
A Metabolomic Analysis of Cirrhotic Ascites


Abstract
:
1. Introduction
2. Results
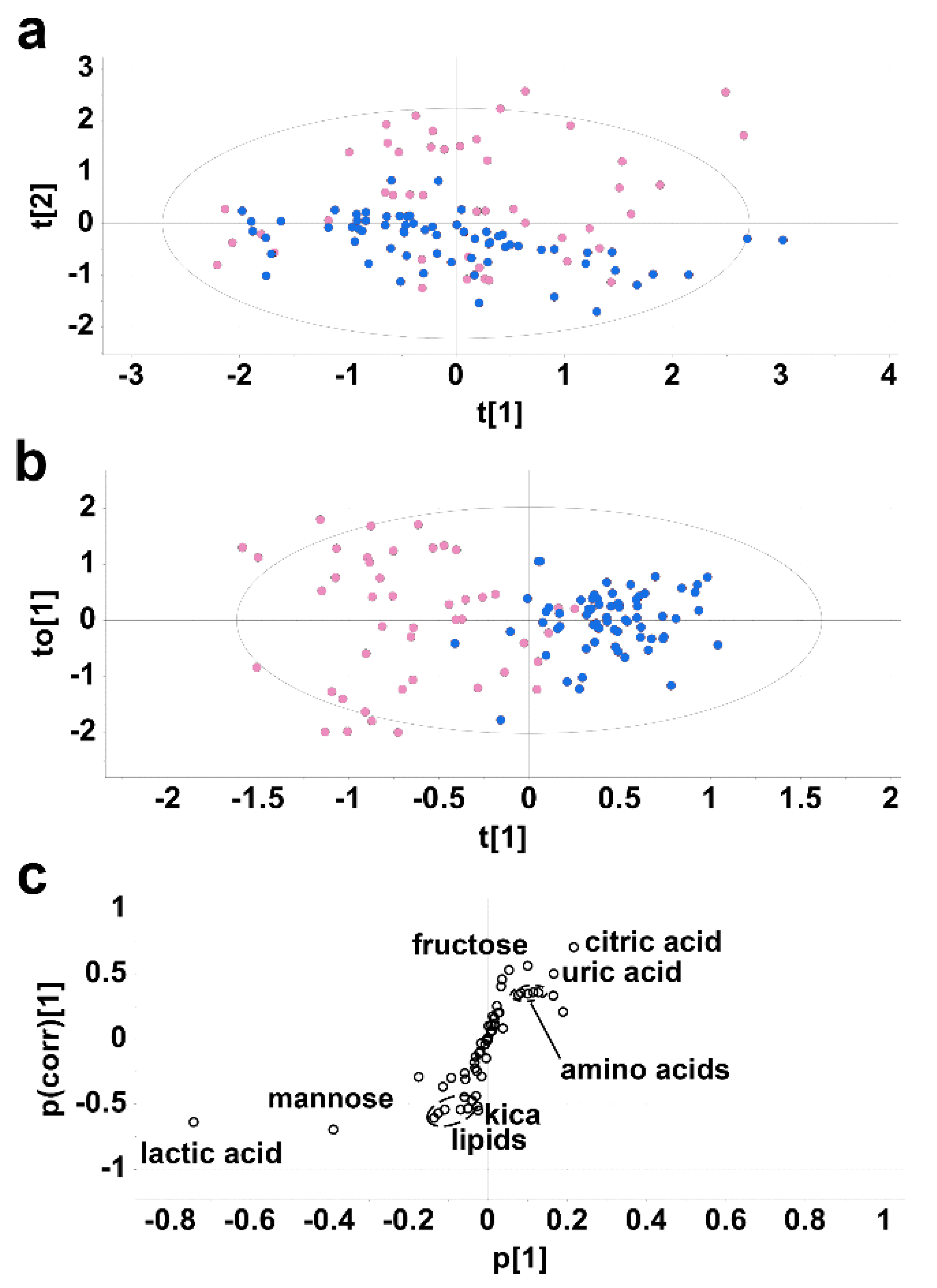
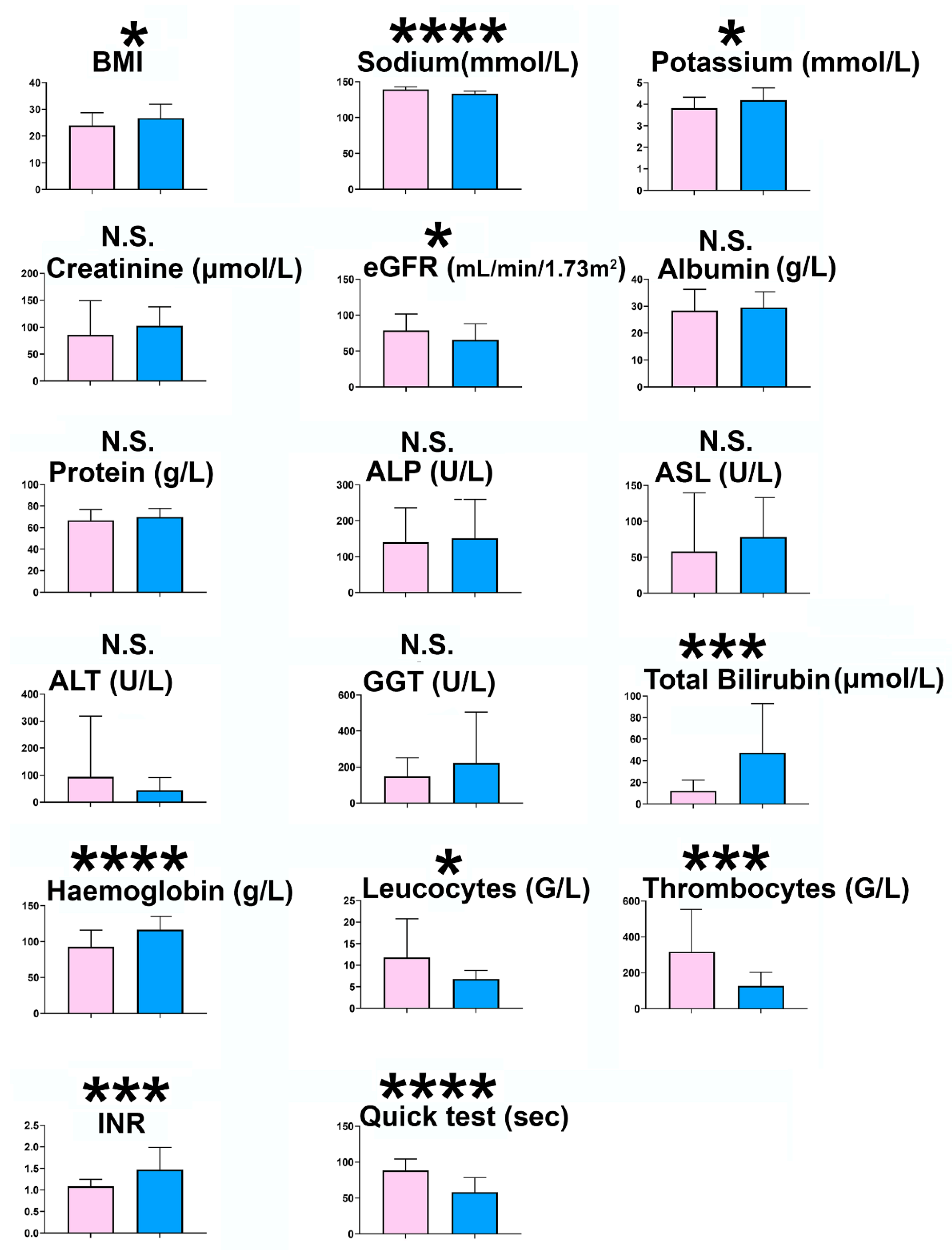
2.1. Experiment 1. Cirrhotic Ascites vs. Postoperative Peritoneal Drainage Fluid
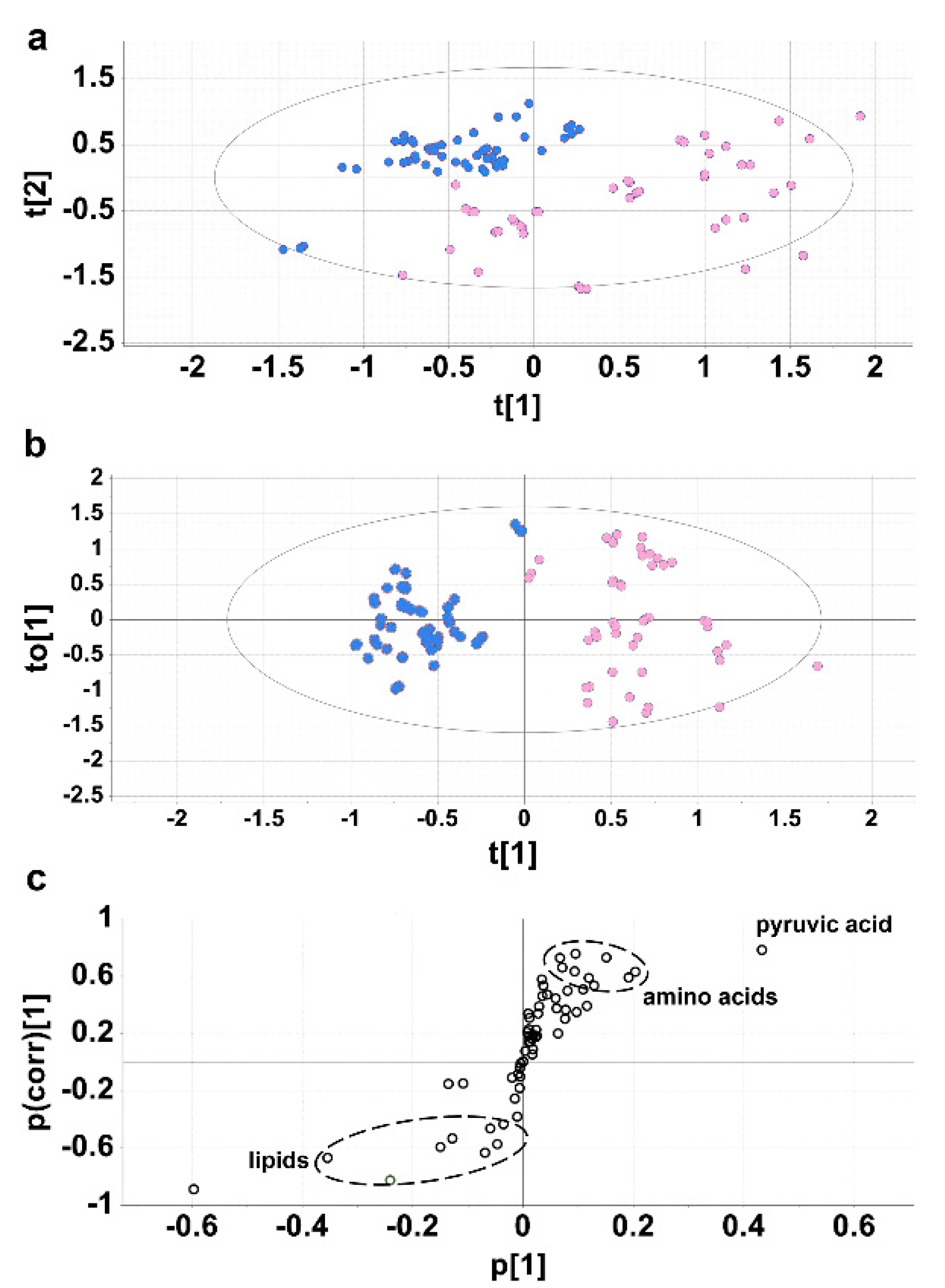
2.2. Experiment 2. Cirrhotic Ascites vs. Paired Plasmas
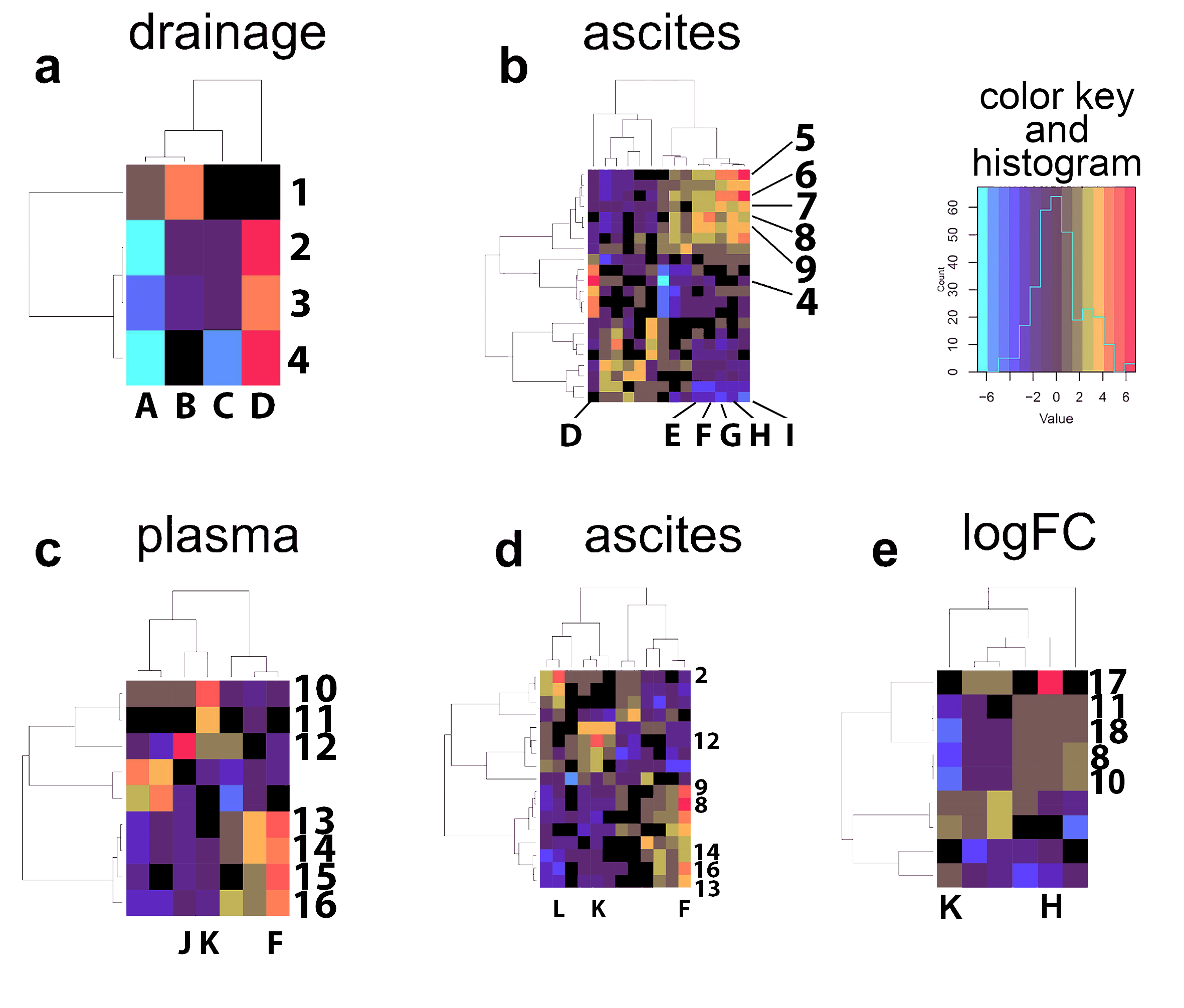
2.3. Bidirectional Clustering of Metabolomic Data Versus Clinical Data
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Gas Chromatography–Mass Spectrometry (GC-MS) Analysis
4.3. Statistics and Data Presentation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Mansour, D.; McPherson, S. Management of decompensated cirrhosis. Clin. Med. 2018, 18, s60–s65. [Google Scholar] [CrossRef] [PubMed]
- Runyon, B.A. Management of adult patients with ascites due to cirrhosis: An update. Hepatology 2009, 49, 2087–2107. [Google Scholar] [CrossRef] [PubMed]
- Spak, I. On the clinical value of chemical analysis of ascites. A study of the main proteins and some enzymes in ascites of differing etiology. Acta Chir. Scand. Suppl. 1960, (Suppl. 261), 1–128. [Google Scholar]
- Henderson, J.M.; Stein, S.F.; Kutner, M.; Wiles, M.-B.; Ansley, J.D.; Rudman, D. Analysis of Twenty-three Plasma Proteins in Ascites the Depletion of Fibrinogen and Plasminogen. Ann. Surg. 1980, 192, 738–742. [Google Scholar] [CrossRef]
- Jungst, D.; Xie, Y.; Gerbes, A.L. Pathophysiology of elevated ascites fluid cholesterol in malignant ascites: Increased ascites to serum relation of proteins and lipoproteins in patients with peritoneal carcinomatosis as compared to patients with cirrhosis of the liver. J. Hepatol. 1992, 14, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Hedenborg, G.; Jönsson, G.; Norman, A.; Wisén, O. Bile constituents in ascitic fluid. Scand. J. Clin. Lab. Investig. 