
Identification of the Primary Structure of Selenium-Containing Polysaccharides Selectively Inhibiting T-Cell Proliferation

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Composition of the Se-Le-30 (Se-Containing Lentinan Analog Isolated from Se-Enriched L. edodes Mycelium by the Chihara Method) Polysaccharide Fraction
2.1.1. Selenium Content
2.1.2. Monosaccharide Composition
2.1.3. Protein Content
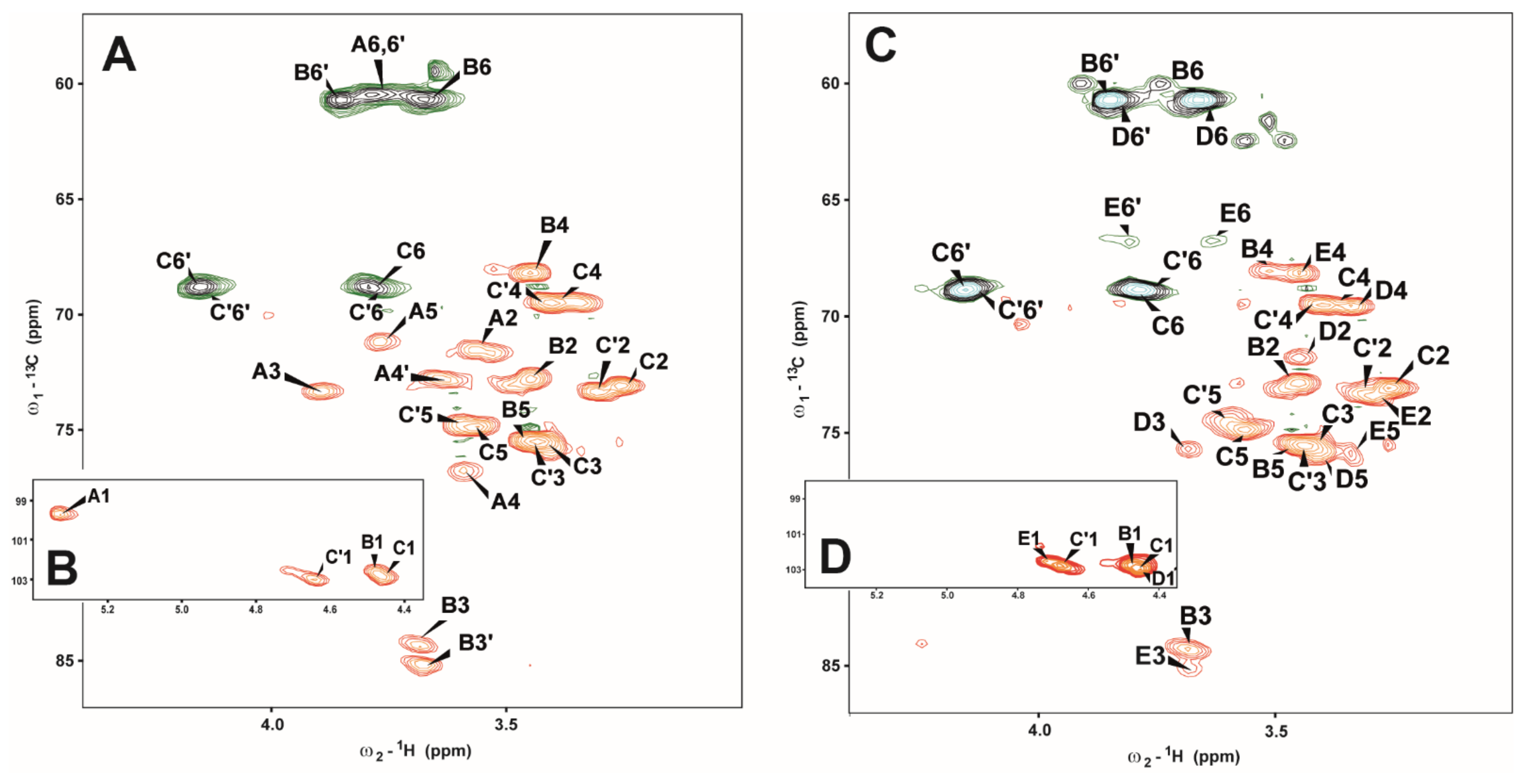
2.2. Structural Analysis on Separated Polysaccharides of Se-Le-30 Crude Extract
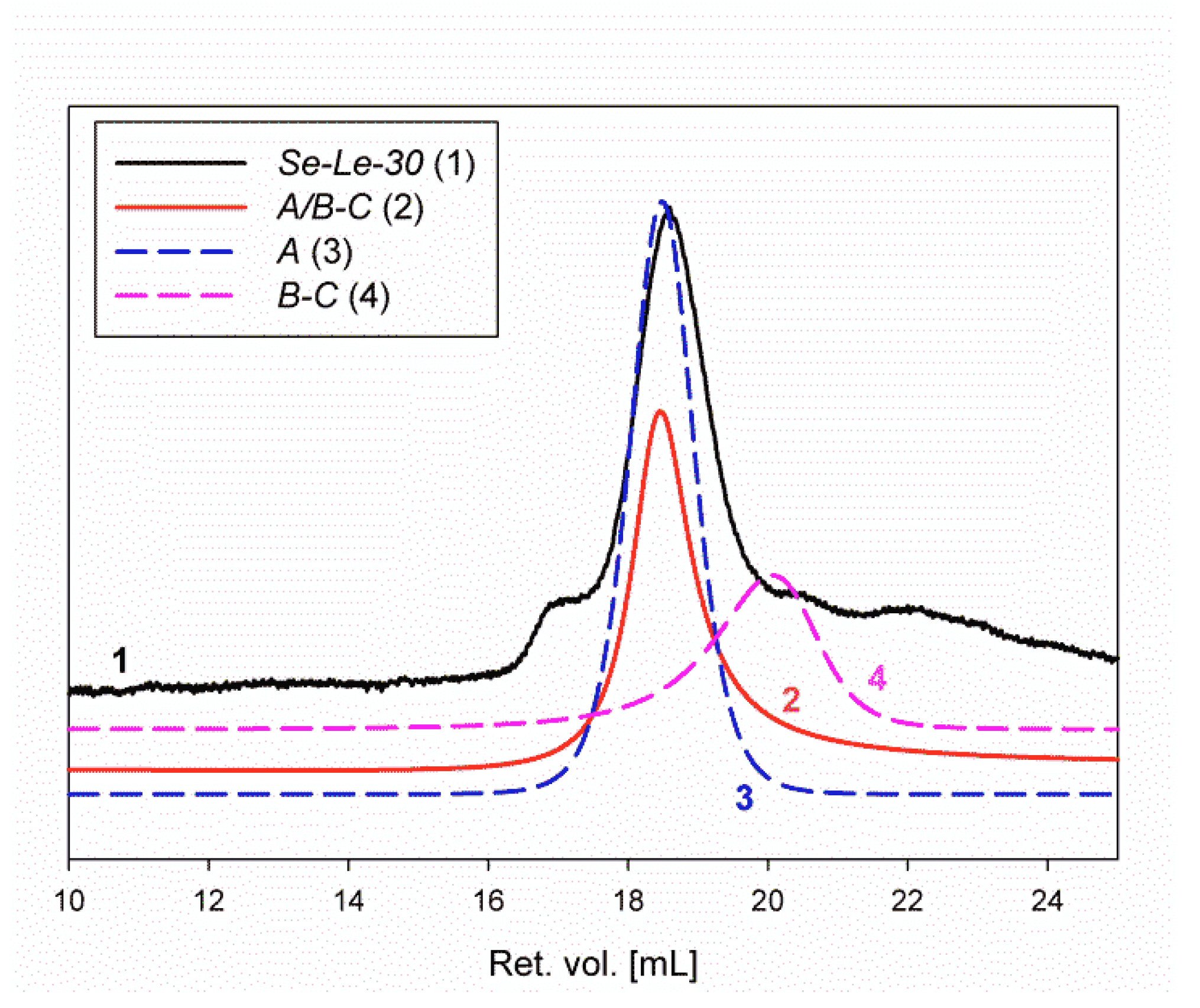
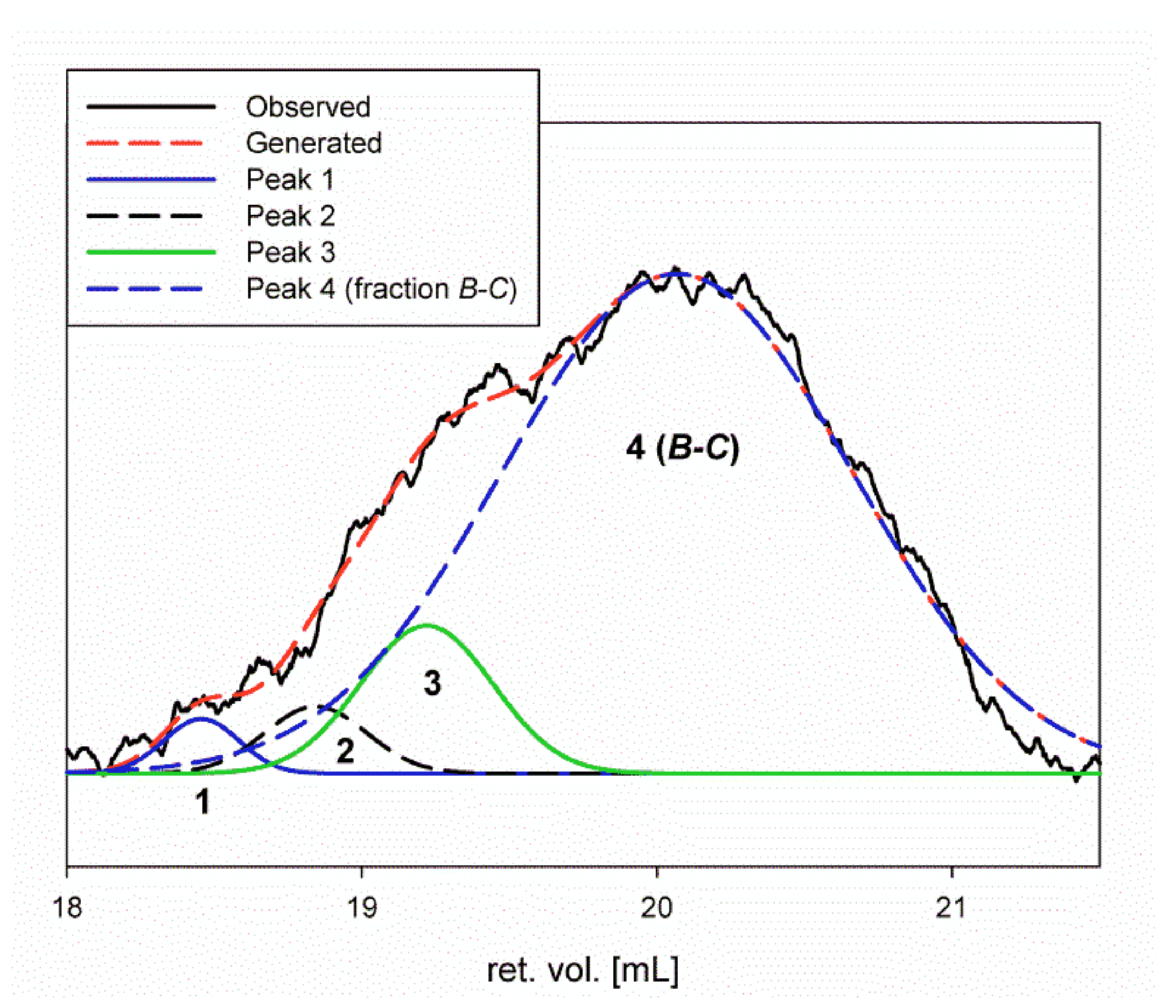
2.3. Homogeneity and Molar Mass Determination of the Lentinan Analog (Se-Le-30) and Isolated Polysaccharides
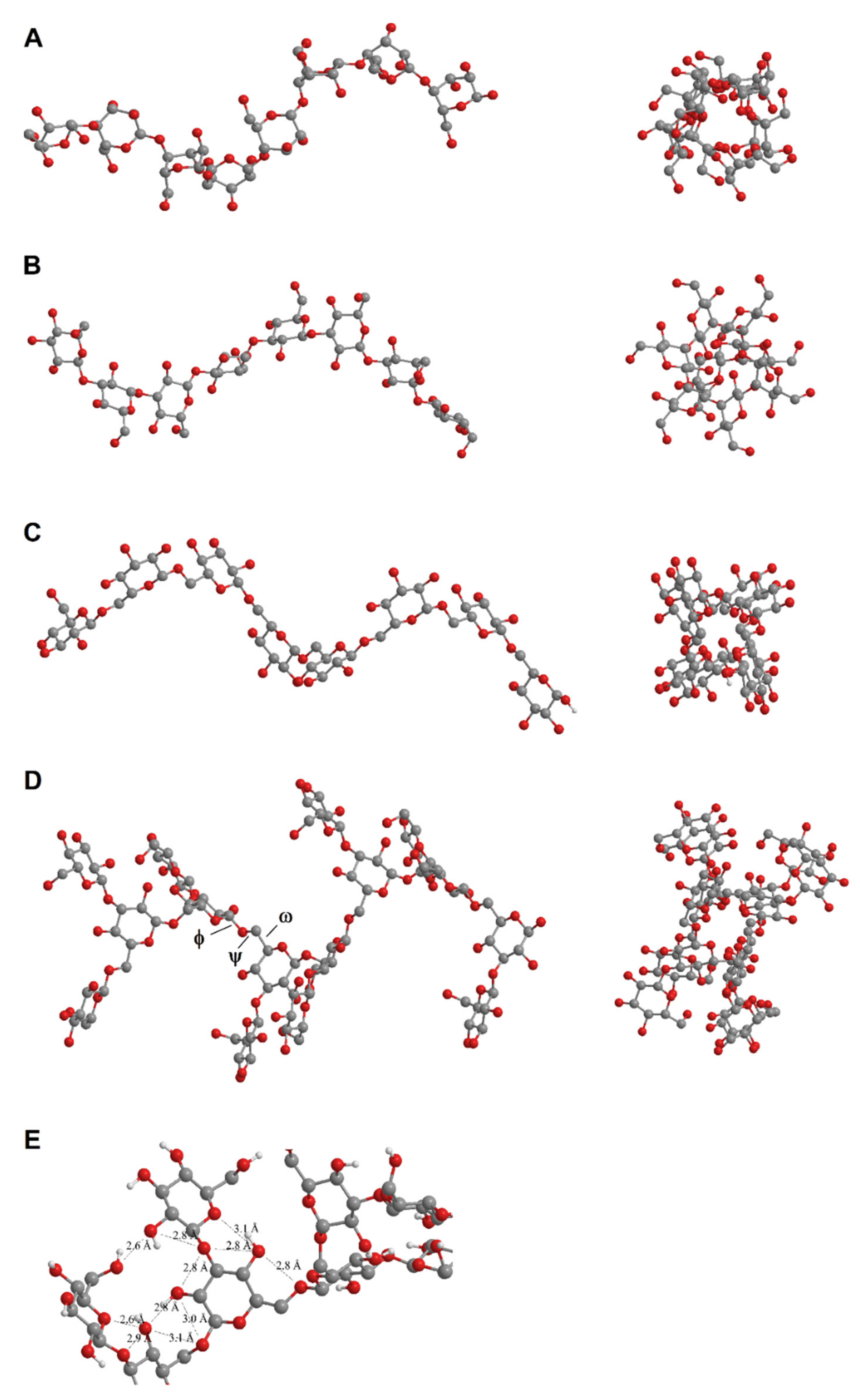
2.4. Molecular Modeling
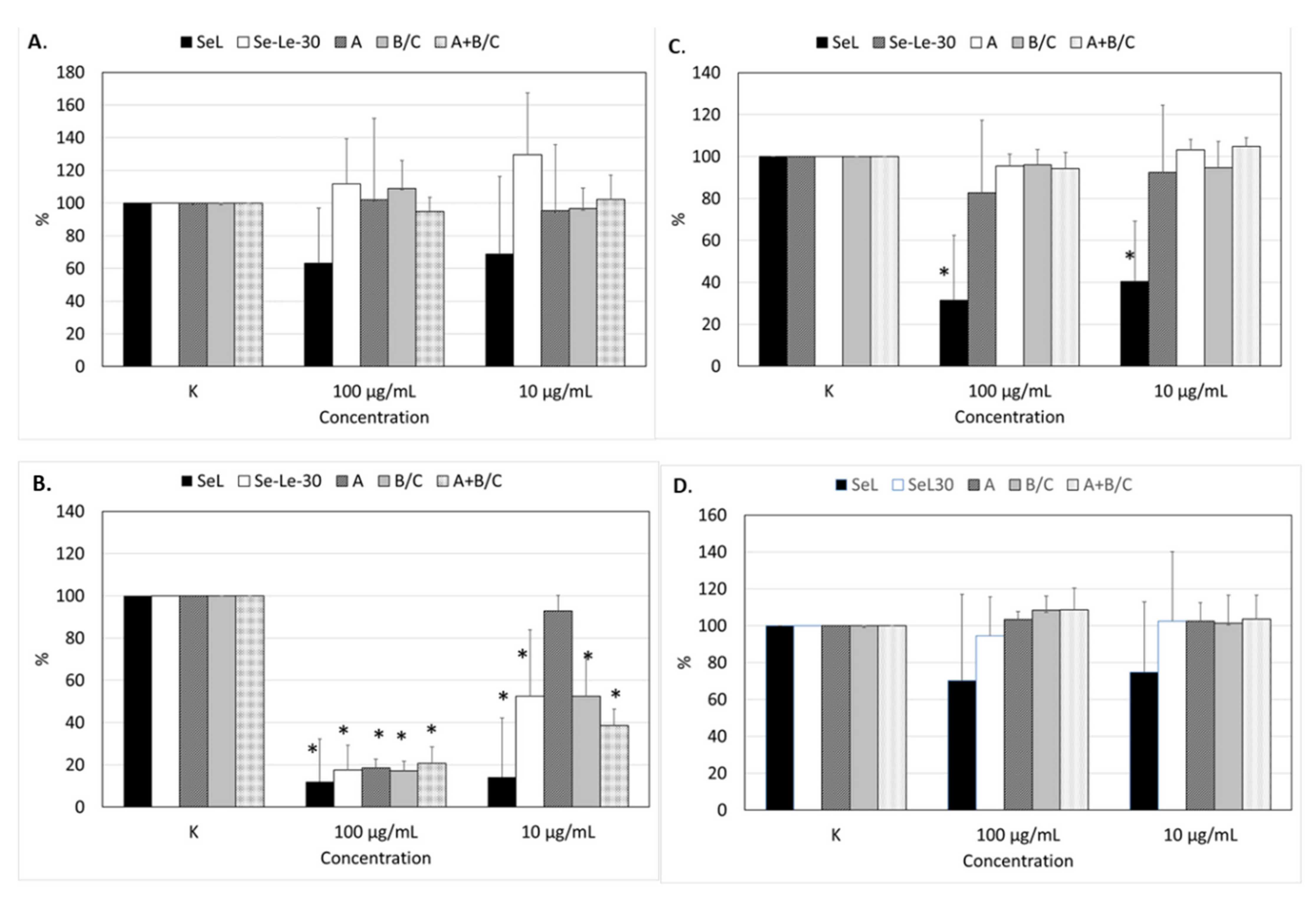
2.5. Effects of Polysaccharides on the Proliferation of Human Peripheral Blood Mononuclear Cells
3. Discussion
3.1. Se-Le-30 Fraction
3.2. Fraction A (I)
3.3. Fraction B-C (III)
3.4. Biological Activity
4. Materials and Methods
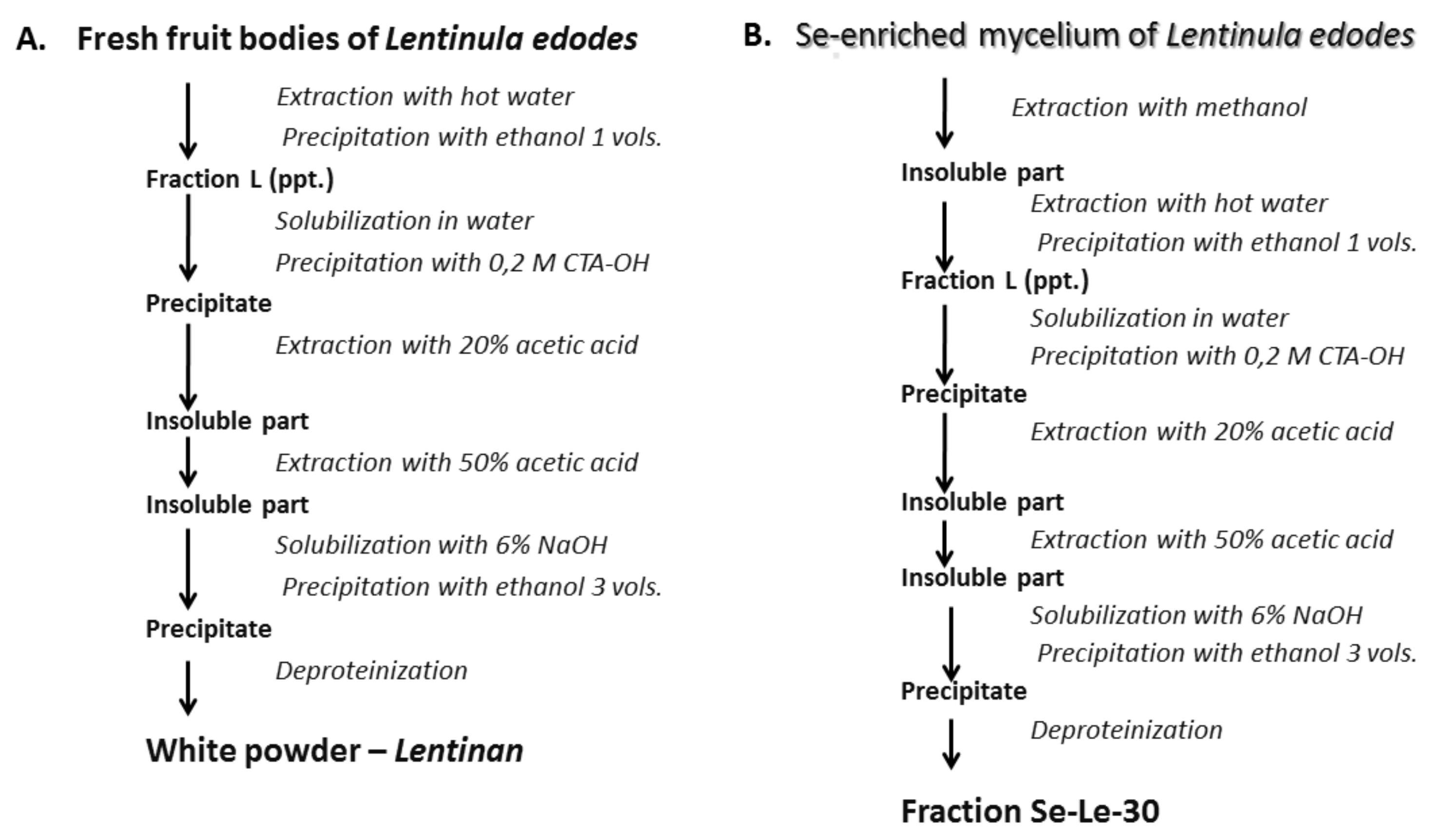
4.1. Biosynthesis and Isolation of Se-Enriched Polysaccharide Fractions
4.1.1. Mushroom Strain and Cultivation Conditions
4.1.2. Extraction of the Se-Enriched Polysaccharide Fraction Using the Chihara Method
4.1.3. Separation of Se-Enriched Polysaccharides Fraction Se-Le-30
4.2. Structural Analysis
4.2.1. Molar Mass Determination by SEC Chromatography with Triple Detection
4.2.2. RP-HPLC Determination of Monosaccharide Composition
4.2.3. RP HPLC Determination of Se Content
4.2.4. Determination of Protein Content in the Lentinan Analogue (Se-Le-30)
4.2.5. Sugar and Methylation Analysis and Determination of the Absolute Configuration
4.2.6. NMR Analysis
4.3. Molecular Modeling
4.4. Effects of Polysaccharides on Human Peripheral Blood Mononuclear Cell Proliferation
