koshino, H.; Uzawa, J.; Fujioka, S.; Kawano, T. Pesthetoxin, a new phytotoxin produced by the gray blight fungus, Pestalotiopsis theae. Biosci. Biotechnol. Biochem. 1998, 62, 1624–1626. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Tian, R.; Liu, S.; Chen, X.; Guo, L.; Che, Y. Pestalotheols A−D, bioactive metabolites from the plant endophytic fungus Pestalotiopsis theae. J. Nat. Prod. 2008, 71, 664–668. [Google Scholar] [CrossRef]
- Ding, G.; Jiang, L.; Guo, L.; Chen, X.; Zhang, H.; Che, Y. Pestalazines and pestalamides, bioactive metabolites from the plant pathogenic fungus Pestalotiopsis theae. J. Nat. Prod. 2008, 71, 1861–1865. [Google Scholar] [CrossRef]
- Liu, L.; Han, Y.; Xiao, J.; Li, L.; Guo, L.; Jiang, X.; Kong, L.; Che, Y. Chlorotheolides A and B, spiroketals generated via Diels–Alder reactions in the endophytic fungus Pestalotiopsis theae. J. Nat. Prod. 2016, 79, 2616–2623. [Google Scholar] [CrossRef]
- Guo, L.; Lin, J.; Niu, S.; Liu, S.; Liu, L. Pestalotiones A–D: Four new secondary metabolites from the plant endophytic fungus Pestalotiopsis theae. Molecules 2020, 25, 470. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Zhang, C.; Gao, Q.; Hou, B.; Liu, L.; Yang, H.; Jiang, X. Chloropupukeananin and pestalofone C regulate autophagy through AMPK and glycolytic Pathway. Chem. Biodivers. 2020, 17, e1900583. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.-F.; Liu, G.-R.; Liu, L. Caryophyllene-type sesquiterpenoids and α-furanones from the plant endophytic fungus Pestalotiopsis theae. Chin. J. Nat. Med. 2020, 18, 261–267. [Google Scholar] [CrossRef]
- Liu, G.; Huo, R.; Zhai, Y.; Liu, L. New bioactive sesquiterpeniods from the plant endophytic fungus Pestalotiopsis theae. Front. Microbiol. 2021, 12, 641504. [Google Scholar] [CrossRef] [PubMed]
- Hou, G.-M.; Xu, X.-M.; Wang, Q.; Li, D.-Y.; Li, Z.-L. Hybrid of dehydroergosterol and nitrogenous alternariol derivative from the fungus Pestalotiopsis uvicola. Steroids 2018, 138, 43–46. [Google Scholar] [CrossRef]
- Bu, S.; Yuan, C.Y.; Xue, Q.; Chen, Y.; Cao, F. Bilobalide suppresses adipogenesis in 3T3-L1 Adipocytes via the AMPK signaling pathway. Molecules 2019, 24, 3503. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.-X.; Kang, J.-C.; Luo, Y.-K.; Zhao, J.-J.; He, J.; Geng, K. A bilobalide-producing endophytic fungus, Pestalotiopsis uvicola from medicinal plant Ginkgo biloba. Curr. Microbiol. 2016, 73, 280–286. [Google Scholar] [CrossRef]
- Zhou, J.-M.; Gu, S.-S.; Mei, W.H.; Zhou, J.; Wang, Z.Z.; Xiao, W. Ginkgolides and bilobalide protect BV2 microglia cells against OGD/reoxygenation injury by inhibiting TLR2/4 signaling pathways. Cell Stress Chaperon. 2016, 21, 1037–1053. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-L.; Huang, L.; Li, H.-Y.; Yang, D.-F.; Li, Z.-Z. Two new compounds from the plant endophytic fungus Pestalotiopsis versicolor. J. Asian Nat. Prod. Res. 2015, 17, 333–337. [Google Scholar] [CrossRef]
- Kesting, J.R.; Olsen, L.; Staerk, D.; Tejesvi, M.V.; Kini, K.R.; Prakash, H.S.; Jaroszewski, J.W. Production of unusual dispiro metabolites in Pestalotiopsis virgatula endophyte cultures: HPLC-SPE-NMR, electronic circular dichroism, and time-dependent density-functional computation study. J. Nat. Prod. 2011, 74, 2206–2215. [Google Scholar] [CrossRef]
- Kesting, J.R.; Staerk, D.; Tejesvi, M.V.; Kini, K.R.; Prakash, H.S.; Jaroszewski, J.W. HPLC-SPE-NMR identification of a novel metabolite containing the benzo[c]oxepin skeleton from the endophytic fungus Pestalotiopsis virgatula culture. Planta Medica 2009, 75, 1104–1106. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, L.; Si, Y.; Jiang, X.; Guo, L.; Che, Y. Virgatolides A–C, Benzannulated spiroketals from the plant endophytic fungus Pestalotiopsis virgatula. Org. Lett. 2011, 13, 2670–2673. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Ding, G.; Li, L.; Cai, X.; Si, Y.; Guo, L.; Che, Y. Isolation, antimicrobial activity, and absolute configuration of the furylidene tetronic acid core of pestalotic acids A–G. Org. Biomol. Chem. 2012, 10, 5307–5314. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, C.; Dong, Y.-J.; Liu, F.-R.; Xu, X.-M.; Li, D.-S.; Li, D.-Y.; Li, Z.-L. Polyketides from Pestalotiopsis zonata and structure revision of pestalrones A and B. J. Asian Nat. Prod. Res. 2019, 21, 134–140. [Google Scholar] [CrossRef]