1988, 48, 543–552. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Li, N.; Xing, F.; Zhao, J.; Yang, T.; Liu, C.; Feng, N. An herbal-compound-based combination therapy that relieves cirrhotic ascites by affecting the L-arginine/nitric oxide pathway: A metabolomics-based systematic study. J. Ethnopharmacol. 2019, 241, 112034. [Google Scholar] [CrossRef]
- Beyoğlu, D.; Zhou, Y.; Chen, C.; Idle, J.R. Mass isotopomer-guided decluttering of metabolomic data to visualize endogenous biomarkers of drug toxicity. Biochem. Pharmacol. 2018, 156, 491–500. [Google Scholar] [CrossRef]
- Sinclair, K.; Dudley, E. Metabolomics and Biomarker Discovery. In Advancements of Mass Spectrometry in Biomedical Research; Springer: Cham, Switzerland, 2019; Volume 1140, pp. 613–633. [Google Scholar] [CrossRef]
- Beyoğlu, D.; Idle, J.R. Metabolomic and Lipidomic Biomarkers for Premalignant Liver Disease Diagnosis and Therapy. Metabolites 2020, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyoğlu, D.; Idle, J.R. Metabolic Rewiring and the Characterization of Oncometabolites. Cancers 2021, 13, 2900. [Google Scholar] [CrossRef] [PubMed]
- Shender, V.O.; Pavlyukov, M.S.; Ziganshin, R.H.; Arapidi, G.P.; Kovalchuk, S.I.; Anikanov, N.; Altukhov, I.A.; Alexeev, D.; Butenko, I.; Shavarda, A.L.; et al. Proteome–Metabolome Profiling of Ovarian Cancer Ascites Reveals Novel Components Involved in Intercellular Communication. Mol. Cell. Proteom. 2014, 13, 3558–3571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharti, S.K.; Wildes, F.; Hung, C.-F.; Wu, T.C.; Bhujwalla, Z.M.; Penet, M.-F. Metabolomic characterization of experimental ovarian cancer ascitic fluid. Metabolomics 2017, 13, 113. [Google Scholar] [CrossRef]
- Zennaro, L.; Nicolè, L.; Vanzani, P.; Cappello, F.; Fassina, A. 1H-NMR spectroscopy metabonomics of reactive, ovarian carcinoma and hepatocellular carcinoma ascites. Pleura Peritoneum 2020, 5, 20200113. [Google Scholar] [CrossRef]
