
Cannabidiol Promotes Neuronal Differentiation Using Akt and Erk Pathways Triggered by Cb1 Signaling
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
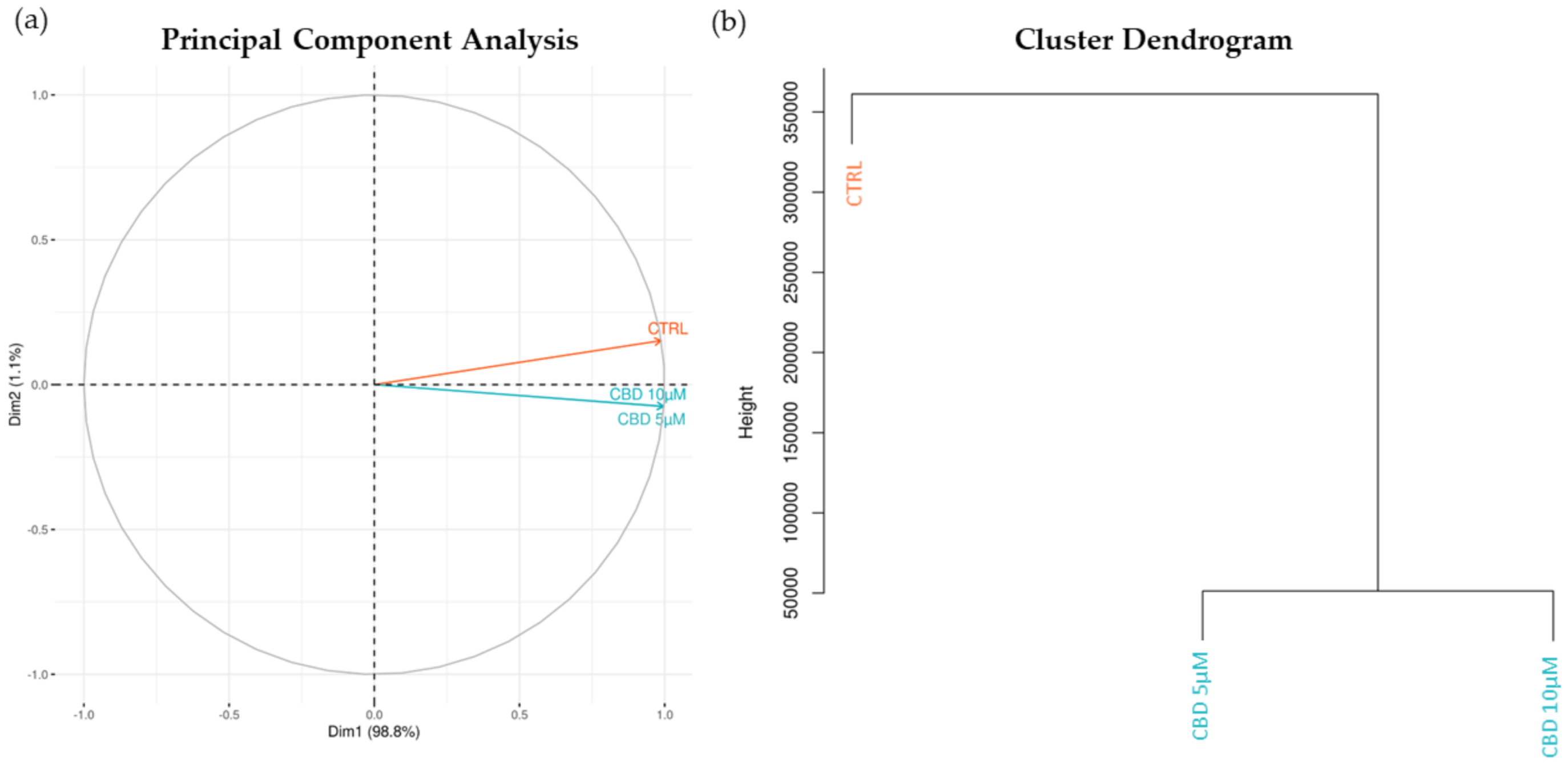
2.1. Transcriptomic Analysis
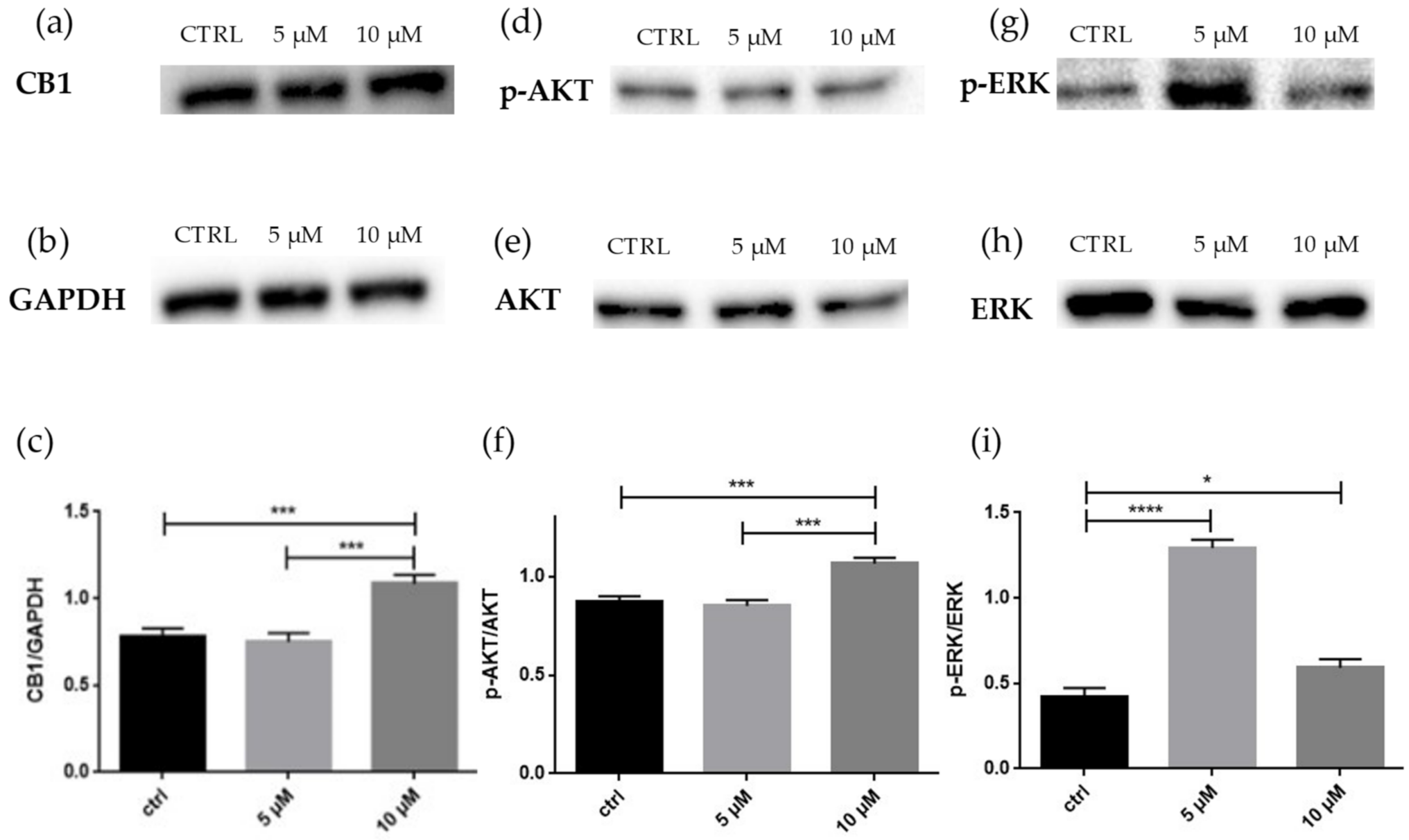
2.2. Western Blot Analyses
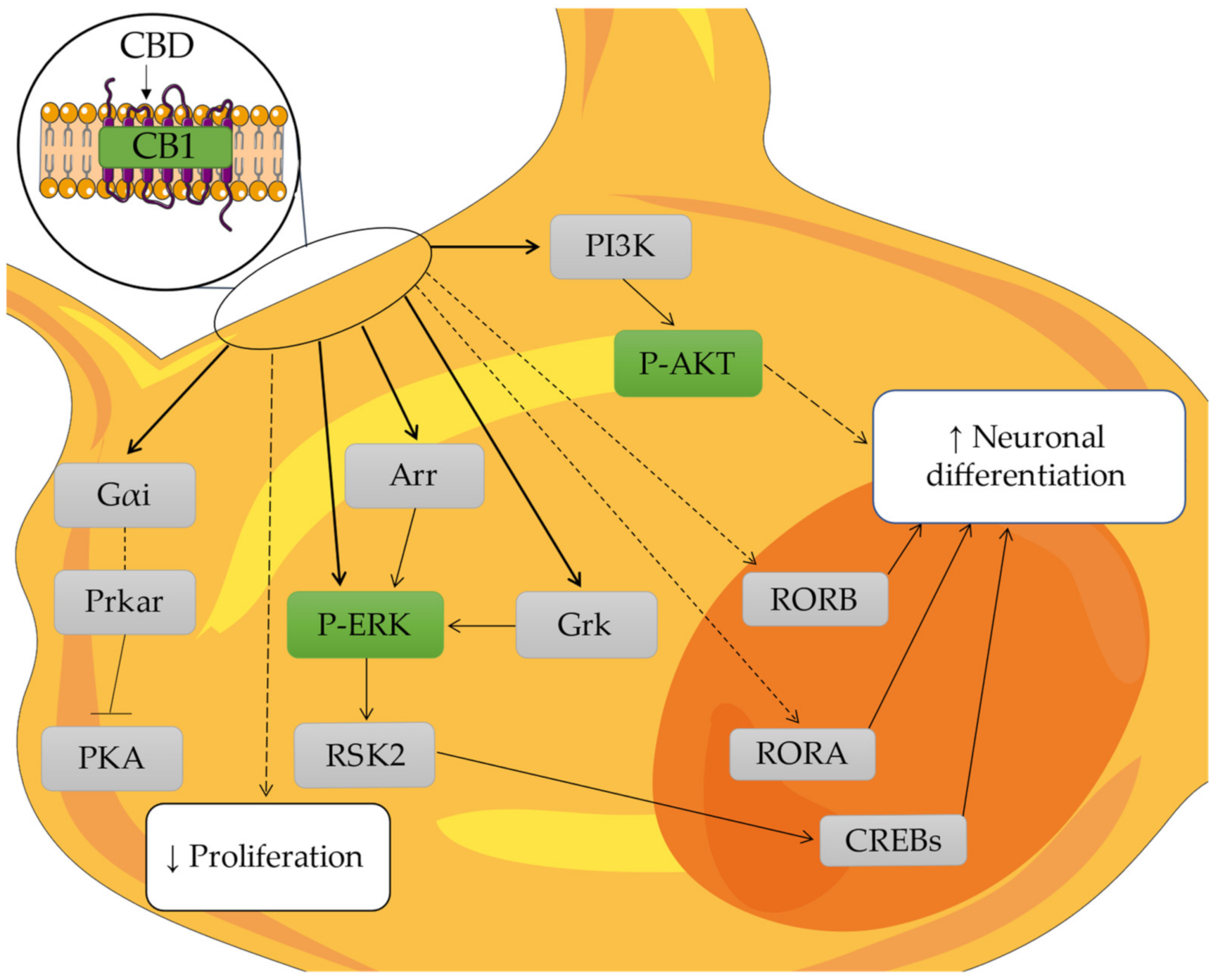
3. Discussion
4. Materials and Methods
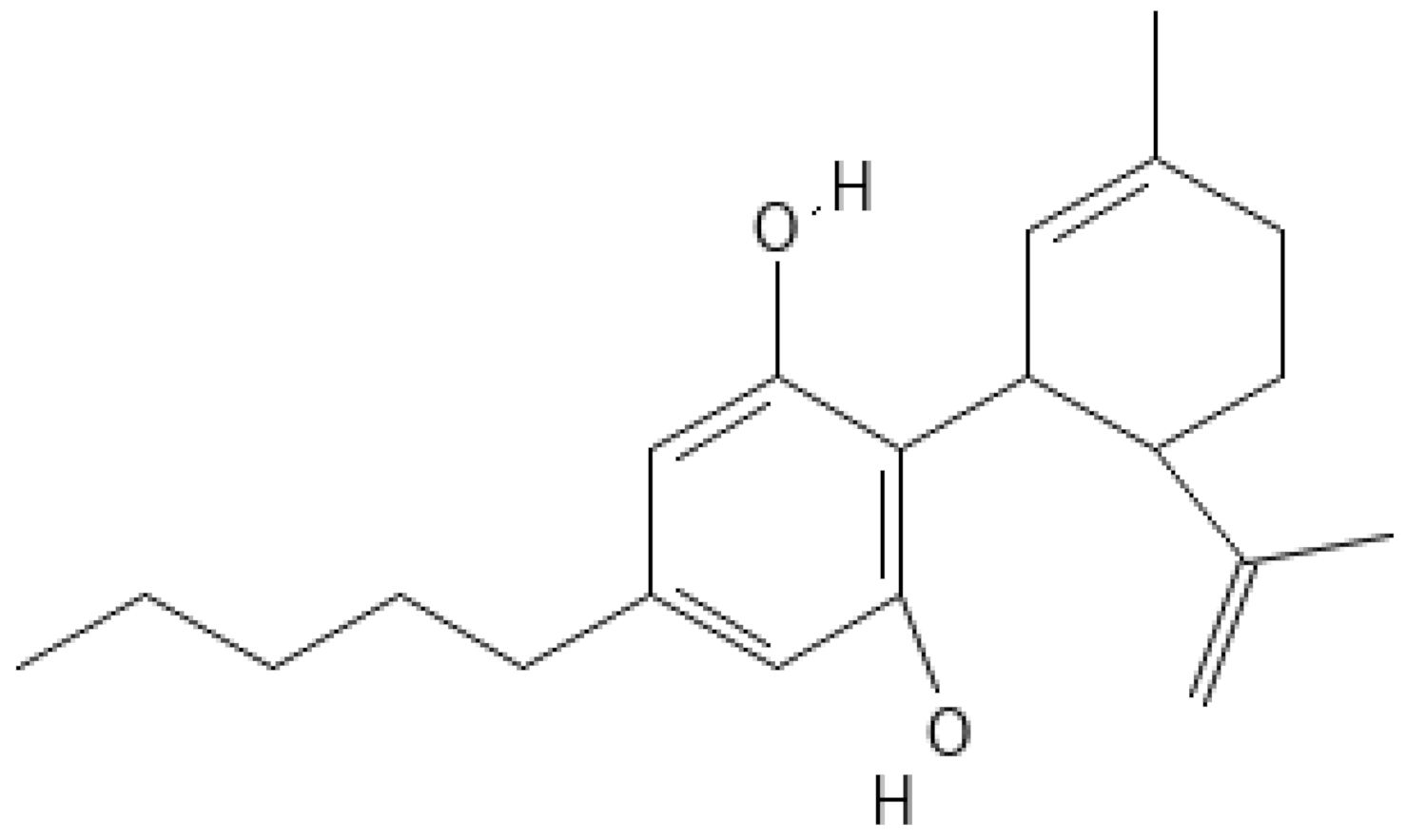
4.1. CBD Extraction from Cannabis Sativa
4.2. NSC-34 Culture and Treatment
4.3. Library Preparation and Bioinformatics Inspection
4.4. Western Blot Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Crocq, M.-A. History of cannabis and the endocannabinoid system. Dialogues Clin. Neurosci. 2020, 22, 223–228. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Oddi, S.; Scipioni, L.; Maccarrone, M. Endocannabinoid system and adult neurogenesis: A focused review. Curr. Opin. Pharmacol. 2020, 50, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; Guzmán, M.; Mackie, K.; Doherty, P.; Harkany, T. Programming of neural cells by (endo)cannabinoids: From physiological rules to emerging therapies. Nat. Rev. Neurosci. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Zorina, Y.; Iyengar, R.; Bromberg, K.D. Chapter 203-Effectors of Gαo. In Handbook of Cell Signaling, 2nd ed.; Bradshaw, R.A., Dennis, E.A., Eds.; Academic Press: San Diego, CA, USA, 2010; pp. 1655–1663. [Google Scholar]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.H.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International union of basic and clinical pharmacology. LXXIX. Cannabinoid receptors and their ligands: Beyond CB1 and CB2. Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef]
- Valeri, A.; Mazzon, E. Cannabinoids and neurogenesis: The promised solution for neurodegeneration? Molecules 2021, 26, 6313. [Google Scholar] [CrossRef]
- Leo, L.M.; Abood, M.E. CB1 cannabinoid receptor signaling and biased signaling. Molecules 2021, 26, 5413. [Google Scholar] [CrossRef]
- Lu, D.; Immadi, S.S.; Wu, Z.; Kendall, D.A. Translational potential of allosteric modulators targeting the cannabinoid CB1 receptor. Acta Pharmacol. Sin. 2019, 40, 324–335. [Google Scholar] [CrossRef]
- Atalay, S.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Antioxidative and anti-inflammatory properties of cannabidiol. Antioxidants 2020, 9, 21. [Google Scholar] [CrossRef]
