Chemical Fingerprinting and Biological Evaluation of the Endemic Chilean Fruit Greigia sphacelata (Ruiz and Pav.) Regel (Bromeliaceae) by UHPLC-PDA-Orbitrap-Mass Spectrometry

 ,
, 

 and
and 
Abstract
:1. Introduction
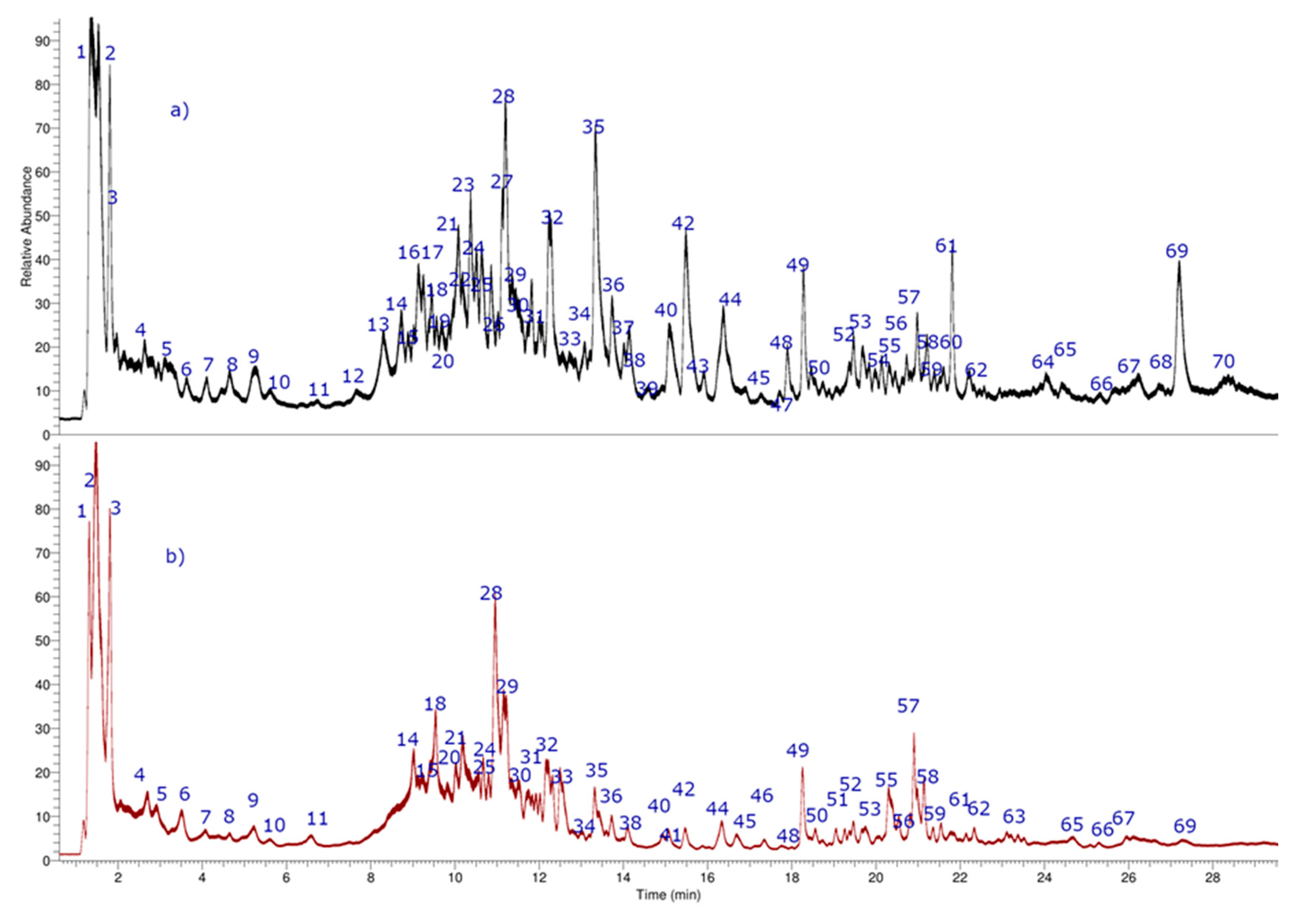
2. Results and Discussion
2.1. Metabolomic Analyses
2.1.1. Coumarins
2.1.2. Flavonoids
2.1.3. Hydrocarbons and Saturated Organic Acids
2.1.4. Oxylipins or Fatty Acids
2.1.5. Iridoids
2.1.6. Terpenes
2.1.7. Benzophenones
2.1.8. Phenolic Acids and Derivatives
2.1.9. Catechins and Proanthocyanidins
2.1.10. Sugar Derivatives
2.2. Total Phenolics, Flavonoid Contents, and Antioxidant Activity of G. sphacelata
2.3. In Vitro Cholinesterase Inhibitory Assay
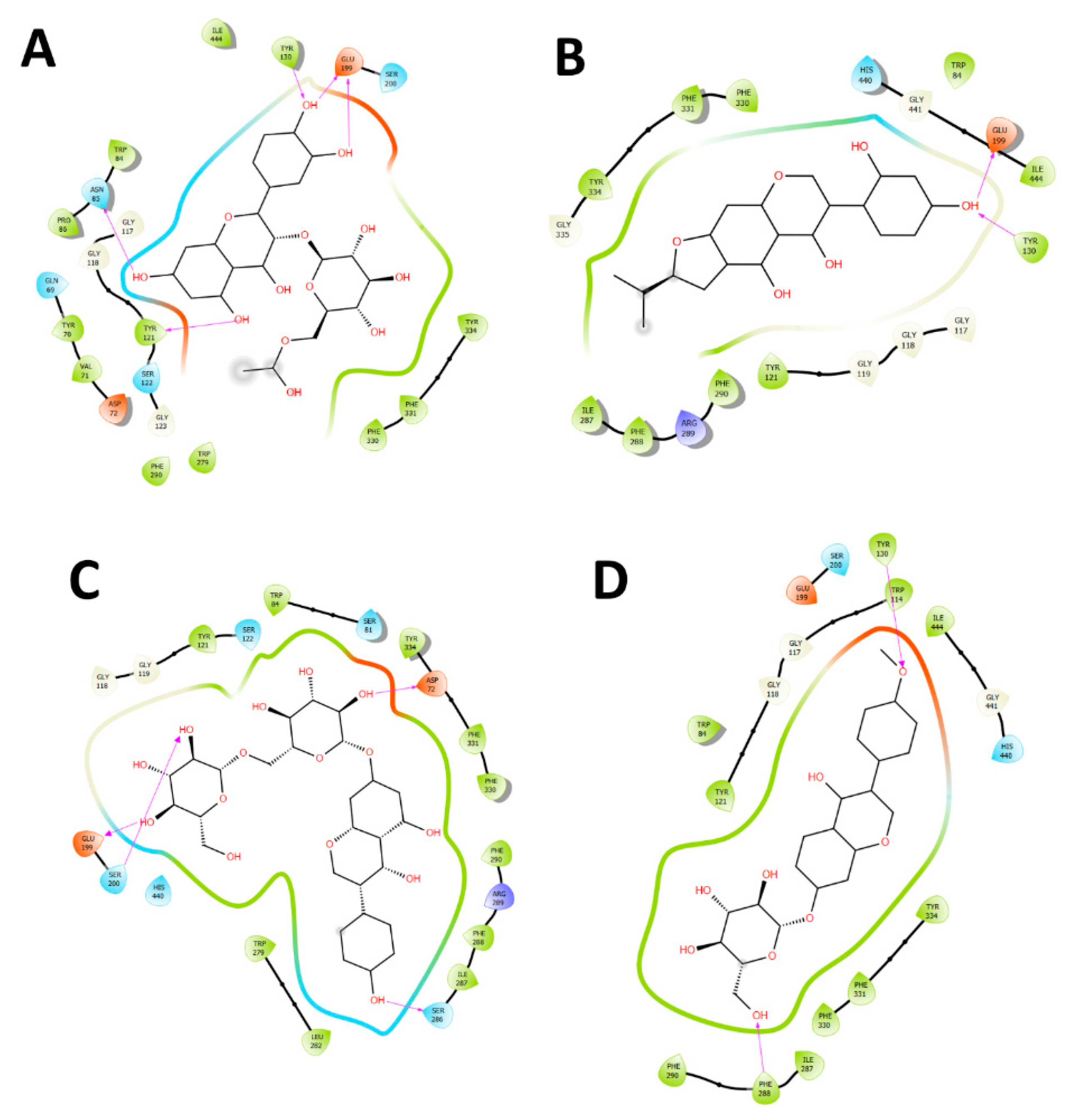
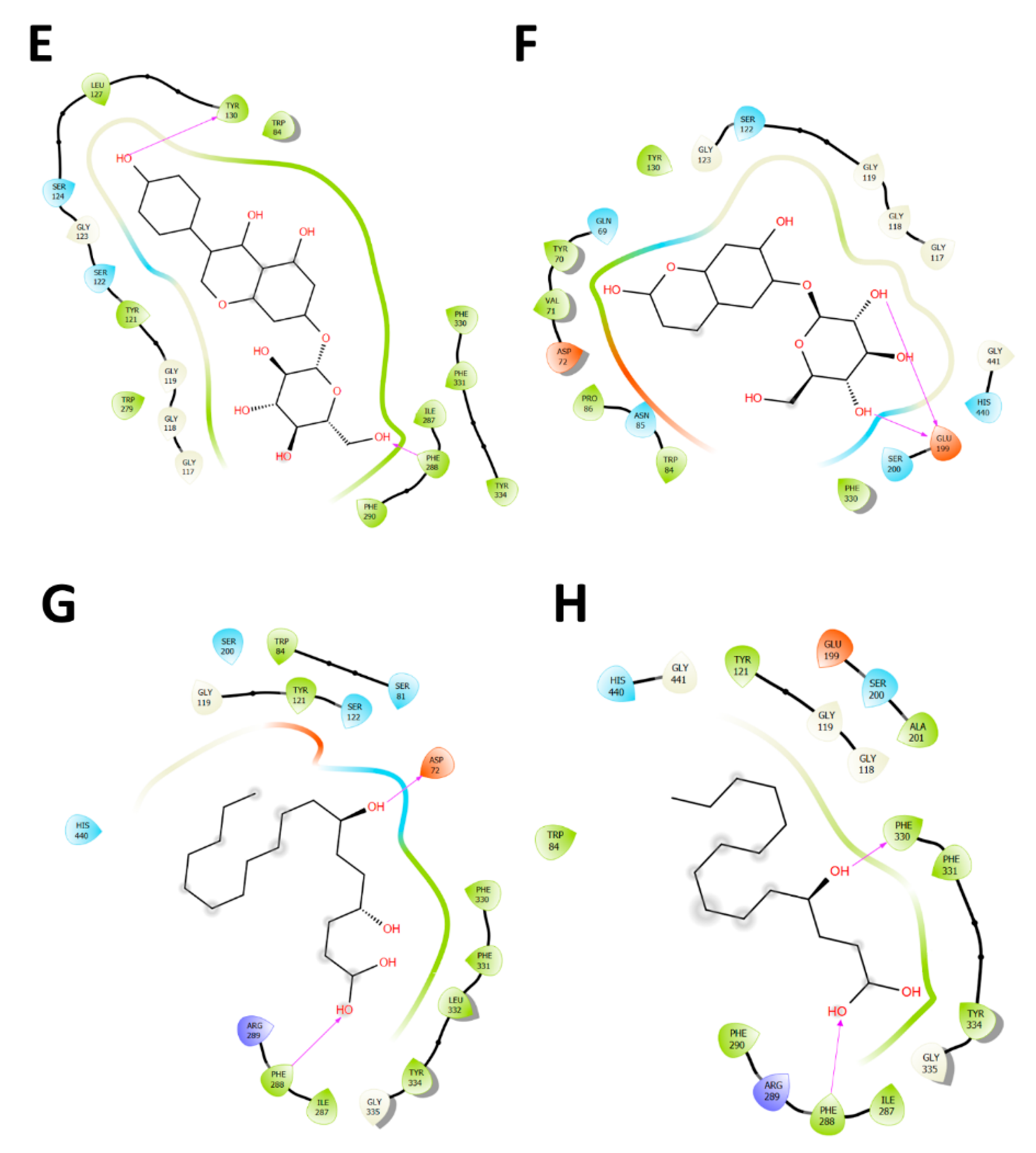
2.4. Docking Studies
2.4.1. Acetylcholinesterase (TcAChE) Docking Results
2.4.2. Butyrylcholinesterase (hBuChE) Docking Results
3. Materials and Methods
3.1. Chemicals and Plant Material
3.2. Fruit Processing
3.3. UHPLC-PDA-Orbitrap-MS
3.4. LC Parameters and MS Parameters
3.5. Total Phenolic (TP) and Total Flavonoid (TF) Content
3.6. Antioxidant Assays
3.6.1. DPPH Cation Radical Discoloration Test
3.6.2. Bleaching Test with the Cationic Radical ABTS•+
3.6.3. Ferric Reduction Ability-Antioxidant Power Test (FRAP)
3.6.4. Superoxide Anion Scavenging Assay
3.7. Cholinesterases (ChE) Inhibitory Activity
3.8. Docking Studies
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Slavin, J.L.; Lloyd, B. Health benefits of fruits and vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, A.W.K.; Aggarwal, B.B.; Barreca, D.; Battino, M.; Belwal, T.; Horbańczuk, O.K.; Berindan-Neagoe, I.; Bishayee, A.; Daglia, M.; Devkota, H.P.; et al. Dietary natural products and their potential to influence health and disease including animal model studies. Anim. Sci. Pap. Rep. 2019, 36, 345–358. [Google Scholar]
- Afrin, S.; Giampieri, F.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Varela-López, A.; Quiles, J.L.; Mezzetti, B.; Battino, M. Chemopreventive and Therapeutic Effects of Edible Berries: A Focus on Colon Cancer Prevention and Treatment. Molecules 2016, 21, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forino, M.; Tartaglione, L.; Dell’Aversano, C.; Ciminiello, P. NMR-based identification of the phenolic profile of fruits of Lycium barbarum (goji berries). Isolation and structural determination of a novel N-feruloyl tyramine dimer as the most abundant antioxidant polyphenol of goji berries. Food Chem. 2016, 194, 1254–1259. [Google Scholar] [CrossRef]
