
Antifungal Activity of Phenolic and Polyphenolic Compounds from Different Matrices of Vitis vinifera L. against Human Pathogens


Abstract
:1. Introduction
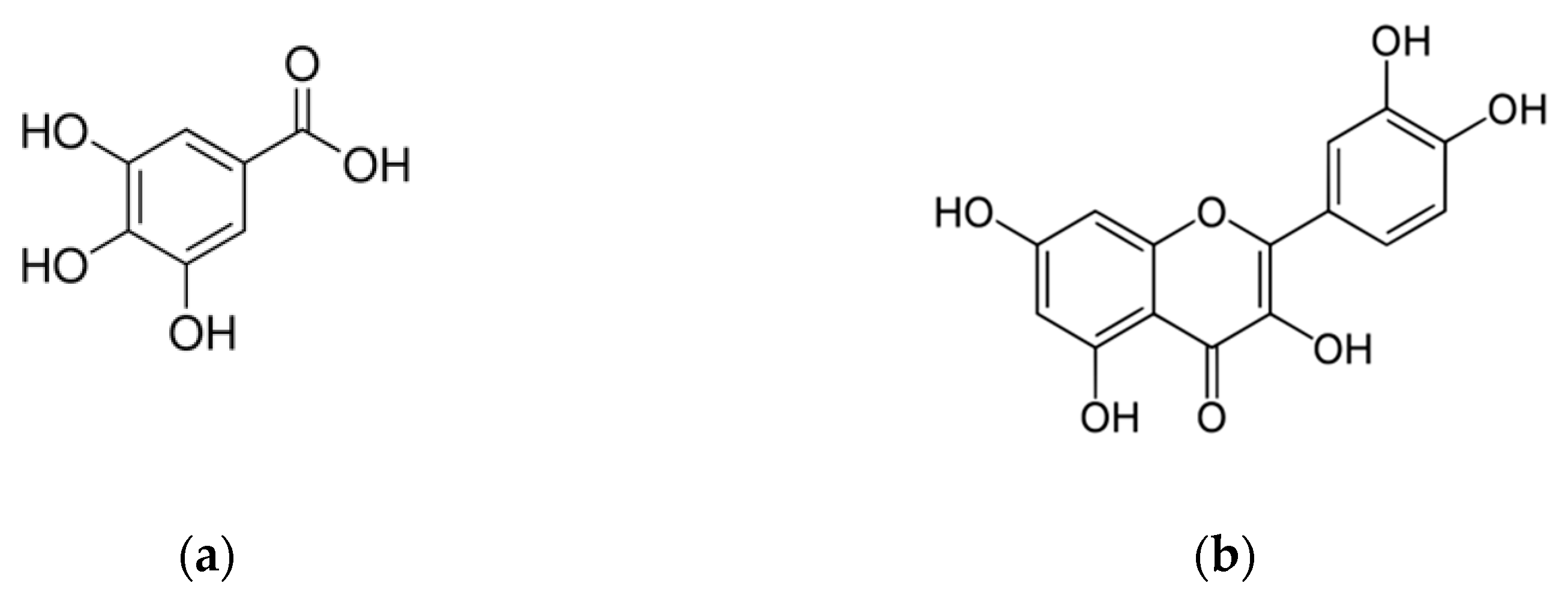
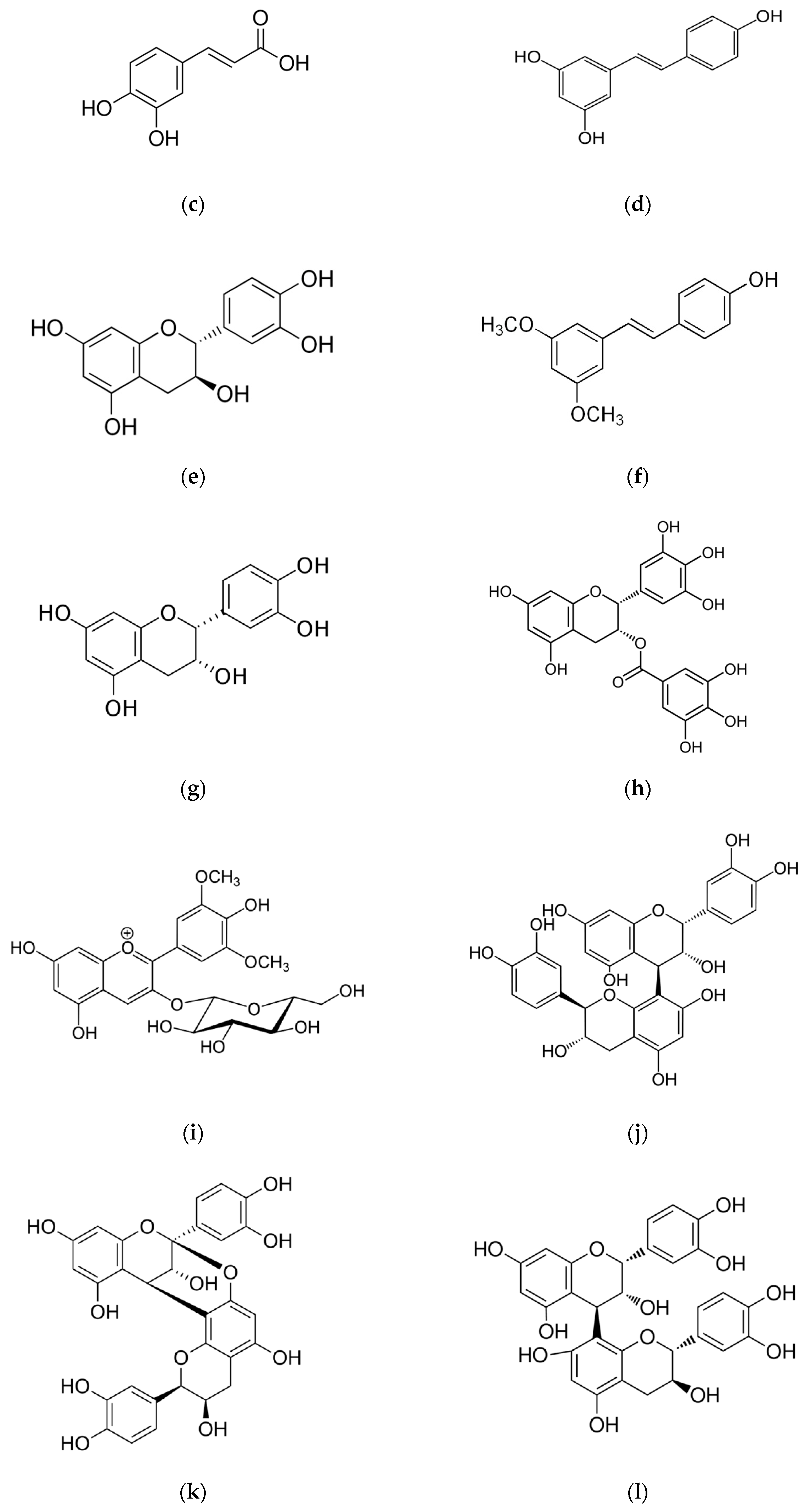
2. Chemical Characterization of Phenols and Polyphenols in Different Matrices of V. vinifera L.: Berry Skins, Seeds, Leaves, Stems, Canes and Not-Fermented and Fermented Pomaces
3. Extraction of Phenolic and Polyphenolic Compounds from V. vinifera and Detection Methods
4. Antifungal Activity of Crude Extracts, Phenolic and Polyphenolic Compounds from Leaves, Stems, Canes of V. vinifera Against Human Pathogenic Fungi
5. Antifungal Activity of Crude Extracts, Phenolic and Polyphenolic Compounds from V. vinifera Unripe Grapes, Berry Skins, Seeds and Not-Fermented and Fermented Pomaces Against Human Pathogenic Fungi
6. Antifungal Mechanisms of Phenolic and Polyphenolic Compounds from V. vinifera
7. Applications
8. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gintjee, T.J.; Donnelley, M.A.; Thompson, G.R. Aspiring Antifungals: Review of Current Antifungal Pipeline Developments. J. Fungi 2020, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, R.S.; Carollo, C.A.; de Magalhães, J.C.; Palumbo, J.M.C.; Boaretto, A.G.; Sá, I.N.; Ferraz, A.C.; Limaa, W.G.; de Siqueira, J.M.; Ferreira, J.M.S. Antibacterial and antifungal activities of phenolic compound-enriched ethyl acetate fraction from Cochlospermum regium (mart. Et. Schr.) Pilger roots: Mechanisms of action and synergism with tannin and gallic acid. S. Afr. J. Bot. 2018, 114, 181–187. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Levitz, S.M. Tackling human fungal infections. Science 2012, 336, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandras, N.; Nostro, A.; Roana, J.; Scalas, D.; Banche, G.; Ghisetti, V.; Del Re, S.; Fucale, G.; Cuffini, A.M.; Tullio, V. Liquid and vapour-phase antifungal activities of essential oils against Candida albicans and non-albicans Candida. BMC Complem. Altern. 2016, 16, 330. [Google Scholar] [CrossRef]
- Tullio, V.; Nostro, A.; Mandras, N.; Dugo, P.; Banche, G.; Cannatelli, M.A.; Cuffini, A.M.; Alonzo, V.; Carlone, N.A. Antifungal activity of essential oils against filamentous fungi determined by broth microdilution and vapour contact methods. J. Appl. Microbiol. 2007, 102, 1544–1550. [Google Scholar] [CrossRef]
- Redondo-Blanco, S.; Fernández, J.; López-Ibáñez, S.; Miguélez, E.; Villar, C.; Lombò, F. Plant phytochemicals in food preservation: Antifungal bioactivity: A review. J. Food Prot. 2020, 83, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Zida, A.; Bamba, S.; Yacouba, A.; Ouedraogo-Traore, R.; Guiguemdé, R.T. Anti-Candida albicans natural products, sources of new antifungal drugs: A review. J. Mycol. Med. 2017, 27, 1–19. [Google Scholar] [CrossRef]
- Friedman, M. Antibacterial, antiviral, and antifungal properties of wines and winery byproducts in relation to their flavonoid content. J. Agric. Food Chem. 2014, 62, 6025–6042. [Google Scholar] [CrossRef]
- Fraternale, D.; Ricci, D.; Verardo, G.; Gorassini, A.; Stocchi, V.; Sestili, P. Activity of Vitis vinifera tendrils extract against phytopathogenic fungi. Nat. Prod. Commun. 2015, 10, 1934578X150100066. [Google Scholar] [CrossRef] [Green Version]
- Olas, B. Berry phenolic antioxidants–implications for human health? Front Pharmacol. 2018, 9, 78. [Google Scholar] [CrossRef]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A concise overview on the chemistry, occurrence, and human health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veiga, M.; Costa, E.M.; Silva, S.; Pintado, M. Impact of plant extracts upon human health: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Panzella, L.; Moccia, F.; Nasti, R.; Marzorati, S.; Verotta, L.; Napolitano, A. Bioactive phenolic compounds from agri-food wastes: An update on green and sustainable extraction methodologies. Front. Nutr. 2020, 7, 60. [Google Scholar] [CrossRef] [PubMed]
- Garrido, J.; Borges, F. Wine and grape polyphenols. A chemical perspective. Food Res. Int. 2013, 54, 1844–1858. [Google Scholar] [CrossRef] [Green Version]
- Haselgrove, L.; Botting, D.; Van Heeswijck, R.; Høj, P.B.; Dry, P.R.; Ford, C.; Land, P.G.I. Canopy microclimate and berry composition: The effect of bunch exposure on the phenolic composition of Vitis vinifera L cv. Shiraz grape berries. Aust. J. Grape Wine Res. 2000, 6, 141–149. [Google Scholar] [CrossRef]
