
Mechanistic Modelling of Biomass Growth, Glucose Consumption and Ethanol Production by Kluyveromyces marxianus in Batch Fermentation
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Data: Culture Medium and Analytical Techniques
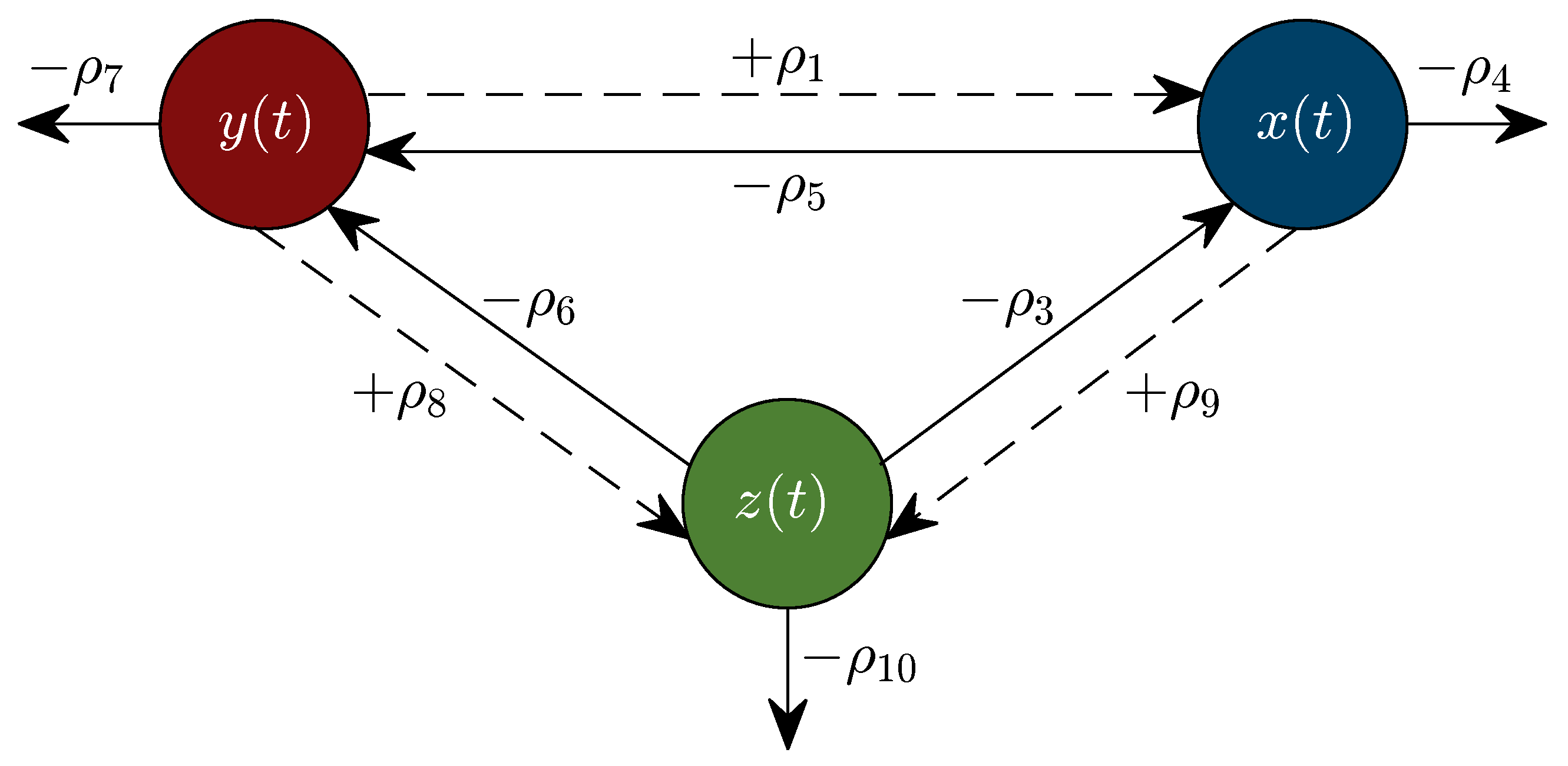
2.2. The KM Mechanistic Model
2.3. Parameter Value Estimation
