
Analysis of Anti-Cancer and Anti-Inflammatory Properties of 25 High-THC Cannabis Extracts

Abstract
:1. Introduction
2. Results
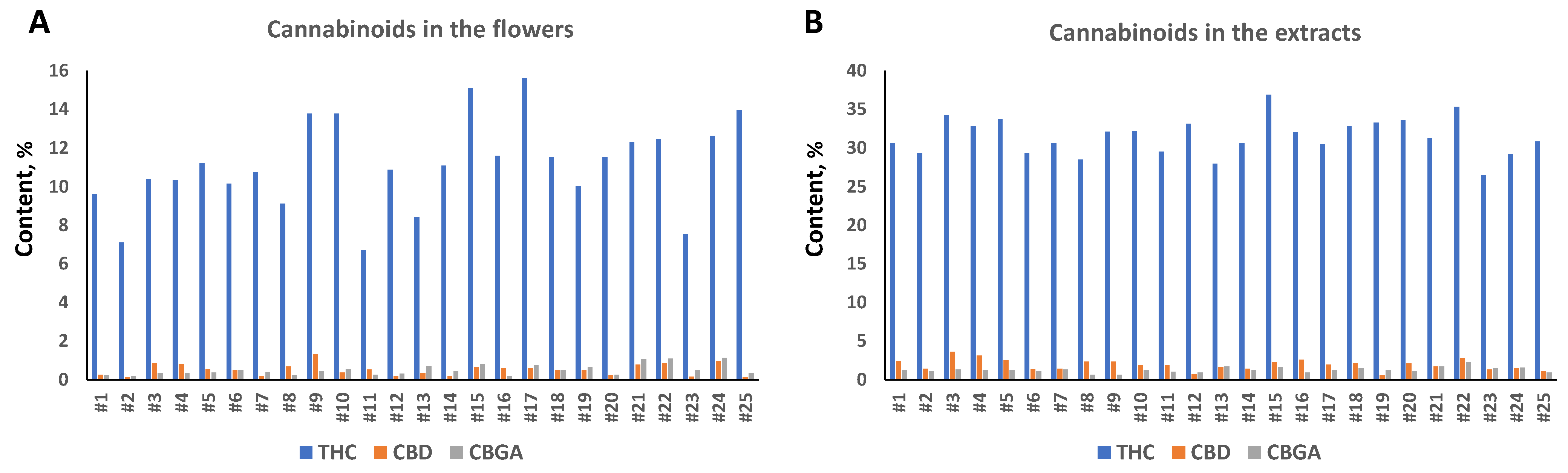
2.1. Analysis of Cannabinoids Content in 25 Cannabis Varieties
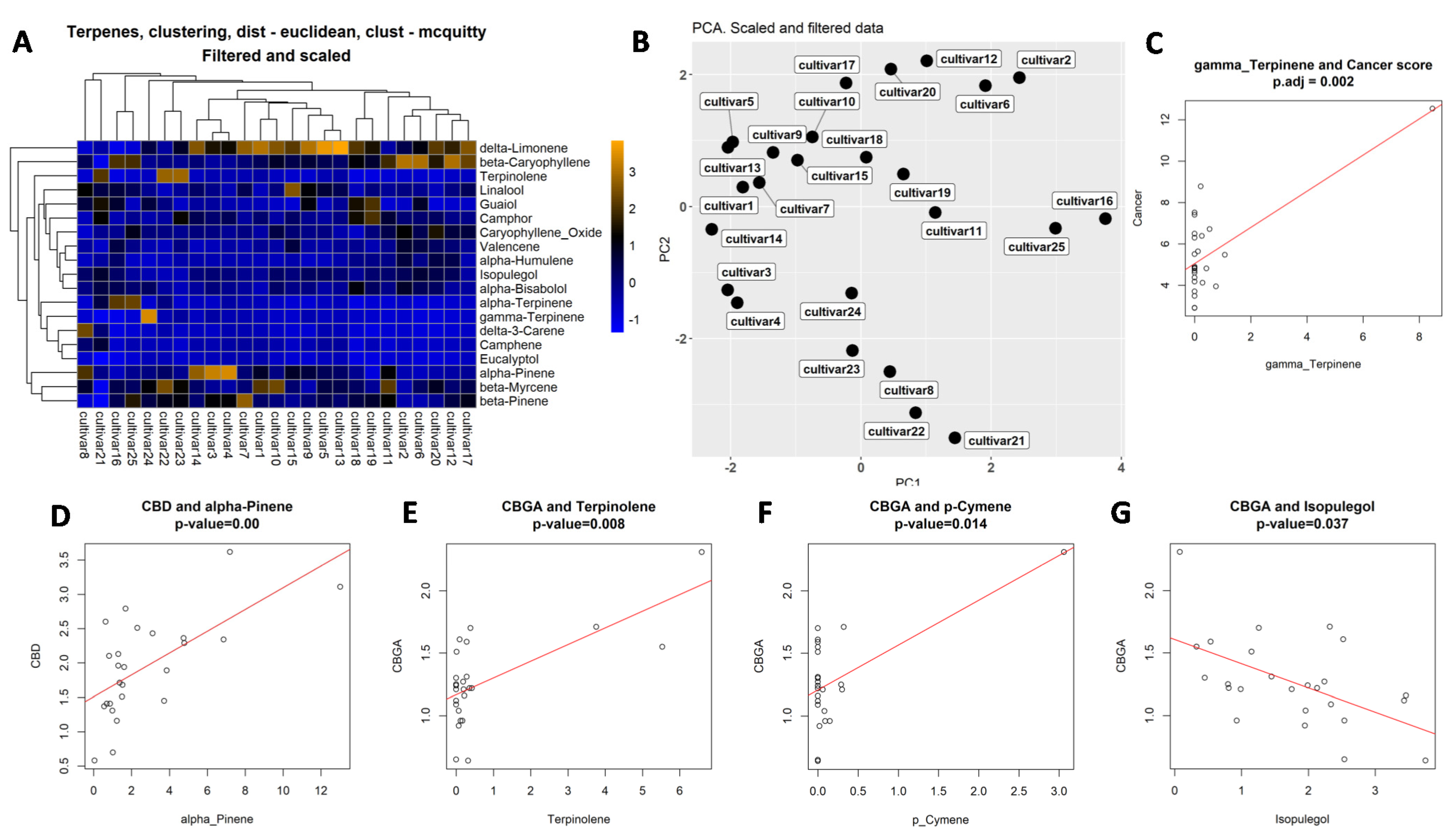
2.2. Analysis of Terpene Content in 25 Cannabis Varieties
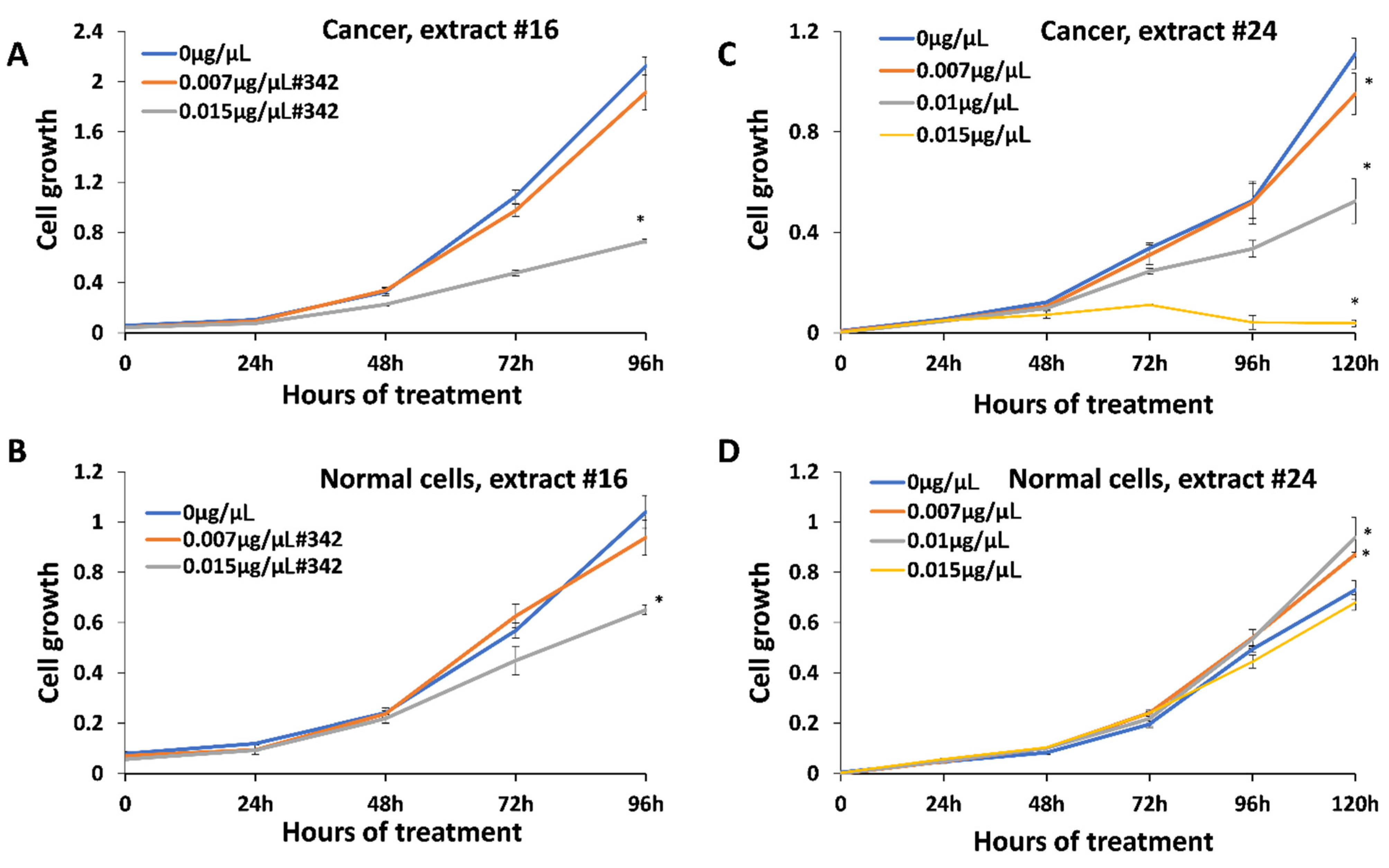
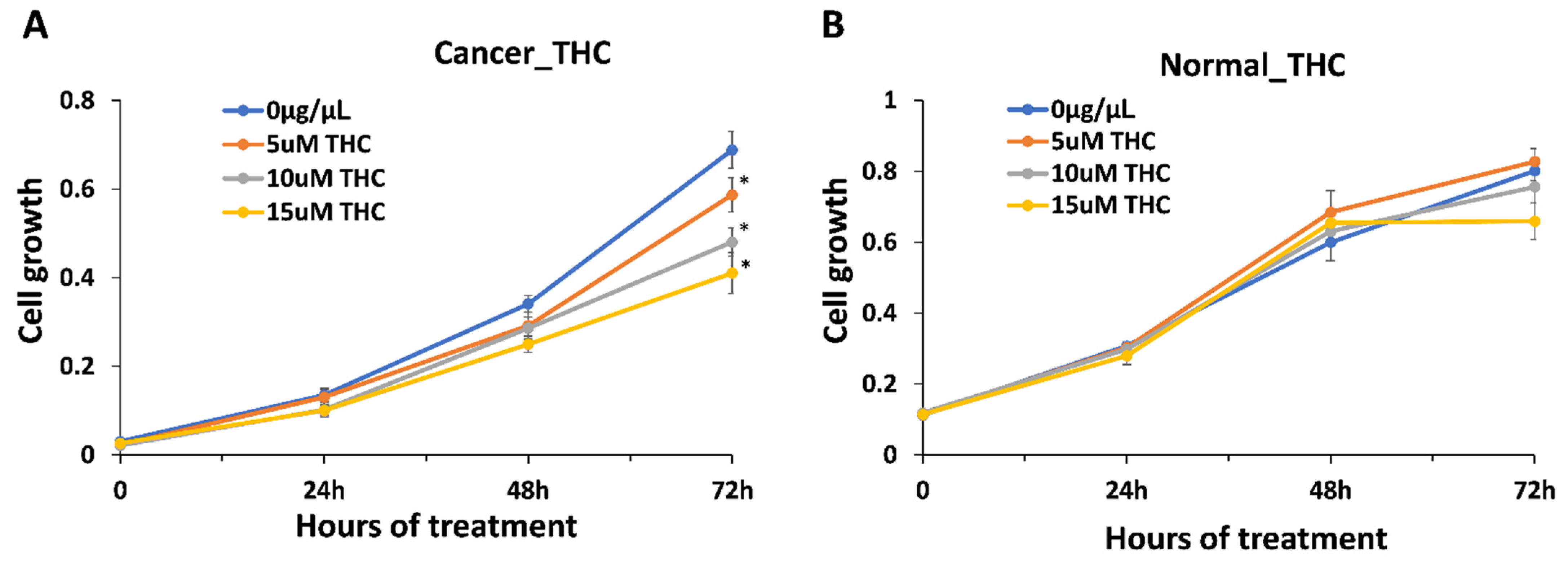
2.3. Inhibitory Effect of Extracts on Cancer Cells
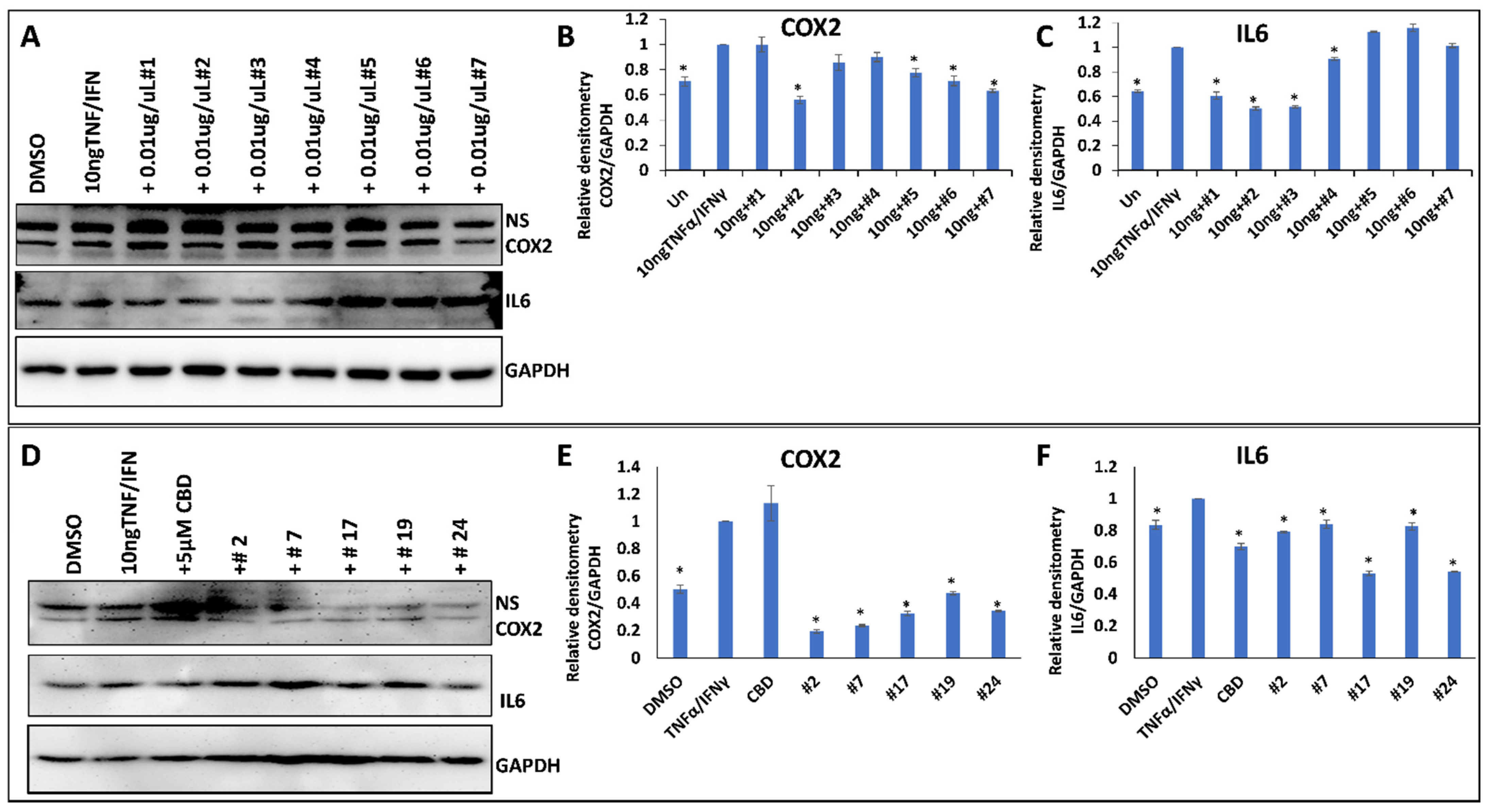
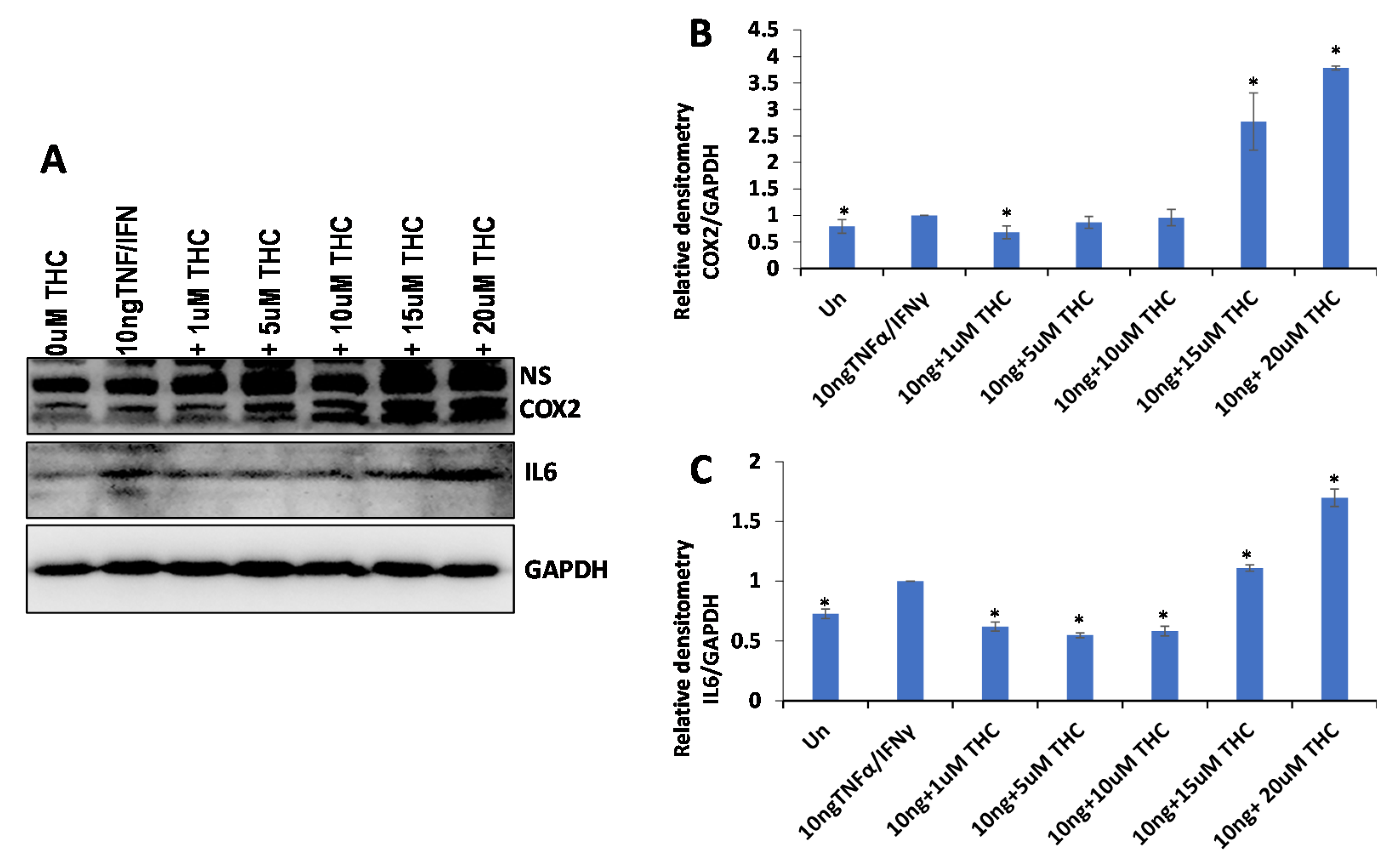
2.4. Anti-Inflammatory Properties of 25 Cannabis Extracts
2.5. Correlation Analysis
3. Methods
3.1. Source of Flowers and Extract Preparation
3.2. Analysis of Cannabinoids
3.3. Analysis of Terpenes
3.4. Cell Culture
3.5. Screening of Anti-Inflammatory Cannabis Extracts
3.6. MTT Assay
3.7. Western Blot Analysis
3.8. Statistical Analysis
3.9. Correlation Analysis
3.10. Clustering Analysis
3.11. PCA Analysis
4. Discussion
4.1. Anti-Inflammatory Effect—THC vs. Other Cannabinoids
4.2. Correlation between Cannabinoids and Terpenes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Lewis, M.M.; Yang, Y.; Wasilewski, E.; Clarke, H.A.; Kotra, L.P. Chemical Profiling of Medical Cannabis Extracts. ACS Omega 2017, 2, 6091–6103. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.; Amaral, C.; Teixeira, N.; Correia-da-Silva, G. Cannabis sativa: Much more beyond Delta(9)-tetrahydrocannabinol. Pharmacol. Res. 2020, 157, 104822. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.J.; Galettis, P.; Schneider, J. The pharmacokinetics and the pharmacodynamics of cannabinoids. Br. J. Clin. Pharmacol. 2018, 84, 2477–2482. [Google Scholar] [CrossRef] [PubMed]