- Simonetti, G.; Santamaria, A.R.; D’Auria, F.D.; Mulinacci, N.; Innocenti, M.; Cecchini, F.; Pericolini, E.; Gabrielli, E.; Panella, S.; Antonacci, D.; et al. Evaluation of anti-Candida activity of Vitis vinifera L. seed extracts obtained from wine and table cultivars. BioMed Res. Int. 2014, 2014, 127021. [Google Scholar] [CrossRef] [Green Version]
- Giannini, B.; Mulinacci, N.; Pasqua, G.; Innocenti, M.; Valletta, A.; Cecchini, F. Phenolics and antioxidant activity in different cultivars/clones of Vitis vinifera L. seeds over two years. Plant Biosyst. 2016, 150, 1408–1416. [Google Scholar] [CrossRef]
- Cheynier, V.; Rigaud, J. HPLC separation and characterization of flavonols in the skins of Vitis vinifera var. Cinsault. Am. J. Enol. Vitic. 1986, 37, 248–252. [Google Scholar]
- Souquet, J.M.; Labarbe, B.; Le Guernevé, C.; Cheynier, V.; Moutounet, M. Phenolic composition of grape stems. J. Agric. Food Chem. 2000, 48, 1076–1080. [Google Scholar] [CrossRef]
- Simonetti, G.; D’Auria, F.D.; Mulinacci, N.; Innocenti, M.; Antonacci, D.; Angiolella, L.; Pasqua, G. Anti-Dermatophyte and Anti-Malassezia Activity of Extracts Rich in Polymeric Flavan-3-ols Obtained from Vitis vinifera Seeds. Phytother. Res. 2017, 31, 124–131. [Google Scholar] [CrossRef]
- Fontana, A.R.; Antoniolli, A.; Bottini, R. Grape pomace as a sustainable source of bioactive compounds: Extraction, characterization, and biotechnological applications of phenolics. J. Agric. Food Chem. 2013, 61, 8987–9003. [Google Scholar] [CrossRef]
- Ghafoor, K.; AL-Juhaimi, F.Y.; Choi, Y.H. Supercritical fluid extraction of phenolic compounds and antioxidants from grape (Vitis labrusca B.) seeds. Plant Foods Hum. Nutr. 2012, 67, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Fuleki, T.; Ricardo da Silva, J.M. Catechin and procyanidin composition of seeds from grape cultivars grown in Ontario. J. Agric. Food Chem. 1997, 45, 1156–1160. [Google Scholar] [CrossRef]
- Cavaliere, C.; Foglia, P.; Marini, F.; Samperi, R.; Antonacci, D.; Laganà, A. The interactive effects of irrigation, nitrogen fertilisation rate, delayed harvest and storage on the polyphenol content in red grape (Vitis vinifera) berries: A factorial experimental design. Food Chem. 2010, 122, 1176–1184. [Google Scholar] [CrossRef]
- Mulinacci, N.; Valletta, A.; Pasqualetti, V.; Innocenti, M.; Giuliani, C.; Bellumori, M.; De Angelis, G.; Carnevale, A.; Locato, V.; Di Venanzio, C.; et al. Effects of ionizing radiation on bio-active plant extracts useful for preventing oxidative damages. Nat. Prod. Res. 2019, 33, 1106–1114. [Google Scholar] [CrossRef]
- Dinicola, S.; Cucina, A.; Pasqualato, A.; D’Anselmi, F.; Proietti, S.; Lisi, E.; Pasqua, G.; Antonacci, D.; Bizzarri, M. Antiproliferative and apoptotic effects triggered by grape seed extract (GSE) versus epigallocatechin and procyanidins on colon cancer cell lines. Int. J. Mol. Sci. 2012, 13, 651–664. [Google Scholar] [CrossRef]
- Giovannelli, L.; Innocenti, M.; Santamaria, A.R.; Bigagli, E.; Pasqua, G.; Mulinacci, N. Antitumoural activity of viniferin-enriched extracts from Vitis vinifera L. cell cultures. Nat. Prod. Res. 2014, 28, 2006–2016. [Google Scholar] [CrossRef]
- Pasqua, G.; Simonetti, G. Antimicrobial and antiviral activities of grape seed extracts. In Grape Seeds; Nova Science Publishers: New York, NY, USA, 2016; pp. 211–224. [Google Scholar]
- Goufo, P.; Singh, R.K.; Cortez, I. A Reference List of Phenolic Compounds (Including Stilbenes) in Grapevine (Vitis vinifera L.) Roots, Woods, Canes, Stems, and Leaves. Antioxidants 2020, 9, 398. [Google Scholar] [CrossRef]
- Moldovan, M.L.; Bogdan, C.; Lurian, S.; Roman, C.; Oniga, I.; Benedec, D. Phenolic content and antioxidant capacity of pomace and canes extracts of some Vitis vinifera varieties cultivated in Romania. Farmacia 2020, 68, 15–21. [Google Scholar] [CrossRef]
- Moldovan, M.L.; Carpa, R.; Fizeșan, I.; Vlase, L.; Bogdan, C.; Iurian, S.M.; Benedec, D.; Pop, A. Phytochemical Profile and Biological Activities of Tendrils and Leaves Extracts from a Variety of Vitis vinifera L. Antioxidants 2020, 9, 373. [Google Scholar] [CrossRef]
- Di Lorenzo, R.; Gambino, C.; Scafidi, P. Summer pruning in table grape. Adv. Hortic. Sci. 2011, 143–150. [Google Scholar]