4.5. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Cheng, S.-C.; Quintin, J.; Cramer, R.A.; Shepardson, K.M.; Saeed, S.; Kumar, V.; Giamarellos-Bourboulis, E.J.; Martens, J.H.A.; Rao, N.A.; Aghajanirefah, A.; et al. MTOR- and HIF-1 -Mediated Aerobic Glycolysis as Metabolic Basis for Trained Immunity. Science 2014, 345, 1250684. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Luo, F.; Ding, C.; Albeituni, S.; Hu, X.; Ma, Y.; Cai, Y.; McNally, L.; Sanders, M.A.; Jain, D.; et al. Dectin-1 Activation by a Natural Product β-Glucan Converts Immunosuppressive Macrophages into an M1-like Phenotype. J. Immunol. 2015, 195, 5055–5065. [Google Scholar] [CrossRef]
- Rathore, H.; Prasad, S.; Kapri, M.; Tiwari, A.; Sharma, S. Medicinal Importance of Mushroom Mycelium: Mechanisms and Applications. J. Funct. Foods 2019, 56, 182–193. [Google Scholar] [CrossRef]
- Latgé, J.-P. Tasting the Fungal Cell Wall: Tasting Fungal Cell Wall. Cell. Microbiol. 2010, 12, 863–872. [Google Scholar] [CrossRef]
- Hardison, S.E.; Brown, G.D. C-Type Lectin Receptors Orchestrate Antifungal Immunity. Nat. Immunol. 2012, 13, 817–822. [Google Scholar] [CrossRef]
- Synytsya, A.; Novák, M. Structural Diversity of Fungal Glucans. Carbohydr. Polym. 2013, 92, 792–809. [Google Scholar] [CrossRef] [PubMed]
- Dalonso, N.; Goldman, G.H.; Gern, R.M.M. β-(1→3),(1→6)-Glucans: Medicinal Activities, Characterization, Biosynthesis and New Horizons. Appl. Microbiol. Biotechnol. 2015, 99, 7893–7906. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Du, B.; Bian, Z.; Xu, B. Beta-Glucans from Edible and Medicinal Mushrooms: Characteristics, Physicochemical and Biological Activities. J. Food Compos. Anal. 2015, 41, 165–173. [Google Scholar] [CrossRef]
- Weis, A.L.; Wasser, S.P. Therapeutic Effects of Substances Occurring in Higher Basidiomycetes Mushrooms: A Modern Perspective. Crit. Rev. Immunol. 1999, 19, 65–96. [Google Scholar] [CrossRef]
- Zheng, R.; Su, J.; Dai, H.; Wu, M. Characterization and Immunomodulating Activities of Polysaccharide from Lentinus edodes. Int. Immunopharmacol. 2005, 5, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Boon, H.; Wong, J. Botanical Medicine and Cancer: A Review of the Safety and Efficacy. Expert Opin. Pharmacother. 2004, 5, 2485–2501. [Google Scholar] [CrossRef]