- Belwal, T.; Bisht, A.; Devkota, H.P.; Ullah, H.; Khan, H.; Pandey, A.; Bhatt, I.D.; Echeverría, J. Phytopharmacology and clinical updates of Berberis species against diabetes and other metabolic diseases. Front. Pharm. 2020, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Areche, C.; Hernandez, M.; Cano, T.; Ticona, J.; Cortes, C.; Simirgiotis, M.; Caceres, F.; Borquez, J.; Echeverría, J.; Sepulveda, B. Corryocactus brevistylus (K. Schum. ex Vaupel) Britton & Rose (Cactaceae): Antioxidant, gastroprotective effects, and metabolomic profiling by ultrahigh-pressure liquid chromatography and electrospray high resolution orbitrap tandem mass spectrometry. Front. Pharm. 2020, 11, 417. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, W.; Zhao, J.; Xi, W. Functional constituents and antioxidant activities of eight Chinese native goji genotypes. Food Chem. 2016, 200, 230–236. [Google Scholar] [CrossRef]
- Simirgiotis, M.J.; Ramirez, J.E.; Schmeda Hirschmann, G.; Kennelly, E.J. Bioactive coumarins and HPLC-PDA-ESI-ToF-MS metabolic profiling of edible queule fruits (Gomortega keule), an endangered endemic Chilean species. Food Res. Int. 2013, 54, 532–543. [Google Scholar] [CrossRef]
- Li, J.; Yuan, C.; Pan, L.; Benatrehina, P.A.; Chai, H.; Keller, W.J.; Naman, C.B.; Kinghorn, A.D. Bioassay-guided isolation of antioxidant and cytoprotective constituents from a maqui berry (Aristotelia chilensis) dietary supplement ingredient as markers for qualitative and quantitative analysis. J. Agric. Food Chem. 2017, 65, 8634–8642. [Google Scholar] [CrossRef]
- Will, B.; Zizka, G.A. Review of the genus Greigia Regel (Bromeliaceae) in Chile. Harv. Pap. Bot. 1999, 4, 225–239. [Google Scholar]
- Flagg, M.L.; Wächter, G.A.; Davis, A.L.; Montenegro, G.; Timmermann, B.N. Two novel flavanones from Greigia sphacelata. J. Nat. Prod. 2000, 63, 1689–1691. [Google Scholar] [CrossRef] [PubMed]
- Donno, D.; Beccaro, G.L.; Mellano, M.G.; Cerutti, A.K.; Bounous, G. Goji berry fruit (Lycium spp.): Antioxidant compound fingerprint and bioactivity evaluation. J. Funct. Foods 2015, 18, 1070–1085. [Google Scholar] [CrossRef]
- Simirgiotis, M.J.; Bórquez, J.; Schmeda-Hirschmann, G. Antioxidant capacity, polyphenolic content and tandem HPLC-DAD-ESI/MS profiling of phenolic compounds from the South American berries Luma apiculata and L. chequen. Food Chem. 2013, 139, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Liu, M.; Li, Y.; Wang, K.; Yang, B.; Deng, L.; Tian, J.; Yang, G.; Zheng, G. UPLC-Q-exactive orbitrap MS analysis for identification of lipophilic components in citri sarcodactylis fructus from different origins in China using supercritical CO2 fluid extraction method. ACS Omega 2020, 5, 11013–11023. [Google Scholar] [CrossRef]
- Barrientos, R.; Fernández-Galleguillos, C.; Pastene, E.; Simirgiotis, M.; Romero-Parra, J.; Ahmed, S.; Echeverría, J. Metabolomic analysis, fast isolation of phenolic compounds, and evaluation of biological activities of the bark from Weinmannia trichosperma Cav. (Cunoniaceae). Front. Pharm. 2020, 11, 780. [Google Scholar] [CrossRef]