- Simonetti, G.; D’Auria, F.D.; Mulinacci, N.; Milella, R.A.; Antonacci, D.; Innocenti, M.; Pasqua, G. Phenolic content and in vitro antifungal activity of unripe grape extracts from agro-industrial wastes. Nat. Prod. Res. 2019, 33, 803–807. [Google Scholar] [CrossRef]
- Lucarini, M.; Durazzo, A.; Kiefer, J.; Santini, A.; Lombardi-Boccia, G.; Souto, E.B.; Romani, A.; Lampe, A.; Ferrari Nicoli, S.; Gabrielli, P.; et al. Grape seeds: Chromatographic profile of fatty acids and phenolic compounds and qualitative analysis by FTIR-ATR spectroscopy. Foods 2020, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Centeno, M.R.; Rosselló, C.; Simal, S.; Garau, M.C.; López, F.; Femenia, A. Physico-chemical properties of cell wall materials obtained from ten grape varieties and their byproducts: Grape pomaces and stems. LWT Food Sci. Technol. 2010, 43, 1580–1586. [Google Scholar] [CrossRef]
- Monrad, J.K.; Howard, L.R.; King, J.W.; Srinivas, K.; Mauromoustakos, A. Subcritical solvent extraction of procyanidins from dried red grape pomace. J. Agric. Food Chem. 2010, 58, 4014–4021. [Google Scholar] [CrossRef]
- González-Centeno, M.R.; Knoerzer, K.; Sabarez, H.; Simal, S.; Rosselló, C.; Femenia, A. Effect of acoustic frequency and power density on the aqueous ultrasonic-assisted extraction of grape pomace (Vitis vinifera L.) a response surface approach. Ultrason. Sonochem. 2014, 21, 2176–2184. [Google Scholar] [CrossRef]
- García-Lomillo, J.; González-San José, M.L. Applications of wine pomace in the food industry: Approaches and functions. Compr. Rev. Food Sci. F 2017, 16, 3–22. [Google Scholar] [CrossRef]
- Cheng, V.J.; Bekhit, A.E.D.A.; McConnell, M.; Mros, S.; Zhao, J. Effect of extraction solvent, waste fraction and grape variety on the antimicrobial and antioxidant activities of extracts from wine residue from cool climate. Food Chem. 2012, 134, 474–482. [Google Scholar] [CrossRef]
- Rinaldi, A.; Jourdes, M.; Teissedre, P.L.; Moio, L. A preliminary characterization of Aglianico (Vitis vinifera L. cv.) grape proanthocyanidins and evaluation of their reactivity towards salivary proteins. Food Chem. 2014, 164, 142–149. [Google Scholar] [CrossRef]
- González-Centeno, M.R.; Jourdes, M.; Femenia, A.; Simal, S.; Rosselló, C.; Teissedre, P.L. Proanthocyanidin composition and antioxidant potential of the stem winemaking byproducts from 10 different grape varieties (Vitis vinifera L.). J. Agric. Food Chem. 2012, 60, 11850–11858. [Google Scholar] [CrossRef]
- Yilmaz, Y.; Toledo, R.T. Oxygen radical absorbance capacities of grape/wine industry byproducts and effect of solvent type on extraction of grape seed polyphenols. J. Food Compos. Anal. 2006, 19, 41–48. [Google Scholar] [CrossRef]
- Youssef, D.; El-Adawi, H. Study on grape seeds extraction and optimization: An approach. J. Appl. Sci. 2006, 6, 2944–2947. [Google Scholar]
- Vergara-Salinas, J.R.; Bulnes, P.; Zúñiga, M.C.; Pérez-Jiménez, J.; Torres, J.L.; Mateos-Martín, M.L.; Agosin, E.; Pérez-Correa, J.R. Effect of pressurized hot water extraction on antioxidants from grape pomace before and after enological fermentation. J. Agric. Food Chem. 2013, 61, 6929–6936. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, H.; Shi, J.; Mittal, G.S.; Kakuda, Y. Extraction of polyphenols from grape seeds and concentration by ultrafiltration. Sep. Purif. Technol. 2006, 48, 176–181. [Google Scholar] [CrossRef]
- Lafka, T.-I.; Sinanoglou, V.; Lazos, E.S. On the extraction and antioxidant activity of phenolic compounds from winery wastes. Food Chem. 2007, 104, 1206–1214. [Google Scholar] [CrossRef]
- Makris, D.P.; Boskou, G.; Andrikopoulos, N.K. Recovery of antioxidant phenolics from white vinification solid by-products employing water/ethanol mixtures. Biores. Technol. 2007, 98, 2963–2967. [Google Scholar] [CrossRef]
- Bucić-Kojić, A.; Planinić, M.; Tomas, S.; Bilić, M.; Velić, D. Study of solid–liquid extraction kinetics of total polyphenols from grape seeds. J. Food Eng. 2007, 81, 236–242. [Google Scholar] [CrossRef]
- Oliveira, D.A.; Salvador, A.A.; Smânia, A., Jr.; Smânia, E.F.; Maraschin, M.; Ferreira, S.R. Antimicrobial activity and composition profile of grape (Vitis vinifera) pomace extracts obtained by supercritical fluids. J. Biotechnol. 2013, 164, 423–432. [Google Scholar] [CrossRef]