- Sullivan, R.; Smith, J.E.; Rowan, N.J. Medicinal Mushrooms and Cancer Therapy: Translating a Traditional Practice into Western Medicine. Perspect. Biol. Med. 2006, 49, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Yeung, K.; Gubili, J. Shiitake Mushroom (Lentinula edodes). J. Soc. Integr. Oncol. 2008, 6, 134–135. [Google Scholar]
- Samuelsen, A.B.C.; Rise, F.; Wilkins, A.L.; Teveleva, L.; Nyman, A.A.T.; Aachmann, F.L. The Edible Mushroom Albatrellus Ovinus Contains a α-l-Fuco-α-d-Galactan, α-d-Glucan, a Branched (1→6)-β-d-Glucan and a Branched (1→3)-β-d-Glucan. Carbohydr. Res. 2019, 471, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Chen, H.; Zhu, W.; Wang, Z. Effect of Different Drying Methods on Physicochemical Properties and Antioxidant Activities of Polysaccharides Extracted from Mushroom Inonotus Obliquus. Food Res. Int. 2013, 50, 633–640. [Google Scholar] [CrossRef]
- Diamantopoulou, P.; Papanikolaou, S.; Komaitis, M.; Aggelis, G.; Philippoussis, A. Patterns of Major Metabolites Biosynthesis by Different Mushroom Fungi Grown on Glucose-Based Submerged Cultures. Bioprocess Biosyst. Eng. 2014, 37, 1385–1400. [Google Scholar] [CrossRef]
- Xu, X.; Li, J.; Hu, Y. Polysaccharides from Inonotus Obliquus Sclerotia and Cultured Mycelia Stimulate Cytokine Production of Human Peripheral Blood Mononuclear Cells in Vitro and Their Chemical Characterization. Int. Immunopharmacol. 2014, 21, 269–278. [Google Scholar] [CrossRef]
- Chien, R.-C.; Yen, M.-T.; Tseng, Y.-H.; Mau, J.-L. Chemical Characteristics and Anti-Proliferation Activities of Ganoderma Tsugae Polysaccharides. Carbohydr. Polym. 2015, 128, 90–98. [Google Scholar] [CrossRef]
- Su, C.-H.; Lai, M.-N.; Lin, C.-C.; Ng, L.-T. Comparative Characterization of Physicochemical Properties and Bioactivities of Polysaccharides from Selected Medicinal Mushrooms. Appl. Microbiol. Biotechnol. 2016, 100, 4385–4393. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, E.; Krzyczkowski, W.; Herold, F.; Łapienis, G.; Ślusarczyk, J.; Suchocki, P.; Kuraś, M.; Turło, J. Biosynthesis of Selenium-Containing Polysaccharides with Antioxidant Activity in Liquid Culture of Hericium Erinaceum. Enzym. Microb. Technol. 2009, 44, 334–343. [Google Scholar] [CrossRef]