- Belozerskaya, T.A.; Gessler, N.N.; Aver’yanov, A.A. Melanin pigments in fungi. In Fungal Metabolites; Merillon, J.-M., Ramawat, K.G., Eds.; Springer: Cham, Switzerland, 2017; pp. 263–291. [Google Scholar] [CrossRef]
- Perez-Cuesta, U.; Aparicio-Fernandez, L.; Guruceaga, X.; Martin-Souto, L.; Abad-Diaz-De-Cerio, A.; Antoran, A.; Buldain, I.; Hernando, F.L.; Ramirez-Garcia, A.; Rementeria, A. Melanin and pyomelanin in Aspergillus fumigatus: From its genetics to host interaction. Int. Microbiol. 2020, 23, 55–63. [Google Scholar] [CrossRef]
- Rusman, Y.; Held, B.W.; Blanchette, R.A.; He, Y.; Salomon, C.E. Cadopherone and colomitide polyketides from Cadophora wood-rot fungi associated with historic expedition huts in Antarctica. Phytochemistry 2018, 148, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.-M.; Li, L.-Y.; Sun, L.-Y.; Sun, B.-D.; Niu, S.-B.; Wang, M.-H.; Zhang, X.-Y.; Sun, W.-S.; Zhang, G.-S.; Deng, H.; et al. spiciferone analogs from an endophytic fungus Phoma betae collected from desert plants in West China. J. Antibiot. 2018, 71, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yan, S.; Huang, Z.; Gao, W.; Zhang, S.; Mo, S.; Lin, S.; Wang, J.; Hu, Z.; Zhang, Y. Inducing new bioactive metabolites production from coculture of Pestalotiopsis sp. and Penicillium bialowiezense. Bioorganic Chem. 2021, 110, 104826. [Google Scholar] [CrossRef]
- Guo, Z.; Zou, Z.-M. Discovery of new secondary metabolites by epigenetic regulation and NMR comparison from the plant endophytic fungus Monosporascus eutypoides. Molecules 2020, 25, 4192. [Google Scholar] [CrossRef]
- Shabir, G.; Saeed, A.; El-Seedi, H.R. Natural isocoumarins: Structural styles and biological activities, the revelations carry on. Phytochemistry 2021, 181, 112568. [Google Scholar] [CrossRef]
- Noor, A.O.; Almasri, D.M.; Bagalagel, A.A.; Abdallah, H.M.; Mohamed, S.G.A.; Mohamed, G.A.; Ibrahim, S.R.M. Naturally occurring isocoumarins derivatives from endophytic fungi: Sources, isolation, structural characterization, biosynthesis, and biological activities. Molecules 2020, 25, 395. [Google Scholar] [CrossRef] [Green Version]
- Harwooda, J.S.; Cutler, H.G.; Jacyno, J.M. Nigrosporolide, a plant growth-inhibiting macrolide from the mould Nigrospora sphaerica. Nat. Prod. Lett. 1995, 6, 181–185. [Google Scholar] [CrossRef]
- Li, J.; Kim, S.G.; Blenis, J. Rapamycin: One drug, many effects. Cell Metab. 2014, 19, 373–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vézina, C.; Kudelski, A.; Sehgal, S.N. Rapamycin (AY-22,989), a new antifungal antibiotic. I. Taxonomy of the producing streptomycete and isolation of the active principle. J. Antibiot. 1975, 28, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-Y.-S.; Borgeson, B.M.; Crews, P. Pitholides A-D, polyketides from a marine tunicate-derived culture of Pithomyces sp. Tetrahedron Lett. 1997, 38, 8449–8452. [Google Scholar] [CrossRef]
- Baker, M.A.; Demoret, R.M.; Ohtawa, M.; Shenvi, R.A. Concise asymmetric synthesis of (−)-bilobalide. Nature 2019, 575, 643–646. [Google Scholar] [CrossRef]
- Lu, J.; Xie, L.; Liu, K.; Zhang, X.; Wang, X.; Dai, X.; Liang, Y.; Cao, Y.; Li, X. Bilobalide: A review of its pharmacology, pharmacokinetics, toxicity, and safety. Phytotherapy Res. 2021, 35, 6114–6130. [Google Scholar] [CrossRef]
