
Cleistocalyx nervosum var. paniala Berry Promotes Antioxidant Response and Suppresses Glutamate-Induced Cell Death via SIRT1/Nrf2 Survival Pathway in Hippocampal HT22 Neuronal Cells

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Antioxidant Capabilities and Resveratrol Content in CNP Extract
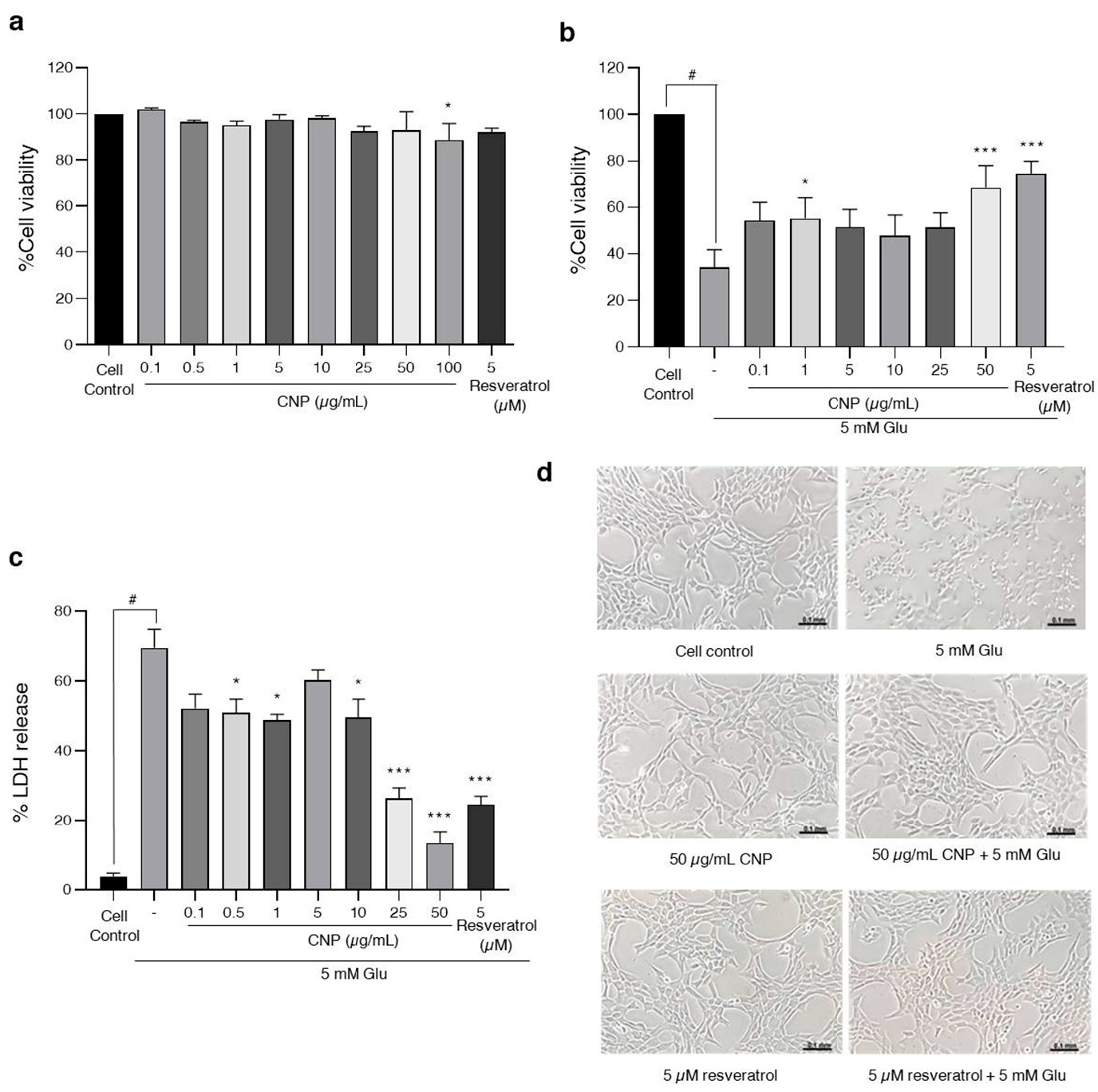
2.2. Cytotoxic Effects of CNP Extract on Viability of Cells in an Experiment Model of Glutamate-Induced Toxicity in HT22 Cells
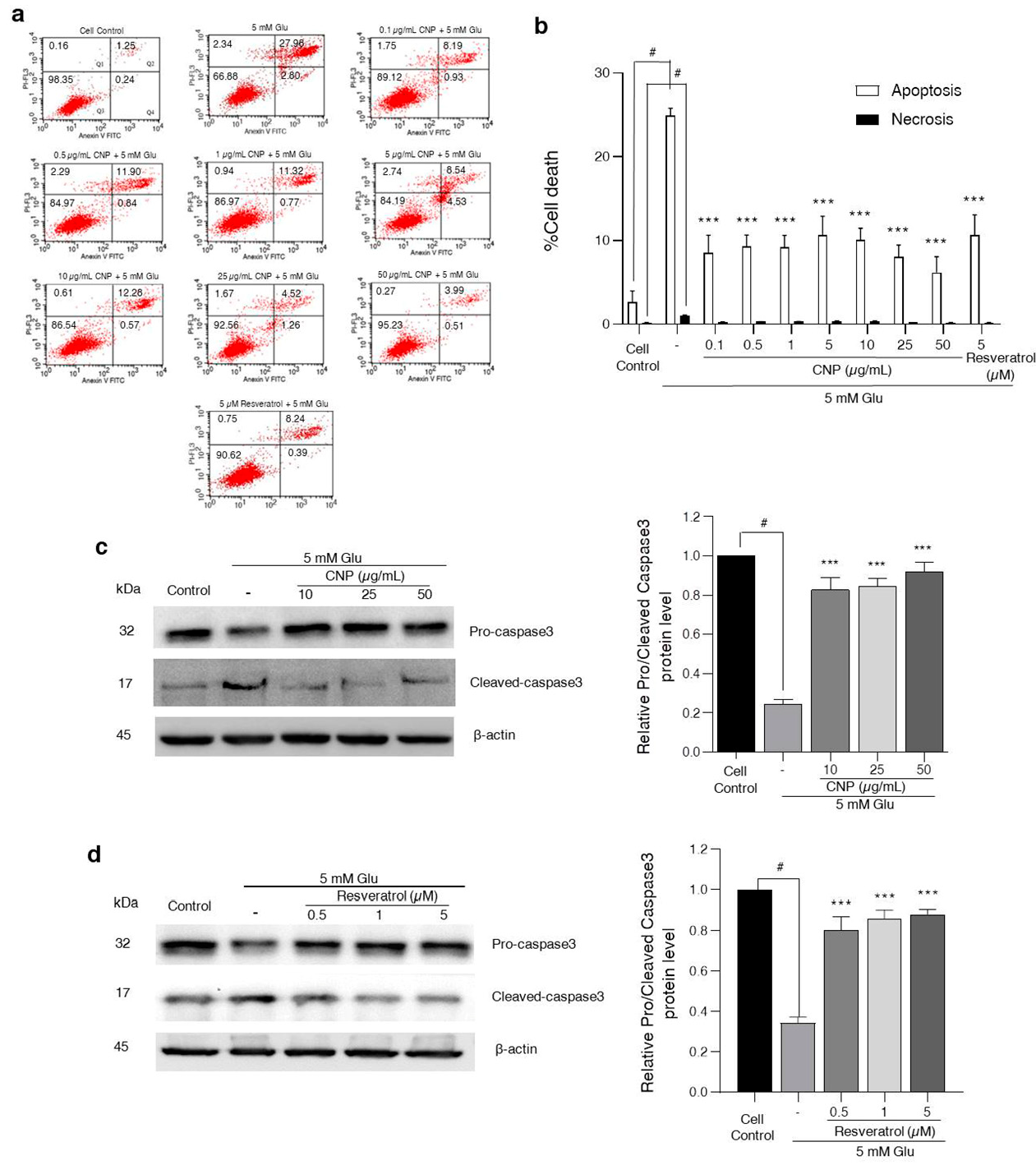
2.3. Effect of CNP Extract on Glutamate-Induced Apoptosis and Caspase-3 Protein Expression in HT22 Cells
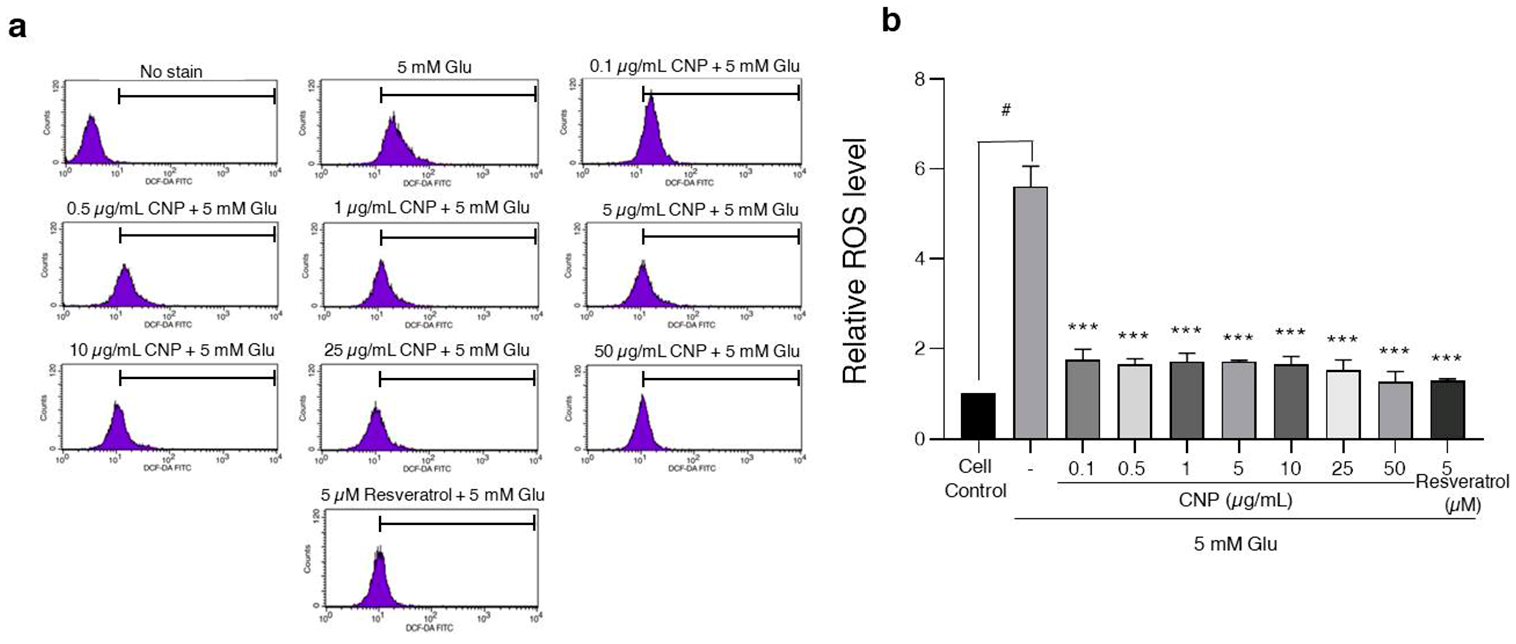
2.4. Inhibition of Glutamate-Induced Intracellular ROS Generation via CNP Extract Treatment of HT22 Cells
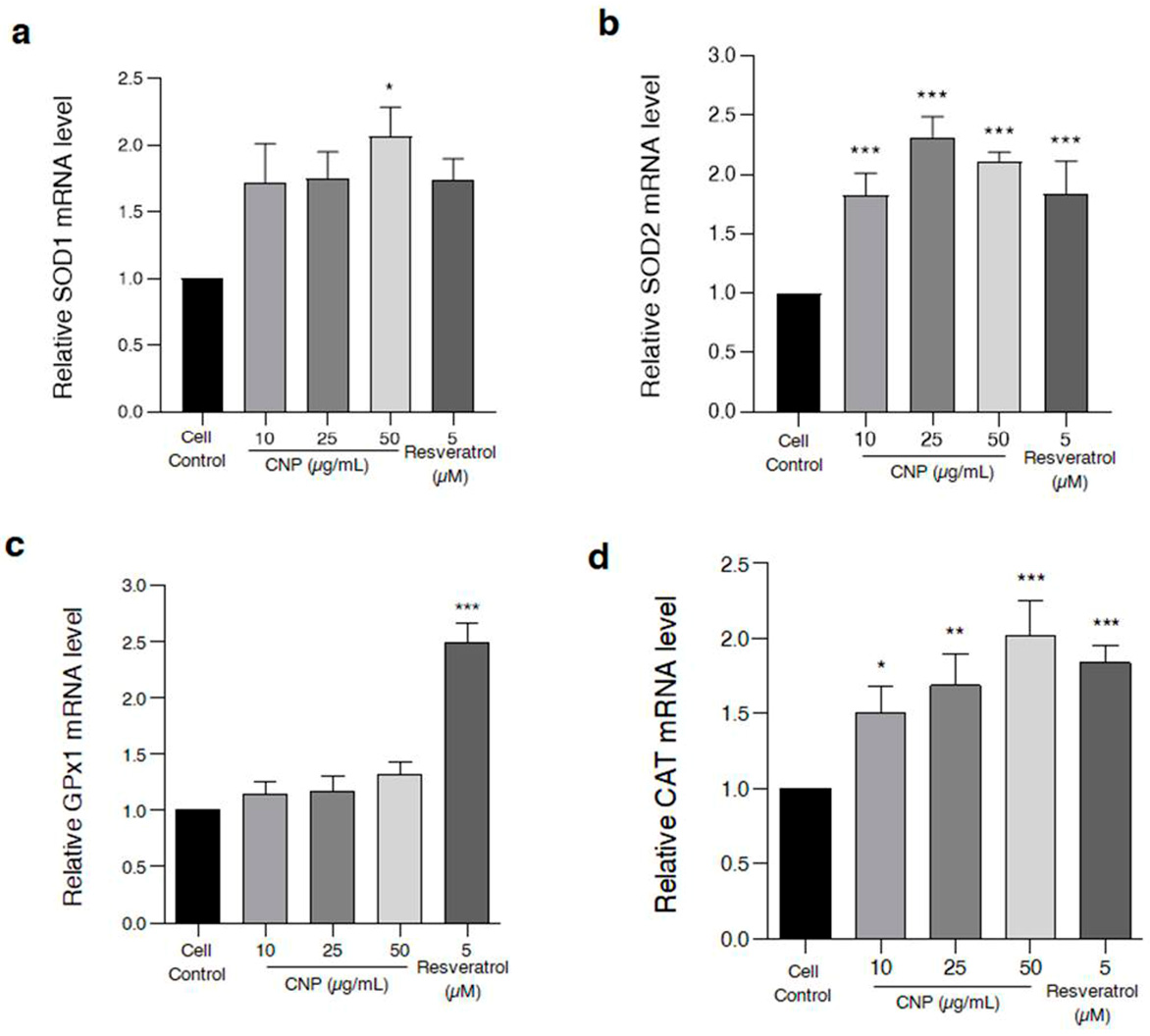
2.5. Effect of CNP Extract on the Cellular Antioxidant Enzymes in HT22 Cells
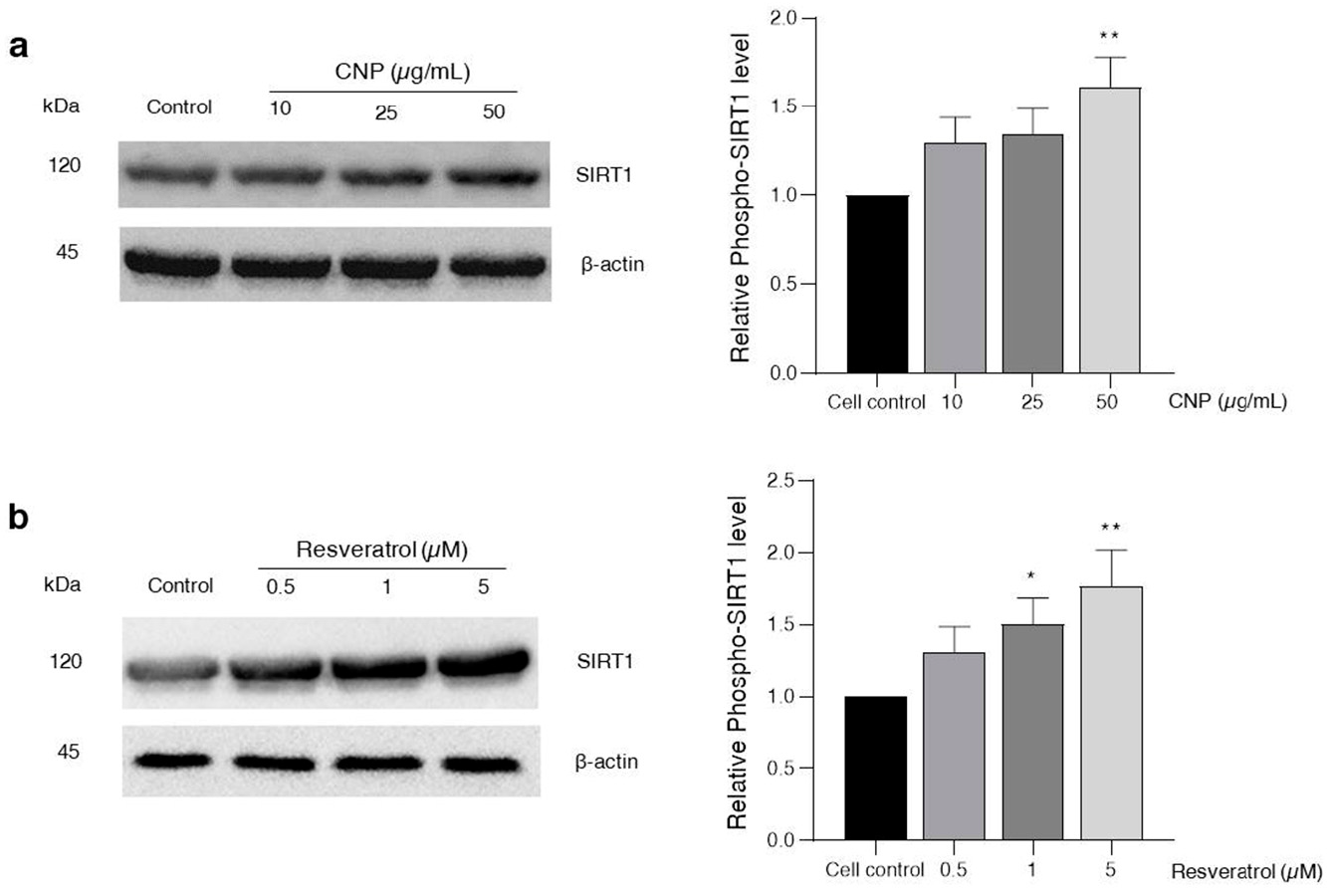
2.6. Effect of CNP Extract on SIRT1 Survival Protein Expression
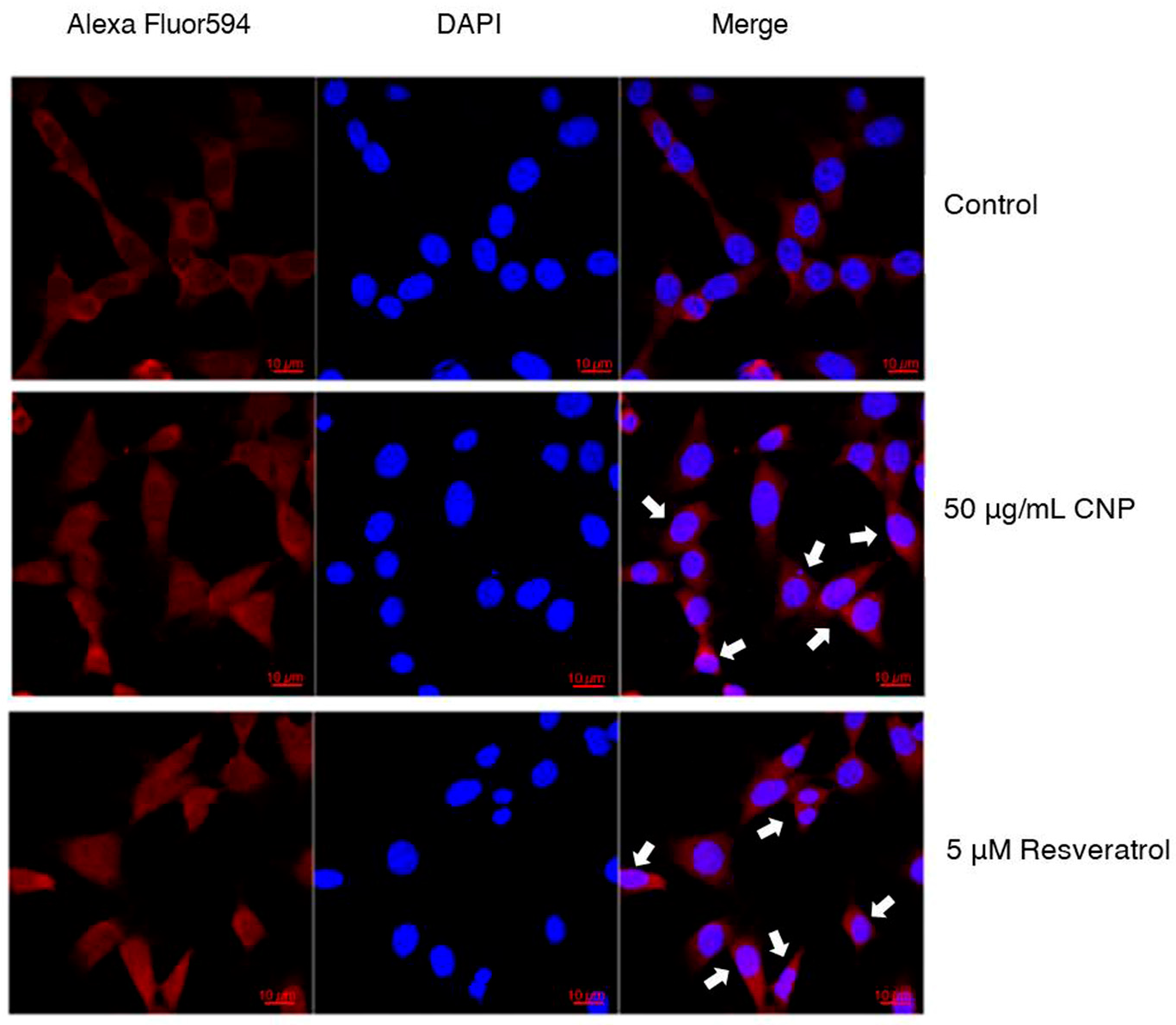
2.7. Effect of CNP Extract on Nrf2 Protein Translocation
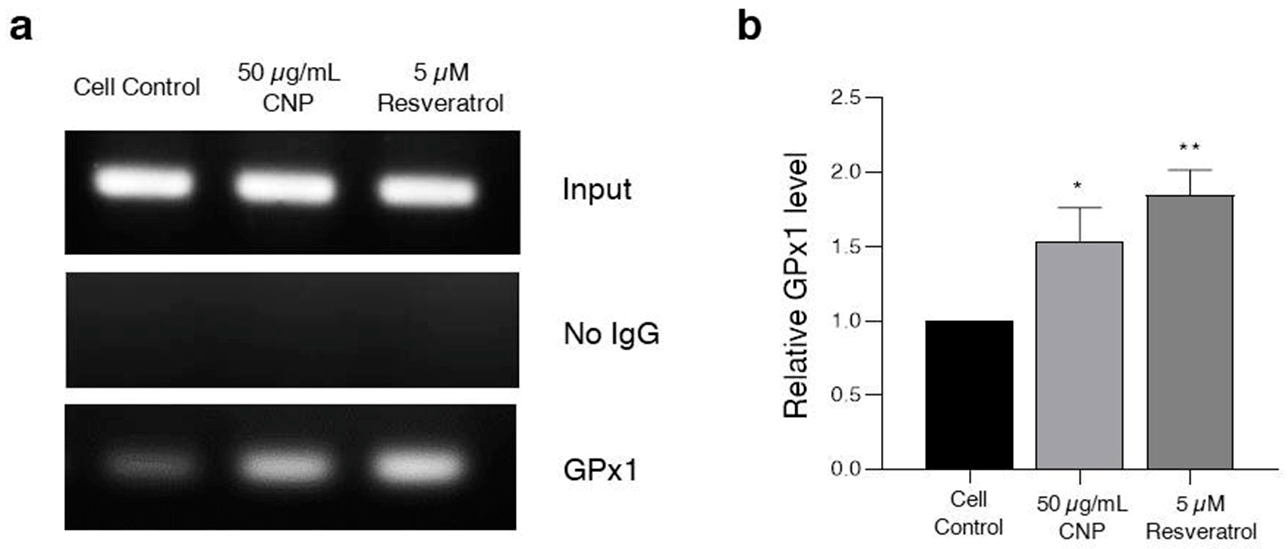
2.8. Effect of CNP Extract on the Ability to Bind the Antioxidant Response Element (ARE) Promoter Element
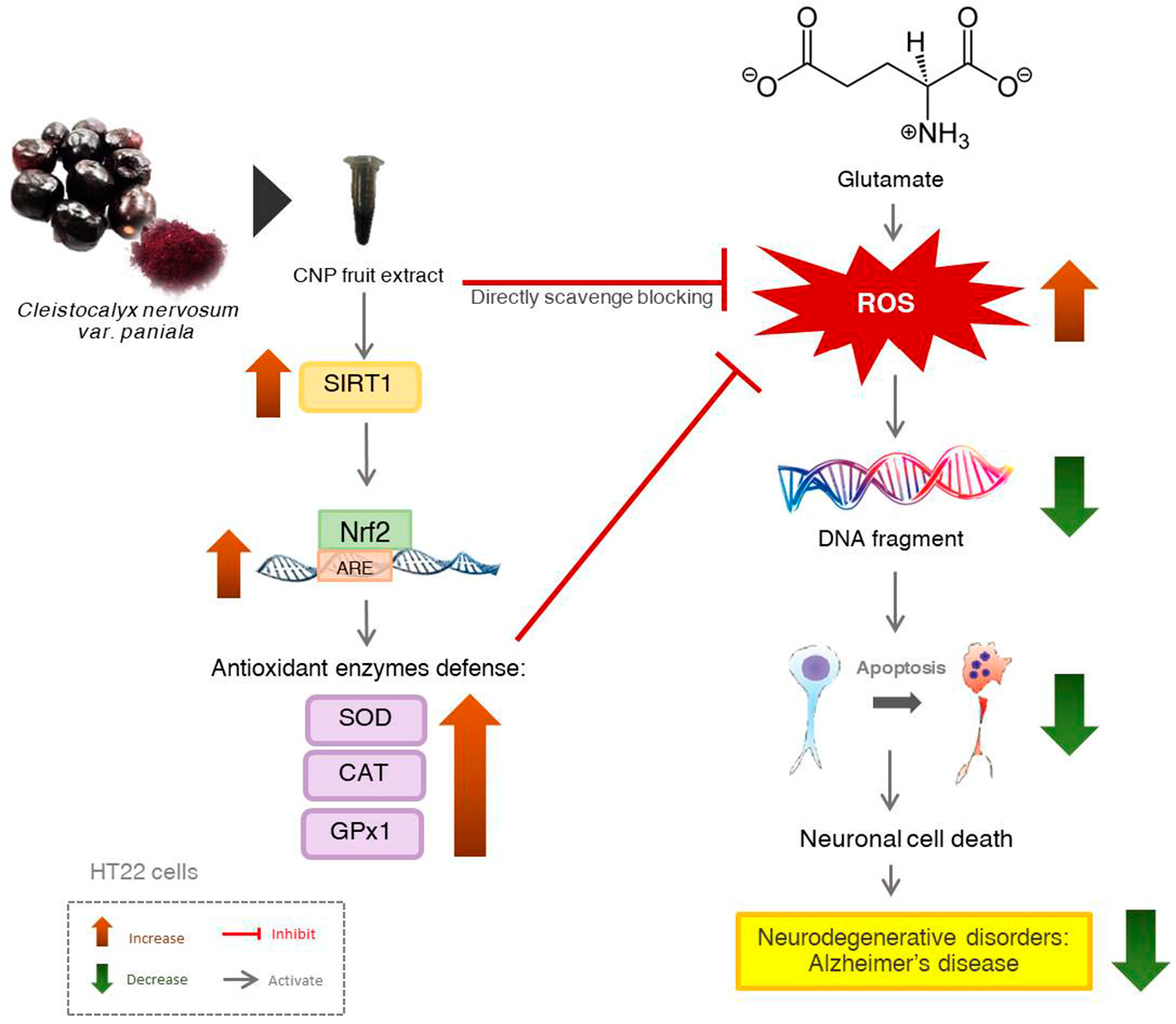
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. CNP Collection and Extraction
4.3. The 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Assay
4.4. The 2,2’-Azinobis-(3-Ethylbenzothiazoline-6-Sulfonic Acid) (ABTS) Assay
4.5. Total Phenolic Content
4.6. Total Flavonoid Content
4.7. Determination of Resveratrol in CNP Extract by HPLC Method
4.8. Cell Culture
4.9. Cell Viability Assay
4.10. Lactase Dehydrogenase (LDH) Assay
4.11. ROS Detection Assay
4.12. Apoptosis Assay
4.13. Gene Expression Analysis by Quantitative Real-Time PCR Analysis (qRT-PCR)
4.14. Western Blot Analysis
4.15. Immunofluorescence Microscopy Assay
4.16. Chromatin Immunoprecipitation (ChIP) Assay
4.17. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Savelieff, M.G.; Nam, G.; Kang, J.; Lee, H.J.; Lee, M.; Lim, M.H. Development of multifunctional molecules as potential therapeutic candidates for Alzheimer’s disease, Parkinson’s disease, and amyotrophic lateral sclerosis in the last decade. Chem. Rev. 2018, 119, 1221–1322. [Google Scholar] [CrossRef]
- Cimler, R.; Maresova, P.; Kuhnova, J.; Kuca, K. Predictions of Alzheimer’s disease treatment and care costs in European countries. PLoS ONE 2019, 14, e0210958. [Google Scholar] [CrossRef] [PubMed]
