
Artemisia anomala Herba Alleviates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis-Like Skin Lesions in Mice and the Production of Pro-Inflammatory Mediators in Tumor Necrosis Factor Alpha-/Interferon Gamma-Induced HaCaT Cells



Abstract
:1. Introduction
2. Results
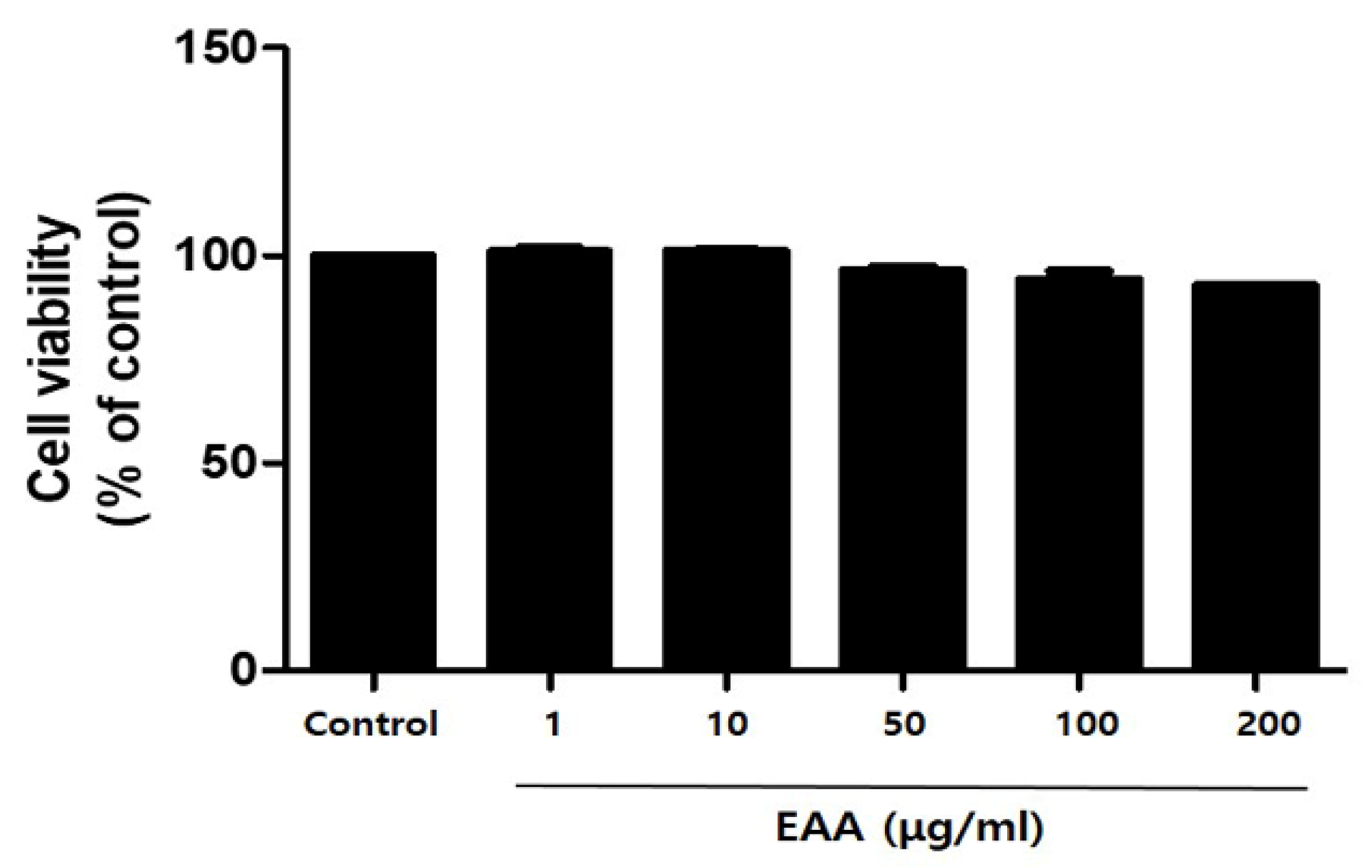
2.1. In Vitro Cell Cytotoxicity in Keratinocytes
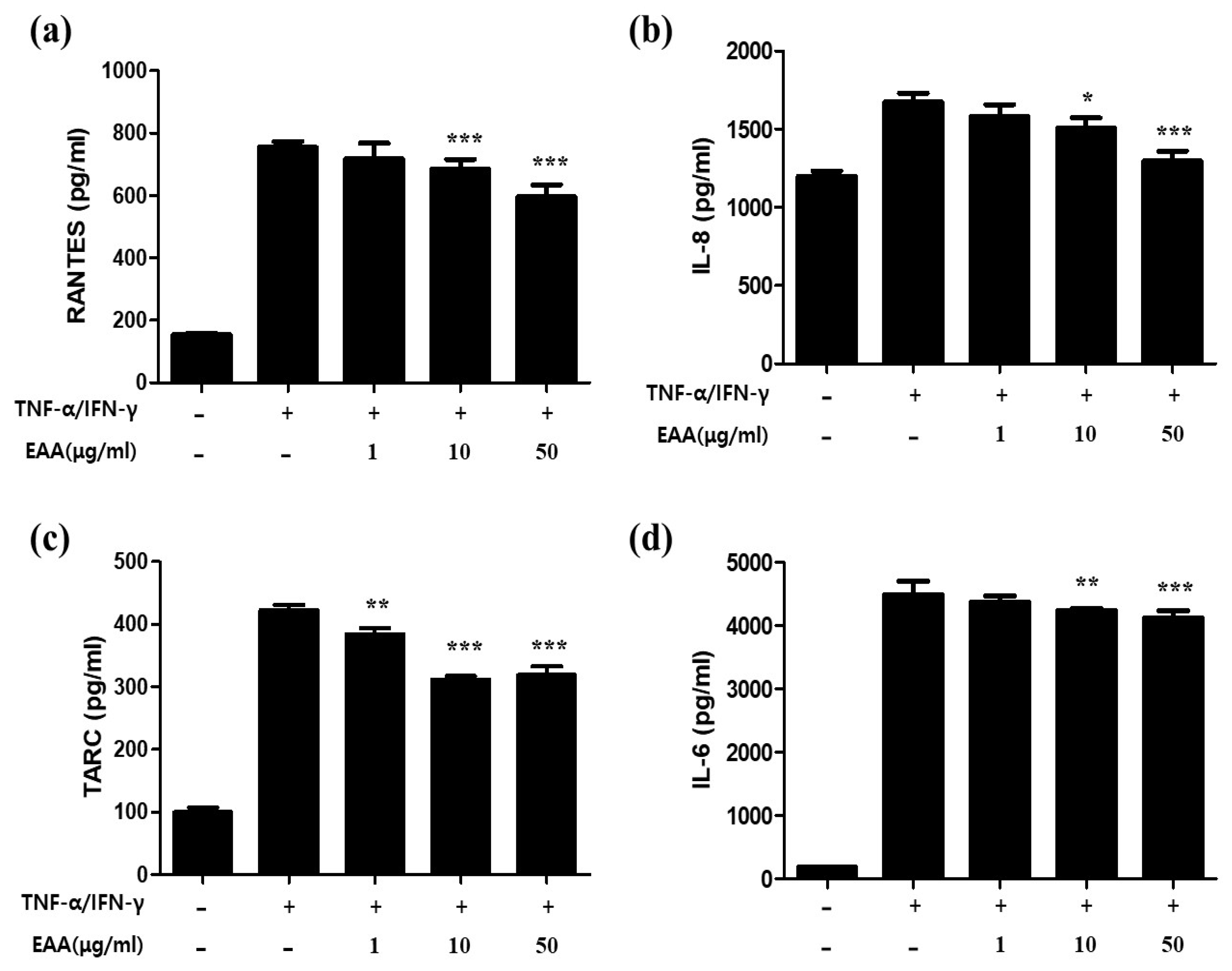
2.2. EAA Suppresses the Production of AD-Related Cytokines and Chemokines in Keratinocytes
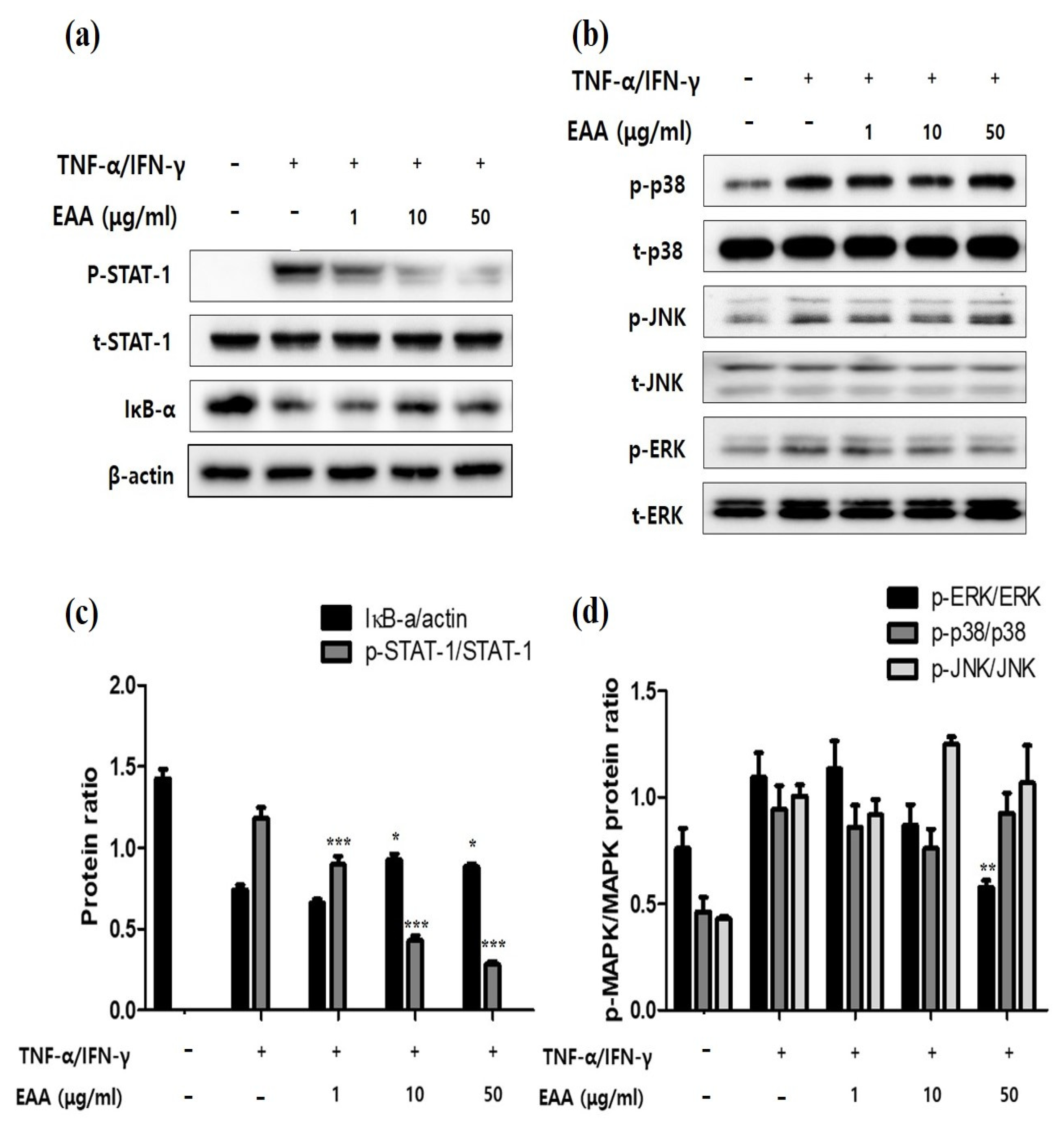
2.3. EAA Inhibits NFκB and STAT-1 Activation in Keratinocytes
2.4. EAA Inhibits MAPKs in Keratinocytes
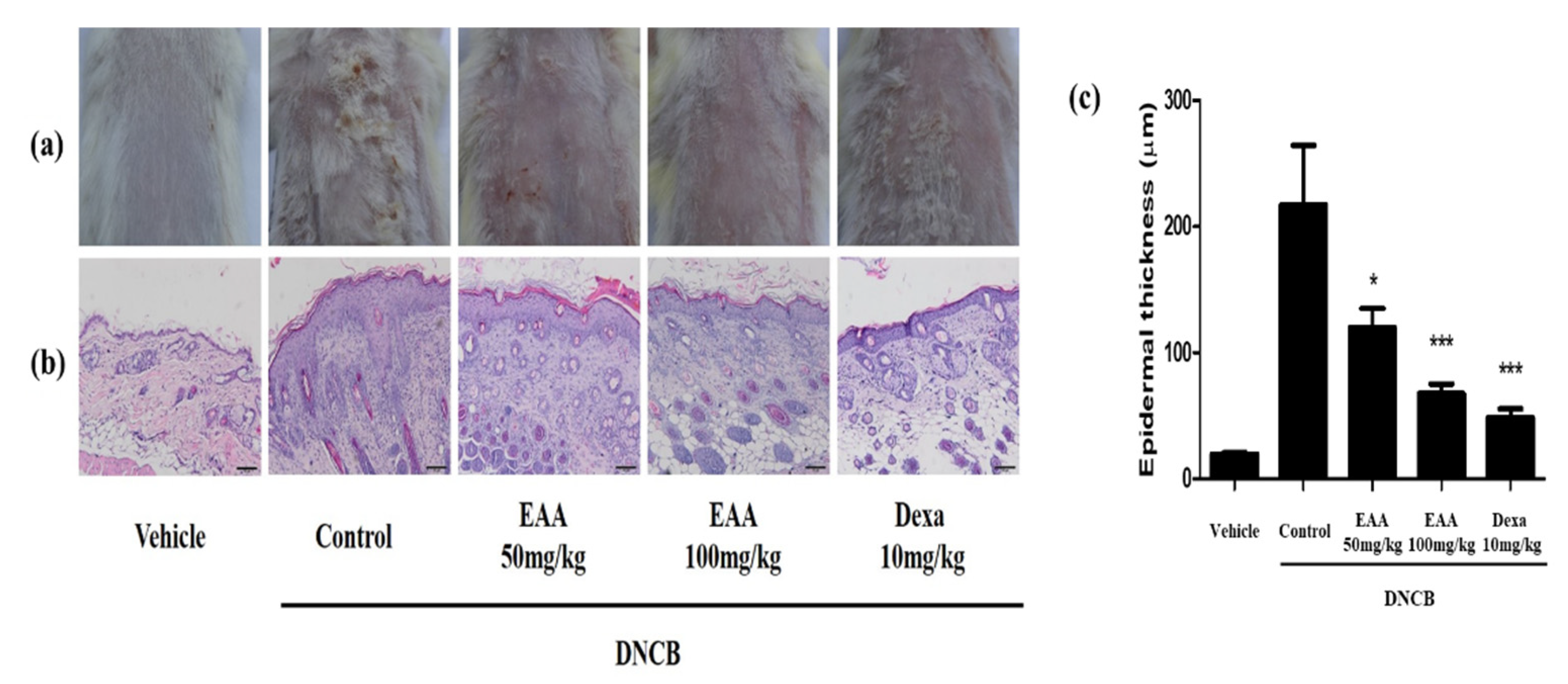
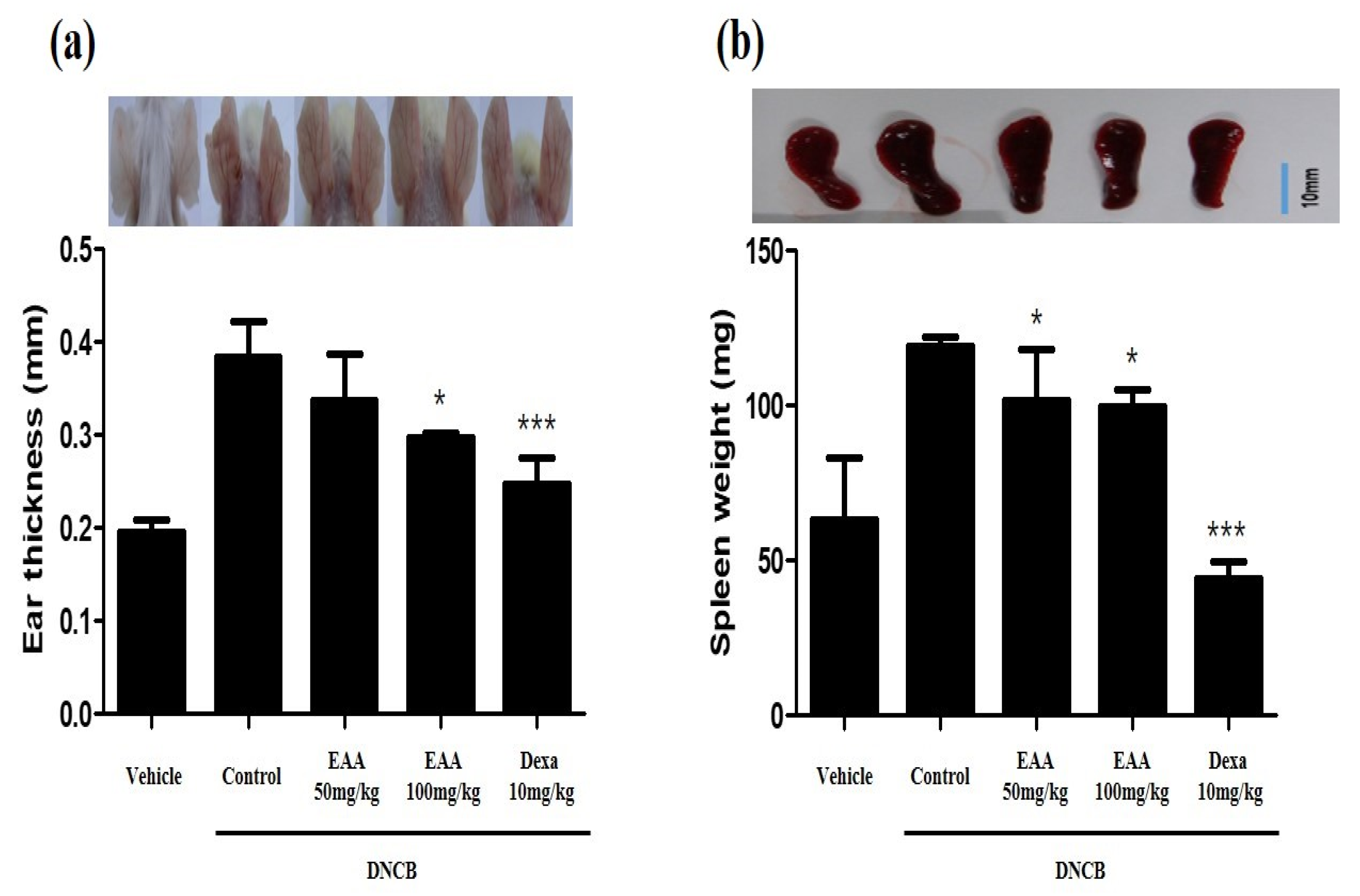
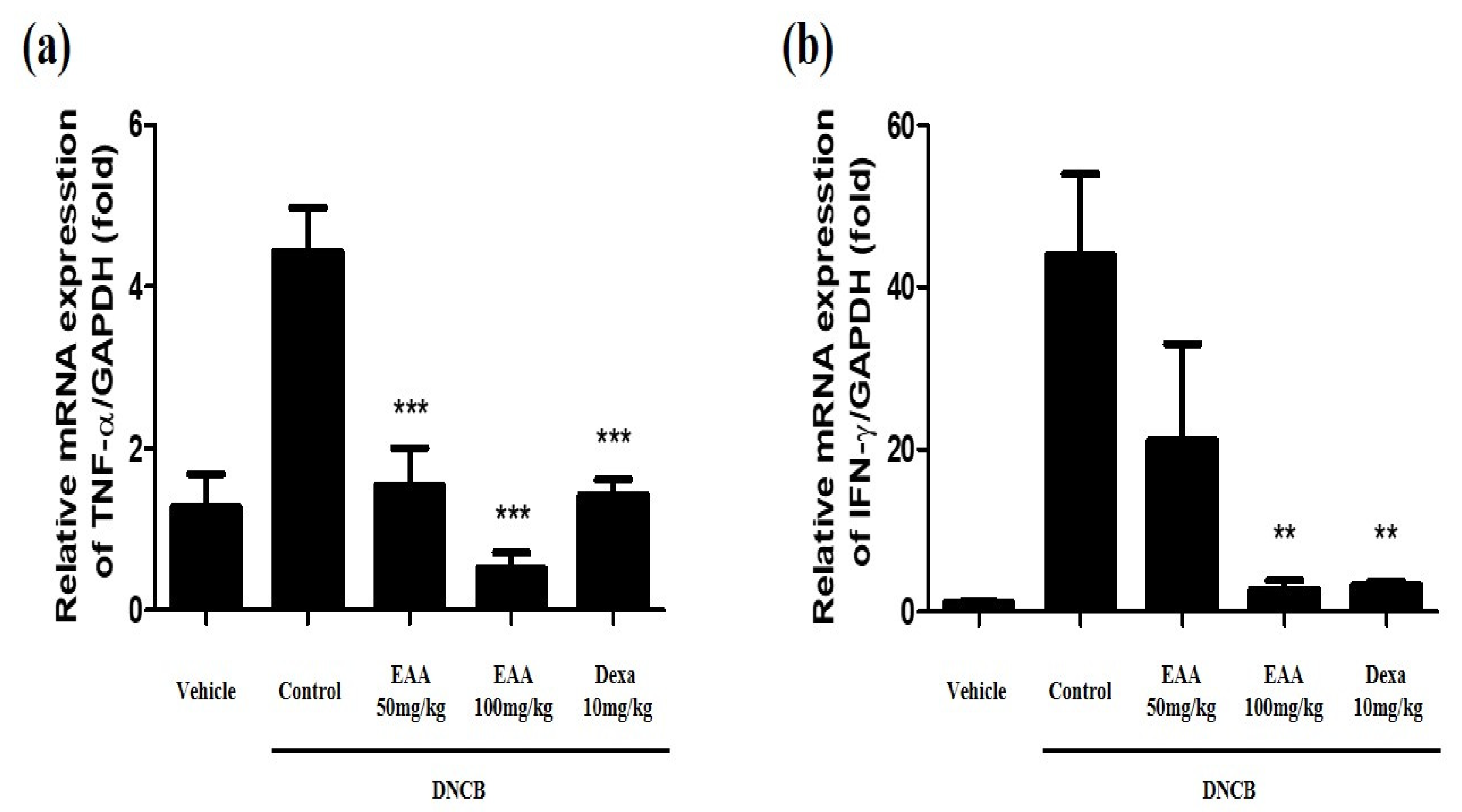
2.5. EAA Reduces the Symptoms of AD in Mice
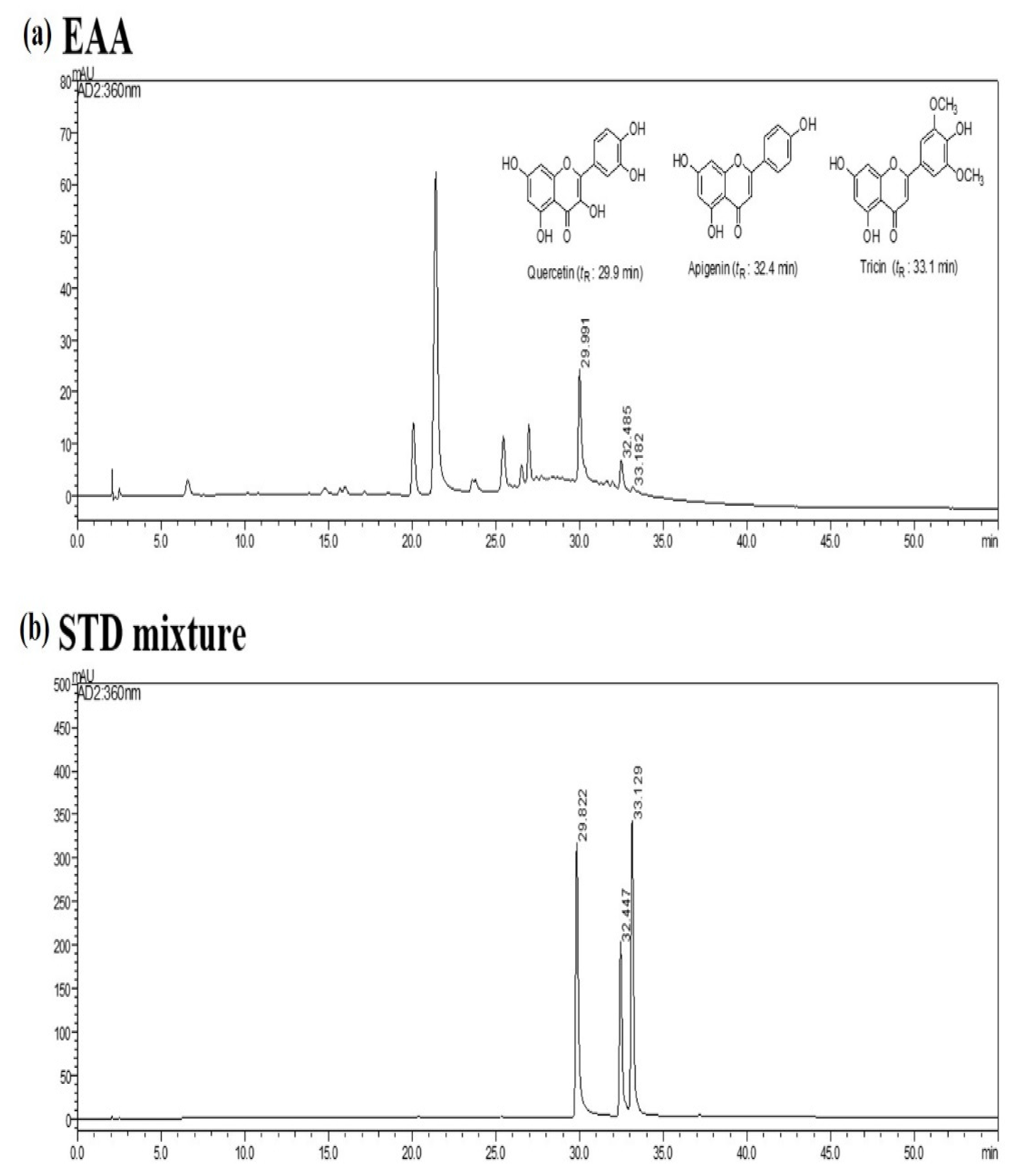
2.6. HPLC Analysis of EAA
3. Discussion
4. Materials and Methods
4.1. Reagents and Cell Culture
4.2. Cell Culture
4.3. Cell Viability Test
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Western Blot Analysis
4.6. Atopic-like Dermatitis Mouse Model
4.7. Histopathological Observation
4.8. Isolation of RNA, cDNA Synthesis, and Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