- MacCallum, C.A.; Russo, E.B. Practical considerations in medical cannabis administration and dosing. Eur. J. Intern. Med. 2018, 49, 12–19. [Google Scholar] [CrossRef]
- Kovalchuk, A.; Wang, B.; Li, D.; Rodriguez-Juarez, R.; Ilnytskyy, S.; Kovalchuk, I.; Kovalchuk, O. Fighting the storm: Could novel anti-TNFalpha and anti-IL-6 C. sativa cultivars tame cytokine storm in COVID-19? Aging 2021, 13, 1571–1590. [Google Scholar] [CrossRef]
- Wang, B.; Li, D.; Fiselier, A.; Kovalchuk, I.; Kovalchuk, O. New AKT-dependent mechanisms of anti-COVID-19 action of high-CBD Cannabis sativa extracts. Cell Death Discov. 2022, 8, 110. [Google Scholar] [CrossRef]
- Wang, B.; Kovalchuk, A.; Li, D.; Rodriguez-Juarez, R.; Ilnytskyy, Y.; Kovalchuk, I.; Kovalchuk, O. In search of preventative strategies: Novel high-CBD cannabis sativa extracts modulate ACE2 expression in COVID-19 gateway tissues. Aging 2020, 12, 22425–22444. [Google Scholar] [CrossRef]
- Russo, E.B. Taming THC: Potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef]
- LaVigne, J.E.; Hecksel, R.; Keresztes, A.; Streicher, J.M. Cannabis sativa terpenes are cannabimimetic and selectively enhance cannabinoid activity. Sci. Rep. 2021, 11, 8232. [Google Scholar] [CrossRef]
- Baek, S.H.; Kim, Y.O.; Kwag, J.S.; Choi, K.E.; Jung, W.Y.; Han, D.S. Boron trifluoride etherate on silica-A modified Lewis acid reagent (VII). Antitumor activity of cannabigerol against human oral epitheloid carcinoma cells. Arch. Pharm. Res. 1998, 21, 353–356. [Google Scholar] [CrossRef]
- Lah, T.T.; Novak, M.; Pena Almidon, M.A.; Marinelli, O.; Žvar Baškovič, B.; Majc, B.; Mlinar, M.; Bošnjak, R.; Breznik, B.; Zomer, R.; et al. Cannabigerol Is a Potential Therapeutic Agent in a Novel Combined Therapy for Glioblastoma. Cells 2021, 10, 340. [Google Scholar] [CrossRef]