- Liu, Q.; Jin, Z.; Xu, Z.; Yang, H.; Li, L.; Li, G.; Li, F.; Gu, S.; Zong, S.; Zhou, J.; et al. Antioxidant effects of ginkgolides and bilobalide against cerebral ischemia injury by activating the Akt/Nrf2 pathway in vitro and in vivo. Cell Stress Chaperon. 2019, 24, 441–452. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhai, H. Bilobalide assuages morphine-induced addiction in hippocampal neuron cells through upregulation of microRNA-101. J. Biochem. Mol. Toxicol. 2020, 34, e22493. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, J.; Tong, L.; Gao, T.; Bai, L.; Xue, Q.; Xing, J.; Wang, Q.; Lyu, H.; Cai, M.; et al. Bilobalide protects against ischemia/reperfusion-induced oxidative stress and inflammatory responses via the MAPK/NF-κB pathways in rats. BMC Musculoskelet. Disord. 2020, 21, 449. [Google Scholar] [CrossRef]
- Smetanina, O.F.; Kuznetsova, T.A.; Gerasimenko, A.V.; Kalinovsky, A.I.; Pivkin, M.V.; Dmitrenok, P.C.; Elyakov, G.B. Metabolites of the marine fungus Humicola fuscoatra KMM 4629. Bull. Acad. Sci. USSR Div. Chem. Sci. 2004, 53, 2643–2646. [Google Scholar] [CrossRef]
- De Souza, E.M.; Da Silva, E.L.; Marinho, A.M. (4S)-4,8-dihydroxy-1-tetralone and other chemical constituents from Pestalotiopsis sp. EJC07, endophytic from Bauhinia guianensis. An. Da Acad. Bras. De Ciencias 2016, 88, 29–33. [Google Scholar] [CrossRef]
- Chakraborty, K.; Salas, S. Antioxidant drimane-type sesquiterpenoid from muricid gastropod Chicoreus ramosus attenuates pro-inflammatory 5-lipoxygenase and carbolytic enzymes. J. Food Biochem. 2019, 43, e12991. [Google Scholar] [CrossRef]
- Chen, C.; Sun, W.; Liu, X.; Wei, M.; Liang, Y.; Wang, J.; Zhu, H.; Zhang, Y. Anti-inflammatory spiroaxane and drimane sesquiterpenoids from Talaromyces minioluteus (Penicillium minioluteum). Bioorganic Chem. 2019, 91, 103166. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Zhang, F.-L.; Feng, T. Sesquiterpenoids Specially Produced by Fungi: Structures, Biological Activities, Chemical and Biosynthesis (2015–2020). J. Fungi 2021, 7, 1026. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.-H.; Mei, W.-L.; Zhao, Y.-X.; Zeng, Y.-B.; Wang, H.; Dai, H.-F. A new drimane sesquiterpenoid glycoside from the seeds of Antiaris toxicaria. J. Asian Nat. Prod. Res. 2011, 13, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Edouarzin, E.; Horn, C.; Paudyal, A.; Zhang, C.; Lu, J.; Tong, Z.; Giaever, G.; Nislow, C.; Veerapandian, R.; Hua, D.H.; et al. Broad-spectrum antifungal activities and mechanism of drimane sesquiterpenoids. Microb. Cell 2020, 7, 146–159. [Google Scholar] [CrossRef]
- Neuhaus, G.F.; Loesgen, S. Antibacterial drimane sesquiterpenes from Aspergillus ustus. J. Nat. Prod. 2020, 84, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Paz, C.; Ortiz, L.; Deuis, J.R.; Vetter, I. Polygodial, a drimane sesquiterpenoid dialdehyde purified from Drimys winteri, inhibits voltage-gated sodium channels. Nat. Prod. Res. 2022, 13, 1–6. [Google Scholar] [CrossRef]
- Paz, C.; Viscardi, S.; Iturra, A.; Marin, V.; Miranda, F.; Barra, P.J.; Mendez, I.; Duran, P. Antifungal effects of drimane sesquiterpenoids isolated from Drimys winteri against Gaeumannomyces graminis var. tritici. Appl. Environ. Microbiol. 2020, 86, e01834-20. [Google Scholar] [CrossRef]
- Mahapatra, S.; Banerjee, D. Structural elucidation and bioactivity of a novel exopolysaccharide from endophytic Fusarium solani SD5. Carbohydr. Polym. 2012, 90, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Mao, W.; Li, Y.; Tian, J.; Xu, J. Structural elucidation of the exopolysaccharide produced by fungus Fusarium oxysporum Y24-2. Carbohydr. Res. 2013, 365, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.; Jana, M.; Mahapatra, S. Production of exopolysaccharide by endophytic Stemphylium sp. Micol. Aplicada Int. 2009, 21, 57–62. [Google Scholar]
- Chen, Y.; Mao, W.; Tao, H.; Zhu, W.; Qi, X.; Chen, Y.; Li, H.; Zhao, C.; Yang, Y.; Hou, Y.; et al. Structural characterization and antioxidant properties of an exopolysaccharide produced by the mangrove endophytic fungus Aspergillus sp. Y16. Bioresour. Technol. 2011, 102, 8179–8184. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.M.; Coffey, A.; Arendt, E.K. Exopolysaccharide producing lactic acid bacteria: Their techno-functional role and potential application in gluten-free bread products. Food Res. Int. 2018, 110, 52–61. [Google Scholar] [CrossRef]