- Burstein, S. Cannabidiol (CBD) and its analogs: A review of their effects on inflammation. Bioorg. Med. Chem. 2015, 23, 1377–1385. [Google Scholar] [CrossRef]
- Haspula, D.; Clark, M.A. Cannabinoid receptors: An update on cell signaling, pathophysiological roles and therapeutic opportunities in neurological, cardiovascular, and inflammatory diseases. Int. J. Mol. Sci. 2020, 21, 7693. [Google Scholar] [CrossRef] [PubMed]
- Tham, M.; Yilmaz, O.; Alaverdashvili, M.; Kelly, M.E.M.; Denovan-Wright, E.M.; Laprairie, R.B. Allosteric and orthosteric pharmacology of cannabidiol and cannabidiol-dimethylheptyl at the type 1 and type 2 cannabinoid receptors. Br. J. Pharmacol. 2019, 176, 1455–1469. [Google Scholar] [CrossRef]
- Soundara Rajan, T.; Giacoppo, S.; Scionti, D.; Diomede, F.; Grassi, G.; Pollastro, F.; Piattelli, A.; Bramanti, P.; Mazzon, E.; Trubiani, O. Cannabidiol activates neuronal precursor genes in human gingival mesenchymal stromal cells. J. Cell. Biochem. 2017, 118, 1531–1546. [Google Scholar] [CrossRef] [PubMed]
- Cashman, N.R.; Durham, H.D.; Blusztajn, J.K.; Oda, K.; Tabira, T.; Shaw, I.T.; Dahrouge, S.; Antel, J.P. Neuroblastoma × spinal cord (NSC) hybrid cell lines resemble developing motor neurons. Dev. Dyn. 1992, 194, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Martet, M.; Mestre, L.; Loría, F.; Guaza, C.; Fernández-Ruiz, J.; de Lago, E. Identification of receptors and enzymes for endocannabinoids in NSC-34 cells: Relevance for in vitro studies with cannabinoids in motor neuron diseases. Neurosci. Lett. 2012, 508, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Aguado, T.; Palazuelos, J.; Monory, K.; Stella, N.; Cravatt, B.; Lutz, B.; Marsicano, G.; Kokaia, Z.; Guzmán, M.; Galve-Roperh, I. The endocannabinoid system promotes astroglial differentiation by acting on neural progenitor cells. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 1551–1561. [Google Scholar] [CrossRef]
- Díaz-Alonso, J.; Guzmán, M.; Galve-Roperh, I. Endocannabinoids via CB1 receptors act as neurogenic niche cues during cortical development. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3229–3241. [Google Scholar] [CrossRef]
- Puighermanal, E.; Marsicano, G.; Busquets-Garcia, A.; Lutz, B.; Maldonado, R.; Ozaita, A. Cannabinoid modulation of hippocampal long-term memory is mediated by mTOR signaling. Nat. Neurosci. 2009, 12, 1152–1158. [Google Scholar] [CrossRef]