4.9. Conditions of Chromatographic
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Doh, E.J.; Paek, S.H.; Lee, G.; Lee, M.Y.; Oh, S.E. Application of Partial Internal Transcribed Spacer Sequences for the Discrimination of Artemisia capillaris from Other Artemisia Species. Evid.-Based Complement. Altern. Med. 2016, 2016, 7043436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes-Lutz, D.; Alviano, D.S.; Alviano, C.S.; Kolodziejczyk, P.P. Screening of chemical composition, antimicrobial and antioxidant activities of Artemisia essential oils. Phytochemistry 2008, 69, 1732–1738. [Google Scholar] [CrossRef]
- Pan, Y.F.; Zhang, D.D.; Ling, S.; Zhang, H.P.; Bian, H.S.; Bian, K. Analysis on anti-vascular inflammatory mechanism in vitro of total flavones from Artemisia anomala. Zhongguo Zhong Yao Za Zhi 2012, 37, 2597–2602. [Google Scholar]
- Tan, X.; Wang, Y.L.; Yang, X.L.; Zhang, D.D. Ethyl Acetate Extract of Artemisia anomala S. Moore Displays Potent Anti-Inflammatory Effect. Evid.-Based Complement. Altern. Med. 2014, 2014, 681352. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Jiang, H.; Li, W.; Qiang, M.; Dong, T.; Li, H. Role of Vitamin C in Skin Diseases. Front. Physiol. 2018, 9, 819. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.; Morello, E.; Garnier, J.; Barrault, C.; Garnier, M.; Burucoa, C.; Lecron, J.C.; Si-Tahar, M.; Bernard, F.X.; Bodet, C. Pseudomonas aeruginosa flagellum is critical for invasion, cutaneous persistence and induction of inflammatory response of skin epidermis. Virulence 2018, 9, 1163–1175. [Google Scholar] [CrossRef] [Green Version]
- Hunter, J.A. Diseases of the skin. Structure and function of skin in relation to therapy. Br. Med. J. 1973, 4, 340–342. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.H.; Yoo, J.M.; Lee, E.; Lee, B.; Cho, W.K.; Park, K.I.; Ma, J.Y. Anti-inflammatory effects of Perillae Herba ethanolic extract against TNF-alpha/IFN-gamma-stimulated human keratinocyte HaCaT cells. J. Ethnopharmacol. 2018, 213, 445. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.W.; Lee, K.S.; Kim, C.W. Curcumin attenuates the expression of IL-1beta, IL-6, and TNF-alpha as well as cyclin E in TNF-alpha-treated HaCaT cells; NF-kappaB and MAPKs as potential upstream targets. Int. J. Mol. Med. 2007, 19, 469–474. [Google Scholar]
- Choi, H.J.; Lee, J.H.; Jung, Y.S. (+)-Nootkatone inhibits tumor necrosis factor alpha/interferon gamma-induced production of chemokines in HaCaT cells. Biochem. Biophys. Res. Commun. 