- Nguyen, P.-T.; Nguyen, T.-T.; Bui, D.-C.; Hong, P.-T.; Hoang, Q.-K.; Nguyen, H.-T. Exopolysaccharide production by lactic acid bacteria: The manipulation of environmental stresses for industrial applications. AIMS Microbiol. 2020, 6, 451–469. [Google Scholar] [CrossRef]
- Farag, M.M.; Moghannem, S.A.; Shehabeldine, A.; Azab, M.S. Antitumor effect of exopolysaccharide produced by Bacillus mycoides. Microb. Pathog. 2020, 140, 103947. [Google Scholar] [CrossRef]
- Asgher, M.; Urooj, Y.; Qamar, S.A.; Khalid, N. Improved exopolysaccharide production from Bacillus licheniformis MS3: Optimization and structural/functional characterization. Int. J. Biol. Macromol. 2019, 151, 984–992. [Google Scholar] [CrossRef]
- Ren, Q.; Tang, Y.; Zhang, L.; Xu, Y.; Liu, N.; Ren, H. Exopolysaccharide produced by Lactobacillus casei promotes the differentiation of CD4+ T cells into Th17 cells in BALB/c mouse Peyer’s patches in vivo and in vitro. J. Agric. Food Chem. 2020, 68, 2664–2672. [Google Scholar] [CrossRef]
- Zhou, M.; Li, Z.; Liu, Y.; Zhang, P.; Hao, X.; Zhu, X. Transcription factors Pmr1 and Pmr2 cooperatively regulate melanin biosynthesis, conidia development and secondary metabolism in Pestalotiopsis microspora. J. Fungi 2021, 8, 38. [Google Scholar] [CrossRef]
- Zhang, Q.; Akhberdi, O.; Wei, D.; Chen, L.; Liu, H.; Wang, D.; Hao, X.; Zhu, X. A MYST Histone Acetyltransferase modulates Conidia development and secondary metabolism in Pestalotiopsis microspora, a taxol producer. Sci. Rep. 2018, 8, 8199. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Xu, Y.; Maine, E.A.; Wijeratne, E.K.; Espinosa-Artiles, P.; Gunatilaka, A.L.; Molnár, I. Functional characterization of the biosynthesis of radicicol, an Hsp90 inhibitor resorcylic acid lactone from Chaetomium chiversii. Chem. Biol. 2008, 15, 1328–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Li, Y.; Wang, H.; Wei, D.; Akhberdi, O.; Liu, Y.; Xiang, B.; Hao, X.; Zhu, X. The AMP-activated protein kinase homolog Snf1 concerts carbon utilization, conidia production and the biosynthesis of secondary metabolites in the taxol-producer Pestalotiopsis microspora. Genes 2018, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhang, P.; Zhou, H.; Liu, X.; Li, S.; Guo, L.; Li, K.; Yin, W. A new regulator RsdA mediating fungal secondary metabolism has a detrimental impact on asexual development in Pestalotiopsis fici. Environ. Microbiol. 2019, 21, 416–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza, W.R.; Morais, E.R.; Krohn, N.G.; Savoldi, M.; Goldman, M.H.S.; Rodrigues, F.; Caldana, C.; Semelka, C.T.; Tikunov, A.P.; Macdonald, J.M.; et al. Identification of metabolic pathways influenced by the G-protein coupled receptors GprB and GprD in Aspergillus nidulans. PLoS ONE 2013, 8, e62088. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhong, K.; Yin, Z.; Hu, J.; Wang, W.; Li, L.; Zhang, H.; Zheng, X.; Wang, P.; Zhang, Z. The seven transmembrane domain protein MoRgs7 functions in surface perception and undergoes coronin MoCrn1-dependent endocytosis in complex with Gα subunit MoMagA to promote cAMP signaling and appressorium formation in Magnaporthe oryzae. PLOS Pathog. 2019, 15, e1007382. [Google Scholar] [CrossRef] [Green Version]
- Pang, X.-M.; Tian, D.; Zhang, T.; Liao, L.-S.; Li, C.-X.; Luo, X.-M.; Feng, J.-X.; Zhao, S. G protein γ subunit modulates expression of plant-biomass-degrading enzyme genes and mycelial-development-related genes in Penicillium oxalicum. Appl. Microbiol. Biotechnol. 2021, 105, 4675–4691. [Google Scholar] [CrossRef]
- Ramanujam, R.; Calvert, M.E.; Selvaraj, P.; Naqvi, N.I. The late endosomal HOPS complex anchors active G-protein signaling essential for pathogenesis in Magnaporthe oryzae. PLOS Pathog. 2013, 9, e1003527. [Google Scholar] [CrossRef] [Green Version]
- Tzima, A.K.; Paplomatas, E.J.; Tsitsigiannis, D.I.; Kang, S. The G protein β subunit controls virulence and multiple growth- and development-related traits in Verticillium dahliae. Fungal Genet. Biol. 2012, 49, 271–283. [Google Scholar] [CrossRef]