- Bohn, J.A.; BeMiller, J.N. (1→3)-β-d-Glucans as Biological Response Modifiers: A Review of Structure-Functional Activity Relationships. Carbohydr. Polym. 1995, 28, 3–14. [Google Scholar] [CrossRef]
- Okazaki, M.; Adachi, Y.; Ohno, N.; Yadomae, T. Structure-Activity Relationship of (1.RAR.3)-.BETA.-D-Glucans in the Induction of Cytokine Production from Macrophages, in Vitro. Biol. Pharm. Bull. 1995, 18, 1320–1327. [Google Scholar] [CrossRef] [Green Version]
- Surenjav, U.; Zhang, L.; Xu, X.; Zhang, X.; Zeng, F. Effects of Molecular Structure on Antitumor Activities of (1→3)-β-d-Glucans from Different Lentinus edodes. Carbohydr. Polym. 2006, 63, 97–104. [Google Scholar] [CrossRef]
- Ohno, N.; Miura, N.N.; Chiba, N.; Adachi, Y.; Yadomae, T. Comparison of the Immunopharmacological Activities of Triple and Single-Helical Schizophyllan in Mice. Biol. Pharm. Bull. 1995, 18, 1242–1247. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, S.; Wang, X.; Zhang, L.; Cheung, P.C.K. Advances in Lentinan: Isolation, Structure, Chain Conformation and Bioactivities. Food Hydrocoll. 2011, 25, 196–206. [Google Scholar] [CrossRef]
- Górska, S.; Maksymiuk, A.; Turło, J. Selenium-Containing Polysaccharides—Structural Diversity, Biosynthesis, Chemical Modifications and Biological Activity. Appl. Sci. 2021, 11, 3717. [Google Scholar] [CrossRef]
- Malinowska, E.; Klimaszewska, M.; Strączek, T.; Schneider, K.; Kapusta, C.; Podsadni, P.; Łapienis, G.; Dawidowski, M.; Kleps, J.; Górska, S.; et al. Selenized Polysaccharides—Biosynthesis and Structural Analysis. Carbohydr. Polym. 2018, 198, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Turło, J.; Gutkowska, B.; Herold, F. Effect of Selenium Enrichment on Antioxidant Activities and Chemical Composition of Lentinula edodes (Berk.) Pegl. Mycelial Extracts. Food Chem. Toxicol. 