- Eslami, H.; Babaei, H.; Mehrbani, S.P.; Aghazadeh, M.; Babaei, Z.; Nezhad, S.K. Evaluation of antifungal effect of grape seed extract (GSE) on Candida glabrata and Candida krusei: In vitro study. Biomed. Res. 2017, 28, 9163–9170. [Google Scholar]
- Va, M.; Eb, J.E. The Journal of Free Radicals and Antioxidants. Photon 2016, 143, 440–445. [Google Scholar]
- Kolouchová, I.; Maťátková, O.; Paldrychová, M.; Kodeš, Z.; Kvasničková, E.; Sigler, K.; Čejková, A.; Šmidrkal, J.; Demnerová, K.; Masák, J. Resveratrol, pterostilbene, and baicalein: Plant-derived anti-biofilm agents. Folia Microbiol. 2018, 63, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Ruhnke, M. Clinical Syndromes: Candida and Candidosis. In Clinically Relevant Mycoses; Springer: Cham, Switzerland, 2019; pp. 45–75. [Google Scholar]
- Simonetti, G.; Palocci, C.; Valletta, A.; Kolesova, O.; Chronopoulou, L.; Donati, L.; Di Nitto, A.; Brasili, E.; Tomai, P.; Gentili, A.; et al. Anti-Candida biofilm activity of pterostilbene or crude extract from non-fermented grape pomace entrapped in biopolymeric nanoparticles. Molecules 2019, 24, 2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.J.; Hwang, I.A.; Sung, W.S.; Kang, H.; Kang, B.S.; Seu, Y.B.; Lee, D.G. Fungicidal effect of resveratrol on human infectious fungi. Arch. Pharm. Res. 2005, 28, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.M.Y. Antimicrobial effect of resveratrol on dermatophytes and bacterial pathogens of the skin. Biochem. Pharmacol. 2002, 63, 99–104. [Google Scholar] [CrossRef]
- Paulo, L.; Oleastro, M.; Gallardo, E.; Queiroz, J.A.; Domingues, F. Antimicrobial properties of resveratrol: A review. In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances; Formatex Research Center: Badajoz, Spain, 2011; Volume 2, pp. 1225–1235. [Google Scholar]
- Khurana, A.; Sardana, K.; Chowdhary, A. Antifungal resistance in dermatophytes: Recent trends and therapeutic implications. Fungal Genet Biol. 2019, 132, 103255. [Google Scholar] [CrossRef]
- Ventoulis, I.; Sarmourli, T.; Amoiridou, P.; Mantzana, P.; Exindari, M.; Gioula, G.; Vyzantiadis, T.A. Bloodstream Infection by Saccharomyces cerevisiae in Two COVID-19 Patients after Receiving Supplementation of Saccharomyces in the ICU. J. Fungi 2020, 6, 98. [Google Scholar] [CrossRef]
- Hay, R.J.; Moore, M.K. Mycology. In Rook’s Textbook of Dermatology, 7th ed.; Burns, T., Breathnach, S., Cox, N., Griffiths, C., Eds.; Blackwell Science: London, UK, 2004; pp. 31.16–31.85. [Google Scholar]
- Houille, B.; Papon, N.; Boudesocque, L.; Bourdeaud, E.; Besseau, S.; Courdavault, V.; Clastre, M. Antifungal activity of resveratrol derivatives against Candida species. J. Nat. Prod. 2014, 77, 1658–1662. [Google Scholar] [CrossRef]
- Weber, K.; Schulz, B.; Ruhnke, M. Resveratrol and its antifungal activity against Candida species. Mycoses 2011, 54, 30–33. [Google Scholar] [CrossRef]
- Barac, A.; Kosmidis, C.; Alastruey-Izquierdo, A.; Salzer, H.J. Chronic pulmonary aspergillosis update: A year in review. Med. Mycol. 2019, 57, S104–S109. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Miri, A.; Sharifi-Rad, M.; Sharifi-Rad, M.; Hoseini-Alfatemi, S.M.; Yazdanpanah, E. Antifungal and antibacterial properties of Grapevine (Vitis vinifera L.) leaves methanolic extract from Iran-in vitro study. Am. Eurasian J. Agric. Environ. Sci. 2014, 14, 1312–1316. [Google Scholar]
- Jediyi, H.; Naamani, K.; Ait Elkoch, A.; Dihazi, A.; Lemjiber, N. A comparative study of phenols composition, antioxidant, and antifungal potency of leaves extract from five Moroccan Vitis vinifera L. varieties. J. Food Saf. 2020, 40, e12756. [Google Scholar] [CrossRef]
- Manimaran, M.; Sivakumari, K.; Ashok, K.; Rajesh, S. Evaluation of the in vitro antimicrobial effect of resveratrol on human pathogens. Evaluation 2017, 2, 124–127. [Google Scholar]
- Filip, V.; Plockova, M.; Šmidrkal, J.; Špičková, Z.; Melzoch, K.; Schmidt, Š. Resveratrol and its antioxidant and antimicrobial effectiveness. Food Chem. 2003, 83, 585–593. [Google Scholar] [CrossRef]
- Peixoto, C.M.; Dias, M.I.; Alves, M.J.; Calhelha, R.C.; Barros, L.; Pinho, S.P.; Ferreira, I.C. Grape pomace as a source of phenolic compounds and diverse bioactive properties. Food Chem. 2018, 253, 132–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yıgıt, D.; Yıgıt, N.; Mavı, A.; Yıldırım, A.; Güleryüz, M. Antioxidant and antimicrobial activities of methanol and water extracts of fruits, leaves and seeds of Vitis vinifera L. cv. Karaerik. Asian J. Chem. 2009, 21, 183–194. [Google Scholar]