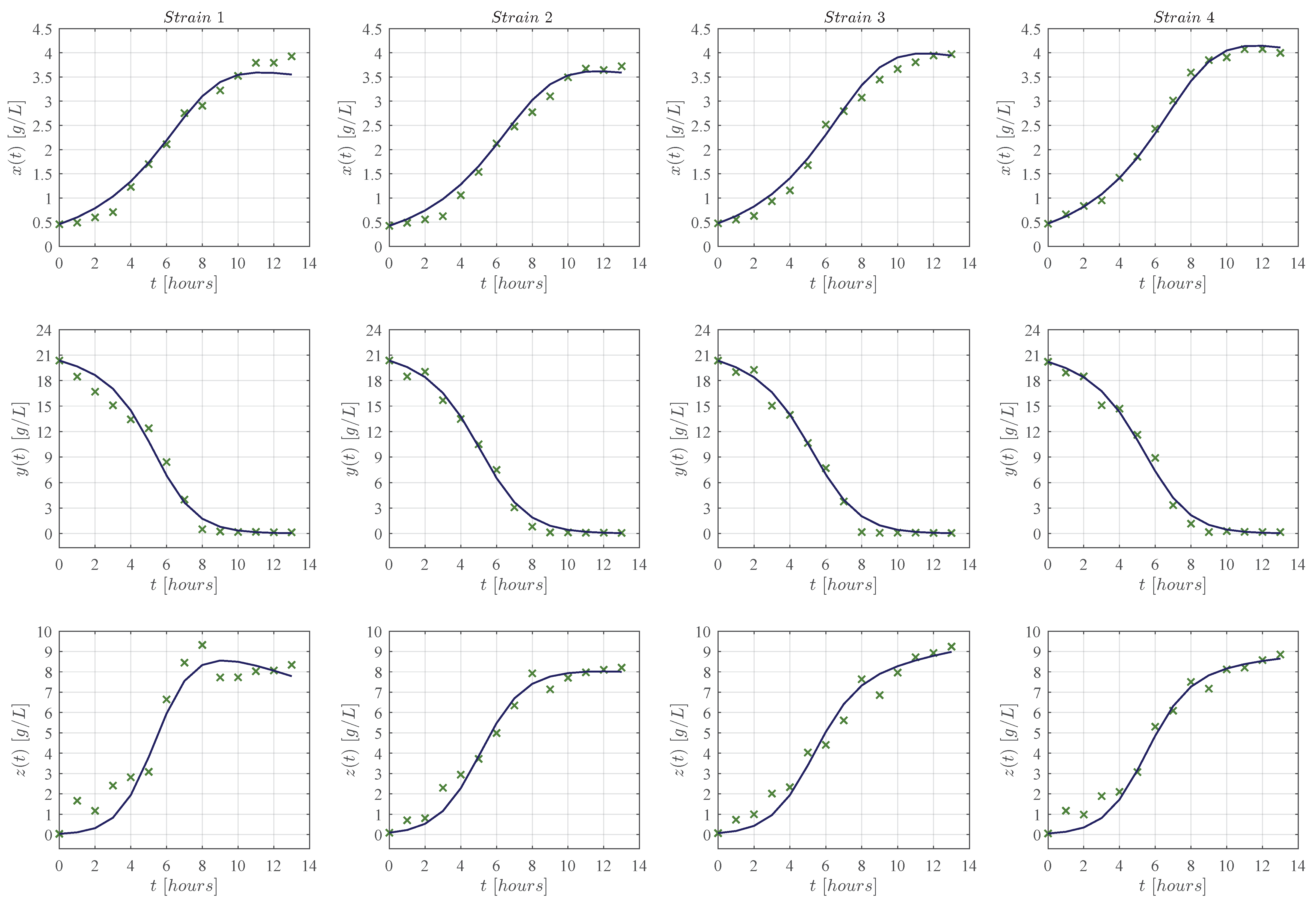
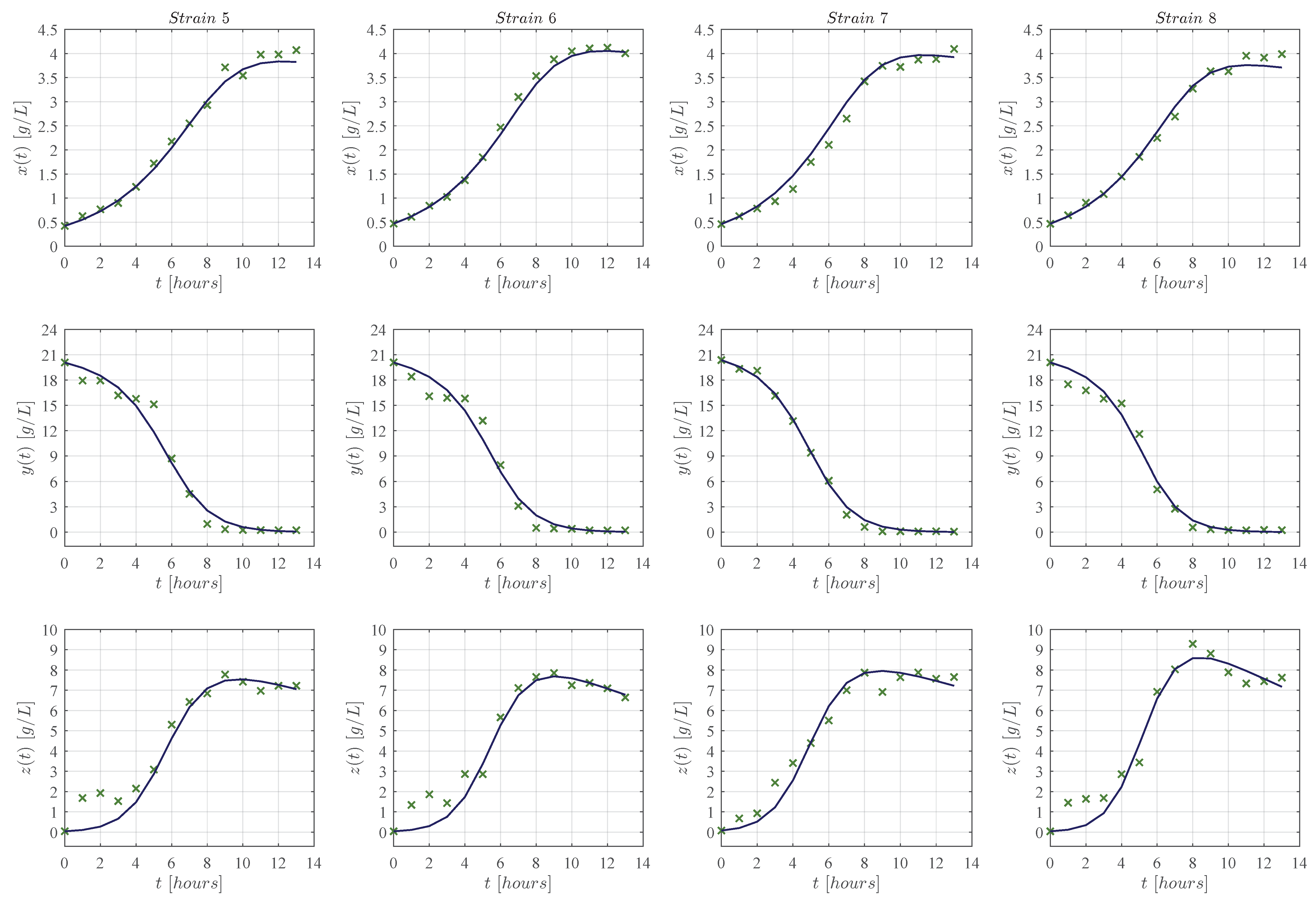