- Laun, A.S.; Shrader, S.H.; Brown, K.J.; Song, Z.-H. GPR3, GPR6, and GPR12 as novel molecular targets: Their biological functions and interaction with cannabidiol. Acta Pharmacol. Sin. 2019, 40, 300–308. [Google Scholar] [CrossRef]
- Nogueras-Ortiz, C.; Yudowski, G.A. The multiple waves of cannabinoid 1 receptor signaling. Mol. Pharmacol. 2016, 90, 620–626. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Peraza, F.; Ahn, K.H.; Nogueras-Ortiz, C.; Mungrue, I.N.; Mackie, K.; Kendall, D.A.; Yudowski, G.A. Mechanisms of biased β-arrestin-mediated signaling downstream from the cannabinoid 1 receptor. Mol. Pharmacol. 2016, 89, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Compagnucci, C.; Di Siena, S.; Bustamante, M.B.; Di Giacomo, D.; Di Tommaso, M.; Maccarrone, M.; Grimaldi, P.; Sette, C. Type-1 (CB1) cannabinoid receptor promotes neuronal differentiation and maturation of neural stem cells. PLoS ONE 2013, 8, e54271. [Google Scholar] [CrossRef] [PubMed]
- Iroegbu, J.D.; Ijomone, O.K.; Femi-Akinlosotu, O.M.; Ijomone, O.M. ERK/MAPK signalling in the developing brain: Perturbations and consequences. Neurosci. Biobehav. Rev. 2021, 131, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Laun, A.S.; Song, Z.H. GPR3 and GPR6, novel molecular targets for cannabidiol. Biochem. Biophys. Res. Commun. 2017, 490, 17–21. [Google Scholar] [CrossRef]
- Tanaka, S.; Miyagi, T.; Dohi, E.; Seki, T.; Hide, I.; Sotomaru, Y.; Saeki, Y.; Antonio Chiocca, E.; Matsumoto, M.; Sakai, N. Developmental expression of GPR3 in rodent cerebellar granule neurons is associated with cell survival and protects neurons from various apoptotic stimuli. Neurobiol. Dis. 2014, 68, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Landeira, B.S.; Santana, T.; Araújo, J.A.M.; Tabet, E.I.; Tannous, B.A.; Schroeder, T.; Costa, M.R. Activity-independent effects of CREB on neuronal survival and differentiation during mouse cerebral cortex development. Cereb. Cortex 2018, 28, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Cañón, E.; Cosgaya, J.M.; Scsucova, S.; Aranda, A. Rapid effects of retinoic acid on CREB and ERK phosphorylation in neuronal cells. Mol. Biol. Cell 2004, 15, 5583–5592. [Google Scholar] [CrossRef]
- Mehta, G.; Sanyal, K.; Abhishek, S.; Rajakumara, E.; Ghosh, S.K. Minichromosome maintenance proteins in eukaryotic chromosome segregation. BioEssays 2022, 44, 2100218. [Google Scholar] [CrossRef]