- Yang, T.; Zheng, X.; Xing, F.; Zhuo, H.; Liu, C. Serum Metabolomic Characteristics of Patients with Liver Cirrhotic Ascites. Integr. Med. Int. 2015, 1, 136–143. [Google Scholar] [CrossRef]
- Beyoğlu, D.; Imbeaud, S.; Maurhofer, O.; Bioulac-Sage, P.; Zucman-Rossi, J.; Dufour, J.-F.; Idle, J.R. Tissue metabolomics of hepatocellular carcinoma: Tumor energy metabolism and the role of transcriptomic classification. Hepatology 2013, 58, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Semmo, N.; Weber, T.; Idle, J.R.; Beyoglu, D. Metabolomics reveals that aldose reductase activity due to AKR1B10 is upregulated in hepatitis C virus infection. J. Viral Hepat. 2015, 22, 617–624. [Google Scholar] [CrossRef]
- Wang, M.; Keogh, A.; Treves, S.; Idle, J.R.; Beyoğlu, D. The metabolomic profile of gamma-irradiated human hepatoma and muscle cells reveals metabolic changes consistent with the Warburg effect. PeerJ 2016, 4, e1624. [Google Scholar] [CrossRef] [Green Version]
- Reeds, P.J. Dispensable and Indispensable Amino Acids for Humans. J. Nutr. 2000, 130, 1835S–1840S. [Google Scholar] [CrossRef] [Green Version]
- Lien, O.G., Jr.; Greenberg, D.M. Identification of α-Aminobutyric Acid Enzymatically Formed from Threonine. J. Biol. Chem. 1953, 200, 367–371. [Google Scholar] [CrossRef]
- Liebich, H.M.; Först, C. Hydroxycarboxylic and oxocarboxylic acids in urine:products from branched-chain amino acid degradation and from ketogenesis. J. Chromatogr. B Biomed. Sci. Appl. 1984, 309, 225–242. [Google Scholar] [CrossRef]
- Bommer, M.; Ward, J.M. A 1-step microplate method for assessing the substrate range of l-α-amino acid aminotransferase. Enzym. Microb. Technol. 2013, 52, 218–225. [Google Scholar] [CrossRef] [PubMed]