- Kumar, K.A.; Vijayalakshmi, K. In vitro anti-microbial activity and phytochemical analysis of selected fruit wastes. Int. J. Curr. Microbiol. App. Sci. 2013, 2, 196–204. [Google Scholar]
- Kiraly-Veghely, Z.; Moricz, A.M.; Ott, P.G.; Katay, G.; Belai, I.; Tyihák, E. Comparison of components from red and white wines for antimicrobial activity by biodetection after OPLC separation. J. Liq. Chromatogr. R. T 2009, 32, 1259–1272. [Google Scholar] [CrossRef]
- Han, Y. Synergic effect of grape seed extract with amphotericin B against disseminated candidiasis due to Candida albicans. Phytomedicine 2007, 14, 733–738. [Google Scholar] [CrossRef]
- Esposito, E.; Campolo, M.; Casili, G.; Lanza, M.; Filippone, A.; Peritore, A.F.; Cuzzocrea, S. Effect of pea protein plus grape seed dry extract on a murine model of Candida albicans induced vaginitis. Future Microbiol. 2018, 13, 1375–1382. [Google Scholar] [CrossRef]
- Shrivastav, V.K.; Shukla, D.; Parashar, D.; Shrivastav, A. Dermatophytes and related keratinophilic fungi isolated from the soil in Gwalior region of India and in vitro evaluation of antifungal activity of the selected plant extracts against these fungi. J. Med. Plant Res. 2013, 7, 2136–2139. [Google Scholar]
- Gupta, A.K.; Batra, R.; Bluhm, R.; Boekhout, T.; Dawson, T.L., Jr. Skin diseases associated with Malassezia species. J. Am. Acad. Dermatol. 2004, 51, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Saunte, D.M.; Gaitanis, G.; Hay, R.J. Malassezia-Associated Skin Diseases, the Use of Diagnostics and Treatment. Front. Cell. Infect. Microbiol. 2020, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Versteeg, S.G. Topical treatment of facial seborrheic dermatitis: A systematic review. Am. J. Clin. Dermatol. 2017, 18, 193–213. [Google Scholar] [CrossRef]
- Pintas, S.K.; Quave, C.L. A Review of Botanicals Exhibiting Antifungal Activity Against Malassezia spp. Implicated in Common Skin Conditions. Curr. Dermatol. Rep. 2019, 8, 279–296. [Google Scholar] [CrossRef]
- Yadav, D.; Kumar, A.; Kumar, P.; Mishra, D. Antimicrobial properties of black grape (Vitis vinifera L.) peel extracts against antibiotic-resistant pathogenic bacteria and toxin producing molds. Indian J. Pharmacol. 2015, 47, 663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghouila, Z.; Laurent, S.; Boutry, S.; Vander Elst, L.; Nateche, F.; Muller, R.N.; Baaliouamer, A. Antioxidant, antibacterial and cell toxicity effects of polyphenols Fromahmeur bouamer grape seed extracts. J. Fundam. Appl. Sci. 2017, 9, 392–420. [Google Scholar] [CrossRef]
- Gago, S.; Denning, D.W.; Bowyer, P. Pathophysiological aspects of Aspergillus colonization in disease. Med. Mycol. 2019, 57, S219–S227. [Google Scholar] [CrossRef] [PubMed]
- Marquez, L.; Quave, C.L. Prevalence and Therapeutic Challenges of Fungal Drug Resistance: Role for Plants in Drug Discovery. Antibiotics 2020, 9, 150. [Google Scholar] [CrossRef] [Green Version]
- Pasqua, G.; Simonetti, G.; D’Auria, F.D.; Santamaria, A.R.; Antonacci, D. Extracts Obtained from Seeds and/or Skins of Vitis vinifera and Related Uses as Antifungal Agents. Italy Patent No. IT1407378B1, 4 April 2014. [Google Scholar]
- Li, D.D.; Zhao, L.X.; Mylonakis, E.; Hu, G.H.; Zou, Y.; Huang, T.K.; Yan, L.; Wang, Y.; Jiang, Y.Y. In vitro and in vivo activities of pterostilbene against Candida albicans biofilms. Antimicrob. Agents Chemother. 2014, 58, 2344–2355. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, D.G. Novel antifungal mechanism of resveratrol: Apoptosis inducer in Candida albicans. Curr. Microbiol. 2015, 70, 383–389. [Google Scholar] [CrossRef]
- Navarro-Martínez, M.D.; García-Cánovas, F.; Rodriguez-Lopez, J.N. Tea polyphenol epigallocatechin-3-gallate inhibits ergosterol synthesis by disturbing folic acid metabolism in Candida albicans. J. Antimicrob. Chemother. 2006, 57, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.J.; Liu, M.; Dawuti, G.; Dou, Q.; Ma, Y.; Liu, H.G.; Aibai, S. Antifungal activity of gallic acid in vitro and in vivo. Phytother. Res. 2017, 31, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, T.A.; Komoto, T.T.; Massaroto, B.G.; Miranda, C.E.S.; Beleboni, R.O.; Marins, M.; Fachin, A.L. Trans-chalcone and quercetin down-regulate fatty acid synthase gene expression and reduce ergosterol content in the human pathogenic dermatophyte Trichophyton rubrum. BMC Complement. Altern. Med. 2013, 13, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheah, H.L.; Lim, V.; Sandai, D. Inhibitors of the glyoxylate cycle enzyme ICL1 in Candida albicans for potential use as antifungal agents. PLoS ONE 2014, 9, e95951. [Google Scholar] [CrossRef] [Green Version]