- Peeri, H.; Shalev, N.; Vinayaka, A.; Nizar, R.; Kazimirsky, G.; Namdar, D.; Anil, S.; Belausov, E.; Brodie, C.; Koltai, H. Specific Compositions of Cannabis sativa Compounds Have Cytotoxic Activity and Inhibit Motility and Colony Formation of Human Glioblastoma Cells In Vitro. Cancers 2021, 13, 1720. [Google Scholar] [CrossRef] [PubMed]
- Nahler, G. Cannabidiol and Other Phytocannabinoids as Cancer Therapeutics. Pharm. Med. 2022, 36, 99–129. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.A.; Dalgleish, A.G.; Liu, W.M. The combination of cannabidiol and Delta9-tetrahydrocannabinol enhances the anticancer effects of radiation in an orthotopic murine glioma model. Mol. Cancer Ther. 2014, 13, 2955–2967. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.B.; Seijo-Vila, M.; Caro-Villalobos, M.; Tundidor, I.; Andradas, C.; García-Taboada, E.; Wade, J.; Smith, S.; Guzmán, M.; Pérez-Gómez, E.; et al. Appraising the “entourage effect”: Antitumor action of a pure cannabinoid versus a botanical drug preparation in preclinical models of breast cancer. Biochem. Pharmacol. 2018, 157, 285–293. [Google Scholar] [CrossRef]
- Ligresti, A.; Moriello, A.S.; Starowicz, K.; Matias, I.; Pisanti, S.; De Petrocellis, L.; Laezza, C.; Portella, G.; Bifulco, M.; Di Marzo, V. Antitumor Activity of Plant Cannabinoids with Emphasis on the Effect of Cannabidiol on Human Breast Carcinoma. J. Pharmacol. Exp. Ther. 2006, 318, 1375–1387. [Google Scholar] [CrossRef]
- Raup-Konsavage, W.M.; Carkaci-Salli, N.; Greenland, K.; Gearhart, R.; Vrana, K.E. Cannabidiol (CBD) Oil Does Not Display an Entourage Effect in Reducing Cancer Cell Viability in vitro. Med. Cannabis Cannabinoids 2020, 3, 95–102. [Google Scholar] [CrossRef]
- Tomko, A.M.; Whynot, E.G.; Ellis, L.D.; Dupré, D.J. Anti-Cancer Potential of Cannabinoids, Terpenes, and Flavonoids Present in Cannabis. Cancers 2020, 12, 1985. [Google Scholar] [CrossRef]