2014, 447, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, X.; Wang, H.; Zhao, B.; Wu, X.; Su, L.; Xie, S.; Wang, Y.; Li, J.; Liu, J.; et al. PKCzeta as a promising therapeutic target for TNFalpha-induced inflammatory disorders in chronic cutaneous wounds. Int. J. Mol. Med. 2017, 40, 1335–1346. [Google Scholar] [CrossRef]
- Kong, L.; Liu, J.; Wang, J.; Luo, Q.; Zhang, H.; Liu, B.; Xu, F.; Pang, Q.; Liu, Y.; Dong, J. Icariin inhibits TNF-alpha/IFN-gamma induced inflammatory response via inhibition of the substance P and p38-MAPK signaling pathway in human keratinocytes. Int. Immunopharmacol. 2015, 29, 401–407. [Google Scholar] [CrossRef]
- Allam, J.P.; Novak, N. The pathophysiology of atopic eczema. Clin. Exp. Dermatol. 2006, 31, 89–93. [Google Scholar] [CrossRef]
- Jeong, S.J.; Lim, H.S.; Seo, C.S.; Kim, J.H.; Jin, S.E.; Yoo, S.R.; Shin, H.K. Traditional herbal formula Jakyakgamcho-tang (Paeonia lactiflora and Glycyrrhiza uralensis) impairs inflammatory chemokine production by inhibiting activation of STAT1 and NF-kappaB in HaCaT cells. Phytomedicine 2015, 22, 326–332. [Google Scholar] [CrossRef]