- Feldman, M.; Tanabe, S.; Howell, A.; Grenier, D. Cranberry proanthocyanidins inhibit the adherence properties of Candida albicans and cytokine secretion by oral epithelial cells. BMC Complement. Altern. Med. 2012, 12, 6. [Google Scholar] [CrossRef] [Green Version]
- Surini, S.; Mubarak, H.; Ramadon, D. Cosmetic serum containing grape (Vitis vinifera L.) seed extract phytosome: Formulation and in vitro penetration study. J. Young Pharm. 2018, 10, S51. [Google Scholar] [CrossRef] [Green Version]
- Valletta, A.; Salvatori, E.; Rita Santamaria, A.; Nicoletti, M.; Toniolo, C.; Caboni, E.; Pasqua, G.; Manes, F. Ecophysiological and phytochemical response to ozone of wine grape cultivars of Vitis vinifera L. Nat. Prod. Res. 2016, 30, 2514–2522. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, M.; Appendino, G.; Efferth, T.; Fürst, R.; Izzo, A.A.; Kayser, O.; Pezzuto, J.M.; Viljoen, A. Best practice in research–overcoming common challenges in phytopharmacological research. J. Ethnopharmacol. 2020, 246, 1122. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |



{kind=link}
{kind=link}
| Flavan-3-ols | Matrices and Cultivars | Extraction | Solvent | Detection | Range | Ref. |
|---|---|---|---|---|---|---|
| Procyanidin A2 | Pomace; cultivar Pinot noir | ME | Methanol/water 50:50 (v/v) | LC–MS | 7.80 mg/g of extract | [39] |
| Pomace; cultivar Pinot Meunir | ME | Methanol/water 50:50 (v/v) | LC–MS | 11.37 mg/g of extract | [39] | |
| Procyanidin B1 | Skin; cultivar Michele Palieri | ME | Methanol/water/formic acid 70:30:1 (v/v/v) | LC–MS | 184.5–247.8 mg/kg WW | [24] |
| Skin; cultivar Red Globe | ME | Methanol/water/formic acid 70:30:1 (v/v/v) | LC–MS | 133.5–160.7 mg/kg WW | [24] | |
| Skin; cultivar Aglianico | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.035 mg/g DW | [40] | |
| Skin; cultivar Merlot | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.034 mg/g DW | [40] | |
| Skin; cultivar Cabernet Sauvignon | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.038 mg/g DW | [40] | |
| Seed; cultivar Michele Palieri | ME | Methanol/water/formic acid 70:30:1 (v/v/v) | LC–MS | 1263.1–1643.8 mg/kg WW | [24] | |
| Seed: cultivar Red Globe | ME | Methanol/water/formic acid 70:30:1 (v/v/v) | LC–MS | 414.8–905.2 mg/kg WW | [24] | |
| Seed; cultivar Aglianico | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.065 mg/g DW | [40] | |
| Seed; cultivar Merlot | ME | Acetone/water 80:20 (v/v) Methanol/water (60:40, v/v) | HPLC/ESI/MS | 0.059 mg/g DW | [40] | |
| Seed; cultivar Cabernet Sauvignon | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.061 mg/g DW | [40] | |
| Stem; cultivar Syrah | ASE | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC-UV-fluo | 132.0 mg/100 g DW | [41] | |
| Stem; cultivar Tempranillo | ASE | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC-UV-fluo | 195.8 mg/100 g DW | [41] | |
| Stem; cultivar Parellada | ASE | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC-UV-fluo | 187.7 mg/100 g DW | [41] | |
| Stem; cultivar Premsal Blanc | ASE | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC-UV-fluo | 121.8 mg/100 g DW | [41] | |
| Procyanidin B2 | Skin; cultivar Michele Palieri | ME | Methanol/water/formic acid 70:30:1 (v/v/v) | LC–MS | 17.1–27.4 mg/kg WW | [24] |
| Skin; cultivar Red Globe | ME | Methanol/water/formic acid 70:30:1 (v/v/v) | LC–MS | 20.2–27.7 mg/kg WW | [24] | |
| Skin; cultivar Aglianico | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.036 mg/g DW | [40] | |
| Skin; cultivar Merlot | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.045 mg/g DW | [40] | |
| Skin; cultivar Cabernet Sauvignon | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.043 mg/g DW | [40] | |
| Seed; cultivar Michele Palieri | ME | Methanol/water/formic acid 70:30:1 (v/v/v) | LC–MS | 1203.6–1636.4 mg/kg WW | [24] | |
| Seed; cultivar Red Globe | ME | Methanol/water/formic acid 70:30:1 (v/v/v) | LC–MS | 394.0–883.1 mg/kg WW | [24] | |
| Seed; cultivar Aglianico | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.078 mg/g DW | [40] | |