- Murata, S.; Shiragami, R.; Kosugi, C.; Tezuka, T.; Yamazaki, M.; Hirano, A.; Yoshimura, Y.; Suzuki, M.; Shuto, K.; Ohkohchi, N.; et al. Antitumor effect of 1, 8-cineole against colon cancer. Oncol. Rep. 2013, 30, 2647–2652. [Google Scholar] [CrossRef]
- Abdalla, A.N.; Shaheen, U.; Abdallah, Q.M.A.; Flamini, G.; Bkhaitan, M.M.; Abdelhady, M.I.S.; Ascrizzi, R.; Bader, A. Proapoptotic Activity of Achillea membranacea Essential Oil and Its Major Constituent 1,8-Cineole against A2780 Ovarian Cancer Cells. Molecules 2020, 25, 1582. [Google Scholar] [CrossRef] [Green Version]
- Jamali, T.; Kavoosi, G.; Ardestani, S.K. In-vitro and in-vivo anti-breast cancer activity of OEO (Oliveria decumbens vent essential oil) through promoting the apoptosis and immunomodulatory effects. J. Ethnopharmacol. 2020, 248, 112313. [Google Scholar] [CrossRef] [PubMed]
- Rolim, T.L.; Meireles, D.R.P.; Batista, T.M.; De Sousa, T.K.G.; Mangueira, V.M.; De Abrantes, R.A.; Pita, J.C.L.R.; Xavier, A.L.; Costa, V.C.O.; Batista, L.M.; et al. Toxicity and antitumor potential of Mesosphaerum sidifolium (Lamiaceae) oil and fenchone, its major component. BMC Complement. Altern. Med. 2017, 17, 347. [Google Scholar] [CrossRef]
- Finlay, D.B.; Sircombe, K.J.; Nimick, M.; Jones, C.; Glass, M. Terpenoids from Cannabis Do Not Mediate an Entourage Effect by Acting at Cannabinoid Receptors. Front. Pharmacol. 2020, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Heblinski, M.; Santiago, M.; Fletcher, C.; Stuart, J.; Connor, M.; McGregor, I.S.; Arnold, J.C. Terpenoids Commonly Found in Cannabis sativa Do Not Modulate the Actions of Phytocannabinoids or Endocannabinoids on TRPA1 and TRPV1 Channels. Cannabis Cannabinoid Res. 2020, 5, 305–317. [Google Scholar] [CrossRef]