2010, 48, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Turlo, J.; Gutkowska, B.; Klimaszewska, M.; Kapusta, C.; Schneider, K.; Sikora, M.; Cieslak, M.; Kazmierczak-Baranska, J.; Gorski, A.; Purchla, S. Selenium-Enriched Polysaccharide Fraction Isolated from Mycelial Culture of Lentinula edodes (Berk.)—Preliminary Analysis of the Structure and Biological Activity. In Proceedings of the 7th International Conference on Mushroom Biology and Mushroom Products (ICMBMP7), Arcachon, France, 4–7 October 2011; pp. 242–246. [Google Scholar]
- Kaleta, B.; Górski, A.; Zagożdżon, R.; Cieślak, M.; Kaźmierczak-Barańska, J.; Nawrot, B.; Klimaszewska, M.; Malinowska, E.; Górska, S.; Turło, J. Selenium-Containing Polysaccharides from Lentinula edodes—Biological Activity. Carbohydr. Polym. 2019, 223, 115078. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, F.; Tomé, D.; Mirand, P.P. Converting Nitrogen into Protein—Beyond 6.25 and Jones’ Factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef]
- Górska, S.; Hermanova, P.; Ciekot, J.; Schwarzer, M.; Srutkova, D.; Brzozowska, E.; Kozakova, H.; Gamian, A. Chemical Characterization and Immunomodulatory Properties of Polysaccharides Isolated from Probiotic Lactobacillus casei LOCK 0919. Glycobiology 2016, 26, 1014–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, E.K.; Maity, K.; Maity, S.; Gantait, S.K.; Behera, B.; Maiti, T.K.; Sikdar, S.R.; Islam, S.S. Chemical Analysis of an Immunostimulating (1→4)-, (1→6)-Branched Glucan from an Edible Mushroom, Calocybe Indica. Carbohydr. Res. 2012, 347, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Chihara, G.; Hamuro, J.; Maeda, Y.Y.; Arai, Y.; Fukuoka, F. Fractionation and Purification of the Polysaccharides with Marked Antitumor Activity, Especially Lentinan, from Lentinus edodes (Berk.) Sing.(an Edible Mushroom). Cancer Res. 1970, 30, 2776–2781. [Google Scholar] [PubMed]