- EASL. EASL clinical practice guidelines on the management of ascites, spontaneous bacterial peritonitis, and hepatorenal syndrome in cirrhosis. J. Hepatol. 2010, 53, 397–417. [Google Scholar] [CrossRef] [PubMed]
- Greco, A.V.; Mingrone, G.; Gasbarrini, G. Free fatty acid analysis in ascitic fluid improves diagnosis in malignant abdominal tumors. Clin. Chim. Acta 1995, 239, 13–22. [Google Scholar] [CrossRef]
- Cistola, D.; Small, D.M. Fatty acid distribution in systems modeling the normal and diabetic human circulation. A 13C nuclear magnetic resonance study. J. Clin. Investig. 1991, 87, 1431–1441. [Google Scholar] [CrossRef] [Green Version]
- Fasano, M.; Curry, S.; Terreno, E.; Galliano, M.; Fanali, G.; Narciso, P.; Notari, S.; Ascenzi, P. The extraordinary ligand binding properties of human serum albumin. IUBMB Life 2005, 57, 787–796. [Google Scholar] [CrossRef]
- Ashbrook, J.; Spector, A.; Santos, E.; Fletcher, J. Long chain fatty acid binding to human plasma albumin. J. Biol. Chem. 1975, 250, 2333–2338. [Google Scholar] [CrossRef]
- Spector, A. Fatty acid binding to plasma albumin. J. Lipid Res. 1975, 16, 165–179. [Google Scholar] [CrossRef]
- Petitpas, I.; Grüne, T.; Bhattacharya, A.A.; Curry, S. Crystal structures of human serum albumin complexed with monounsaturated and polyunsaturated fatty acids. J. Mol. Biol. 2001, 314, 955–960. [Google Scholar] [CrossRef]
- van der Vusse, G.J. Albumin as Fatty Acid Transporter. Drug Metab. Pharmacokinet. 2009, 24, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.A.; Rinaldi, S.; Scalbert, A.; Ferrari, P.; Achaintre, D.; Gunter, M.J.; Appleby, P.N.; Key, T.J.; Travis, R.C. Plasma concentrations and intakes of amino acids in male meat-eaters, fish-eaters, vegetarians and vegans: A cross-sectional analysis in the EPIC-Oxford cohort. Eur. J. Clin. Nutr. 2016, 70, 