| Seed; cultivar Merlot | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.085 mg/g DW | [40] | |
| Seed; cultivar Cabernet Sauvignon | ME | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC/ESI/MS | 0.083 mg/g DW | [40] | |
| Stem; cultivar Tempranillo | ASE | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC-UV-fluo | 9.4 mg/100 g DW | [41] | |
| Stem; cultivar Premsal Blanc | ASE | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC-UV-fluo | 4.0 mg/100 g DW | [41] | |
| Total flavan-3-ols | Pomace; cultivar Labrusca | ASE | Ethanol/water 50:50 (v/v) | HPLC-ESI-MS/MS | 3253–5708 mg/100 g DW | [36] |
| Stem; cultivar Syrah | ASE | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC-UV-fluo | 269.7 mg/100 g DW | [41] | |
| Stem; cultivar Tempranillo | ASE | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC-UV-fluo | 366.3 mg/100 g DW | [41] | |
| Stem; cultivar Premsal Blanc | ASE | Acetone/water 80:20 (v/v) Methanol/water 60:40 (v/v) | HPLC-UV-fluo | 214.2 mg/100 g DW | [41] | |
| Seed; cultivar Michele Palieri | ME | Ethanol/water/formic acid 70:30:1 (v/v/v) | HPLC/ESI/MS | 638.5 mg/g of extract | [16] | |
| Unripe grape; cultivar Don Mariano | ME | Ethanol/water/formic acid 70:30:1 (v/v/v) | HPLC/ESI/MS | 2666 µg/g FW | [33] | |
| Unripe grape; cultivar Alphonse Lavalle | ME | Ethanol/water/formic acid 70:30:1 (v/v/v) | HPLC/ESI/MS | 6773 µg/g FW | [33] | |
| Unripe grape; cultivar Italia | ME | Ethanol/water/formic acid 70:30:1 (v/v/v) | HPLC/ESI/MS | 3390 µg/g FW | [33] | |
| Unripe grape; cultivar Michele Palieri | ME | Ethanol/water/formic acid 70:30:1 (v/v/v) | HPLC/ESI/MS | 3596 µg/g FW | [33] | |
| Unripe grape; cultivar Red Globe | ME | Ethanol/water/formic acid 70:30:1 (v/v/v) | HPLC/ESI/MS | 2300 µg/g FW | [33] | |
| Unripe grape; cultivar Almeria | ME | Ethanol/water/formic acid 70:30:1 (v/v/v) | HPLC/ESI/MS | 4149 µg/g FW | [33] |
| Matrices | Extraction | Detection | Compounds | Microorganism | MIC µg/mL | Ref. | |
|---|---|---|---|---|---|---|---|
| Seeds | Cultivar Michele Palieri Turi (BA) Italy, 2010 treated with water 2000 m3/hectare and nitrogen fertilization 120 kg/hectare | Ethanol/water 70:30 (v/v) | HPLC/ESI/MS | Flavan-3-ols 820 mg/g | Candida albicans ATCC 10231 | 8 | [16] |
| HPLC/ESI/MS | Candida krusei DSM 6128 | 4 | [16] | ||||
| HPLC/ESI/MS | Candida parapsilosis ATCC 22019 | 8 | [16] | ||||
| HPLC/ESI/MS | Candida glabrata PMC 0805 | 4 | [16] | ||||
| HPLC/ESI/MS | Candida tropicalis DSM 11953 | 16 | [16] | ||||
| HPLC/DAD/ESI/MS | Trichophyton mentagrophytes DSM 4870 | 32 | [20] | ||||
| HPLC/DAD/ESI/MS | Microsporum gypseum PMC 7331 | 64 | [20] | ||||
| HPLC/DAD/ESI/MS | Microsporum canis PMC 7426 | 64 | [20] | ||||
| Cultivar Michele Palieri Turi (BA) Italy, 2012 | Ethanol/water | HPLC/DAD/ESI/MS | Flavan-3-ols 383.4 mg/g | Malassezia furfur DSM 6171 | 16 | [20] | |
| Ethanol | HPLC/DAD/ESI/MS | Flavan-3-ols 272.0 mg/g | Malassezia furfur DSM 6171 | 64 | [20] | ||
| Methanol | HPLC/DAD/ESI/MS | Flavan-3-ols 288.2 mg/g | Malassezia furfur DSM 6171 | 32 | [20] | ||
| Nd | Ethanol/water 70:30 (v/v) | Nd | Nd | Candida glabrata BSM 11226 | 50 | [50] | |
| Nd | Ethanol/water 70:30 (v/v) | Nd | Nd | Candida krusei BSM 70079 | 50 | [50] | |
| Wine residue obtained after 1 week of pre-fermentation from Pinot noir, Lincoln University, Canterbury, New Zealand, between 2008 and 2009 | Ethanol/water 50:50 (v/v) | LC–MS | Catechin 14.31 mg/g | Candida albicans ATCC 10261 | 390 | [39] | |
| Acetone/water 50:50 (v/v) | LC–MS | Catechin 13.41mg/g | Candida albicans ATCC 10261 | 780 | [39] | ||
| Methanol/water 50:50 (v/v) | LC–MS | Catechin 15.18 mg/g | Candida albicans ATCC 10261 | 780 | [39] | ||
| Nd | Ethanol | Nd | Nd | Candida albicans MTCC 227 | 1000 | [70] | |
| Nd | Ethanol | Nd | Nd | Candida tropicalis MTCC 184 | 500 | [70] | |
| Nd | Ethanol | Nd | Nd | Cryptococcus neoformans clinical isolate | 1000 | [70] | |
| Pomace | Wine residue obtained after 1 week of pre-fermentation from Pinot noir, Lincoln University, Canterbury, New Zealand, between 2008 and 2009 | Ethanol/water 50:50 (v/v) | LC–MS | Catechin 8.97 mg/g | Candida albicans ATCC 10261 | 390 | [39] |
| Acetone/water 50:50 (v/v) | LC–MS | Catechin 10.01 mg/g | Candida albicans ATCC 10261 | >1000 | [39] | ||