- Spínola, V.; Mendes, B.; Câmara, J.S.; Castilho, P.C. An improved and fast UHPLC-PDA methodology for determination of L-ascorbic and dehydroascorbic acids in fruits and vegetables. Evaluation of degradation rate during storage. Anal. Bioanal. Chem. 2012, 403, 1049–1058. [Google Scholar]
- Lim, V.; Gorji, S.G.; Daygon, V.D.; Fitzgerald, M. Untargeted and targeted metabolomic profiling of australian indigenous fruits. Metabolites 2020, 10, 114. [Google Scholar] [CrossRef] [Green Version]
- Dinda, B. Pharmacology of Iridoids. In Pharmacology and Applications of Naturally Occurring Iridoids; Dinda, B., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 145–254. ISBN 978-3-030-05575-2. [Google Scholar]
- Chang, J.; Xuan, L.J.; Xu, Y.M.; Zhang, J.S. Seven new sesquiterpene glycosides from the root bark of Dictamnus dasycarpus. J. Nat. Prod. 2001, 64, 935–938. [Google Scholar] [CrossRef]
- Ramirez, J.E.; Zambrano, R.; Sepúlveda, B.; Kennelly, E.J.; Simirgiotis, M.J. Anthocyanins and antioxidant capacities of six Chilean berries by HPLC–HR-ESI-ToF-MS. Food Chem. 2015, 176, 106–114. [Google Scholar] [CrossRef]
- Rodríguez, K.; Ah-Hen, K.; Vega-Gálvez, A.; López, J.; Quispe-Fuentes, I.; Lemus-Mondaca, R.; Gálvez-Ranilla, L. Changes in bioactive compounds and antioxidant activity during convective drying of murta (Ugni molinae T.) berries. Int. J. Food Sci. Technol. 2014, 49, 990–1000. [Google Scholar] [CrossRef]
- Genskowsky, E.; Puente, L.A.; Pérez-Álvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Determination of polyphenolic profile, antioxidant activity and antibacterial properties of maqui [Aristotelia chilensis (Molina) Stuntz] a Chilean blackberry. J. Sci. Food Agric. 2016, 96, 4235–4242. [Google Scholar] [CrossRef] [PubMed]
- Gironés-Vilaplana, A.; Baenas, N.; Villaño, D.; Speisky, H.; García-Viguera, C.; Moreno, D.A. Evaluation of Latin-American fruits rich in phytochemicals with biological effects. J. Funct. Foods 2014, 7, 599–608. [Google Scholar] [CrossRef]
- Xu, J.; Guo, Y.; Xie, C.; Jin, D.-Q.; Gao, J.; Gui, L. Isolation and neuroprotective activities of acylated iridoids from Valeriana jatamansi. Chem. Biodivers. 2012, 9, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y.; Guo, Y.; Guo, P.; Yamakuni, T.; Ohizumi, Y. Isolation, structural elucidation, and neuroprotective effects of iridoids from Valeriana jatamansi. Biosci. Biotechnol. Biochem. 2012, 76, 1401–1403. [Google Scholar] [CrossRef] [Green Version]
- Jugran, A.K.; Rawat, S.; Bhatt, I.D.; Rawal, R.S. Valeriana jatamansi: An herbaceous plant with multiple medicinal uses. Phyther. Res. 2019, 33, 482–503. [Google Scholar] [CrossRef]
- De Jesus, R.A.P.; Cechinel-Filho, V.; Oliveira, A.E.; Schlemper, V. Analysis of the antinociceptive properties of marrubiin isolated from Marrubium vulgare. Phytomedicine 2000, 7, 111–115. [Google Scholar] [CrossRef]