- Shivji, M.K.K.; Kenny, M.K.; Wood, R.D. Proliferating cell nuclear antigen is required for DNA excision repair. Cell 1992, 69, 367–374. [Google Scholar] [CrossRef]
- Graefe, C.; Eichhorn, L.; Wurst, P.; Kleiner, J.; Heine, A.; Panetas, I.; Abdulla, Z.; Hoeft, A.; Frede, S.; Kurts, C.; et al. Optimized Ki-67 staining in murine cells: A tool to determine cell proliferation. Mol. Biol. Rep. 2019, 46, 4631–4643. [Google Scholar] [CrossRef]
- Fu, F.; Li, L.S.; Li, R.; Deng, Q.; Yu, Q.X.; Yang, X.; Pan, M.; Han, J.; Zhen, L.; Zhang, L.N.; et al. All-trans-retinoid acid induces the differentiation of P19 cells into neurons involved in the PI3K/Akt/GSK3β signaling pathway. J. Cell. Biochem. 2020, 121, 4386–4396. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-Y.; Chang, C.-T.; Han, Y.-T.; Liao, C.-P.; Yu, J.-Y.; Wang, T.-W. Ginkgolide B promotes neuronal differentiation through the Wnt/β-catenin pathway in neural stem cells of the postnatal mammalian subventricular zone. Sci. Rep. 2018, 8, 14947. [Google Scholar] [CrossRef] [PubMed]
- Grünblatt, E.; Bartl, J.; Walitza, S. Methylphenidate enhances neuronal differentiation and reduces proliferation concomitant to activation of Wnt signal transduction pathways. Transl. Psychiatry 2018, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Adelstein, R.S.; Kawamoto, S. Identification of neuronal nuclei (NeuN) as Fox-3, a new member of the Fox-1 gene family of splicing factors*. J. Biol. Chem. 2009, 284, 31052–31061. [Google Scholar] [CrossRef] [PubMed]
- Rosskothen-Kuhl, N.; Illing, R.B. Gap43 transcription modulation in the adult brain depends on sensory activity and synaptic cooperation. PLoS ONE 2014, 9, e92624. [Google Scholar] [CrossRef]
- Bustos, F.J.; Ampuero, E.; Jury, N.; Aguilar, R.; Falahi, F.; Toledo, J.; Ahumada, J.; Lata, J.; Cubillos, P.; Henríquez, B.; et al. Epigenetic editing of the Dlg4/PSD95 gene improves cognition in aged and Alzheimer’s disease mice. Brain 2017, 140, 3252–3268. [Google Scholar] [CrossRef]
- Ogorodnikov, A.; Levin, M.; Tattikota, S.; Tokalov, S.; Hoque, M.; Scherzinger, D.; Marini, F.; Poetsch, A.; Binder, H.; Macher-Göppinger, S.; et al. Transcriptome 3′end organization by PCF11 links alternative polyadenylation to formation and neuronal differentiation of neuroblastoma. Nat. Commun. 2018, 9, 5331. [Google Scholar] [CrossRef] [PubMed]