| Methanol/water 50:50 (v/v) | LC–MS | Catechin 11.81 mg/g | Candida albicans ATCC 10261 | >1000 | [39] | ||
| Merlot pomace, vintage 2009, Vale dos Vinhedos–Rio Grande do Sul, Brazil | Supercritical fluid extraction SFE SC-CO2 50 °C/200 bar | RF-HPLC-UV | Gallic acid 1159 | Candida albicans ATCC 14053 | 500 | [49] | |
| RF-HPLC-UV | Candida parapsilosis ATCC 22019 | 1000 | [49] | ||||
| RF-HPLC-UV | Candida krusei ATCC 6258 | 500 | [49] | ||||
| non-fermented pomace cultivar Italia grown in the experimental farm of Crea-Utv, Turi (Bari) Italy | Ethanol/water 70:30 (v/v) | HPLC/ESI/MS | Procyanidins 90.67 mg/g | Candida albicans ATCC 10231 | 12.5 | [54] | |
| HPLC/ESI/MS | Candida albicans 3135 | 6.25 | [54] | ||||
| HPLC/ESI/MS | Candida albicans ATCC 20891 | 1.4 | [54] | ||||
| HPLC/ESI/MS | Candida albicans ATCC 10261 | 12.5 | [54] | ||||
| Skin | Wine residue obtained after 1 week of pre-fermentation from Pinot noir, Lincoln University, Canterbury, New Zealand, between 2008 and 2009 | Ethanol/water 50:50 (v/v) | LC–MS | Malvidin-3-glucoside 8.36 mg/g | Candida albicans ATCC 10261 | 780 | [39] |
| Acetone/water 50:50 (v/v) | LC–MS | Malvidin-3-glucoside 10.64 mg/g | Candida albicans ATCC 10261 | 780 | [39] | ||
| Methanol/water 50:50 (v/v) | LC–MS | Malvidin-3-glucoside 9.95 mg/g | Candida albicans ATCC10261 | 780 | [39] | ||
| Unripe grape | Alphonse Lavalle experimental farm of Crea-Utv, Turi (Bari) Italy | Ethanol/water 70:30 (v/v) | HPLC/ESI/MS | Polymeric flavan-3-ols 3896 μg/g FW | Candida albicans ATCC 10231 | 64 | [33] |
| Candida krusei DSM 6128 | 64 | [33] | |||||
| Candida glabrata PMC 0805 | 64 | [33] | |||||
| Candida parapsilosis ATCC 22019 | 64 | [33] | |||||
| Candida tropicalis DSM 11953 | 128 | [33] | |||||
| Microsporum gypseum PMC 7331 | 16 | [33] | |||||
| Triciphyton mentagrophytes PMC 6503 | 64 | [33] | |||||
| Tricophyton rubrum PMC 6612 | 64 | [33] | |||||
| Leaves | 2014 from Zabol region (Coordinates: 31°1′43″ N, 61°30′4″ E), in Sistan and Baluchestan Provinces of Iran. | Methanol | Nd | Nd | Candida albicans ATCC 10231 | 80.9 | [64] |
| Nd | Nd | Aspergillus niger ATCC 9142 | 75.4 | [64] |
| Chemical Compounds | Molecular Weight |
|---|---|
| Gallic acid | 170.1 |
| Caffeic acid | 180.2 |
| Catechin | 290.3 |
| Epicatechin | 290.3 |
| Epicatechin-3-gallate | 442.4 |
| Quercetin | 448.4 |
| Resveratrol | 228.2 |
| Pterostilbene | 256.3 |
| Malvidin-3-O-glucoside | 493.4 |
| Procyanidin A2 | 576.5 |
| Procyanidin B1 | 578.5 |
| Procyanidin B2 | 578.5 |
| Compounds | Fungi | Antifungal Mechanisms | Ref. |
|---|---|---|---|
| Pterostilbene | Candida albicans | Down regulator of Ras/cAMP pathway and the ergosterol biosynthesis. | [84] |
| Resveratrol | Candida albicans | Activator of metacaspase and cytochrome c release inducing apoptosis | [85] |
| Epigallocatechin-3-gallate | Candida albicans | Inhibitor of dihydrofolate reductase (Ki = 0.7 µM). Synergy with inhibitors of the ergosterol biosynthesis pathway | [86] |
| Gallic acid | Trichophyton rubrum | Inhibitor of ergosterol biosynthesis reducing the activity of sterol 14α-demethylase P450 (CYP51) and squalene epoxidase | [87] |
| Quercetin | Trichophyton rubrum | Reduces ergosterol levels and modulates the expression of FAS1 and ERG6 | [88] |
| Caffeic acid | Candida albicans | Inhibits isocitrate lyase enzyme activity | [89] |
| Proanthocyanidins | Candida albicans | Reduce the adherence properties of fungus attenuating the inflammatory response, interfering with NF-κB p65 activation and the phosphorylation of specific signal intracellular kinases. | [90] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simonetti, G.; Brasili, E.; Pasqua, G. Antifungal Activity of Phenolic and Polyphenolic Compounds from Different Matrices of Vitis vinifera L. against Human Pathogens. Molecules 2020, 25, 3748. https://doi.org/10.3390/molecules25163748
Simonetti G, Brasili E, Pasqua G. Antifungal Activity of Phenolic and Polyphenolic Compounds from Different Matrices of Vitis vinifera L. against Human Pathogens. Molecules. 2020; 25(16):3748. https://doi.org/10.3390/molecules25163748
Chicago/Turabian StyleSimonetti, Giovanna, Elisa Brasili, and Gabriella Pasqua. 2020. "Antifungal Activity of Phenolic and Polyphenolic Compounds from Different Matrices of Vitis vinifera L. against Human Pathogens" Molecules 25, no. 16: 3748. https://doi.org/10.3390/molecules25163748