- Ismail, M.; Hasan, H.; El-Orfali, Y.; Ismail, H.; Khawaja, G. Anti-Inflammatory, Antioxidative, and Hepatoprotective Effects of Trans Delta9-Tetrahydrocannabinol/Sesame Oil on Adjuvant-Induced Arthritis in Rats. Evid. Based Complement. Altern. Med. 2018, 2018, 9365464. [Google Scholar] [CrossRef]
- Palomares, B.; Garrido-Rodriguez, M.; Gonzalo-Consuegra, C.; Gómez-Cañas, M.; Saen-oon, S.; Soliva, R.; Collado, J.A.; Fernández-Ruiz, J.; Morello, G.; Calzado, M.A.; et al. Delta(9)-Tetrahydrocannabinolic acid alleviates collagen-induced arthritis: Role of PPARgamma and CB1 receptors. Br. J. Pharmacol. 2020, 177, 4034–4054. [Google Scholar] [CrossRef]
- Motwani, M.P.; Bennett, F.; Norris, P.; Maini, A.A.; George, M.J.; Newson, J.; Henderson, A.; Hobbs, A.J.; Tepper, M.; White, B.; et al. Potent Anti-Inflammatory and Pro-Resolving Effects of Anabasum in a Human Model of Self-Resolving Acute Inflammation. Clin. Pharmacol. Ther. 2018, 104, 675–686. [Google Scholar] [CrossRef]
- Suryavanshi, S.V.; Zaiachuk, M.; Pryimak, N.; Kovalchuk, I.; Kovalchuk, O. Cannabinoids Alleviate the LPS-Induced Cytokine Storm via Attenuating NLRP3 Inflammasome Signaling and TYK2-Mediated STAT3 Signaling Pathways In Vitro. Cells 2022, 11, 1391. [Google Scholar] [CrossRef]
- Borisov, N.; Ilnytskyy, Y.; Byeon, B.; Kovalchuk, O.; Kovalchuk, I. System, Method and Software for Calculation of a Cannabis Drug Efficiency Index for the Reduction of Inflammation. Int. J. Mol. Sci. 2020, 22, 388. [Google Scholar] [CrossRef]
- Paula-Freire, L.; Andersen, M.; Gama, V.; Molska, G.; Carlini, E. The oral administration of trans-caryophyllene attenuates acute and chronic pain in mice. Phytomedicine 2014, 21, 356–362. [Google Scholar] [CrossRef]