- Yap, A.-T.; Ng, M.-L. An Improved Method for the Isolation of Lentinan from the Edible and Medicinal Shiitake Mushroom, Lentinus edodes (Berk.) Sing.(Agaricomycetideae). Int. J. Med. Mushrooms 2001, 3, 9–19. [Google Scholar] [CrossRef]
- Joye, I.J. Starch. In Encyclopedia of Food Chemistry; Elsevier: Amsterdam, The Netherlands, 2019; pp. 256–264. ISBN 978-0-12-814045-1. [Google Scholar]
- McCleary, B.V.; Draga, A. Measurement of β-Glucan in Mushrooms and Mycelial Products. J. Aoac Int. 2016, 99, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Sjöö, M.; Nilsson, L. Starch in Food: Structure, Function and Applications, 2nd ed.; Woodhead Publishing, an Imprint of Elsevier: Duxford, UK, 2018; pp. 873–893. ISBN 978-0-08-100896-6. [Google Scholar]
- Aimanianda, V.; Clavaud, C.; Simenel, C.; Fontaine, T.; Delepierre, M.; Latgé, J.-P. Cell Wall Beta-(1,6)-Glucan of Saccharomyces cerevisiae: Structural Characterization and in Situ Synthesis. J. Biol. Chem. 2009, 284, 13401–13412. [Google Scholar] [CrossRef] [Green Version]
- Klis, F.M.; Boorsma, A.; De Groot, P.W.J. Cell Wall Construction in Saccharomyces cerevisiae. Yeast 2006, 23, 185–202. [Google Scholar] [CrossRef]
- Lesage, G.; Bussey, H. Cell Wall Assembly in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 317–343. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Cai, C.; Zheng, M.; Hao, J.; Wang, Y.; Hu, M.; Fan, L.; Yu, G. Alkaline Extraction, Structural Characterization, and Bioactivities of (1→6)-β-d-Glucan from Lentinus edodes. Molecules 2019, 24, 1610. [Google Scholar] [CrossRef] [Green Version]
- Lindahl-Kiessling, K.; Peterson, R.D.A. The Mechanism of Phytohemagglutinin (PHA) Action. Exp. Cell Res. 1969, 55, 85–87. [Google Scholar] [CrossRef]
- Van Wauwe, J.P.; De Mey, J.R.; Goossens, J.G. OKT3: A Monoclonal Anti-Human T Lymphocyte Antibody with Potent Mitogenic Properties. J. Immunol. 1980, 124, 2708–2713. [Google Scholar] [CrossRef]
- Burkus, Z.; Temelli, F. Rheological Properties of Barley β-Glucan. Carbohydr. Polym. 2005, 59, 459–465. [Google Scholar] [CrossRef]