- Maziuk, B.; Ballance, H.I.; Wolozin, B. Dysregulation of RNA binding protein aggregation in neurodegenerative disorders. Front. Mol. Neurosci. 2017, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Atluri, V.; Kaushik, A.; Yndart, A.; Nair, M. Alzheimer’s disease: Pathogenesis, diagnostics, and therapeutics. Int. J. Nanomed. 2019, 14, 5541–5554. [Google Scholar] [CrossRef] [PubMed]
- Perluigi, M.; Di Domenico, F.; Barone, E.; Butterfield, D.A. mTOR in Alzheimer disease and its earlier stages: Links to oxidative damage in the progression of this dementing disorder. Free Radic. Biol. Med. 2021, 169, 382–396. [Google Scholar] [CrossRef]
- Takahashi, R.H.; Nagao, T.; Gouras, G.K. Plaque formation and the intraneuronal accumulation of β-amyloid in Alzheimer’s disease. Pathol. Int. 2017, 67, 185–193. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Boyd-Kimball, D. Oxidative stress, amyloid-β peptide, and altered key molecular pathways in the pathogenesis and progression of Alzheimer’s disease. J. Alzheimer’s Dis. 2018, 62, 1345–1367. [Google Scholar] [CrossRef]
- Mett, J. The Impact of Medium Chain and Polyunsaturated ω-3-Fatty Acids on Amyloid-β Deposition, Oxidative Stress and Metabolic Dysfunction Associated with Alzheimer’s disease. Antioxidants 2021, 10, 1991. [Google Scholar] [CrossRef]
- Siano, G.; Varisco, M.; Scarlatti, A.; Caiazza, M.C.; Dunville, K.; Cremisi, F.; Cattaneo, A. Gene expression of disease-related genes in alzheimer’s disease is impaired by tau aggregation. J. Mol. Biol. 2020, 432, 166675. [Google Scholar] [CrossRef]
- Pagliusi, S.R.; Gerrard, P.; Abdallah, M.; Talabot, D.; Catsicas, S. Age-related changes in expression of AMPA-selective glutamate receptor subunits: Is calcium-permeability altered in hippocampal neurons? Neuroscience 1994, 61, 429–433. [Google Scholar] [CrossRef]
- Cenini, G.; Lloret, A.; Cascella, R. Oxidative stress in neurodegenerative diseases: From a mitochondrial point of view. Oxid. Med. Cell. Longev. 2019, 2019, 2105607. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.U.; Wali, A.F.; Ahmad, A.; Shakeel, S.; Rasool, S.; Ali, R.; Khan, R. Neuroprotective strategies for neurological disorders by natural products: An update. Curr. Neuropharmacol. 2019, 17, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Jansom, C.; Bhamarapravati, S.; Itharat, A. Major anthocyanin from ripe berries of Cleistocalyx nervosum var. paniala. Thammasat. Med. J. 2008, 8, 365. [Google Scholar]
- Patthamakanokporn, O.; Puwastien, P.; Nitithamyong, A.; Sirichakwal, P.P. Changes of antioxidant activity and total phenolic compounds during storage of selected fruits. J. Food Compost. Anal. 2008, 21, 241–248. [Google Scholar] [CrossRef]
- Sukprasansap, M.; Chanvorachote, P.; Tencomnao, T. Cleistocalyx nervosum var. paniala berry fruit protects neurotoxicity against endoplasmic reticulum stress-induced apoptosis. Food Chem. Toxicol. 2017, 103, 279–288. [Google Scholar] [CrossRef]