- Javed, H.; Meeran, M.F.N.; Azimullah, S.; Eddin, L.B.; Dwivedi, V.D.; Jha, N.K.; Ojha, S. α-Bisabolol, a Dietary Bioactive Phytochemical Attenuates Dopaminergic Neurodegeneration through Modulation of Oxidative Stress, Neuroinflammation and Apoptosis in Rotenone-Induced Rat Model of Parkinson’s disease. Biomolecules 2020, 10, 1421. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, D.; Chen, Y.; Yang, M. β-Caryophyllene inhibits high glucose-induced oxidative stress, inflammation and extracellular matrix accumulation in mesangial cells. Int. Immunopharmacol. 2020, 84, 106556. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Zhang, T.; Yin, N.; Ma, X.; Zhao, G.; Wu, H.; Qiu, C.; Deng, G. Geraniol alleviates LPS-induced acute lung injury in mice via inhibiting inflammation and apoptosis. Oncotarget 2017, 8, 71038–71053. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.J.; Lee, D.-U.; Shin, H.M. Inhibitory Effect of Valencene on the Development of Atopic Dermatitis-Like Skin Lesions in NC/Nga Mice. Evid.-Based Complement. Altern. Med. 2016, 2016, 9370893. [Google Scholar] [CrossRef]
- Cabrera, C.L.R.; Keir-Rudman, S.; Horniman, N.; Clarkson, N.; Page, C. The anti-inflammatory effects of cannabidiol and cannabigerol alone, and in combination. Pulm. Pharmacol. Ther. 2021, 69, 102047. [Google Scholar] [CrossRef]
- Anil, S.M.; Shalev, N.; Vinayaka, A.C.; Nadarajan, S.; Namdar, D.; Belausov, E.; Shoval, I.; Mani, K.A.; Mechrez, G.; Koltai, H. Cannabis compounds exhibit anti-inflammatory activity in vitro in COVID-19-related inflammation in lung epithelial cells and pro-inflammatory activity in macrophages. Sci. Rep. 2021, 11, 1462. [Google Scholar] [CrossRef]
- Hanus, L.O.; Hod, Y. Terpenes/Terpenoids in Cannabis: Are They Important? Med. Cannabis Cannabinoids 2020, 3, 25–60. [Google Scholar] [CrossRef]