- Teh, S.S.; Ee, G.C.L.; Mah, S.H.; Ahmad, Z. Structure–activity relationship study of secondary metabolites from Mesua beccariana, Mesua ferrea and Mesua congestiflora for anti-cholinesterase activity. Med. Chem. Res. 2016, 25, 819–823. [Google Scholar] [CrossRef]
- Woo, M.H.; Nguyen, D.H.; Choi, J.S.; Park, S.E.; Thuong, P.T.; Min, B.S.; Le, D.D. Chemical constituents from the roots of Kadsura coccinea with their protein tyrosine phosphatase 1B and acetylcholinesterase inhibitory activities. Arch. Pharm. Res. 2020, 43, 204–213. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, G.N.; Kwak, J.H.; Jeong, H.R.; Jeong, C.-H.; Heo, H.J. Neuronal cell protection and acetylcholinesterase inhibitory effect of the phenolics in chestnut inner skin. Food Sci. Biotechnol. 2011, 20, 311–318. [Google Scholar] [CrossRef]
- Gilani, A.H.; Ghayur, M.N.; Saify, Z.S.; Ahmed, S.P.; Choudhary, M.I.; Khalid, A. Presence of cholinomimetic and acetylcholinesterase inhibitory constituents in betel nut. Life Sci. 2004, 75, 2377–2389. [Google Scholar] [CrossRef]
- Floris, S.; Fais, A.; Rosa, A.; Piras, A.; Marzouki, H.; Medda, R.; González-Paramás, A.M.; Kumar, A.; Santos-Buelga, C.; Era, B. Phytochemical composition and the cholinesterase and xanthine oxidase inhibitory properties of seed extracts from the Washingtonia filifera palm fruit. RSC Adv. 2019, 9, 21278–21287. [Google Scholar] [CrossRef] [Green Version]
- Simirgiotis, J.M.; Quispe, C.; Bórquez, J.; Areche, C.; Sepúlveda, B. Fast detection of phenolic compounds in extracts of easter pears (Pyrus communis) from the Atacama Desert by ultrahigh-performance liquid chromatography and mass spectrometry (UHPLC–Q/Orbitrap/MS/MS). Molecules 2016, 21, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simirgiotis, M.J.; Quispe, C.; Bórquez, J.; Schmeda-Hirschmann, G.; Avendaño, M.; Sepúlveda, B.; Winterhalter, P. Fast high resolution Orbitrap MS fingerprinting of the resin of Heliotropium taltalense Phil. from the Atacama Desert. Ind. Crop. Prod. 2016, 85, 159–166. [Google Scholar] [CrossRef]
- Quesada-Romero, L.; Fernández-Galleguillos, C.; Bergmann, J.; Amorós, M.-E.; Jiménez-Aspee, F.; González, A.; Simirgiotis, M.; Rossini, C. Phenolic fingerprinting, antioxidant, and deterrent potentials of Persicaria maculosa Extracts. Molecules 2020, 25, 3054. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Kuskoski, E.M.; Asuero, A.G.; García-Parilla, M.C.; Troncoso, A.M.; Fett, R. Actividad antioxidante de pigmentos antociánicos. Ciênc. E Tecnol. Aliment. 2004, 24, 691–693. [Google Scholar] [CrossRef] [Green Version]
- Benzie, I.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Mocan, A.; Moldovan, C.; Zengin, G.; Bender, O.; Locatelli, M.; Simirgiotis, M.; Atalay, A.; Vodnar, D.C.; Rohn, S.; Crișan, G. UHPLC-QTOF-MS analysis of bioactive constituents from two Romanian Goji (Lycium barbarum L.) berries cultivars and their antioxidant, enzyme inhibitory, and real-time cytotoxicological evaluation. Food Chem. Toxicol. 2018, 115, 414–424. [Google Scholar] [CrossRef]