306–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalbert, A.; Brennan, L.; Fiehn, O.; Hankemeier, T.; Kristal, B.S.; Van Ommen, B.; Pujos-Guillot, E.; Verheij, E.; Wishart, D.; Wopereis, S. Mass-spectrometry-based metabolomics: Limitations and recommendations for future progress with particular focus on nutrition research. Metabolomics 2009, 5, 435–458. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.; Ohnishi, H.; Kawade, Y.; Muto, Y.; Takahashi, Y. Augmented utilization of branched-chain amino acids by skeletal muscle in decompensated liver cirrhosis in special relation to ammonia detoxication. Gastroenterol. Jpn. 1981, 16, 64–70. [Google Scholar] [CrossRef]
- Meloun, B.; Morávek, L.; Kostka, V. Complete amino acid sequence of human serum albumin. FEBS Lett. 1975, 58, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Gatta, A.; Verardo, A.; Bolognesi, M. Hypoalbuminemia. Intern. Emerg. Med. 2012, 7 (Suppl. S3), S193–S199. [Google Scholar] [CrossRef] [PubMed]
- Hirschberger, J.; Goldberg, M.; Sauer, U.G. Glutamine and Glutamate in Ascitic Fluid of Dogs. Eur. J. Clin. Chem. Clin. Biochem. 1993, 31, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Kragh-Hansen, U.; Chuang, V.T.G.; Otagiri, M. Practical Aspects of the Ligand-Binding and Enzymatic Properties of Human Serum Albumin. Biol. Pharm. Bull. 2002, 25, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, K.; Kuga, N.; Takamura, N.; Furuya, Y.; Hidaka, M.; Iwakiri, T.; Nishii, R.; Okumura, M.; Kodama, H.; Kawai, K.; et al. Inhibitory Effects of Amino-Acid Fluids on Drug Binding to Site II of Human Serum Albumin in Vitro. Biol. Pharm. Bull. 2005, 28, 549–552. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, J.K.; Gartland, K.P.R. 1H NMR studies on protein binding of histidine, tyrosine and phenylalanine in blood plasma. NMR Biomed. 1989, 2, 77–82. [Google Scholar] [CrossRef]