- Isgrò, M.A.; Bottoni, P.; Scatena, R. Neuron-specific enolase as a biomarker: Biochemical and clinical aspects. In Advances in Cancer Biomarkers: From Biochemistry to Clinic for a Critical Revision; Scatena, R., Ed.; Springer: Dordrecht, The Netherland, 2015; pp. 125–143. [Google Scholar]
- Kojetin, D.J.; Burris, T.P. REV-ERB and ROR nuclear receptors as drug targets. Nat. Rev. Drug Discov. 2014, 13, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Kim, H.; Baek, S.H. Unraveling the physiological roles of retinoic acid receptor-related orphan receptor α. Exp. Mol. Med. 2021, 53, 1278–1286. [Google Scholar] [CrossRef]
- Liu, H.; Aramaki, M.; Fu, Y.; Forrest, D. Chapter eight-Retinoid-related orphan receptor β and transcriptional control of neuronal differentiation. In Current Topics in Developmental Biology; Forrest, D., Tsai, S., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 125, pp. 227–255. [Google Scholar]
- Bojesen, I.N.; Hansen, H.S. Membrane transport of anandamide through resealed human red blood cell membranes. J. Lipid Res. 2005, 46, 1652–1659. [Google Scholar] [CrossRef] [Green Version]
- Bragança, V.A.N.; França, T.G.; de Jesus, A.C.S.P.S.; Palheta, I.C.; Melo, F.P.A.; Neves, P.A.P.F.G.; Lima, A.B.; Borges, R.S. Impact of conformational and solubility properties on psycho-activity of cannabidiol (CBD) and tetrahydrocannabinol (THC). Chem. Data Collect. 2020, 26, 100345. [Google Scholar] [CrossRef]
- Fezza, F.; Oddi, S.; Di Tommaso, M.; De Simone, C.; Rapino, C.; Pasquariello, N.; Dainese, E.; Finazzi-Agrò, A.; Maccarrone, M. Characterization of biotin-anandamide, a novel tool for the visualization of anandamide accumulation. J. Lipid Res. 2008, 49, 1216–1223. [Google Scholar] [CrossRef]
- Kaczocha, M.; Glaser, S.T.; Deutsch, D.G. Identification of intracellular carriers for the endocannabinoid anandamide. Proc. Natl. Acad. Sci. USA 2009, 106, 6375–6380. [Google Scholar] [CrossRef] [PubMed]
- Oddi, S.; Fezza, F.; Pasquariello, N.; D’Agostino, A.; Catanzaro, G.; De Simone, C.; Rapino, C.; Finazzi-Agrò, A.; Maccarrone, M. Molecular identification of albumin and Hsp70 as cytosolic anandamide-binding proteins. Chem. Biol. 2009, 16, 624–632. [Google Scholar] [CrossRef]