- Adamo, C.; Barone, V. Toward reliable density functional methods without adjustable parameters: The PBE0 model Seeking for parameter-free double-hybrid functionals: The PBE0-DH model accurate excitation energies from time-dependent density functional theory: Assessing the PBE0 model toward reliable density functional methods without adjustable parameters: The PBE0 model. Cit. J. Chem. Phys. 1999, 110, 2889. [Google Scholar] [CrossRef]
- Petersson, G.A.; Bennett, A.; Tensfeldt, T.G.; Al-Laham, M.A.; Shirley, W.A.; Mantzaris, J. A complete basis set model chemistry. I. The total energies of closed-shell atoms and hydrides of the first-row elements. J. Chem. Phys. 1988, 89, 2193–2218. [Google Scholar] [CrossRef]
Sample Availability: The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Tentative Identification | [M − H]− Ions | Retention Time (min) | UV Max (nm) | Theoretical Mass (m/z) | Measured Mass (m/z) | Accuracy (ppm) | Metabolite Type | MS2 Ions (ppm) | Fruit Part |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Syringaldehyde syringate | C18H17O9− | 1.36 | 315 | 377.08578 | 377.08671 | −2.44 | Pa | 215.03258 | P; S |
| 2 | Quinic acid | C7H11O6− | 1.45 | 191.05501 | 191.05621 | 3.16 | Oa | 135.02919 | P; S | |
| 3 | Citric acid | C6H7O7− | 1.52 | 187 | 191.01863 | 191.01915 | 3.12 | Oa | 162.89121, 111.00782 | P; S |
| 4 | Dehydroascorbic acid | C6H5O6− | 2.20 | 176 | 173.00806 | 173.00879 | 4.18 | Oa | P; S | |
| 5 | Isocitric acid | C6H7O7− | 2.74 | 191.01863 | 191.01923 | 3.76 | Oa | 162.89110, 111.00810 | P; S | |
| 6 | Diffutidin | C17H18O5− | 3.45 | 301.10705 | 301.10614 | −3.02 | Pa | 153.01875 | P; S | |
| 7 | Homocitric acid | C10H17O4− | 4.05 | 205.03542 | 205.03491 | −2.43 | Oa | P; S | ||
| 8 | Vanilloyl-glucoside | C14H17O9− | 4.34 | 287 | 329.08810 | 329.08671 | 4.24 | Pa | 151.03874, 123.02314 | P; S |
| 9 | Euonyminol | C15H25O10− | 5.26 | 365.14422 | 365.14560 | 3.76 | Te | P; S | ||
| 10 | Pantothenic acid | C9H16NO5− | 5.255 | 218.10230 | 218.10313 | 3.82 | Oa | 125.85231 | P; S | |
| 11 | Ebuloside | C21H31O10− | 6.50 | 175 | 443.19263 | 443.19117 | 3.27 | Ir | P; S | |
| 12 | Nonioside k | C17H29O11− | 7.85 | 409.17144 | 409.17191 | 3.58 | Sd | P | ||
| 13 | Glucosyringic acid | C15H19O10− | 8.56 | 325 | 359.09841 | 359.09824 | -0.47 | Pa | 197.0455 (syringic acid) | P |
| 14 | Acetylshikonin | C18H17O6− | 9.01 | 305 | 329.10162 | 329.10114 | −0.47 | Pa | 171.07063 | P; S |
| 15 | Allyl-sucrose | C15H25O11− | 9.12 | 381.14023 | 381.14066 | 2.86 | Sd | P; S | ||
| 16 | Feruloyl-O-galactarate | C16H17O11− | 9.25 | 246–310 | 385.07797 | 385.07654 | 3.64 | Pa | 194.0577, 133.8732 | P |
| 17 | Cinnamoyl catalpol | C24H27O11− | 9.29 | 198 | 491.15589 | 491.15479 | −1.74 | Ir | 175.06085 | P |
