
On the Emergence of the Deviation from a Poisson Law in Stochastic Mathematical Models for Radiation-Induced DNA Damage: A System Size Expansion

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- (i)
- to derive a system size expansion for the master equation governing GSMstudying both the macroscopic limit and the fluctuations around the average;
- (ii)
- to show how the nonlinear terms accounting for DNA clustering give rise to a non-Poissonian behavior;
- (iii)
- to shed light on another insightful connection between existing radiobiological models.
2. Material and Methods
2.1. The Generalized Stochastic Microdosimetric Model and the Microdosimetric Kinetic Model
3. Theory and Calculations
3.1. Macroscopic Description for the GSM
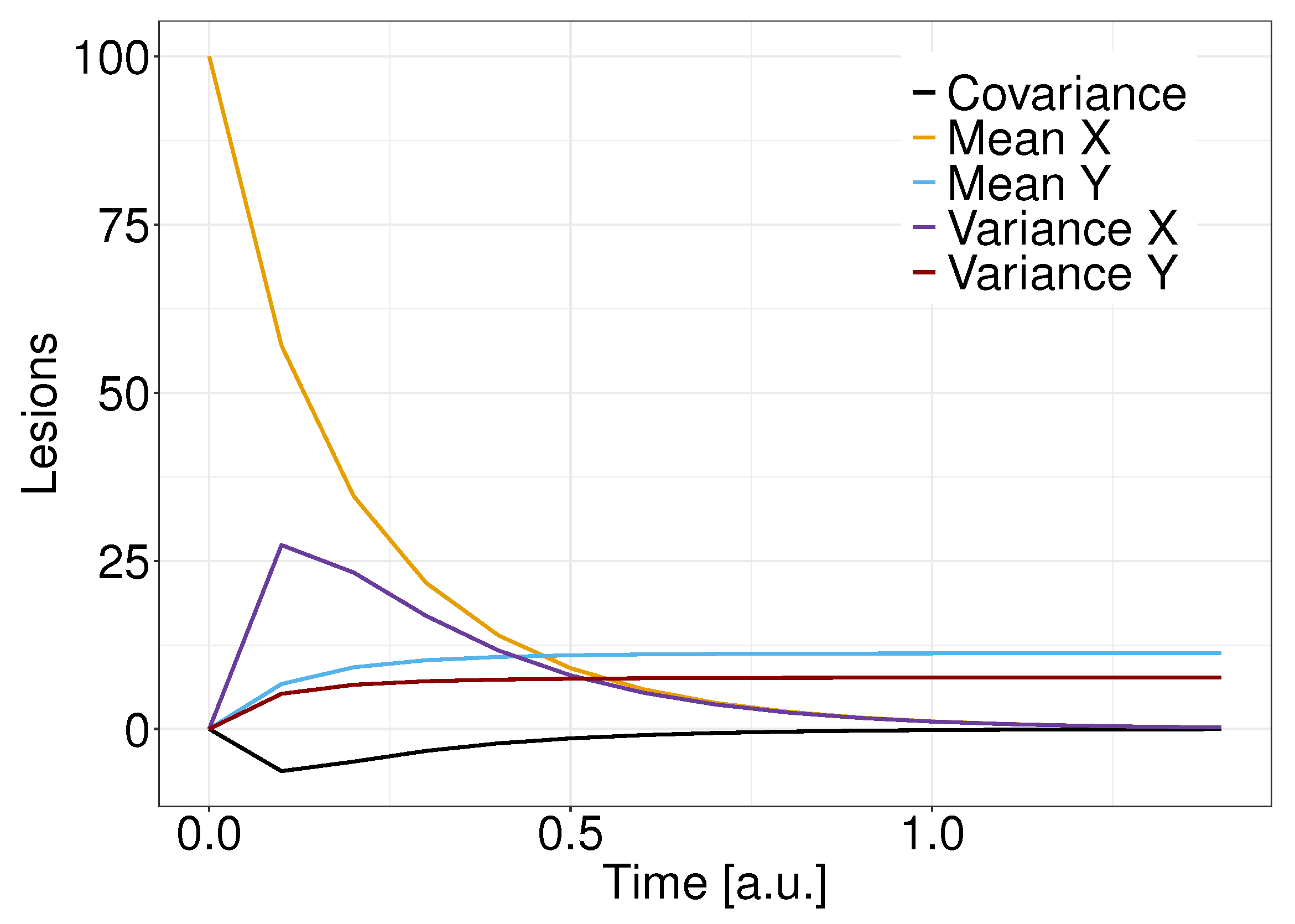
3.2. The Linear Noise Approximation and Moments Estimates