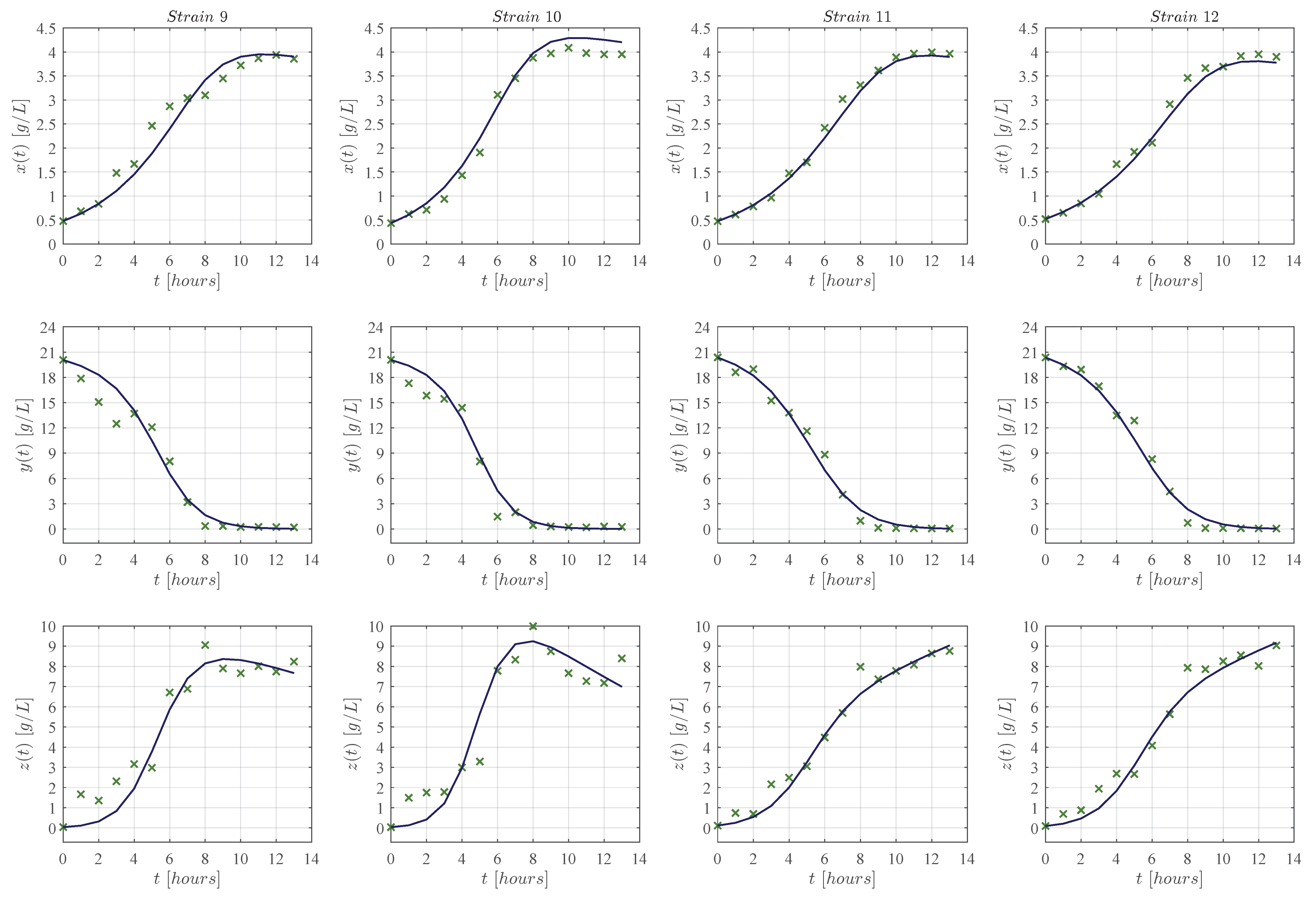
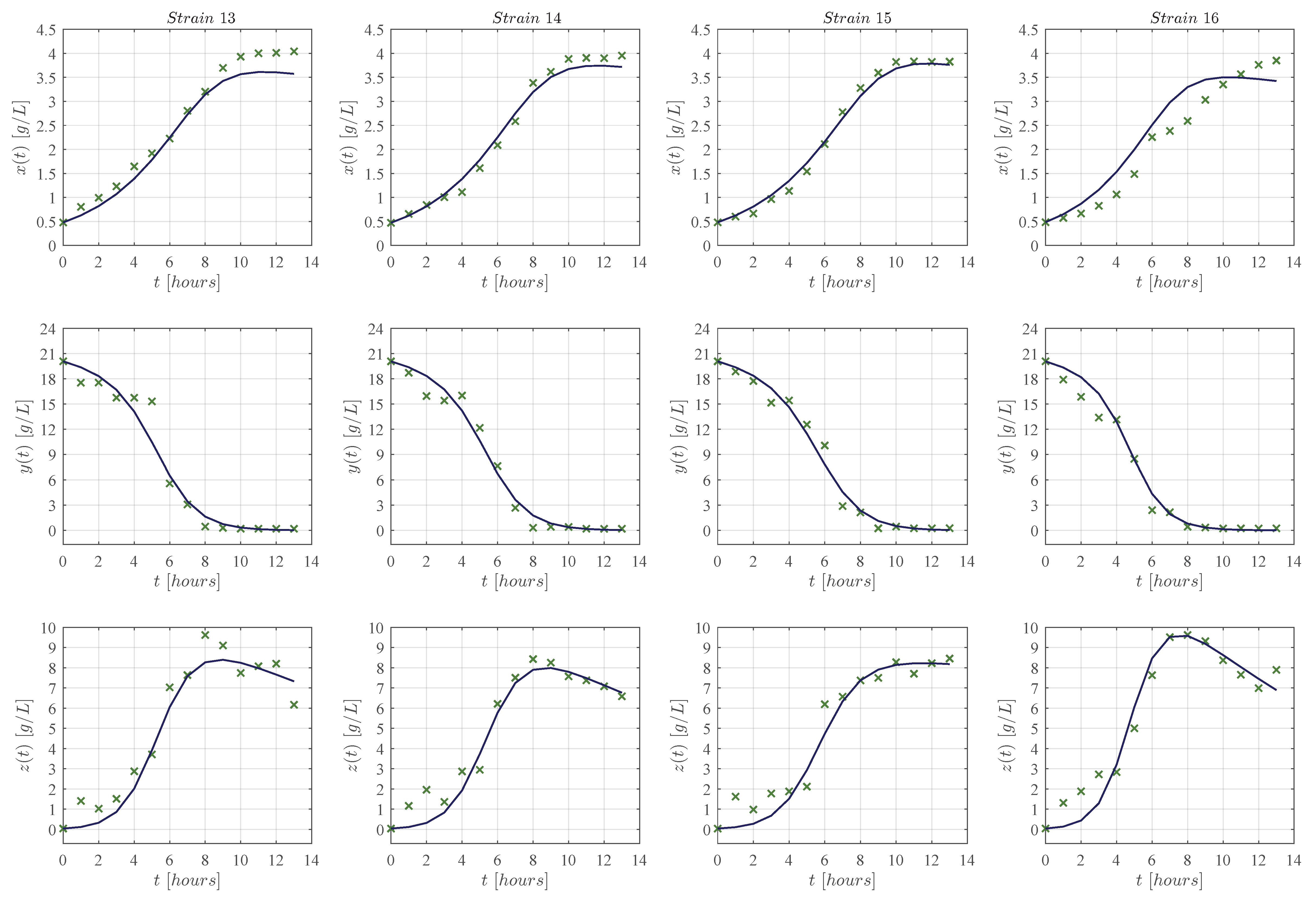