- Almond, A. Towards Understanding the Interaction between Oligosaccharides and Water Molecules. Carbohydr. Res. 2005, 340, 907–920. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cui, S.W.; Cheung, P.C.K.; Wang, Q. Antitumor Polysaccharides from Mushrooms: A Review on Their Isolation Process, Structural Characteristics and Antitumor Activity. Trends Food Sci. Technol. 2007, 18, 4–19. [Google Scholar] [CrossRef]
- Turło, J.; Gutkowska, B.; Herold, F.; Gajzlerska, W.; Dawidowski, M.; Dorociak, A.; Zobel, A. Biological Availability and Preliminary Selenium Speciation in Selenium-Enriched Mycelium of Lentinula edodes (Berk.). Food Biotechnol. 2011, 25, 16–29. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Huang, Y.; Peng, H.; Lam, J.W.Y.; Xu, Z.; Leung, F.S.M.; Mays, J.W.; Tang, B.Z. Linear or Branched Structure? Probing Molecular Architectures of Fullerene–Styrene Copolymers by Size Exclusion Chromatographs with Online Right-Angle Laser-Light Scattering and Differential Viscometric Detectors. Polymer 2004, 45, 4811–4817. [Google Scholar] [CrossRef]
- Turło, J.; Gutkowska, B.; Herold, F.; Łuczak, I. Investigation of the Kinetics of Selenium Accumulation by Lentinula Edodes (Berk.) Mycelial Culture by Use of Reversed-Phase High-Performance Liquid Chromatography with Fluorimetric Detection. Acta Chromatogr. 2009, 21, 1–11. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sawardeker, J.S.; Sloneker, J.H.; Jeanes, A. Quantitative Determination of Monosaccharides as Their Alditol Acetates by Gas Liquid Chromatography. Anal. Chem. 1965, 37, 1602–1604. [Google Scholar] [CrossRef]
- Ciucanu, I.; Kerek, F. A Simple and Rapid Method for the Permethylation of Carbohydrates. Carbohydr. Res. 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Gerwig, G.J.; Kamerling, J.P.; Vliegenthart, J.F.G. Determination of the Absolute Configuration of Monosaccharides in Complex Carbohydrates by Capillary g.l.c. Carbohydr. Res. 1979, 77, 1–7. [Google Scholar] [CrossRef] [Green Version]
- SPARKY 3; Vision 3.1; Enhanced Software for Biomolecular NMR Spectroscopy; University of California Research: San Francisco, CA, USA, 2000.