- Yan, H.; Zhou, Z.; Shim, W.B. Two regulators of G-protein signaling (RGS) proteins FlbA1 and FlbA2 differentially regulate fumonisin B1 biosynthesis in Fusarium verticillioides. Curr. Genet. 2021, 67, 305–315. [Google Scholar] [CrossRef]
- Chen, L.; Li, Y.; Zhang, Q.; Akhberdi, O.; Wei, D.; Pan, J.; Zhu, X. Seamless deletion of a large DNA fragment in the taxol-producing fungus Pestalotiopsis microspora. Mycoscience 2017, 58, 35–39. [Google Scholar] [CrossRef]
- Chen, L.; Wei, D.; Zhang, Q.; Yu, X.; Wang, Y.; Zhu, X. Orotidine 5′-phosphate decarboxylase-based reusable in situ genetic editing system: Development and application in taxol-producing Pestalotiopsis microspora. Eng. Life Sci. 2015, 15, 542–549. [Google Scholar] [CrossRef]
- Xu, X.; Huang, R.; Yin, W.-B. An optimized and efficient CRISPR/Cas9 system for the endophytic fungus Pestalotiopsis fici. J. Fungi 2021, 7, 809. [Google Scholar] [CrossRef] [PubMed]
- Subban, K.; Subramani, R.; Srinivasan, V.P.M.; Johnpaul, M.; Chelliah, J. Salicylic acid as an effective elicitor for improved taxol production in endophytic fungus Pestalotiopsis microspora. PLoS ONE 2019, 14, e0212736. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.S.; Shindia, A.A.; AbouZeid, A.; Koura, A.; Hassanein, S.E.; Ahmed, R.M. Triggering the biosynthetic machinery of taxol by Aspergillus flavipes via cocultivation with Bacillus subtilis: Proteomic analyses emphasize the chromatin remodeling upon fungal-bacterial interaction. Environ. Sci. Pollut. Res. 2021, 28, 39866–39881. [Google Scholar] [CrossRef]
- Nützmann, H.-W.; Fischer, J.; Scherlach, K.; Hertweck, C.; Brakhage, A.A. Distinct amino acids of histone H3 control secondary metabolism in Aspergillus nidulans. Appl. Environ. Microbiol. 2013, 79, 6102–6109. [Google Scholar] [CrossRef] [Green Version]
- Nützmann, H.-W.; Reyes-Dominguez, Y.; Scherlach, K.; Schroeckh, V.; Horn, F.; Gacek, A.; Schümann, J.; Hertweck, C.; Strauss, J.; Brakhage, A.A. Bacteria-induced natural product formation in the fungus Aspergillus nidulans requires Saga/Ada-mediated histone acetylation. Proc. Natl. Acad. Sci. USA 2011, 108, 14282–14287. [Google Scholar] [CrossRef] [Green Version]
- Schroeckh, V.; Scherlach, K.; Nützmann, H.-W.; Shelest, E.; Schmidt-Heck, W.; Schuemann, J.; Martin, K.; Hertweck, C.; Brakhage, A.A. Intimate bacterial–fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 2009, 106, 14558–14563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, J.; Müller, S.Y.; Netzker, T.; Jäger, N.; Gacek-Matthews, A.; Scherlach, K.; Stroe, M.C.; García-Altares, M.; Pezzini, F.; Schoeler, H.; et al. Chromatin mapping identifies BasR, a key regulator of bacteria-triggered production of fungal secondary metabolites. eLife 2018, 7, e40969. [Google Scholar] [CrossRef]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites—Strategies to activate silent gene clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
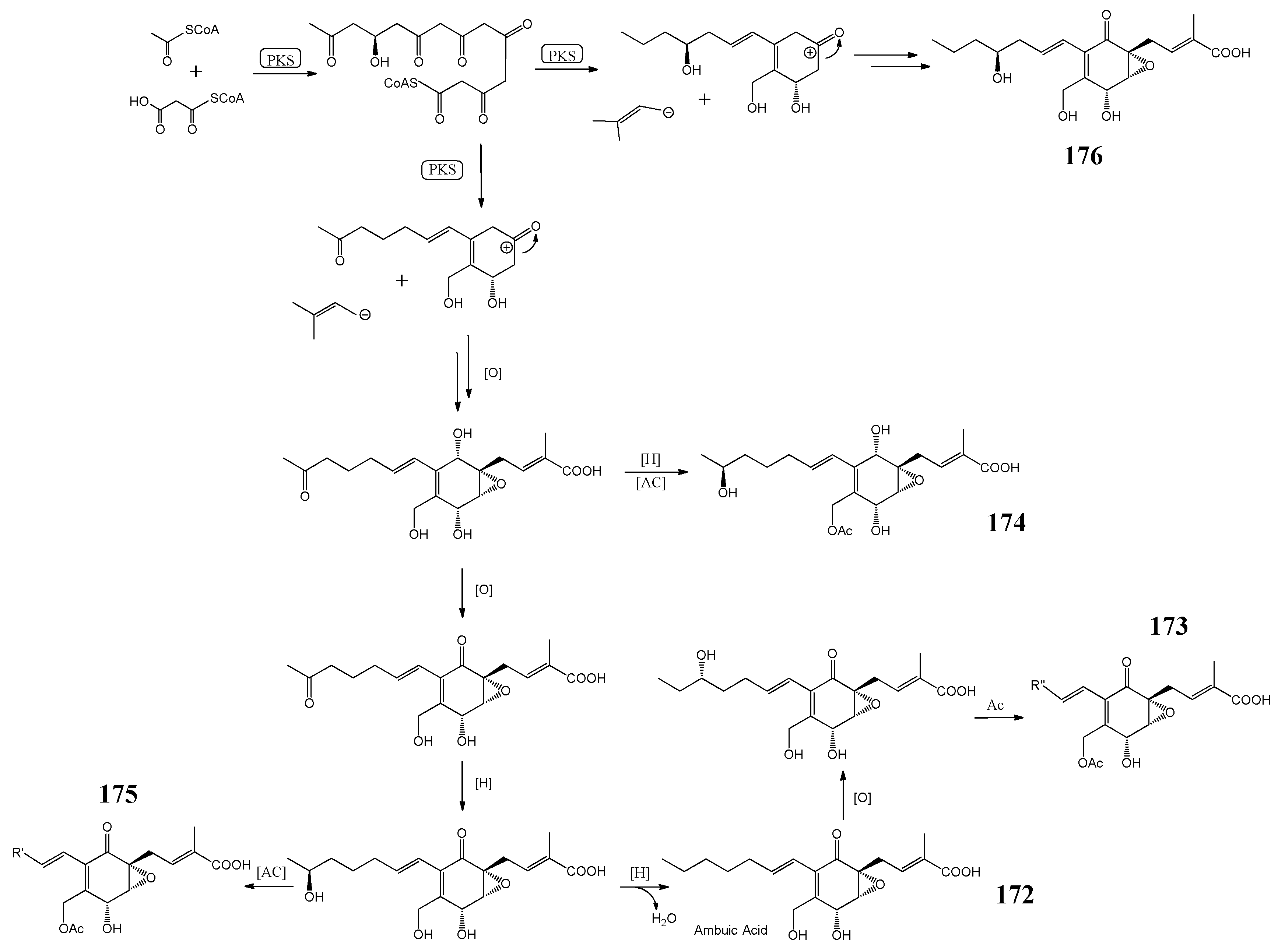{kind=link}
| Fungal Species | Metabolites | Bioactivity | References |
|---|---|---|---|
| P. adusta | pestalachlorides A–C | antifungal | [69] |
| diterpenoid | [70] | ||
| P. besseyi | furanones | [71] | |
| P. breviseta | taxol | anticancer | [16,17] |
| P. crassiuscula | a new coumarin and six known compounds | [72] | |
| P. diploclisia | pestalotioquinols G and H, pestalotioquinol A, phomonitroester, (R)-4,6,8-trihydroxy-3,4-dihydronaphthalen-1(2H)-one, and scylatone | antimalarial and cytotoxic activity | [73] |
| P. disseminata | 6-hydroxypunctaporonin E, 6-hydroxypunctaporonin B, and 6-hydroxypunctaporonin A | anti-bacteria | [74] |
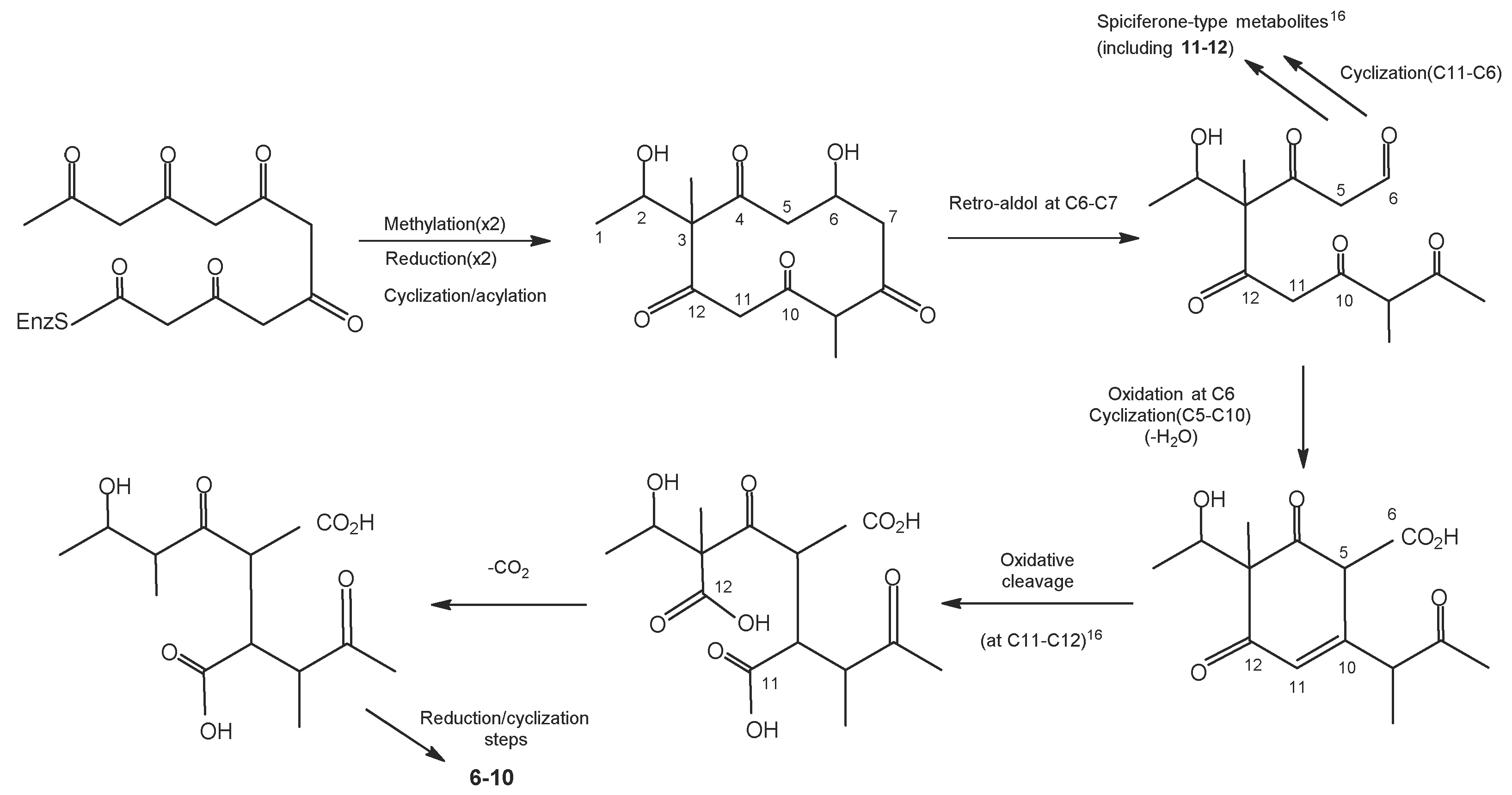
| disseminins A–E, spiciferones D and E | [75] | ||
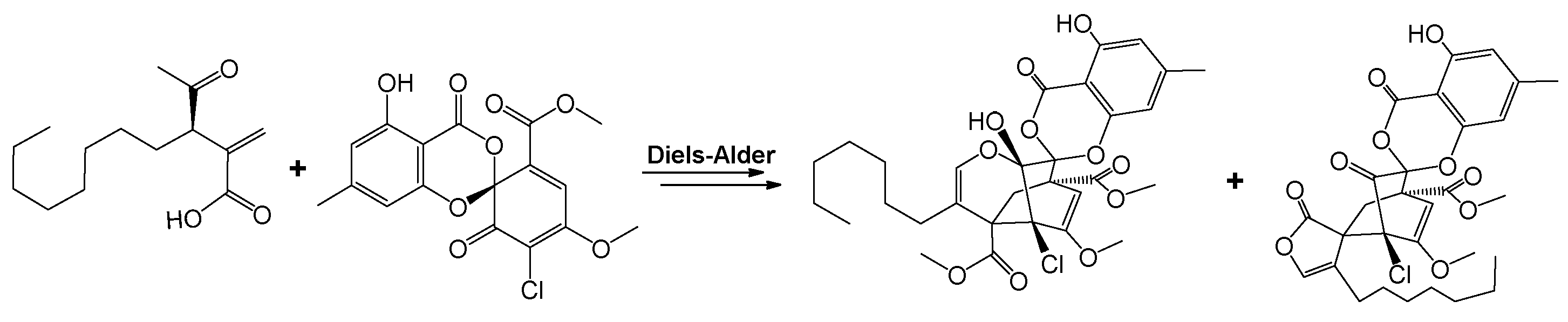
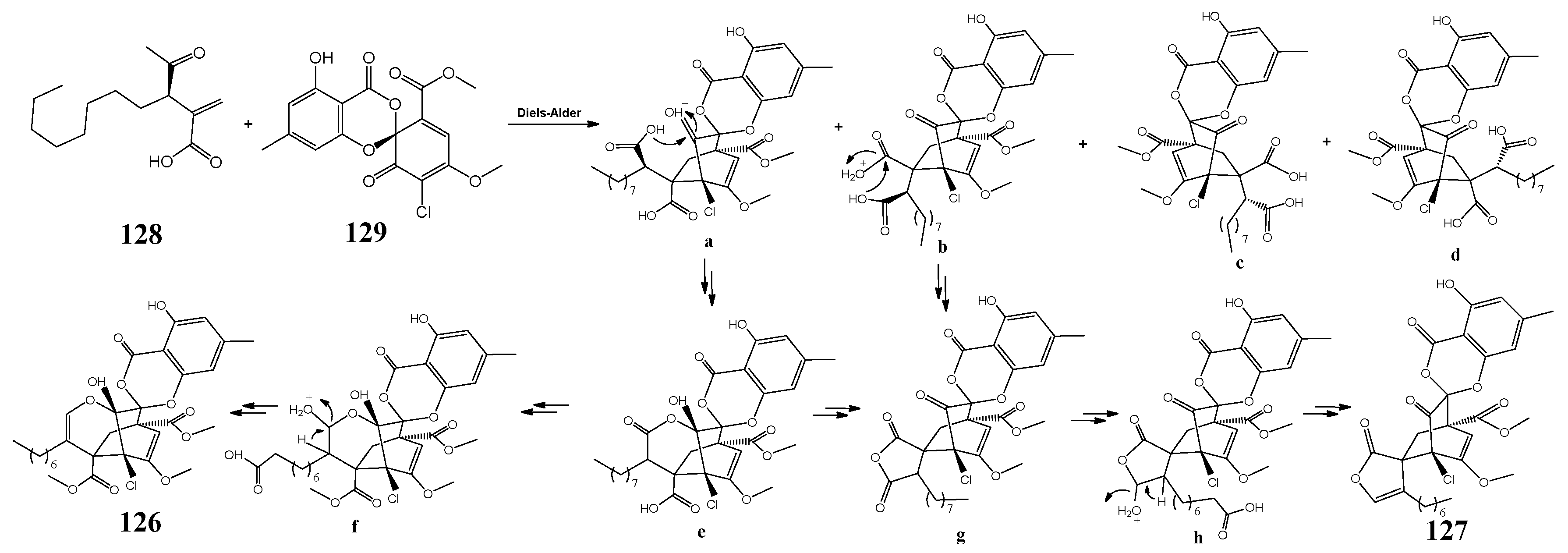
| P. fici | chloropupukeananin and chloropestolides | antimicrobial, antitumor, and anti-HIV activities | [76] |