- Varshney, A.; Sen, P.; Ahmad, E.; Rehan, M.; Subbarao, N.; Khan, R.H. Ligand binding strategies of human serum albumin: How can the cargo be utilized? Chirality 2010, 22, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Yeggoni, D.P.; Subramanyam, R. Binding studies of l-3,4-dihydroxyphenylalanine with human serum albumin. Mol. BioSyst. 2014, 10, 3101–3110. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.; Iwakiri, Y. The lymphatic vascular system in liver diseases: Its role in ascites formation. Clin. Mol. Hepatol. 2013, 19, 99–104. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Nemkov, T.; Moore, H.B.; Moore, E.E.; Wither, M.; Nydam, T.; Slaughter, A.; Silliman, C.C.; Banerjee, A.; Hansen, K.C. Metabolomics of trauma-associated death: Shared and fluid-specific features of human plasma vs lymph. Blood Transfus. 2016, 14, 185–194. [Google Scholar] [CrossRef]
- Idle, J.R.; Seipel, K.; Bacher, U.; Pabst, T.; Beyoğlu, D. (2R,3S)-Dihydroxybutanoic Acid Synthesis as a Novel Metabolic Function of Mutant Isocitrate Dehydrogenase 1 and 2 in Acute Myeloid Leukemia. Cancers 2020, 12, 2842. [Google Scholar] [CrossRef]
- Tetko, I.V.; Tanchuk, V.Y.; Kasheva, T.N.; Villa, A.E. Estimation of Aqueous Solubility of Chemical Compounds Using E-State Indices. J. Chem. Inf. Comput. Sci. 2001, 41, 1488–1493. [Google Scholar] [CrossRef]
- Wang, J.; Shen, X.; Jain, R.; Wang, J.; Yuan, Q.; Yan, Y. Establishing a novel biosynthetic pathway for the production of 3,4-dihydroxybutyric acid from xylose in Escherichia coli. Metab. Eng. 2017, 41, 39–45. [Google Scholar] [CrossRef]
- Fell, V.; Lee, C.; Pollitt, R. The occurrence of (S)-3,4-dihydroxybutyrate in human blood and urine. Biochem. Med. 1975, 13, 40–45. [Google Scholar] [CrossRef]