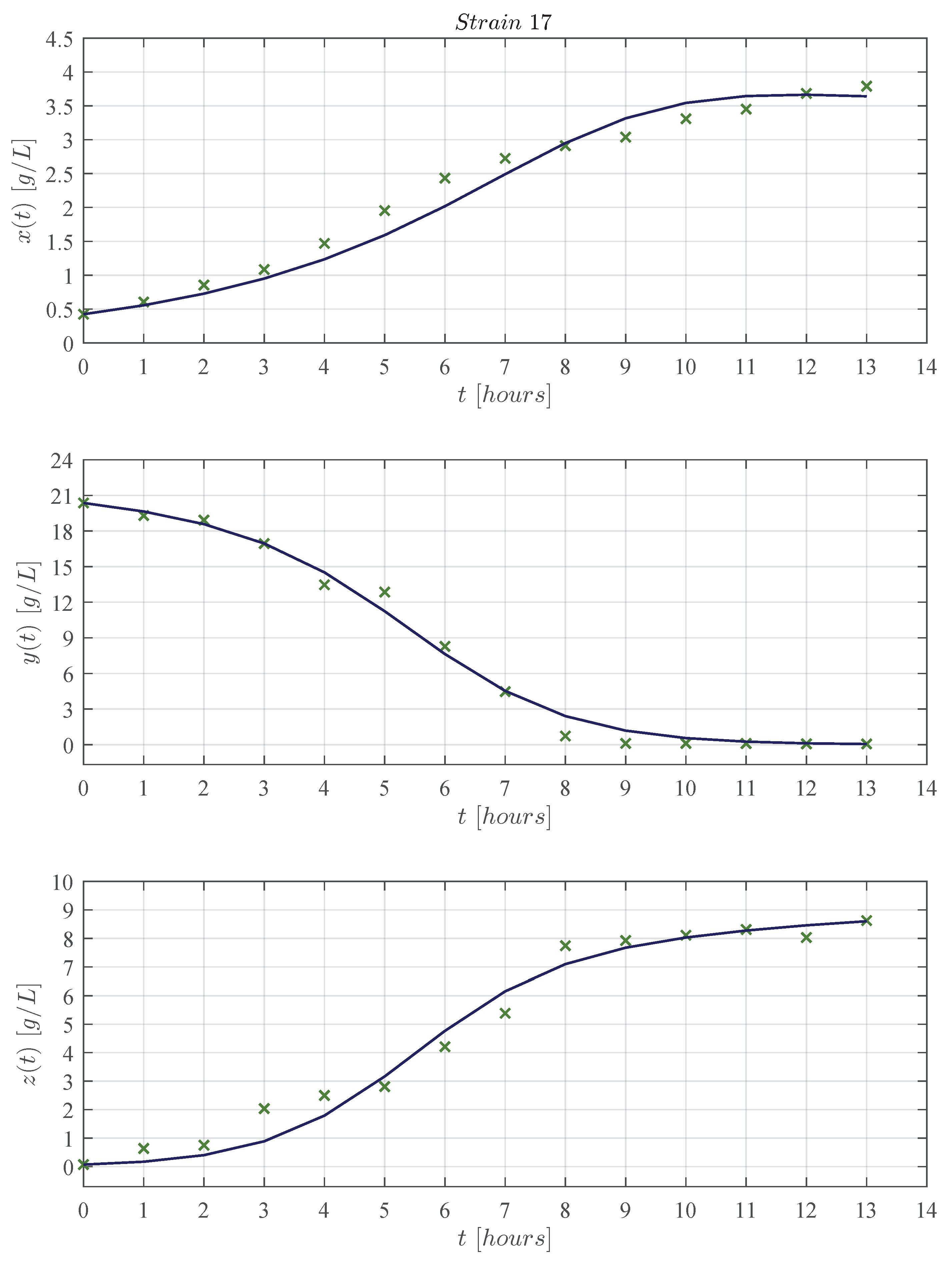
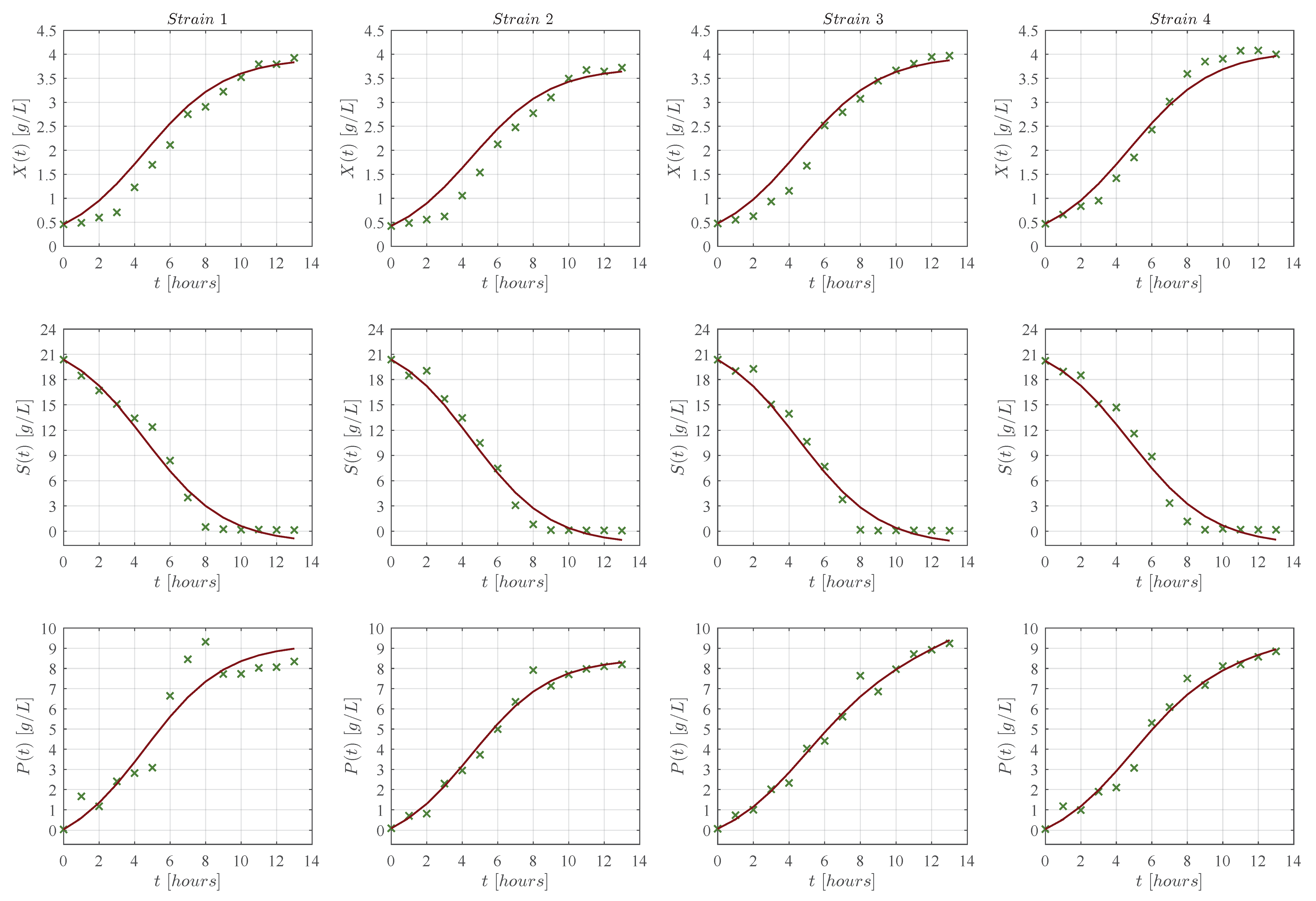