| pestaloficiols A–E | inhibitory effects on HIV-1 replication | [77] | |
| isosulochrin, ficipyrone A, pestheic acid, iso-A82775C, pestaloficiol M, RES1214-1, and iso-A82775C | [78] | ||
| pestalofones A–E, isosulochrin, isosulochrin dehydrate, and iso-A82775C | inhibitory effects on HIV-1 replication and antifungal activity | [79] | |
| chloropestolide A | antitumor | [80] | |
| chloropupukeanolides C–E | cytotoxicity | [81] | |
| pestheic acid | [82] | ||
| chloropupukeananin | [83] | ||
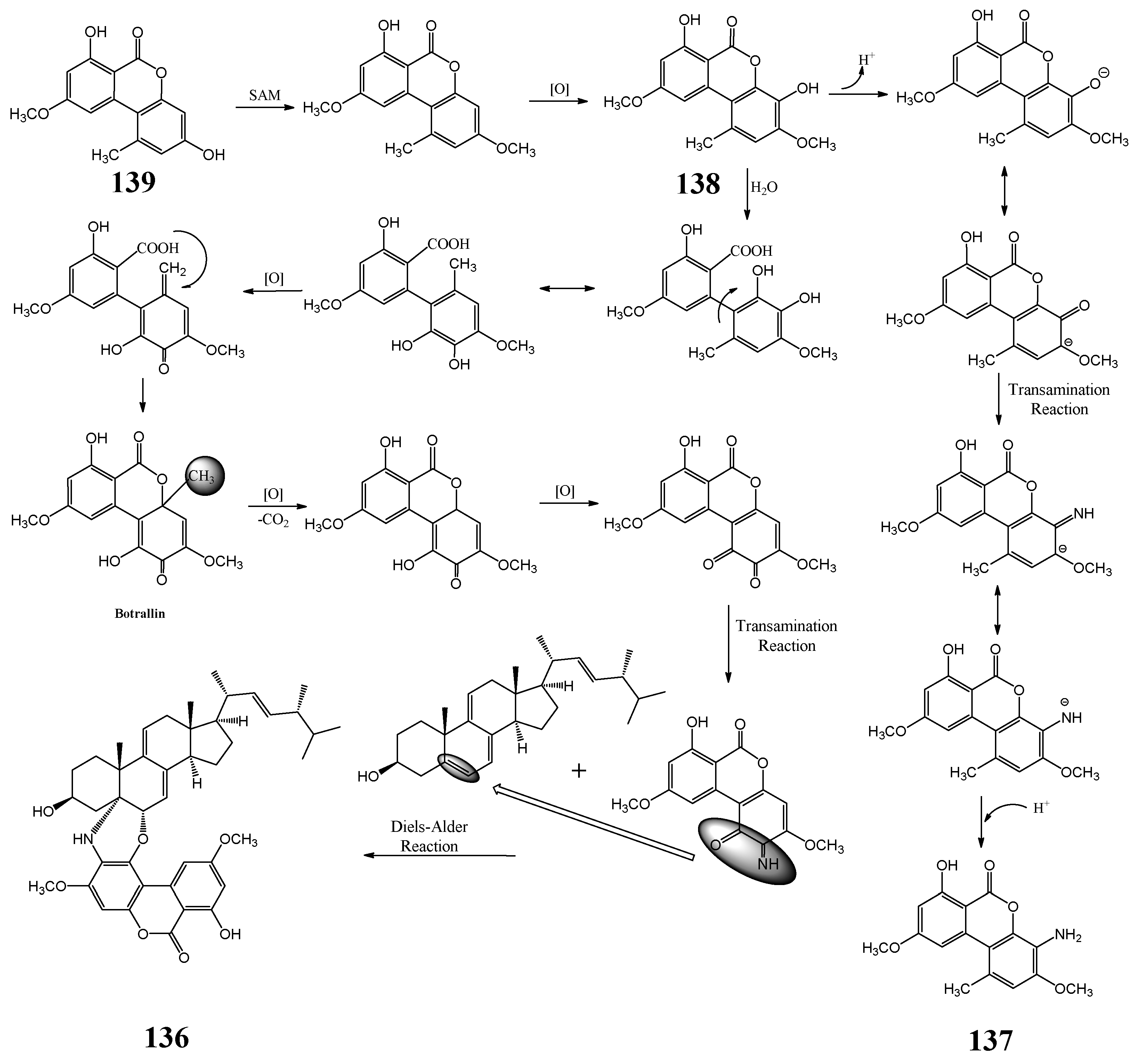
| melanin | [84] | ||
| chloropupukeanolides A and B, chloropupukeanone A | anti-HIV-1 and cytotoxic activity | [85] | |
| isoprenylated chromone derivatives | [86] | ||
| Chloropestolides B–G | [87] | ||
| pestaloficins A–E | [88] | ||
| pestaloficiols Q–S | [89] | ||
| DHN melanin | [90] | ||
| 2H-pyran-2-one and 2H-furan-2-one derivatives | [91] | ||
| P. flavidula | spiroketal derivatives | cytotoxicity | [92] |
| P. foedan | pestafolide A, pestaphthalide A and B | antifungal activity | [93] |
| (−)-(4S, 8S)-foedanolide and (+)-(4R, 8R)-foedanolide | [94] | ||
| monoterpene derivatives | antifungal | [95] | |
| P. guepinii | metabolites of ciprofloxacin and norfloxacin | [96] | |
| culture broth extract | inhibit actinomycete growth | [97] | |
| pestheic acid or dihidromaldoxin | genotoxicity and mutagenicity | [98] | |
| alpha-pyrones | [99] | ||
| P. hainanensis | taxol | [19] | |
| caryophyllene-Type Sesquiterpenes | [100] | ||
| P. heterocorni | 7-hydroxy-5-methoxy-4,6-dimethyl-7-O-α-L-rhamnosyl-phthalide and 7-hydroxy-5-methoxy-4,6-dimethyl-7-O-β-D-glucopyranosyl-phthalide | [101] | |
| heterocornols A–L, methyl-(2-formyl-3-hydroxyphenyl) propanoate, cladoacetal A, xylarinol A, agropyrenol, vaccinol G, (R)-3-hydroxy-1-[(R)-4-hydroxy-1,3-dihydroisobenzofuran-1-yl]butan-2-one, and (R)-3-hydroxy-1-[(S)-4-hydroxy-1,3-dihydroisobenzo -furan-1-yl]butan-2-one | cytotoxicity and antifungal | [29] | |
| pestaloisocoumarins A and B, isopolisin B, pestalotiol A, gamahorin, pestalachloride B, pestalachloride E, pestalalactone atropisomers (8a/8b), | cytotoxicity | [30] | |
| heterocornols M and N, heterocornols O and P | anticancer | [28] | |
| P. humus | pestynol | antibacterial and antifungal activity, weak cytotoxicity | [102] |
| pestiocandin | antibacterial and antifungal activity | [103] | |
| P. jesteri | jesterone and hydroxy-jesterone | selective antimycotic activity | [104] |
| P. karstenii | pestalrone A, pestalrone B, pestalotin, hydroxypestalotin | cytotoxic activity, antiprotozoal activity | [42] |
| P. leucothës | BS, GS, and YS | Immunomodulatory | [105] |
| immunomodulatory effects | [106] | ||
| P. mangiferae | 4-(2,4,7-trioxa-bicyclo [4.1.0] heptane-3-yl) phenol | antibacterial and antifungal activity | [107] |
| P. microspora | taxol | antiproliferative activity | [11,108] |
| ɑ-pyrone | [109] | ||
| (+)-dendocarbin L, (+)-sydonic acid, and (+)-sydowic acid | cytotoxicity | [68] | |
| isopestacin | antifungal and antioxidant activities | [110] | |
| pestalotiollide B | [49,111,112,113] | ||
| pestalotiollide B, melanin | [114] | ||
| 7-epi-10-deacetyltaxol | induces apoptosis | [115] | |
| taxol, pestalotiollide B, 1, 8-dihydroxy naphthalene melanin | antitumor | [13] | |
| melanin | [116] | ||
| ursolic acid | [117] | ||
| pestalotioprolides C, D–H, 7-O-methylnigrosporolide, pestalotioprolide B, seiricuprolide, nigrosporolide, and 4,7-dihydroxy-13-tetradeca-2,5,8-trienolide | cytotoxic | [118] | |
| dibenzodioxocinons | inhibitors of cholesterol ester transfer protein | [119,120] | |
| pestalotioquinols A and B | neuroprotective | [121] | |
| pitholide E, pitholide B, pitholide D, pestalotin, PC-2, tyrosol, 4-oxo-4H-pyran-3-acetic acid | [67] | ||
| 2H-pyranone and isocoumarin derivatives | antifungal | [66] | |
| Microsporols A–C, ambuic acid | 5-lipoxygenase (5-LOX) inhibitory effects | [122] | |
| P. neglecta | ambuic acid derivatives | inhibitory activity against the NO production | [123] |