- Bala, L.; Sharma, A.; Yellapa, R.K.; Roy, R.; Choudhuri, G.; Khetrapal, C.L. 1H NMR spectroscopy of ascitic fluid: Discrimination between malignant and benign ascites and comparison of the results with conventional methods. NMR Biomed. 2008, 21, 606–614. [Google Scholar] [CrossRef]
- Beyoglu, D.; Idle, J.R. Metabolomic insights into the mode of action of natural products in the treatment of liver disease. Biochem. Pharmacol. 2020, 180, 114171. [Google Scholar] [CrossRef]
- Miller, R.G., Jr. Simultaneous Statistical Inference; Springer: New York, NY, USA, 1981; p. 299. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite (Reported Previously in Ascites; Y = Yes, N = No) | Control Peritoneal Fluid | Cirrhotic Ascites | * p Value | Ascites/ Control Ratio |
|---|---|---|---|---|
| METABOLIC INTERMEDIATES | ||||
| Lactic acid (Y) | 1071 | 451 | <0.002 | 0.4 |
| Glycolic acid (N) | 4 | 6 | 0.007 | 1.5 |
| Citric acid (Y) | 47 | 124 | <0.002 | 2.6 |
| Uric acid (Y) | 62 | 138 | <0.002 | 2.2 |
| AMINO ACIDS AND AMINO ACID METABOLITES | ||||
| Glutamic acid (Y) | 9 | 2 | <0.002 | 0.2 |
| Glutamine (Y) | 36 | 61 | <0.002 | 1.7 |
| Proline (N) | 113 | 149 | 0.005 | 1.3 |
| Histidine (Y) | 7 | 13 | 0.007 | 1.9 |
| Tyrosine (Y) | 58 | 107 | <0.002 | 1.8 |
| 2-Ketoisocaproic acid (N) | 2 | 0.8 | <0.002 | 0.4 |
| Pipecolic acid (N) | 1 | 3 | <0.002 | 3.0 |
| SUGARS AND SUGAR METABOLITES | ||||
| Erythritol (N) | 2 | 5 | <0.002 | 2.5 |
| Mannitol (N) | 110 | 21 | 0.002 | 0.2 |
| Gluconic acid (N) | 3 | 2 | <0.002 | 0.7 |
| Myo-inositol (N) | 20 | 42 | <0.002 | 2.1 |
| Scyllo-inositol (N) | 1 | 5 | <0.002 | 5.0 |
| Fructose (N) | 5 | 18 | <0.002 | 3.6 |
| LIPIDS | ||||
| Lauric acid (12:0) (Y) | 2 | 0.8 | <0.002 | 0.4 |
| Myristic acid (14:0) (Y) | 4 | 1 | <0.002 | 0.3 |
| Palmitic acid (16:0) (Y) | 41 | 12 | <0.002 | 0.3 |
| Linoleic acid (18:2 9Z,12Z) (Y) | 5 | 2 | <0.002 | 0.4 |
| Oleic acid (18:1 9Z) (Y) | 27 | 7 | <0.002 | 0.3 |