- Huang, W.C.; Dai, Y.W.; Peng, H.L.; Kang, C.W.; Kuo, C.Y.; Liou, C.J. Phloretin ameliorates chemokines and ICAM-1 expression via blocking of the NF-kappaB pathway in the TNF-alpha-induced HaCaT human keratinocytes. Int. Immunopharmacol. 2015, 27, 32–37. [Google Scholar] [CrossRef]
- Ramana, K.V.; Singhal, S.S.; Reddy, A.B. Therapeutic potential of natural pharmacological agents in the treatment of human diseases. Biomed. Res. Int. 2014, 2014, 573452. [Google Scholar] [CrossRef]
- Park, H.J.; Jang, Y.J.; Yim, J.H.; Lee, H.K.; Pyo, S. Ramalin Isolated from Ramalina Terebrata Attenuates Atopic Dermatitis-like Skin Lesions in Balb/c Mice and Cutaneous Immune Responses in Keratinocytes and Mast Cells. Phytother. Res. 2016, 30, 1978–1987. [Google Scholar] [CrossRef]
- Werfel, T. The role of leukocytes, keratinocytes, and allergen-specific IgE in the development of atopic dermatitis. J. Investig. Dermatol. 2009, 129, 1878–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, Y.Y.; Kim, Y.S.; Kim, H.K. Illicium verum extract inhibits TNF-alpha- and IFN-gamma-induced expression of chemokines and cytokines in human keratinocytes. J. Ethnopharmacol. 2012, 144, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Do, H.J.; Lee, E.; Yim, N.H.; Cho, W.K.; Park, K.I.; Ma, J.Y. Jageum-Jung improves 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in mice and suppresses pro-inflammatory chemokine production by inhibiting TNF-alpha/IFN-gamma-induced STAT-1 and NFkappaB signaling in HaCaT cells. J. Ethnopharmacol. 