- Charoensin, S.; Taya, S.; Wongpornchai, S.; Wongpoomchai, R. Assessment of genotoxicity and antigenotoxicity of an aqueous extract of Cleistocalyx nervosum var. paniala in in vitro and in vivo models. Interdiscip. Toxicol. 2012, 5, 201. [Google Scholar] [CrossRef]
- Inboot, W.; Taya, S.; Chailungka, A.; Meepowpan, P.; Wongpoomchai, R. Genotoxicity and antigenotoxicity of the methanol extract of Cleistocalyx nervosum var. paniala seed using a Salmonella mutation assay and rat liver micronucleus tests. Mol. Cell. Toxicol. 2012, 8, 19–24. [Google Scholar] [CrossRef]
- Taya, S.; Punvittayagul, C.; Inboot, W.; Fukushima, S.; Wongpoomchai, R. Cleistocalyx nervosum extract ameliorates chemical-induced oxidative stress in early stages of rat hepatocarcinogenesis. Asian Pac. J. Cancer Prev. 2014, 15, 2825–2830. [Google Scholar] [CrossRef]
- Prasanth, M.I.; Brimson, J.M.; Chuchawankul, S.; Sukprasansap, M.; Tencomnao, T. Antiaging, stress resistance, and neuroprotective efficacies of Cleistocalyx nervosum var. paniala fruit extracts using Caenorhabditis elegans model. Oxid. Med. Cell. Longev. 2019, 2019, 7024785. [Google Scholar] [CrossRef]
- Manosroi, J.; Chankhampan, C.; Kumguan, K.; Manosroi, W.; Manosroi, A. In vitro anti-aging activities of extracts from leaves of Ma Kiang (Cleistocalyx nervosum var. paniala). Pharm. Biol. 2015, 53, 862–869. [Google Scholar] [CrossRef]
- Sriwanthana, B.; Treesangsri, W.; Boriboontrakul, B.; Niumsakul, S.; Chavalittumrong, P. In vitro effects of Thai medicinal plants on human lymphocyte activity. In Vitro 2007, 29, 1. [Google Scholar]
- Pallàs, M.; Porquet, D.; Vicente, A.; Sanfeliu, C. Resveratrol: New avenues for a natural compound in neuroprotection. Curr. Pharm. Des. 2013, 19, 6726–6731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijekoon, C.; Netticadan, T.; Siow, Y.L.; Sabra, A.; Yu, L.; Raj, P.; Prashar, P. Associations among Bioactive Molecules, Antioxidant Activity and Resveratrol Production in Vitis vinifera Fruits of North America. Molecules 2022, 27, 336. [Google Scholar] [CrossRef] [PubMed]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-inflammatory action and mechanisms of resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef] [PubMed]
- Nasiri, M.; Ahmadizad, S.; Hedayati, M.; Zarekar, T.; Seydyousefi, M.; Faghfoori, Z. Trans-resveratrol supplement lowers lipid peroxidation responses of exercise in male Wistar rats. Int. J. Vitam. Nutr. Res. 2021, 91, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Yan, Y.; He, X.Y.; Yang, H.; Liang, B.; Wang, J.; Yu, H. Effects of resveratrol on the mechanisms of antioxidants and estrogen in Alzheimer’s disease. BioMed Res. Int. 2019, 20, 8983752. [Google Scholar] [CrossRef]