- André, E.; Conquet, F.; Steinmayr, M.; Stratton, S.C.; Porciatti, V.; Becker-André, M. Disruption of retinoid-related orphan receptor beta changes circadian behavior, causes retinal degeneration and leads to vacillans phenotype in mice. EMBO J. 1998, 17, 3867–3877. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, H.; Ng, L.; Kim, J.-W.; Hao, H.; Swaroop, A.; Forrest, D. Feedback Induction of a photoreceptor-specific Isoform of retinoid-related orphan nuclear receptor β by the rod transcription factor NRL*. J. Biol. Chem. 2014, 289, 32469–32480. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.R.; Heck, N.; Lohof, A.M.; Rochefort, C.; Morel, M.P.; Wehrlé, R.; Doulazmi, M.; Marty, S.; Cannaya, V.; Avci, H.X.; et al. Mature purkinje cells require the retinoic acid-related orphan receptor-α (RORα) to maintain climbing fiber mono-innervation and other adult characteristics. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 9546–9562. [Google Scholar] [CrossRef]
- Janmaat, S.; Akwa, Y.; Doulazmi, M.; Bakouche, J.; Gautheron, V.; Liere, P.; Eychenne, B.; Pianos, A.; Luiten, P.; Groothuis, T.; et al. Age-related purkinje cell death is steroid dependent: RORα haplo-insufficiency impairs plasma and cerebellar steroids and Purkinje cell survival. Age 2011, 33, 565–578. [Google Scholar] [CrossRef]
- Gold, D.A.; Baek, S.H.; Schork, N.J.; Rose, D.W.; Larsen, D.D.; Sachs, B.D.; Rosenfeld, M.G.; Hamilton, B.A. RORalpha coordinates reciprocal signaling in cerebellar development through sonic hedgehog and calcium-dependent pathways. Neuron 2003, 40, 1119–1131. [Google Scholar] [CrossRef]
- Taglialatela-Scafati, O.; Pagani, A.; Scala, F.; De Petrocellis, L.; Di Marzo, V.; Grassi, G.; Appendino, G. Cannabimovone, a cannabinoid with a rearranged terpenoid skeleton from hemp. Eur. J. Org. Chem. 2010, 2010, 2067–2072. [Google Scholar] [CrossRef]
- Gugliandolo, A.; Pollastro, F.; Grassi, G.; Bramanti, P.; Mazzon, E. In vitro model of neuroinflammation: Efficacy of cannabigerol, a non-psychoactive cannabinoid. Int. J. Mol. Sci. 2018, 19, 1992. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, S.; Chiricosta, L.; Gugliandolo, A.; Pizzicannella, J.; Diomede, F.; Bramanti, P.; Trubiani, O.; Mazzon, E. Extracellular vesicles derived from human gingival mesenchymal stem cells: A transcriptomic analysis. Genes 2020, 11, 118. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | CTRL vs. 5 µM Fold Change | CTRL vs. 5 µM q-Value | CTRL vs. 10 µM Fold Change | CTRL vs. 10 µM q-Value |