Moments Estimates for a Stochastic Initial Condition
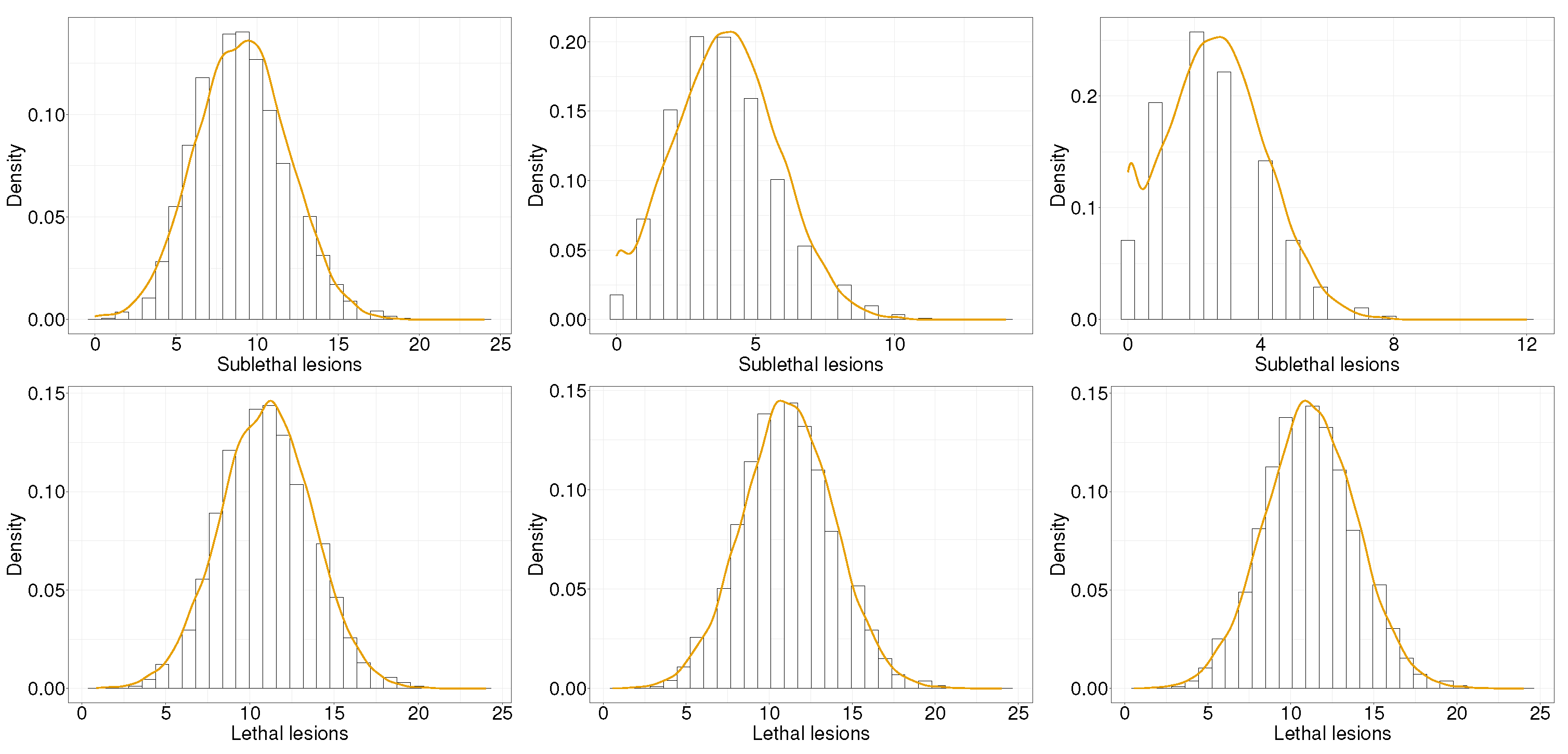
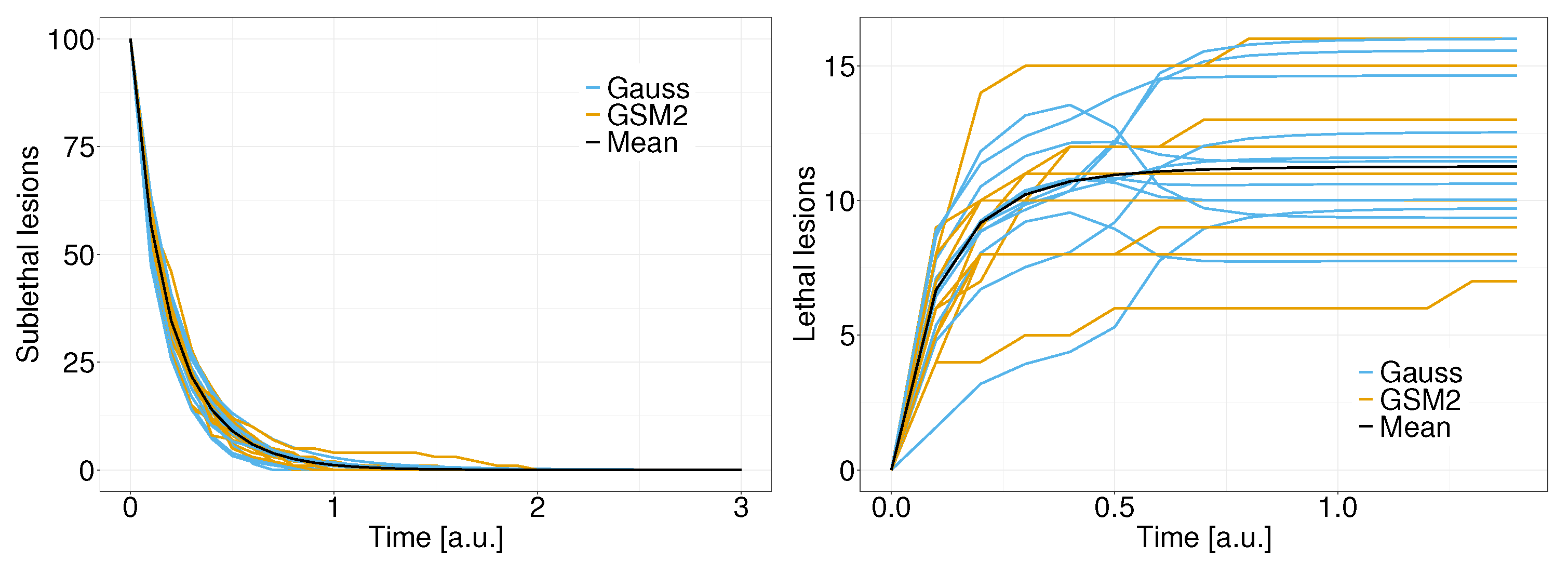
4. Results
5. Discussion
6. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thariat, J.; Hannoun-Levi, J.M.; Sun Myint, A.; Vuong, T.; Gérard, J.P. Past, present, and future of radiotherapy for the benefit of patients. Nat. Rev. Clin. Oncol. 2013, 10, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Paganetti, H. Nuclear physics in particle therapy: A review. Rep. Prog. Phys. 2016, 79, 096702. [Google Scholar] [CrossRef]
- Durante, M.; Loeffler, J.S. Charged particles in radiation oncology. Nat. Rev. Clin. Oncol. 2010, 7, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Bellinzona, V.; Cordoni, F.; Missiaggia, M.; Tommasino, F.; Scifoni, E.; La Tessa, C.; Attili, A. Linking Microdosimetric Measurements to Biological Effectiveness in Ion Beam Therapy: A review of theoretical aspects of MKM and other models. Front. Phys. 2021, 8, 578492. [Google Scholar] [CrossRef]
- Hawkins, R.B. A statistical theory of cell killing by radiation of varying linear energy transfer. Radiat. Res. 1994, 140, 366–374. [Google Scholar] [CrossRef]
- Hawkins, R.B.; Inaniwa, T. A microdosimetric-kinetic model for cell killing by protracted continuous irradiation including dependence on LET I: Repair in cultured mammalian cells. Radiat. Res. 2013, 180, 584–594. [Google Scholar] [CrossRef]
- Kellerer, A.M.; Rossi, H.H. The theory of dual radiation action. In Current Topics in Radiation Research Quarterly; North-Holland Publishing Company: Amsterdam, The Netherlands, 1974; pp. 85–158. Available online: https://www.osti.gov/biblio/4611340 (accessed on 13 June 2023).