- Quarta, A.; Gaballo, A.; Pradhan, B.; Patra, S.; Jena, M.; Ragusa, A. Beneficial oxidative stress-related trans-resveratrol effects in the treatment and prevention of breast cancer. Appl. Sci. 2021, 11, 11041. [Google Scholar] [CrossRef]
- Ahmed, R.F.; El Awdan, S.A.; Jaleel, G.A.A.; Saleh, D.O.; Farid, O.A.A. Correlation between brain neurotransmitters and insulin sensitivity: Neuro-preservative role of resveratrol against high fat, high fructose-induced insulin resistance. J. Appl. Pharm. Sci. 2020, 10, 26–36. [Google Scholar]
- Bastianetto, S.; Ménard, C.; Quirion, R. Neuroprotective action of resveratrol. Biochim. Biophys. Acta-Mol. Basis Dis. 2015, 1852, 1195–1201. [Google Scholar] [CrossRef]
- Gerszon, J.; Rodacka, A.; Puchała, M. Antioxidant properties of resveratrol and its protective effects in neurodegenerative diseases. Adv. Cell Biol. 2014, 4, 97–117. [Google Scholar] [CrossRef]
- Rege, S.D.; Geetha, T.; Griffin, G.D.; Broderick, T.L.; Babu, J.R. Neuroprotective effects of resveratrol in Alzheimer disease pathology. Front. Aging Neurosci. 2014, 6, 218. [Google Scholar] [CrossRef] [PubMed]
- Xue, F.; Huang, J.-W.; Ding, P.-Y.; Zang, H.-G.; Kou, Z.-J.; Li, T.; Fan, J.; Peng, Z.-W.; Yan, W.-J. Nrf2/antioxidant defense pathway is involved in the neuroprotective effects of Sirt1 against focal cerebral ischemia in rats after hyperbaric oxygen preconditioning. Behav. Brain Res. 2016, 309, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Sun, Z.; Han, X.; Li, S.; Jiang, X.; Chen, S.; Lu, H. Neuroprotective effect of resveratrol via activation of Sirt1 signaling in a rat model of combined diabetes and Alzheimer’s disease. Front. Neurosci. 2020, 13, 1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukprasansap, M.; Chanvorachote, P.; Tencomnao, T. Cyanidin-3-glucoside activates Nrf2-antioxidant response element and protects against glutamate-induced oxidative and endoplasmic reticulum stress in HT22 hippocampal neuronal cells. BMC Complement. Altern. Med. 2020, 20, 46. [Google Scholar] [CrossRef]
- Huo, Y.; Yang, D.; Lai, K.; Tu, J.; Zhu, Y.; Ding, W.; Yang, S. Antioxidant Effects of Resveratrol in Intervertebral Disk. J. Investig. Surg. 2022, 35, 1135–1144. [Google Scholar] [CrossRef]
- Ma, X.; Deng, D.; Chen, W. Inhibitors and Activators of SOD, GSH-Px, and CAT. Enzym. Inhib. Act. 2017, 29, 207–224. [Google Scholar]
- Ighodaro, O.; Akinloye, O. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Sacitharan, P.K.; Bou-Gharios, G.; Edwards, J.R. SIRT1 directly activates autophagy in human chondrocytes. Cell Death Discov. 2020, 6, 41. [Google Scholar] [CrossRef]