- Aizpurua-Olaizola, O.; Soydaner, U.; Öztürk, E.; Schibano, D.; Simsir, Y.; Navarro, P.; Etxebarria, N.; Usobiaga, A. Evolution of the Cannabinoid and Terpene Content during the Growth of Cannabis sativa Plants from Different Chemotypes. J. Nat. Prod. 2016, 79, 324–331. [Google Scholar] [CrossRef]
- Hillig, K.W.; Mahlberg, P.G. A chemotaxonomic analysis of cannabinoid variation in Cannabis (Cannabaceae). Am. J. Bot. 2004, 91, 966–975. [Google Scholar] [CrossRef]
- Jin, D.; Henry, P.; Shan, J.; Chen, J. Identification of Chemotypic Markers in Three Chemotype Categories of Cannabis Using Secondary Metabolites Profiled in Inflorescences, Leaves, Stem Bark, and Roots. Front. Plant Sci. 2021, 12, 699530. [Google Scholar] [CrossRef]
- Bautista, J.L.; Yu, S.; Tian, L. Flavonoids in Cannabis sativa: Biosynthesis, Bioactivities, and Biotechnology. ACS Omega 2021, 6, 5119–5123. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer, % | Normal, % | Cancer, % | Normal, % | |
|---|---|---|---|---|
| #1 | 76.08 | 47.40 | ||
| #2 | 79.21 | 12.00 | ||
| #3 | 85.12 | 39.10 | ||
| #4 | 79.38 | 52.20 | ||
| #5 | 86.50 | 53.90 | ||
| #6 | 77.17 | 36.40 | ||
| #7 | 81.85 | 31.90 | ||
| #8 | 84.59 | 40.80 | ||
| #9 | 81.72 | 40.80 | ||
| #10 | 84.13 | 37.30 | ||
| #11 | 79.59 | 43.50 | ||
| #12 | 78.77 | 41.40 | ||
| #13 | 78.31 | 33.80 | ||
| #14 | 71.35 | 35.20 | ||
| #15 | 82.27 | 36.90 | ||
| #16 | 65.52 | 37.50 | ||
| #17 | 65.75 | 20.10 | ||
| #18 | 79.25 | 48.70 | ||
| #19 | 75.73 | 16.80 | ||
| #20 | 73.05 | 40.00 | ||
| #21 | 88.61 | 21.80 | 95.86 | 24.50 |
| #22 | 84.35 | 18.00 | 95.23 | 32.70 |
| #23 | 74.68 | −11.00 | 90.28 | 5.90 |
| #24 | 92.03 | 10.10 | 96.50 | 6.85 |
| #25 | 86.70 | 16.20 | 96.44 | 20.05 |