2018, 221, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Gold, M.S.; Kemp, A.S. Atopic disease in childhood. Med. J. Aust. 2005, 182, 298–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, H.; Watanabe, N.; Geba, G.P.; Sperl, J.; Tsudzuki, M.; Hiroi, J.; Matsumoto, M.; Ushio, H.; Saito, S.; Askenase, P.W.; et al. Development of atopic dermatitis-like skin lesion with IgE hyperproduction in NC/Nga mice. Int. Immunol. 1997, 9, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, N.; Shiraishi, N.; Igeta, K.; Itoh, T.; Chikumoto, T.; Nagao, M.; Kim, J.F.; Nagai, H. Inhibition of scratching behavior associated with allergic dermatitis in mice by tacrolimus, but not by dexamethasone. Eur. J. Pharmacol. 2006, 546, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Kabashima, K. New concept of the pathogenesis of atopic dermatitis: Interplay among the barrier, allergy, and pruritus as a trinity. J. Dermatol. Sci. 2013, 70, 3–11. [Google Scholar] [CrossRef]
- Brenninkmeijer, E.E.; Spuls, P.I.; Legierse, C.M.; Lindeboom, R.; Smitt, J.H.; Bos, J.D. Clinical differences between atopic and atopiform dermatitis. J. Am. Acad. Dermatol. 2008, 58, 407–414. [Google Scholar] [CrossRef]