- Albani, D.; Polito, L.; Batelli, S.; De Mauro, S.; Fracasso, C.; Martelli, G.; Colombo, L.; Manzoni, C.; Salmona, M.; Caccia, S. The SIRT1 activator resveratrol protects SK-N-BE cells from oxidative stress and against toxicity caused by α-synuclein or amyloid-β (1-42) peptide. J. Neurochem. 2009, 110, 1445–1456. [Google Scholar] [CrossRef]
- Da Cunha, M.D.S.B.; Arruda, S.F. Tucum-do-Cerrado (Bactris setosa Mart.) may promote anti-aging effect by upregulating SIRT1-Nrf2 pathway and attenuating oxidative stress and inflammation. Nutrients 2017, 9, 1243. [Google Scholar] [CrossRef]
- Serafini, M.M.; Catanzaro, M.; Fagiani, F.; Simoni, E.; Caporaso, R.; Dacrema, M.; Lanni, C. Modulation of Keap1/Nrf2/ARE signaling pathway by curcuma-and garlic-derived hybrids. Front. Pharmacol. 2020, 10, 1597. [Google Scholar] [CrossRef] [PubMed]
- Kulawiak, B.; Szewczyk, A. Glutamate-induced cell death in HT22 mouse hippocampal cells is attenuated by paxilline, a BK channel inhibitor. Mitochondrion 2012, 12, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Weiner, M.W. Further insights into Alzheimer disease pathogenesis. Nat. Rev. Neurol. 2013, 9, 65–66. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Panahi, Y.; Javadi, B.; Sahebkar, A. The underlying role of oxidative stress in neurodegeneration: A mechanistic review. CNS Neurol. Disord. Drug Targets 2018, 17, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Indo, H.P.; Davidson, M.; Yen, H.-C.; Suenaga, S.; Tomita, K.; Nishii, T.; Higuchi, M.; Koga, Y.; Ozawa, T.; Majima, H.J. Evidence of ROS generation by mitochondria in cells with impaired electron transport chain and mitochondrial DNA damage. Mitochondrion 2007, 7, 106–118. [Google Scholar] [CrossRef]
- Breyer, A.; Elstner, M.; Gillessen, T.; Weiser, D.; Elstner, E. Glutamate-induced cell death in neuronal HT22 cells is attenuated by extracts from St. John’s wort (Hypericum perforatum L.). Phytomedicine 2007, 14, 250–255. [Google Scholar] [CrossRef]
- Byrnes, K.R.; Loane, D.J.; Faden, A.I. Metabotropic glutamate receptors as targets for multipotential treatment of neurological disorders. Neurotherapeutics 2009, 6, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Weiner, M.W.; Veitch, D.P.; Aisen, P.S.; Beckett, L.A.; Cairns, N.J.; Green, R.C.; Harvey, D.; Jack, C.R.; Jagust, W.; Liu, E. The Alzheimer’s disease neuroimaging initiative: A review of papers published since its inception. Alzheimer’s Dement. 2013, 9, 111–194. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Daly, I.; Zhang, Y.; Wang, X.; Cichocki, A. An optimized ERP brain–computer interface based on facial expression changes. J. Neural Eng. 2014, 11, 036004. [Google Scholar] [CrossRef] [PubMed]