| crude methanol and ethyl acetate extract | antibacterial activity | [124] | |
| pestalotiochromenoic acids A–D, pestalotiochromones A and B | liver X receptors modulators | [125] | |
| ambuic acid | anti-inflammatory action | [126] | |
| benzophenones | inhibit pancreatic cancer cells | [31] | |
| Ene-yne Hydroquinones | [26] | ||
| pestalopyrones A–D | [127] | ||
| pestallic acids F and G, pestalotiopyrone N, neopestalone, sesquicaranoic acid B, monocycloalternarene B, pestalone, 2,4-dihydroxy-3,5,6-trimethyl benzoic acid, and citreorosein | [27] | ||
| P. palmarump | sinopestalotiollides A–D, 3′-O-methyldehydroisopenicillide, ∆1′3′-1′-dehydroxypenicillide, dehydroisopenicillide, 2′-hydroxy-3′,4′-didehydropenicillide, scirpyrones A, 5,6-dihydro-4-methoxy-2H-pyran-2-one, LL-P880α, (6S,1′S,2′R)-LL-P880b, photipyrone B, PC-2, (1′S,2′R)-LL-P880γ, necpyrone C | cytotoxic | [128] |
| P. pauciseta | taxol | anticancer | [15] |
| P. photiniae | phthalide derivatives | against plant pathogens | [129] |
| photipyrones A, B, C | modest inhibitory effects on the growth of MDA-MB-231 | [130] | |
| 4-(3′,3′-dimethylallyloxy)-5-methyl-6-methoxyphthalide | induced G1 cell cycle arrest and apoptosis in a dose-dependent manner | [131] | |
| 4-(3′,3′-Dimethylallyloxy)-5-methyl-6-methoxy-phthalide | cytotoxicity | [132] | |
| three new phthalide derivatives and six known phthalide derivatives | antifungal activities | [133] | |
| photinides A–F | cytotoxic | [52] | |
| Pestalotiopsis sp. | RES-1214-1 and -2 | non-peptidic endothelin type A receptor antagonists | [134] |
| Pestalotiopsis sp. | pestalotiopamide E | [135] | |
| Pestalotiopsis sp. | pestalotiopens A and B | [136] | |
| Pestalotiopsis sp. | pestalactams A–C | [137] | |
| Pestalotiopsis sp. | pestaloficiol J, (±)-pestaloficiol X | [138] | |
| Pestalotiopsis sp. | pestaloquinols A and B | [139] | |
| Pestalotiopsis sp. | demethylincisterol A3, dankasterone B, (22E, 24R)-ergosta-7,9, 22-triene-3β, 5α, 6α-triol, ergosta-5,7,22-trien-3-ol, 5,8-epidioxy-5,8-ergosta-6,22E-dien-3-ol, stigmastan-3-one, stigmast-4-en-3-one, stigmast-4-en-6-ol-3-one, flufuran, (2-cis, 4-trans)-abscisic acid, similanpyrone B. | induce G0/G1 cell cycle arrest and apoptosis in human cancer cells | [140] |
| Pestalotiopsis sp. | pestaloporonins | [141] | |
| Pestalotiopsis sp. | polyketide-terpene hybrid metabolites | [142] | |
| Pestalotiopsis sp. | pestaloporinates A–G and 14-acetylhumulane | [143] | |
| Pestalotiopsis sp. | isocoumarin derivatives | antifungal | [144] |
| Pestalotiopsis sp. | pestalotiopsolide A, taedolidol and 6-epitaedolidol | [41] | |
| Pestalotiopsis sp. | (±)-pestalachloride D | antibacterial | [145] |
| Pestalotiopsis sp. | cytosporones J-N, pestalasins A-E, pestalotiopsoid A, cyclosporine C, dothiorelone B, and 3-hydroxymethyl-6,8-dimethoxycoumarin (13). | [146] | |
| Pestalotiopsis sp. | (+)- and (−)-pestaloxazine A | antiviral | [147] |
| Pestalotiopsis sp. | pestalotiopsins A and B | immunosuppressive | [148] |
| Pestalotiopsis sp. | ambuic acid and torreyanic acid derivatives | antimicrobial activity | [149] |
| Pestalotiopsis sp. | pestalotiopsones A–F | moderate cytotoxicity | [150] |
| Pestalotiopsis sp. AcBC2 | pestalols A–E, 4-hydroxyphenethyl 2-(4-hydroxyphenyl) acetate, r-hydroxyphenyl acetic acid methyl ester, transharzialactones A and F, 3-hydroxy-3-methyl-d-lactone, 3β,5α, 9α-trihydroxy-7, 22-en-ergost-6-one, and 3b-hydroxy-sterol | cytotoxicity, inhibitory activities against Influenza A virus subtype (H3N2), Swine Flu (H1N1) viruses, tuberculosis | [151] |
| Pestalotiopsis sp. BC55 | exopolysaccharide | [152] | |
| Pestalotiopsis sp. cr013 | pestalpolyols A–D | cytotoxic | [153] |
| Pestalotiopsis sp. cr014 | pestalotic acids A–I | antibacterial | [154] |
| Pestalotiopsis sp. PG52 | pestalpolyols E–H | cytotoxic activities | [155] |
| Pestalotiopsis sp. EJC07 | (4S)-4,8-dihydroxy-1-tetralone, uracil, uridine, p-hydroxybenzoic acid, ergosterol, ergosterol peroxide, cerevisterol and ducitol | [74] | |