- Tan, H.Y.; Zhang, A.L.; Chen, D.; Xue, C.C.; Lenon, G.B. Chinese herbal medicine for atopic dermatitis: A systematic review. J. Am. Acad. Dermatol. 2013, 69, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Schooling, C.M.; Au Yeung, S.L.; Freeman, G.; Cowling, B.J. The effect of statins on testosterone in men and women, a systematic review and meta-analysis of randomized controlled trials. BMC Med. 2013, 11, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.; Panda, S. Use of Topical Corticosteroids in Dermatology: An Evidence-based Approach. Indian J. Dermatol. 2017, 62, 237–250. [Google Scholar]
- Yang, J.H.; Yoo, J.M.; Cho, W.K.; Ma, J.Y. Ethanol Extract of Sanguisorbae Radix Inhibits Mast Cell Degranulation and Suppresses 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis-Like Skin Lesions. Mediat. Inflamm. 2016, 2016, 2947390. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regression Equation | Linear Range | Content | ||
|---|---|---|---|---|
| Compounds | tR (min) | (y = ax + b, R2) | (mg/mL) | (mg/g) |
| Quercetin | 29.9 | y = 0.494179x + 4.1, 0.9995 | 10–200 | 14.1 ± 0.7 |
| Apigenin | 32.4 | y = 0.124202x + 2.8, 0.9996 | 10–200 | 5.2 ± 0.4 |
| Tricin | 33.1 | y = 0.932455x + 3.8, 0.9998 | 10–200 | 0.4 ± 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.-H.; Kim, K.-Y.; Kim, Y.-W.; Park, K.-I. Artemisia anomala Herba Alleviates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis-Like Skin Lesions in Mice and the Production of Pro-Inflammatory Mediators in Tumor Necrosis Factor Alpha-/Interferon Gamma-Induced HaCaT Cells. Molecules 2021, 26, 5427. https://doi.org/10.3390/molecules26175427
Yang J-H, Kim K-Y, Kim Y-W, Park K-I. Artemisia anomala Herba Alleviates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis-Like Skin Lesions in Mice and the Production of Pro-Inflammatory Mediators in Tumor Necrosis Factor Alpha-/Interferon Gamma-Induced HaCaT Cells. Molecules. 2021; 26(17):5427. https://doi.org/10.3390/molecules26175427
Chicago/Turabian StyleYang, Ju-Hye, Kwang-Youn Kim, Young-Woo Kim, and Kwang-Il Park. 2021. "Artemisia anomala Herba Alleviates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis-Like Skin Lesions in Mice and the Production of Pro-Inflammatory Mediators in Tumor Necrosis Factor Alpha-/Interferon Gamma-Induced HaCaT Cells" Molecules 26, no. 17: 5427. https://doi.org/10.3390/molecules26175427