- Tobaben, S.; Grohm, J.; Seiler, A.; Conrad, M.; Plesnila, N.; Culmsee, C. Bid-mediated mitochondrial damage is a key mechanism in glutamate-induced oxidative stress and AIF-dependent cell death in immortalized HT-22 hippocampal neurons. Cell Death Differ. 2011, 18, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Bensalem, J.; Dal-Pan, A.; Gillard, E.; Calon, F.; Pallet, V. Protective effects of berry polyphenols against age-related cognitive impairment. Nutr. Aging 2015, 3, 89–106. [Google Scholar] [CrossRef]
- Duangjan, C.; Rangsinth, P.; Zhang, S.; Gu, X.; Wink, M.; Tencomnao, T. Vitis vinifera leaf extract protects against glutamate-induced oxidative toxicity in HT22 hippocampal neuronal cells and increases stress resistance properties in Caenorhabditis elegans. Front. Nutr. 2021, 8, 250. [Google Scholar]
- Pluta, R. Alzheimer’s disease connected genes in the post-ischemic hippocampus and temporal cortex. Genes 2022, 13, 1059. [Google Scholar] [CrossRef] [PubMed]
- Engwa, G.A. Free radicals and the role of plant phytochemicals as antioxidants against oxidative stress-related diseases. In Phytochemicals: Source of Antioxidants and Role in Disease Prevention. BoD–Books on Demand; Intechopen: London, UK, 2018; Volume 7, pp. 49–74. [Google Scholar]
- Sharma, G.N.; Gupta, G.; Sharma, P. A comprehensive review of free radicals, antioxidants, and their relationship with human ailments. Alcohol Crit. Rev. Eukaryot. Gene Expr. 2018, 28, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Si, H.; Liu, D. Dietary antiaging phytochemicals and mechanisms associated with prolonged survival. J. Nutr. Biochem. 2014, 25, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Grabska-Kobylecka, I.; Kaczmarek-Bak, J.; Figlus, M.; Prymont-Przyminska, A.; Zwolinska, A.; Sarniak, A.; Nowak, D. The presence of caffeic acid in cerebrospinal fluid: Evidence that dietary polyphenols can cross the blood-brain barrier in humans. Nutrients 2020, 12, 1531. [Google Scholar] [CrossRef]
- De Lange, E.; Vd Berg, D.; Bellanti, F.; Voskuyl, R.; Syvänen, S. P-glycoprotein protein expression versus functionality at the blood-brain barrier using immunohistochemistry, microdialysis and mathematical modeling. Eur. J. Pharm. Sci. 2018, 124, 61–70. [Google Scholar] [CrossRef]
- Katila, N.; Duwa, R.; Bhurtel, S.; Khanal, S.; Maharjan, S.; Jeong, J.H.; Yook, S. Enhancement of blood–brain barrier penetration and the neuroprotective effect of resveratrol. J. Control. Release 2022, 346, 1–19. [Google Scholar] [CrossRef]
- Robb, E.L.; Stuart, J.A. Trans-Resveratrol as a neuroprotectant. Molecules 2010, 15, 1196–1212. [Google Scholar] [CrossRef]