| 18 | Cinnamoyl catalpol isomer | C16H21O9− | 9.49 | 198 | 357.11801 | 357.11938 | −1.74 | Ir | 258.09830 | P; S |
| 19 | Linamarin | C10H15NO6− | 9.65 | 246.09821 | 246.09825 | 4.21 | Sd | 258.09830 | P | |
| 20 | Amurensin | C26H29O12− | 10.03 | 202–284 | 533.16431 | 533.16425 | −1.96 | Fl | 503.14331, 341.1015 | P; S |
| 21 | 2-caffeoylisocitric acid | C15H13O10− | 9.98 | 353.07032 | 353.05173 | 3.97 | Pa | 179.06425, 154.99808 | P; S | |
| 22 | Methyl 2,3,4-tris-O-[2-(carboxymethyl) ethyl]- glucose derivative | C26H31O13− | 10.12 | 479.17592 | 479.17749 | 3.27 | Sd | P | ||
| 23 | Methyl 2,3,4-tris-O-[2-(carboxymethyl) ethyl]-glucose | C19H31O12− | 10.43 | 451.18228 | 451.18100 | 2.83 | Sd | P | ||
| 24 | Dimethyl--hydroxyl-oct-2,7-die- noate-glucopyranosyl-glucose | C22H35O14− | 10.55 | 523.20213 | 523.20331 | 2.24 | Sd | 241.07164, 125.02764 | P; S | |
| 25 | Aesculetin-7-O-glucuronide | C15H13O10− | 10.62 | 232–279–329 | 353.05140 | 353.05130 | -0.28 | Co | 177.0193 (Aesculetin) | P; S |
| 26 | Hexamethyl-glucopiranuronosyl-glucoside methyl ester | C19H33O12− | 10.83 | 453.19800 | 453.19794 | 2.96 | Sd | P | ||
| 27 | 7-Hydroxycoumarin glucuronide | C15H13O9− | 10.95 | 255–355 | 337.05541 | 337.05661 | 3.56 | Co | P | |
| 28 | Quercetin-3-O-glucoside-acetate | C23H21O13− | 11.03 | 255–355 | 505.09767 | 505.09912 | 2.87 | Fl | 301.02755 (Quercetin) | P; S |
| 29 | Tectoridin | C22H21O11− | 11.23 | 281 | 461.10894 | 461.10941 | 3.39 | Fl; Is | 300.2632 (tectorigenin) | P; S |
| 30 | Escopoletin 7-O-glucuronide | C16H15O10− | 11.35 | 320 | 367.06597 | 367.06729 | 3.59 | Co | 171.06593 | P; S |
| 31 | Hexenyl-xylopiranosyl-glucose | C17H29O10− | 11.71 | 393.17706 | 393.17552 | 3.91 | Sd | P; S | ||
| 32 | Lupinisoflavone A | C16H15O9− | 12.12 | 285 | 351.07245 | 351.07106 | 3.96 | Fl; Is | 321.0763, 241.0502 | P; S |
| 33 | 2,2,3-Tris (2,3-dihydroxypropanoyl) decanoic acid | C19H31O11− | 12.43 | 435.18609 | 435.18750 | 3.23 | Fa | 287.15002 | P; S | |
| 34 | Trimeric proanthocyanidin C2 | C45H37O18− | 12.96 | 283 | 865.19859 | 865.19451 | −4.71 | Ca | 577.1342, (dimer), 289.0714 catechin, (C15H14O6−) | P; S |
| 35 | Genistein-7-O-di-glucoside | C27H29O15− | 13.20 | 230–238–333 | 593.15122 | 593.15033 | −1.65 | Fl; Is | 269.0452 (genistein) | P; S |
| 36 | Genistein-7-O-di-galactoside | C27H29O15− | 13.52 | 230–238–333 | 593.15120 | 593.15033 | −1.63 | Fl; Is | 269.0451 (genistein) | P; S |
| 37 | Procyanidin B1 | C30H23O12− | 13.83 | 285 | 577.13445 | 577.13373 | 1.16 | Ca | 289.0714 catechin, (C15H14O6−) | P |
| 38 | Genistein-7-O-glucoside | C21H19O10− | 14.03 | 230–238–333 | 431.09847 | 431.09824 | −0.53 | Fl; Is | 269.0452 (genistein) | P; S |
| 39 | Secolonitoside | C21H35O12− | 14.35 | 479.21230 | 479.21350 | 2.50 | Sd | P | ||