| Pestalotiopsis sp. FT172 | pestallic acids A–E | anti-proliferative | [156] |
| Pestalotiopsis sp. HC02 | pestalotines A and B | [38] | |
| Pestalotiopsis sp. HHL101 | pestalotiopisorin B | [157] | |
| Pestalotiopsis sp. HQD-6 | pestalotiopin B and pestalotiopyrone N | very weak cytotoxic | [158] |
| Pestalotiopsis sp. IQ-011 | cuautepestalorin, cytosporin M, cytosporin N, oxopestalochromane, pestalone | inhibitory properties against α-glucosidase from S. cerevisiae | [159] |
| Pestalotiopsis sp. M-23 | drimane sesquiterpenoids, 2α-hydroxyisodrimeninol, a new isochromone derivative | weak antibacterial | [160] |
| Pestalotiopsis sp. PSU-MA69 | pestalochromones A–C, pestalotethers A–D, pestaloxanthone, pestalolide | antifungal activity against Candida albicans and Cryptococcus neoformans | [161] |
| Pestalotiopsis sp. Z233 | 1β,5α,6α,14-tetraacetoxy-9α-benzoyloxy-7β H-eudesman-2β,11-diol and 4α,5α-diacetoxy-9α-benzoyloxy-7βH-eudesman-1β,2β,11,14-tetraol | tyrosinase inhibitory activities | [162] |
| Pestalotiopsis spp. | ambuic acid | antifungal | [163] |
| Pestalotiopsis spp. | chromones, cytosporones, polyketides, terpenoids and coumarins | [164] | |
| P. sydowiana | 1-O-methyldehydroisopenicillide, pestalotiollide B, pestalotiollide A, dehydroisopenicillide, 6-hydroxymethyl-4-methoxy-5,6-dihydro-2H-pyran-2-one, pestalotiopyrone D, pestalotiopyrone E, pestalotiopyrone G, LL-P880b, and photipyrone B | 20S proteasome inhibitory activities | [165] |
| cyclo(-Leu-Pro) and 4-hydroxyphenylacetamide | antimicrobial, | [33] | |
| P. terminaliae | taxol | anticancer | [18] |
| P. theae | chloroisosulochrin, ficipyrone A and pestheic acid | strong activity against respiratory syncytial virus | [166] |
| pesthetoxin | [167] | ||
| pestalotheols A–D | inhibitory effect on HIV-1 LAI replication | [168] | |
| pestalazines and pestalamides | inhibitory effects on HIV-1 replication and antifungal activity | [169] | |
| chlorotheolides A and B, 1-undecene-2,3-dicarboxylic acid, maldoxin | [170] | ||
| pestalotiones A–D | [171] | ||
| chloropupukeananin and pestalofone C | regulate autophagy through AMPK and Glycolytic Pathway | [172] | |
| pestathenols A and B, pestatheranone A, punctaporonins O, P, and R, ficifuranone B, and decarestrictine D | [173] | ||
| pestalothenin A–C | cytotoxic and antibacterial activity | [174] | |
| P. uvicola | pestauvicomorpholine A and three alternariol analogues | cytotoxicity against mouse melanoma (B16-BL6) cell line | [175] |
| bilobalide | suppresses adipogenesis in 3T3-L1 adipocytes via the AMPK signaling pathway; protect BV2 microglia cells against OGD/reoxygenation injury | [176,177,178] | |
| P. vaccinii | vaccinols J-S, trans-sordarial, trans-sordariol, cis-sordariol, 4-hydroxyphthalide, pestalotiopin A | [32] | |
| P. versicolor | 4,6-dihydroxy-7-formyl-3-methyl coumarin, 6-[(7S,8R)-8-propyloxiran-1-yl]-4-methoxy-pyran-2-one | devoid of significant antifungal activity against F. solani, Ustilago maydis, and C. albicans | [179] |
| P. virgatula | pestalospiranes A and B | [180] | |
| 9-hydroxybenzo[C]oxepin-3[1H]-one | [181] | ||
| virgatolides A–C | [182] | ||
| P. yunnanensis | pestalotic acids A–G | significant antimicrobial activity | [183] |
| P. zonata | pestalrones A and B, pestalotin, hydroxypestalotin, pestazonatic acid, necpyrones A–B | [184] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Wang, Y.; Yang, Y. Pestalotiopsis Diversity: Species, Dispositions, Secondary Metabolites, and Bioactivities. Molecules 2022, 27, 8088. https://doi.org/10.3390/molecules27228088
Wu C, Wang Y, Yang Y. Pestalotiopsis Diversity: Species, Dispositions, Secondary Metabolites, and Bioactivities. Molecules. 2022; 27(22):8088. https://doi.org/10.3390/molecules27228088
Chicago/Turabian StyleWu, Chu, Yun Wang, and Yujie Yang. 2022. "Pestalotiopsis Diversity: Species, Dispositions, Secondary Metabolites, and Bioactivities" Molecules 27, no. 22: 8088. https://doi.org/10.3390/molecules27228088