- Arbo, B.D.; André-Miral, C.; Nasre-Nasser, R.G.; Schimith, L.E.; Santos, M.G.; Costa-Silva, D.; Hort, M.A. Resveratrol derivatives as potential treatments for Alzheimer’s and Parkinson’s disease. Front. Aging Neurosci. 2020, 12, 103. [Google Scholar] [CrossRef]
- Kim, D.-W.; Kim, Y.-M.; Kang, S.-D.; Han, Y.-M.; Pae, H.-O. Effects of resveratrol and trans-3, 5, 4′-trimethoxystilbene on glutamate-induced cytotoxicity, heme oxygenase-1, and sirtuin 1 in HT22 neuronal cells. Biomol. Ther. 2012, 20, 306. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Yang, W.; Laaksonen, O.; Nylander, M.; Kallio, H.; Yang, B. Role of flavonols and proanthocyanidins in the sensory quality of sea buckthorn (Hippophaë rhamnoides L.) Berries. J. Agric. Food Chem. 2017, 65, 9871–9879. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, L.; Roriz-Cruz, M. Sirtuin 1 and Alzheimer’s disease: An up-to-date review. Neuropeptides 2018, 71, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Charoensin, S. Antioxidant and anticancer activities of Moringa oleifera leaves. J. Med. Plant Res. 2014, 8, 318–325. [Google Scholar]
- Chen, X.; Drew, J.; Berney, W.; Lei, W. Neuroprotective natural products for Alzheimer’s disease. Cells 2021, 10, 1309. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Boyd-Kimball, D. Redox proteomics and amyloid β-peptide: Insights into Alzheimer disease. J. Neurochem. 2019, 151, 459–487. [Google Scholar] [CrossRef] [Green Version]
- Essa, M.M.; Vijayan, R.K.; Castellano-Gonzalez, G.; Memon, M.A.; Braidy, N.; Guillemin, G.J. Neuroprotective effect of natural products against Alzheimer’s disease. Neurochem. Res. 2012, 37, 1829–1842. [Google Scholar] [CrossRef]
- Ren, Z.; He, H.; Zuo, Z.; Xu, Z.; Wei, Z.; Deng, J. The role of different SIRT1-mediated signaling pathways in toxic injury. Cell. Mol. Biol. Lett. 2019, 24, 36. [Google Scholar] [CrossRef]
- Xu, J.; Jackson, C.W.; Khoury, N.; Escobar, I.; Perez-Pinzon, M.A. Brain SIRT1 mediates metabolic homeostasis and neuroprotection. Front. Endocrinol. 2018, 9, 702. [Google Scholar] [CrossRef]
- Feng, X.; Liang, N.; Zhu, D.; Gao, Q.; Peng, L.; Dong, H.; Yue, Q.; Liu, H.; Bao, L.; Zhang, J. Resveratrol inhibits β-amyloid-induced neuronal apoptosis through regulation of SIRT1-ROCK1 signaling pathway. PLoS ONE 2013, 8, 59888. [Google Scholar] [CrossRef]
- Li, Z.; Han, X. Resveratrol alleviates early brain injury following subarachnoid hemorrhage: Possible involvement of the AMPK/SIRT1/autophagy signaling pathway. Biol. Chem. 2018, 399, 1339–1350. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Yamamoto, M. The molecular mechanisms regulating the KEAP1-NRF2 pathway. Mol. Cell. Biol. 2020, 40, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Guo, T.; Li, G.; Sun, S.; He, S.; Cheng, B.; Shi, B.; Shan, A. Dietary resveratrol improves antioxidant status of sows and piglets and regulates antioxidant gene expression in placenta by Keap1-Nrf2 pathway and Sirt1. J. Anim. Sci. Biotechnol. 2018, 9, 34. [Google Scholar] [CrossRef]