| Stearic acid (18:0) (N) | 14 | 6 | <0.002 | 0.4 |
| Cholesterol (Y) | 31 | 16 | <0.002 | 0.5 |
| Metabolite (Reported Previously in Ascites; Y = Yes, N = No) | Plasma (Median) | Ascites (Median) | * p Value | Ascites/ Plasma Ratio |
|---|---|---|---|---|
| AMINO ACIDS | ||||
| Glycine (Y) | 31 | 43 | 0.004 | 1.4 |
| Valine (Y) | 61 | 73 | 0.004 | 1.2 |
| Proline (N) | 85 | 136 | 0.002 | 1.6 |
| Threonine (N) | 12 | 22 | 0.002 | 1.8 |
| Methionine (Y) | 9 | 13 | 0.004 | 1.4 |
| Lysine (Y) | 17 | 22 | 0.004 | 1.3 |
| Glutamine (Y) | 34 | 62 | 0.02 | 1.8 |
| Phenylalanine (Y) | 27 | 34 | 0.03 | 1.3 |
| Tyrosine (Y) | 65 | 113 | 0.004 | 1.7 |
| OTHER INTERMEDIARY METABOLITES | ||||
| Pyruvic acid (Y) | 328 | 532 | 0.004 | 1.6 |
| Fumaric acid (N) | 1 | 3 | 0.02 | 3.0 |
| Sorbitol (N) | 0.4 | 0.6 | 0.02 | 1.5 |
| Phosphate (N) | 371 | 49 | 0.007 | 0.1 |
| LIPIDS | ||||
| Myristic acid (14:0) (Y) | 5 | 1 | 0.002 | 0.2 |
| Palmitic acid (16:0) (Y) | 59 | 12 | 0.002 | 0.2 |
| Oleic acid (18:1 9Z) (Y) | 45 | 6 | 0.002 | 0.1 |
| Vaccenic acid (18:1 11E) (N) | 4 | 0.7 | 0.002 | 0.2 |
| Stearic acid (18:0) (N) | 12 | 5 | 0.002 | 0.4 |
| Cholesterol (Y) | 97 | 21 | 0.002 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beyoğlu, D.; Simillion, C.; Storni, F.; De Gottardi, A.; Idle, J.R. A Metabolomic Analysis of Cirrhotic Ascites. Molecules 2022, 27, 3935. https://doi.org/10.3390/molecules27123935
Beyoğlu D, Simillion C, Storni F, De Gottardi A, Idle JR. A Metabolomic Analysis of Cirrhotic Ascites. Molecules. 2022; 27(12):3935. https://doi.org/10.3390/molecules27123935
Chicago/Turabian StyleBeyoğlu, Diren, Cedric Simillion, Federico Storni, Andrea De Gottardi, and Jeffrey R. Idle. 2022. "A Metabolomic Analysis of Cirrhotic Ascites" Molecules 27, no. 12: 3935. https://doi.org/10.3390/molecules27123935