| Average | 79.67 | 32.03 | 94.86 | 18.00 |
| Cultivar Name | WI38 | WI38 | HSIEC | |||
|---|---|---|---|---|---|---|
| COX2 | IL6 | COX2 | IL6 | COX2 | IL6 | |
| #1 | 1.00 | 1.65 | ||||
| #2 | 1.79 | 1.99 | 5.16 | 1.26 | 0.50 | 1.61 |
| #3 | 1.17 | 1.93 | ||||
| #4 | 1.11 | 1.10 | ||||
| #5 | 1.29 | 0.89 | ||||
| #6 | 1.40 | 0.86 | ||||
| #7 | 1.58 | 0.99 | 4.24 | 1.19 | 0.48 | 1.74 |
| #8 | 0.89 | 2.12 | ||||
| #9 | 1.10 | 1.65 | ||||
| #10 | 0.71 | 1.87 | ||||
| #11 | 0.48 | 1.53 | ||||
| #12 | 0.33 | 1.53 | ||||
| #13 | 0.26 | 1.96 | ||||
| #14 | 0.80 | 0.76 | ||||
| #15 | 0.81 | 0.59 | ||||
| #16 | 0.65 | 0.71 | ||||
| #17 | 1.39 | 1.24 | 3.09 | 1.89 | 1.71 | 1.84 |
| #18 | 0.76 | 0.56 | ||||
| #19 | 1.00 | 0.33 | 2.11 | 1.21 | 1.78 | 1.53 |
| #20 | 0.83 | 1.16 | ||||
| #21 | 0.93 | 0.47 | ||||
| #22 | 1.26 | 3.43 | ||||
| #23 | 1.05 | 0.91 | ||||
| #24 | 1.22 | 1.78 | 2.90 | 1.85 | 0.43 | 1.30 |
| #25 | 1.63 | 1.07 | ||||
| Average | 1.02 | 1.32 | 3.50 | 1.48 | 0.98 | 1.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Ilnytskyy, Y.; Ghasemi Gojani, E.; Kovalchuk, O.; Kovalchuk, I. Analysis of Anti-Cancer and Anti-Inflammatory Properties of 25 High-THC Cannabis Extracts. Molecules 2022, 27, 6057. https://doi.org/10.3390/molecules27186057
Li D, Ilnytskyy Y, Ghasemi Gojani E, Kovalchuk O, Kovalchuk I. Analysis of Anti-Cancer and Anti-Inflammatory Properties of 25 High-THC Cannabis Extracts. Molecules. 2022; 27(18):6057. https://doi.org/10.3390/molecules27186057
Chicago/Turabian StyleLi, Dongping, Yaroslav Ilnytskyy, Esmaeel Ghasemi Gojani, Olga Kovalchuk, and Igor Kovalchuk. 2022. "Analysis of Anti-Cancer and Anti-Inflammatory Properties of 25 High-THC Cannabis Extracts" Molecules 27, no. 18: 6057. https://doi.org/10.3390/molecules27186057