- Han, J.; Huang, Z.; Tang, S.; Lu, C.; Wan, H.; Zhou, J.; Li, Y.; Ming, T.; Wang, Z.J.; Su, X. The novel peptides ICRD and LCGEC screened from tuna roe show antioxidative activity via Keap1/Nrf2-ARE pathway regulation and gut microbiota modulation. Food Chem. 2020, 327, 127094. [Google Scholar] [CrossRef]
- Hannan, M.; Dash, R.; Sohag, A.A.M.; Haque, M.; Moon, I.S. Neuroprotection against oxidative stress: Phytochemicals targeting TrkB signaling and the Nrf2-ARE antioxidant system. Front. Mol. Neurosci. 2020, 13, 116. [Google Scholar] [CrossRef]
- Sarubbo, F.; Esteban, S.; Miralles, A.; Moranta, D. Effects of resveratrol and other polyphenols on Sirt1: Relevance to brain function during aging. Curr. Neuropharmacol. 2018, 16, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Prasanth, D.; Murahari, M.; Chandramohan, V.; Panda, S.P.; Atmakuri, L.R.; Guntupalli, C. In silico identification of potential inhibitors from Cinnamon against main protease and spike glycoprotein of SARS-CoV-2. J. Biomol. Struct. Dyn. 2020, 39, 4618–4632. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DPPH 1 | ABTS 1 | Total Phenolic Content 2 | Total Flavonoid Content 3 | Resveratrol Content 4 |
|---|---|---|---|---|
| 37.65 ± 4.72 | 56.82 ± 0.86 | 383.07 ± 1.83 | 43.71 ± 1.47 | 1.51 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nantacharoen, W.; Baek, S.J.; Plaingam, W.; Charoenkiatkul, S.; Tencomnao, T.; Sukprasansap, M. Cleistocalyx nervosum var. paniala Berry Promotes Antioxidant Response and Suppresses Glutamate-Induced Cell Death via SIRT1/Nrf2 Survival Pathway in Hippocampal HT22 Neuronal Cells. Molecules 2022, 27, 5813. https://doi.org/10.3390/molecules27185813
Nantacharoen W, Baek SJ, Plaingam W, Charoenkiatkul S, Tencomnao T, Sukprasansap M. Cleistocalyx nervosum var. paniala Berry Promotes Antioxidant Response and Suppresses Glutamate-Induced Cell Death via SIRT1/Nrf2 Survival Pathway in Hippocampal HT22 Neuronal Cells. Molecules. 2022; 27(18):5813. https://doi.org/10.3390/molecules27185813
Chicago/Turabian StyleNantacharoen, Wanchanok, Seung Joon Baek, Waluga Plaingam, Somsri Charoenkiatkul, Tewin Tencomnao, and Monruedee Sukprasansap. 2022. "Cleistocalyx nervosum var. paniala Berry Promotes Antioxidant Response and Suppresses Glutamate-Induced Cell Death via SIRT1/Nrf2 Survival Pathway in Hippocampal HT22 Neuronal Cells" Molecules 27, no. 18: 5813. https://doi.org/10.3390/molecules27185813