|---|---|---|---|---|
| Cnr1 | 0.47 | 3.97 × 10−8 | 0.70 | 1.03 × 10−17 |
| Gpr3 | - | >0.05 | −1.18 | 5.95 × 10−4 |
| Gpr6 | 0.89 | 1.05 × 10−2 | - | >0.05 |
| Gnai1 | 1.45 | 5.75 × 10−4 | - | >0.05 |
| Gnai2 | 0.17 | 2.46 × 10−7 | 0.46 | 5.42 × 10−54 |
| Gnai3 | - | >0.05 | 0.29 | 3.55 × 10−10 |
| Gnas | −0.22 | 1.11 × 10−98 | 0.08 | 7.73 × 10−16 |
| Grk2 | - | >0.05 | −0.17 | 9.88 × 10−4 |
| Grk5 | - | >0.05 | 0.31 | 4.50 × 10−2 |
| Grk6 | 0.49 | 6.17 × 10−9 | 0.39 | 6.81 × 10−6 |
| Arrb1 | 0.36 | 1.78 × 10−3 | 0.54 | 5.94 × 10−7 |
| Arrb2 | 0.14 | 2.22 × 10−2 | - | >0.05 |
| Genes | CTRL vs. 5 µM Fold Change | CTRL vs. 5 µM q-Value | CTRL vs. 10 µM Fold Change | CTRL vs. 10 µM q-Value |
|---|---|---|---|---|
| Prkaca | - | >0.05 | 0.50 | 3.88 × 10−26 |
| Prkar1a | - | >0.05 | 0.42 | 1.85 × 10−72 |
| Prkar1b | 0.83 | 3.67 × 10−2 | - | >0.05 |
| Prkar2a | 0.29 | 8.07 × 10−4 | 0.59 | 1.50 × 10−13 |
| Prkar2b | 0.27 | 1.74 × 10−3 | 0.34 | 5.47 × 10−5 |
| Genes | CTRL vs. 5 µM Fold Change | CTRL vs. 5 µM q-Value | CTRL vs. 10 µM Fold Change | CTRL vs. 10 µM q-Value |
|---|---|---|---|---|
| Mapk3 | 0.32 | 1.23 × 10−11 | 0.67 | 1.61 × 10−52 |
| Mapk1 | 0.23 | 2.34 × 10−6 | 0.58 | 4.72 × 10−38 |
| Rps6ka3 | 0.11 | 3.68 × 10−3 | 0.54 | 9.21 × 10−66 |
| Creb1 | −0.17 | 4.50 × 10−2 | - | >0.05 |
| Atf4 | - | >0.05 | 0.28 | 4.67 × 10−23 |
| Creb5 | - | >0.05 | −1.2 | 8.90 × 10−3 |
| Genes | CTRL vs. 5 µM Fold Change | CTRL vs. 5 µM q-Value | CTRL vs. 10 µM Fold Change | CTRL vs. 10 µM q-Value |
|---|---|---|---|---|
| Cdc6 | −2.53 | 6.14 × 10−5 | −2.10 | 2.41 × 10−4 |
| Mcm3 | −0.15 | 1.17 × 10−3 | −0.13 | 3.78 × 10−3 |
| Mcm5 | −0.41 | 4.26 × 10−8 | −0.72 | 2.43 × 10−20 |
| Mcm6 | −0.25 | 3.67 × 10−5 | −0.17 | 6.11 × 10−3 |
| Mcm9 | - | >0.05 | 0.39 | 3.02 × 10−2 |
| Mcm10 | −0.35 | 3.32 × 10−2 | −0.56 | 5.43 × 10−4 |
| Pcna | −0.47 | 1.02 × 10−2 | −0.65 | 3.40 × 10−4 |
| Mki67 | −0.49 | 2.92 × 10−8 | −0.40 | 5.75 × 10−6 |
| Genes | CTRL vs. 5 µM Fold Change | CTRL vs. 5 µM q-Value | CTRL vs. 10 µM Fold Change | CTRL vs. 10 µM q-Value |
|---|---|---|---|---|
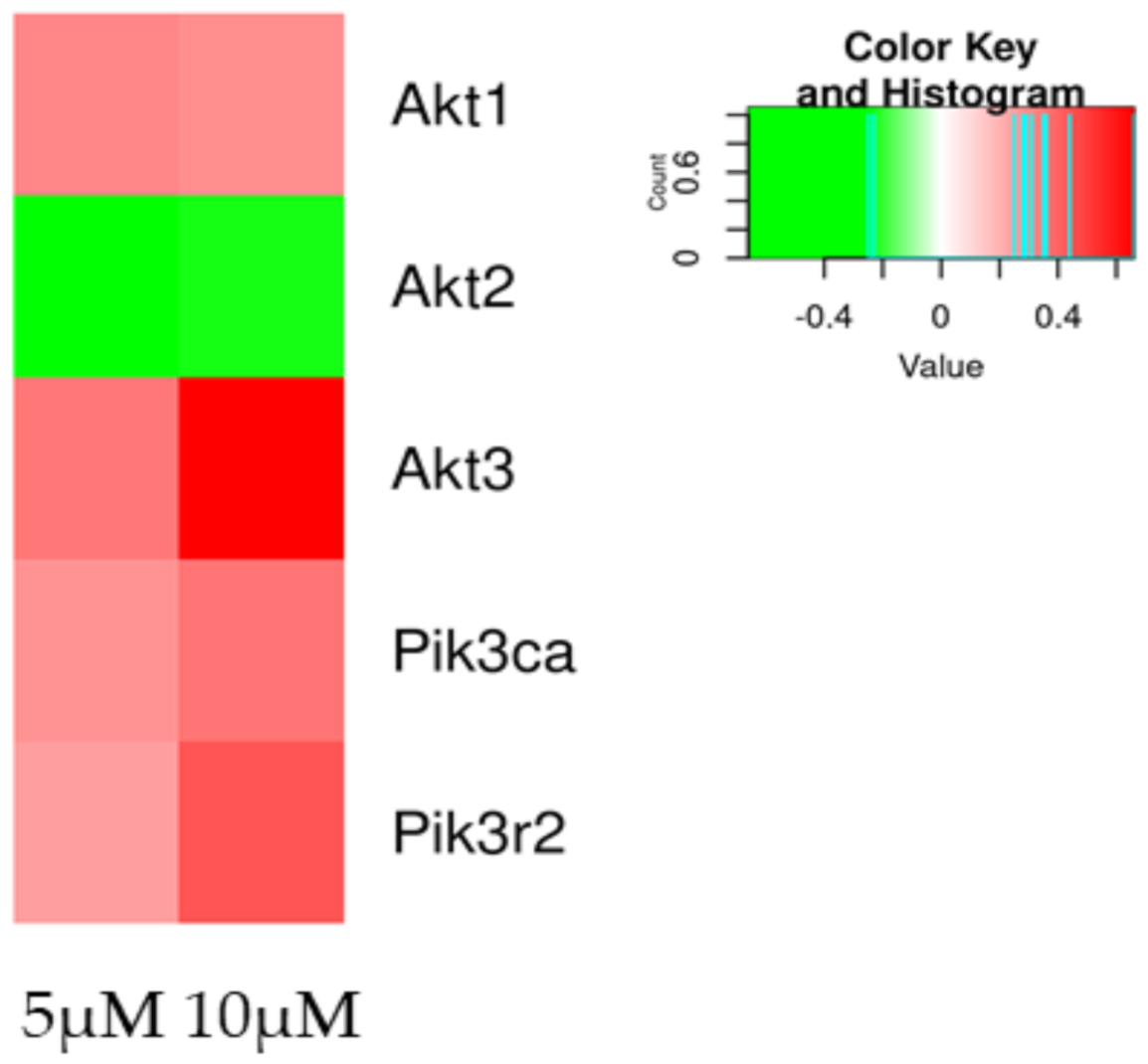
| Akt1 | 0.31 | 3.45 × 10−23 | 0.29 | 2.72 × 10−20 |
| Akt2 | −0.23 | 5.43 × 10−5 | −0.25 | 0.000172 |
| Akt3 | 0.35 | 3.51 × 10−34 | 0.66 | 4.74 × 10−131 |