| 40 | Trehalose undecylenoate | C23H39O12− | 15.13 | 507.24360 | 507.24454 | 1.84 | Sd | 457.06708 | P; S | |
| 41 | Proanthocyanin tetramer | C60H49O24− | 15.32 | 285 | 1153.26191 | 1153.25146 | −9.05 | Ca | 577.1337 (dimer) | S |
| 42 | Bis-2-hydroxyethyl phtalate | C12H13O6− | 15.52 | 315 | 253.07173 | 253.07066 | 4.21 | Pa | P; S | |
| 43 | Daidzein-7-O-galactoside | C21H19O9− | 15.63 | 281 | 415.10236 | 415.10327 | 3.00 | Fl; Is | 253.05065 | P |
| 44 | Procyanidin A1 | C30H23O12− | 16.45 | 282 | 575.11958 | 575.11877 | −1.42 | Ca | 289.07157 (catechin) | P; Se |
| 45 | Daidzein-7-O-glucoside | C21H19O9− | 16.75 | 230–285–326 | 415.10236 | 415.10361 | 3.00 | Fl; Is | 273.04050 | P; S |
| 46 | Ononin (formononetin 7-O-glucoside) | C22H21O9− | 17.26 | 220–285–326 | 429.11917 | 429.11880 | −0.86 | Fl; Is | 267.06601, 251.0362 (formononetin) | P |
| 47 | 1-O-trans-p-coumaroylglycerol | C12H13O6− | 17.84 | 212–326 | 253.07184 | 253.07150 | −1.34 | Pa | 119.04922, 120.05251 | S |
| 48 | 1, 3-O-di-trans-p-coumaroylglycerol | C21H19O9− | 18.03 | 212–326 | 415.10355 | 415.10333 | 2.86 | Pa | 119.04920, 120.05248 | P; S |
| 49 | Catechin* | C15H13O6− | 18.32 | 285 | 289.07162 | 289.07164 | 0.06 | Fl; Ca | 260.06575, 245.08210 | P; S |
| 50 | Tetrahydroxy-tetradecadienoic acid | C14H23O6− | 18.45 | 265 | 287.15012 | 287.15001 | 4.50 | Fa | P; S | |
| 51 | Monic acid A | C17H27O7− | 19.03 | 215 | 343.17513 | 343.17657 | 4.21 | Te | P; S | |
| 52 | Pentahydroxy-octadecatetranoic acid | C18H27O7− | 19.47 | 189 | 355.17514 | 355.17648 | 3.81 | Fa | Pu; Se | |
| 53 | Dictamnoside N | C18H29O8− | 19.75 | 175 | 373.18569 | 373.18707 | 3.69 | Te | 217.88326 | P; S |
| 54 | Evodinnol | C14H16O4− | 19.83 | 325 | 247.09649 | 247.09760 | 4.49 | Pa | 133.02882 | P |
| 55 | Dihydroxy-pentadecanoic acid | C15H29O4− | 20.36 | 215 | 273.20724 | 273.20734 | 0.36 | Fa | P; S | |
| 56 | 12-hydroxyoctadecanoic acid | C18H36O3− | 20.52 | 225 | 299.25946 | 299.25807 | 4.64 | Fa | P; S | |
| 57 | Congestiflorone | C28H31O4− | 20.98 | 243 | 431.22169 | 431.22061 | −2.49 | Be | 365.17944, 269.17584, 152.99516 | P; S |
| 58 | Hydroxy-pentadecaenoic acid | C15H27O4− | 21.06 | 234 | 271.19144 | 271.19173 | 1.06 | Fa | P; S | |
| 59 | Oct-1-en-3-yl-arabinosyl-glucopyranoside | C19H33O10− | 21.57 | 421.20821 | 421.20816 | −0.11 | Sa | P; S | ||
| 60 | Marrubiin | C20H27O4− | 21.73 | 225 | 331.19196 | 331.19039 | 4.73 | Te | 207.19521 | P |
| 61 | Quillaic acid | C30H45O5− | 21.85 | 214 | 485.32788 | 485.32615 | 3.56 | Te | 405.32125, 249.03424 | P; S |
| 62 | Dihydroxy-octadecaenoic acid | C18H33O4− | 22.05 | 225 | 313.23883 | 313.23734 | −4.76 | Fa | P; S | |
| 63 | Pentadecanedioic acid | C15H27O4− | 23.25 | 235 | 271.19307 | 271.19165 | 4.66 | Fa | S | |
| 64 | Trihydroxy-octadecadienoic acid | C18H31O5− | 24.21 | 235 | 327.21770 | 327.21780 | 0.30 | Fa | P | |