- Herr, L.; Friedrich, T.; Durante, M.; Scholz, M. A comparison of kinetic photon cell survival models. Radiat. Res. 2015, 184, 494–508. [Google Scholar] [CrossRef]
- Pfuhl, T.; Friedrich, T.; Scholz, M. Prediction of cell survival after exposure to mixed radiation fields with the local effect model. Radiat. Res. 2020, 193, 130–142. [Google Scholar] [CrossRef]
- Cordoni, F.; Missiaggia, M.; Attili, A.; Welford, S.; Scifoni, E.; La Tessa, C. Generalized stochastic microdosimetric model: The main formulation. Phys. Rev. E 2021, 103, 012412. [Google Scholar] [CrossRef]
- Hawkins, R.B. A microdosimetric-kinetic model for the effect of non-Poisson distribution of lethal lesions on the variation of RBE with LET. Radiat. Res. 2003, 160, 61–69. [Google Scholar] [CrossRef]
- Scholz, M.; Kellerer, A.; Kraft-Weyrather, W.; Kraft, G. Computation of cell survival in heavy ion beams for therapy. Radiat. Environ. Biophys. 1997, 36, 59–66. [Google Scholar] [CrossRef]
- Friedrich, T.; Scholz, U.; Elsässer, T.; Durante, M.; Scholz, M. Calculation of the biological effects of ion beams based on the microscopic spatial damage distribution pattern. Int. J. Radiat. Biol. 2012, 88, 103–107. [Google Scholar] [CrossRef]
- Hawkins, R.B. A microdosimetric-kinetic model of cell killing by irradiation from permanently incorporated radionuclides. Radiat. Res. 2018, 189, 104–116. [Google Scholar] [CrossRef]
- Inaniwa, T.; Suzuki, M.; Furukawa, T.; Kase, Y.; Kanematsu, N.; Shirai, T.; Hawkins, R.B. Effects of dose-delivery time structure on biological effectiveness for therapeutic carbon-ion beams evaluated with microdosimetric kinetic model. Radiat. Res. 2013, 180, 44–59. [Google Scholar] [CrossRef]
- Sato, T.; Furusawa, Y. Cell survival fraction estimation based on the probability densities of domain and cell nucleus specific energies using improved microdosimetric kinetic models. Radiat. Res. 2012, 178, 341–356. [Google Scholar] [CrossRef]
- Kase, Y.; Kanai, T.; Matsumoto, Y.; Furusawa, Y.; Okamoto, H.; Asaba, T.; Sakama, M.; Shinoda, H. Microdosimetric measurements and estimation of human cell survival for heavy-ion beams. Radiat. Res. 2006, 166, 629–638. [Google Scholar] [CrossRef]
- Cordoni, F.G.; Missiaggia, M.; Scifoni, E.; La Tessa, C. Cell Survival Computation via the Generalized Stochastic Microdosimetric Model (GSM2); Part I: The Theoretical Framework. Radiat. Res. 2022, 197, 218–232. [Google Scholar] [CrossRef]
- Cordoni, F.G.; Missiaggia, M.; La Tessa, C.; Scifoni, E. Multiple levels of stochasticity accounted for in different radiation biophysical models: From physics to biology. Int. J. Radiat. Biol. 2022, 99, 807–822. [Google Scholar] [CrossRef] [PubMed]
- Missiaggia, M.; Cordoni, F.G.; Scifoni, E.; La Tessa, C. Cell Survival Computation via the Generalized Stochastic Microdosimetric Model (GSM2)-Part II: Numerical results. Radiat. Res. 2022; submitted. [Google Scholar]