| Pik3ca | 0.28 | 0.00229 | 0.36 | 5.13 × 10−5 |
| Pik3r2 | 0.25 | 4.88 × 10−7 | 0.44 | 5.73 × 10−21 |
| Genes | CTRL vs. 5 µM Fold Change | CTRL vs. 5 µM q-Value | CTRL vs. 10 µM Fold Change | CTRL vs. 10 µM q-Value |
|---|---|---|---|---|
| Ctnnb1 | −0.22 | 5.50 × 10−16 | − | >0.05 |
| Lef1 | 0.51 | 3.70 × 10−14 | 0.75 | 3.12 × 10−31 |
| Chd8 | - | >0.05 | 0.19 | 9.56 × 10−10 |
| Ctbp2 | 0.14 | 1.54 × 10−2 | 0.35 | 1.41 × 10−11 |
| Tle3 | 0.35 | 2.22 × 10−23 | 0.51 | 2.26 × 10−51 |
| Neurod1 | - | >0.05 | −0.52 | 9.96 × 10−3 |
| Genes | CTRL vs. 5 µM Fold Change | CTRL vs. 5 µM q-Value | CTRL vs. 10 µM Fold Change | CTRL vs. 10 µM q-Value |
|---|---|---|---|---|
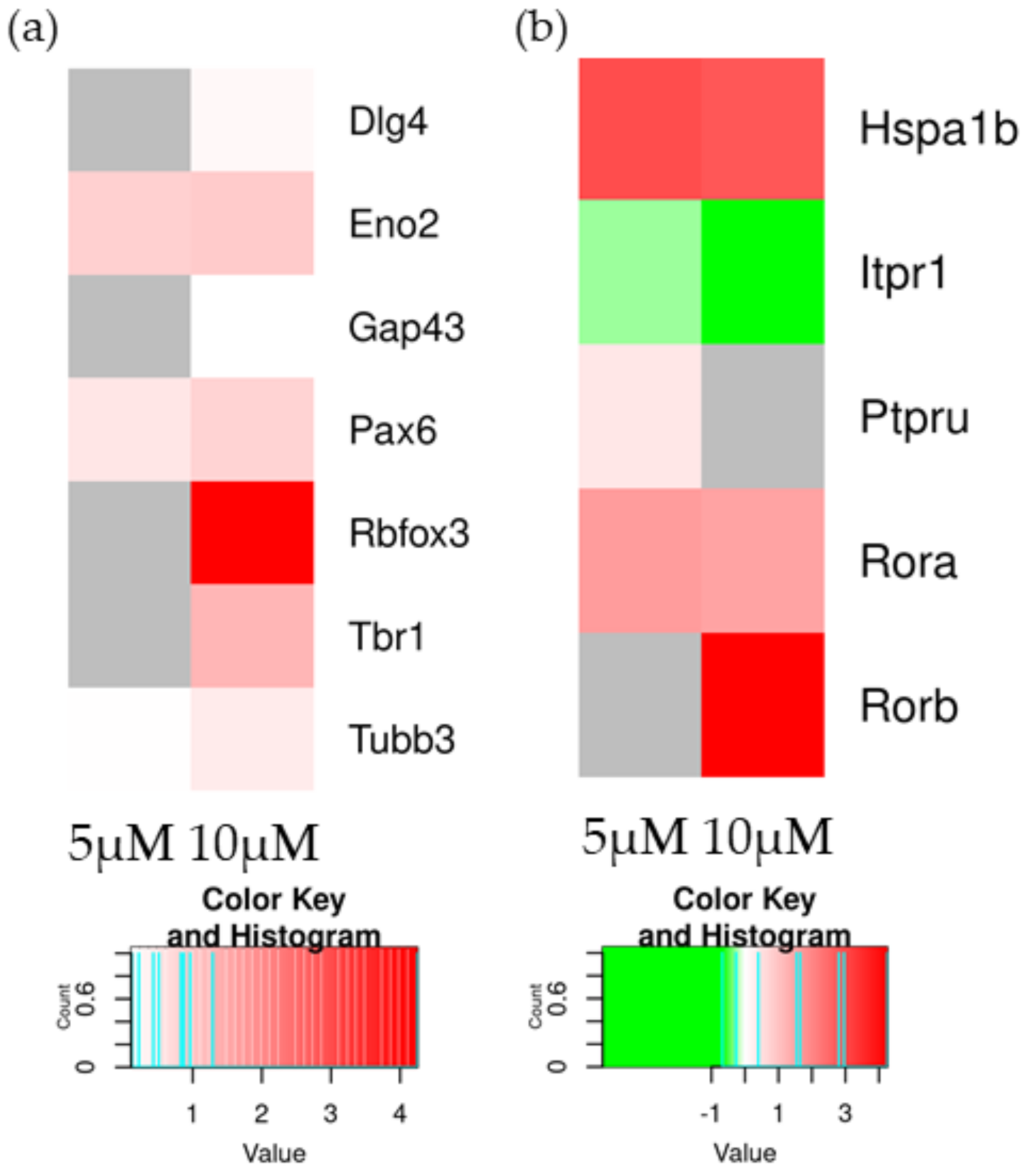
| Rbfox3 | - | >0.05 | 4.25 | 1.17 × 10−2 |
| Tubb3 | 0.12 | 1.23 × 10−44 | 0.43 | 0 |
| Gap43 | - | >0.05 | 0.11 | 2.06 × 10−2 |
| Pax6 | 0.51 | 7.14 × 10−14 | 0.83 | 2.64 × 10−38 |
| Dlg4 | - | >0.05 | 0.22 | 1.41 × 10−3 |
| Tbr1 | - | >0.05 | 1.29 | 4.94 × 10−2 |
| Eno2 | 0.87 | 1.57 × 10−13 | 0.96 | 1.94 × 10−16 |
| Genes | CTRL vs. 5 µM Fold Change | CTRL vs. 5 µM q-Value | CTRL vs. 10 µM Fold Change | CTRL vs. 10 µM q-Value |
|---|---|---|---|---|
| Hspa1b | 2.96 | 1.50 × 10−02 | 2.80 | 1.91 × 10−02 |
| Rorb | - | >0.05 | 4.25 | 1.17 × 10−02 |
| Itpr1 | −0.27 | 3.52 × 10−02 | −0.68 | 5.51 × 10−08 |
| Ptpru | 0.40 | 3.77 × 10−03 | − | >0.05 |
| Rora | 1.65 | 1.34 × 10−03 | 1.54 | 2.53 × 10−03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blando, S.; Raffaele, I.; Chiricosta, L.; Valeri, A.; Gugliandolo, A.; Silvestro, S.; Pollastro, F.; Mazzon, E. Cannabidiol Promotes Neuronal Differentiation Using Akt and Erk Pathways Triggered by Cb1 Signaling. Molecules 2022, 27, 5644. https://doi.org/10.3390/molecules27175644
Blando S, Raffaele I, Chiricosta L, Valeri A, Gugliandolo A, Silvestro S, Pollastro F, Mazzon E. Cannabidiol Promotes Neuronal Differentiation Using Akt and Erk Pathways Triggered by Cb1 Signaling. Molecules. 2022; 27(17):5644. https://doi.org/10.3390/molecules27175644
Chicago/Turabian StyleBlando, Santino, Ivana Raffaele, Luigi Chiricosta, Andrea Valeri, Agnese Gugliandolo, Serena Silvestro, Federica Pollastro, and Emanuela Mazzon. 2022. "Cannabidiol Promotes Neuronal Differentiation Using Akt and Erk Pathways Triggered by Cb1 Signaling" Molecules 27, no. 17: 5644. https://doi.org/10.3390/molecules27175644