| 65 | Trihydroxy-octadecaenoic acid | C18H33O5− | 24.68 | 222 | 329.23332 | 329.23341 | 0.27 | Fa | P; S | |
| 66 | 2,3-dihydroxypropyl dodecanoate | C15H29O4− | 25.36 | 198 | 273.20715 | 273.20724 | 4.42 | Fa | P; S | |
| 67 | Tetrahydroxy-heptadecatrienoic acid | C17H27O6− | 26.03 | 285 | 327.1813 | 327.18142 | 3.89 | Fa | P; S | |
| 68 | Jatamanvaltrate H | C22H33O9− | 26.19 | 215 | 441.21335 | 441.21329 | 3.26 | Ir | 389.62607, 311.22302, 135.04443 | P |
| 69 | Hydroxy-pentadecanoic acid | C15H29O3− | 27.45 | 235 | 257.21221 | 257.21222 | 0.03 | Fa | P; S | |
| 70 | Unknown | C14H29O8− | 28.33 | 220 | 325.18569 | 325.18442 | −3.92 | Fa | P |
| Sample | TPC a | TFC b | DPPH c | ABTS d | FRAP e | O2− f (%) |
|---|---|---|---|---|---|---|
| Pulp | 45.44 ± 0.67 | 35.57 ± 0.86 | 487.11 ± 26.22 | 190.32 ± 6.23 | 169.08 ± 9.81 | 76.46 ± 3.18 b |
| Seeds | 37.21 ± 0.45 | 28.32 ± 0.35 | 354.51 ± 34.16 | 140.49 ± 3.58 | 147.84 ± 4.35 | 67.02 ± 2.23 b |
| Sample | AChE (IC50) | BChE (IC50) |
|---|---|---|
| Pulp | 4.49 ± 0.08 | 73.86 ± 0.09 |
| Seeds | 4.38 ± 0.04 | 78.57 ± 0.06 |
| Galantamine | 0.27 ± 0.03 | 3.82 ± 0.02 |
| Compound | Binding Energy (kcal/mol) Acetylcholinesterase (TcAChE) | Binding Energy (kcal/mol) Butyrylcholinesterase (hBChE) |
|---|---|---|
| Quercetin-3-O-glucoside-acetate | −9.46 | −8.31 |
| Lupinisoflavone | −9.36 | −7.99 |
| Genistein-7-O-di-glucoside | −9.18 | −6.89 |
| Ononin (formononetin 7-O-glucoside) | −7.45 | −6.44 |
| Genistein-7-O-glucoside | −7.24 | −5.86 |
| Aesculetin-7-O-glucuronide | −6.67 | −6.85 |
| Dihydroxy-octadecaenoic acid | −4.71 | −5.76 |
| Hydroxy-pentadecanoic acid | −4.81 | −4.87 |
| Galantamine | −11.81 | −9.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrientos, R.E.; Ahmed, S.; Cortés, C.; Fernández-Galleguillos, C.; Romero-Parra, J.; Simirgiotis, M.J.; Echeverría, J. Chemical Fingerprinting and Biological Evaluation of the Endemic Chilean Fruit Greigia sphacelata (Ruiz and Pav.) Regel (Bromeliaceae) by UHPLC-PDA-Orbitrap-Mass Spectrometry. Molecules 2020, 25, 3750. https://doi.org/10.3390/molecules25163750
Barrientos RE, Ahmed S, Cortés C, Fernández-Galleguillos C, Romero-Parra J, Simirgiotis MJ, Echeverría J. Chemical Fingerprinting and Biological Evaluation of the Endemic Chilean Fruit Greigia sphacelata (Ruiz and Pav.) Regel (Bromeliaceae) by UHPLC-PDA-Orbitrap-Mass Spectrometry. Molecules. 2020; 25(16):3750. https://doi.org/10.3390/molecules25163750
Chicago/Turabian StyleBarrientos, Ruth E., Shakeel Ahmed, Carmen Cortés, Carlos Fernández-Galleguillos, Javier Romero-Parra, Mario J. Simirgiotis, and Javier Echeverría. 2020. "Chemical Fingerprinting and Biological Evaluation of the Endemic Chilean Fruit Greigia sphacelata (Ruiz and Pav.) Regel (Bromeliaceae) by UHPLC-PDA-Orbitrap-Mass Spectrometry" Molecules 25, no. 16: 3750. https://doi.org/10.3390/molecules25163750