{kind=link}
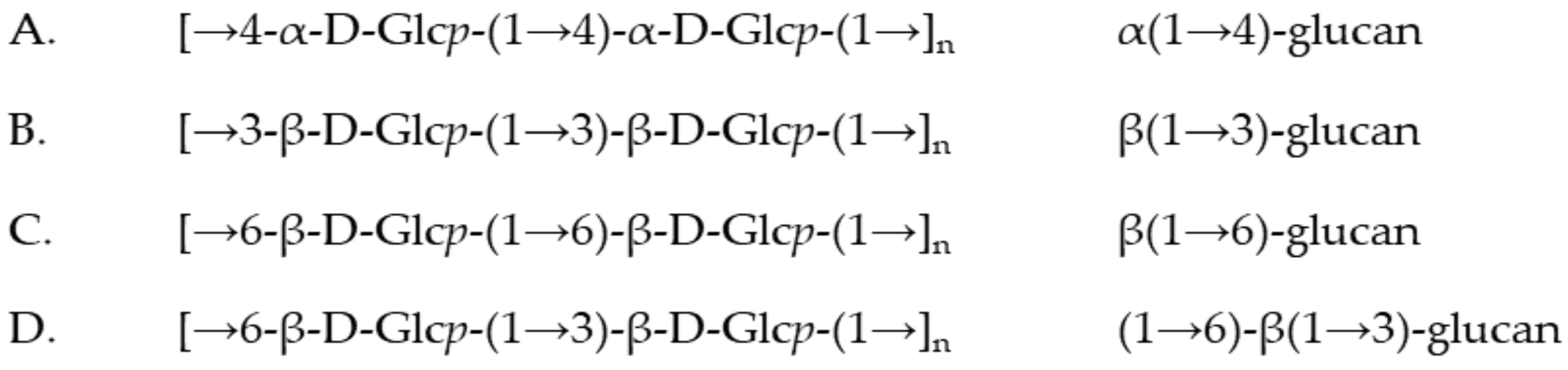{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sugar Residue | Chemical Shifts (ppm) | |||||
|---|---|---|---|---|---|---|
| H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6, H6′ C6 | |
| A →4)-α-d-Glcp-(1→ | 5.33 99.7 | 3.54 71.6 | 3.90 73.4 | 3.57 76.7 | 3.77 71.2 | 3.77, 3.80 60.5 |
| B →3)-β-d-Glcp-(1→ | 4.48 102.6 | 3.45 72.8 | 3.69 84.5 | 3.48 69.2 | 3.46 75.5 | 3.67, 3.85 60.7 |
| C →6)-β-d-Glcp-(1→ | 4.46 103.0 | 3.26 73.1 | 3.39 75.8 | 3.40 69.4 | 3.56 74.8 | 3.80, 4.15 68.8 |
| C’ →6)-β-d-Glcp-(1→ | 4.63 103.0 | 3.31 73.3 | 3.44 75.4 | 3.40 69.5 | 3.56 74.8 | 3.77, 4.13 68.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klimaszewska, M.; Górska, S.; Łapienis, G.; Kaleta, B.; Górska, S.; Kaszowska, M.; Dawidowski, M.; Gamian, A.; Zagożdżon, R.; Górski, A.; et al. Identification of the Primary Structure of Selenium-Containing Polysaccharides Selectively Inhibiting T-Cell Proliferation. Molecules 2021, 26, 5404. https://doi.org/10.3390/molecules26175404
Klimaszewska M, Górska S, Łapienis G, Kaleta B, Górska S, Kaszowska M, Dawidowski M, Gamian A, Zagożdżon R, Górski A, et al. Identification of the Primary Structure of Selenium-Containing Polysaccharides Selectively Inhibiting T-Cell Proliferation. Molecules. 2021; 26(17):5404. https://doi.org/10.3390/molecules26175404
Chicago/Turabian StyleKlimaszewska, Marzenna, Sabina Górska, Grzegorz Łapienis, Beata Kaleta, Sandra Górska, Marta Kaszowska, Maciej Dawidowski, Andrzej Gamian, Radoslaw Zagożdżon, Andrzej Górski, and et al. 2021. "Identification of the Primary Structure of Selenium-Containing Polysaccharides Selectively Inhibiting T-Cell Proliferation" Molecules 26, no. 17: 5404. https://doi.org/10.3390/molecules26175404