- Cordoni, F.G. A spatial measure-valued model for radiation-induced DNA damage kinetics and repair under protracted irradiation condition. arXiv 2023, arXiv:2303.14784. [Google Scholar]
- Van Kampen, N.G. Stochastic Processes in Physics and Chemistry; Elsevier: Amsterdam, The Netherlands, 1992; Volume 1. [Google Scholar]
- Gardiner, C.W. Handbook of Stochastic Methods; Springer: Berlin/Heidelberg, Germany, 1985; Volume 3. [Google Scholar]
- Zhao, L.; Mi, D.; Sun, Y. A novel multitarget model of radiation-induced cell killing based on the Gaussian distribution. J. Theor. Biol. 2017, 420, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Rossi, H.H.; Zaider, M. Saturation in dual radiation action. In Quantitative Mathematical Models in Radiation Biology; Springer: Berlin/Heidelberg, Germany, 1988; pp. 111–118. [Google Scholar]
- Rossi, H.H.; Zaider, M. Microdosimetry and Its Applications; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Vassiliev, O.N. Formulation of the multi-hit model with a non-Poisson distribution of hits. Int. J. Radiat. Oncol. Biol. Phys. 2012, 83, 1311–1316. [Google Scholar] [CrossRef]
- Manganaro, L.; Russo, G.; Cirio, R.; Dalmasso, F.; Giordanengo, S.; Monaco, V.; Muraro, S.; Sacchi, R.; Vignati, A.; Attili, A. A Monte Carlo approach to the microdosimetric kinetic model to account for dose rate time structure effects in ion beam therapy with application in treatment planning simulations. Med. Phys. 2017, 44, 1577–1589. [Google Scholar] [CrossRef] [PubMed]
- Karatzas, I.; Shreve, S.E. Brownian motion. In Brownian Motion and Stochastic Calculus; Springer: Berlin/Heidelberg, Germany, 1998; pp. 47–127. [Google Scholar]
- Gillespie, D.T. The chemical Langevin equation. J. Chem. Phys. 2000, 113, 297–306. [Google Scholar] [CrossRef]
- Albeverio, S.; Cordoni, F.; Di Persio, L.; Pellegrini, G. Asymptotic expansion for some local volatility models arising in finance. Decis. Econ. Financ. 2019, 42, 527–573. [Google Scholar] [CrossRef]
- Cordoni, F.; Di Persio, L. Small noise expansion for the Lévy perturbed Vasicek model. Int. J. Pure Appl. Math. 2015, 98, 291–301. [Google Scholar] [CrossRef]
- Weinan, E.; Li, T.; Vanden-Eijnden, E. Applied Stochastic Analysis; American Mathematical Society: Providence, RI, USA, 2019; Volume 199. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordoni, F.G. On the Emergence of the Deviation from a Poisson Law in Stochastic Mathematical Models for Radiation-Induced DNA Damage: A System Size Expansion. Entropy 2023, 25, 1322. https://doi.org/10.3390/e25091322
Cordoni FG. On the Emergence of the Deviation from a Poisson Law in Stochastic Mathematical Models for Radiation-Induced DNA Damage: A System Size Expansion. Entropy. 2023; 25(9):1322. https://doi.org/10.3390/e25091322
Chicago/Turabian StyleCordoni, Francesco Giuseppe. 2023. "On the Emergence of the Deviation from a Poisson Law in Stochastic Mathematical Models for Radiation-Induced DNA Damage: A System Size Expansion" Entropy 25, no. 9: 1322. https://doi.org/10.3390/e25091322