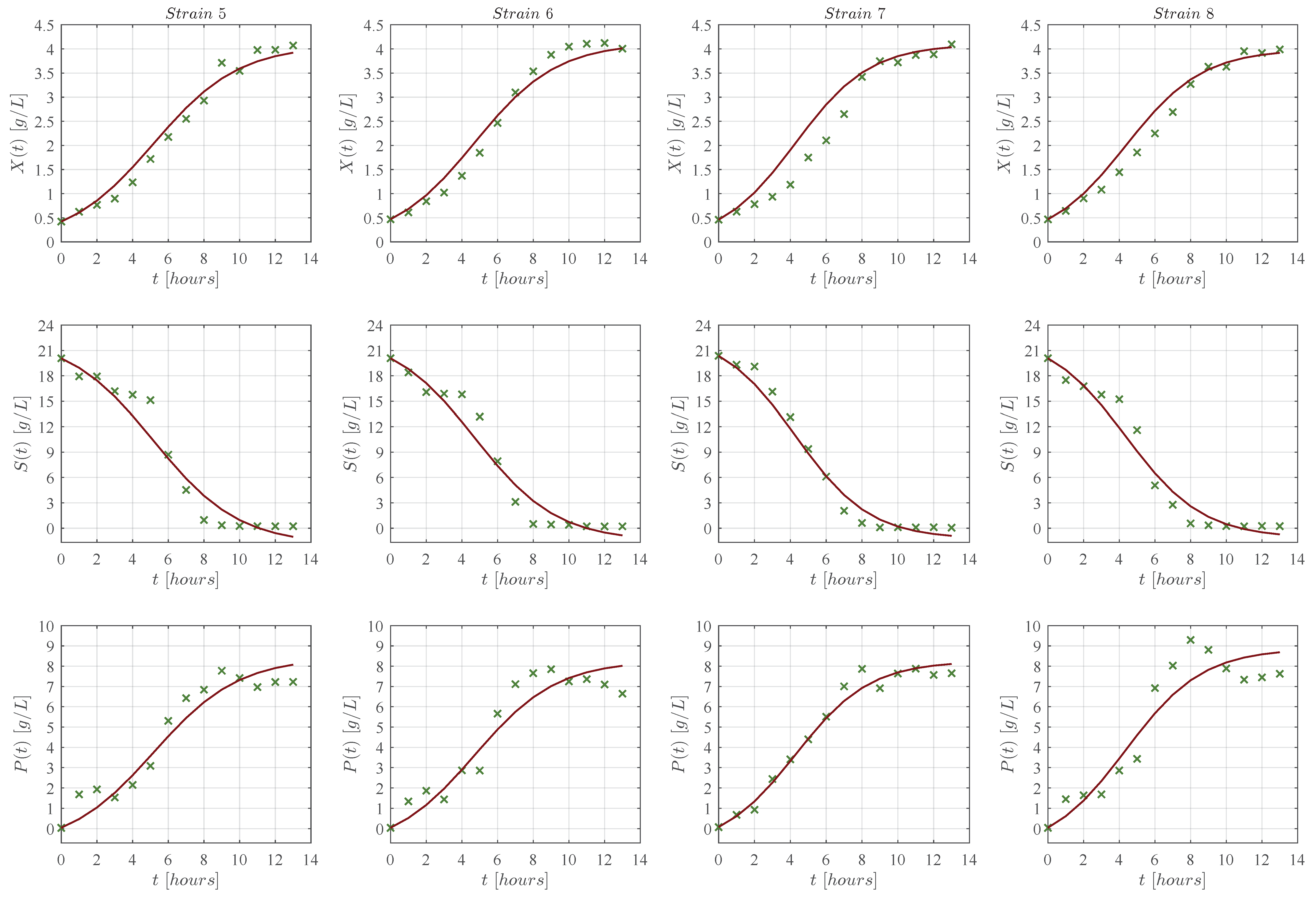
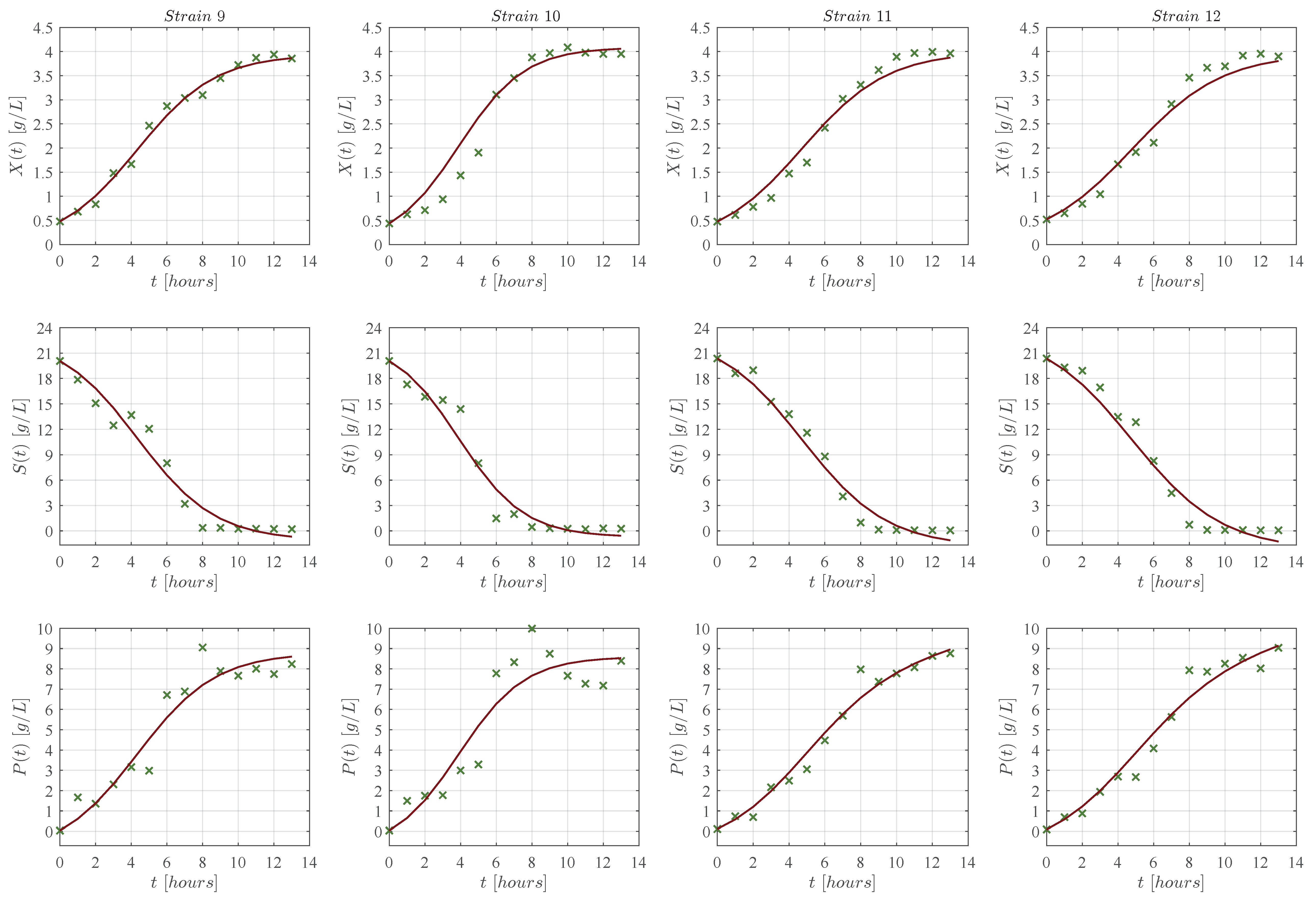
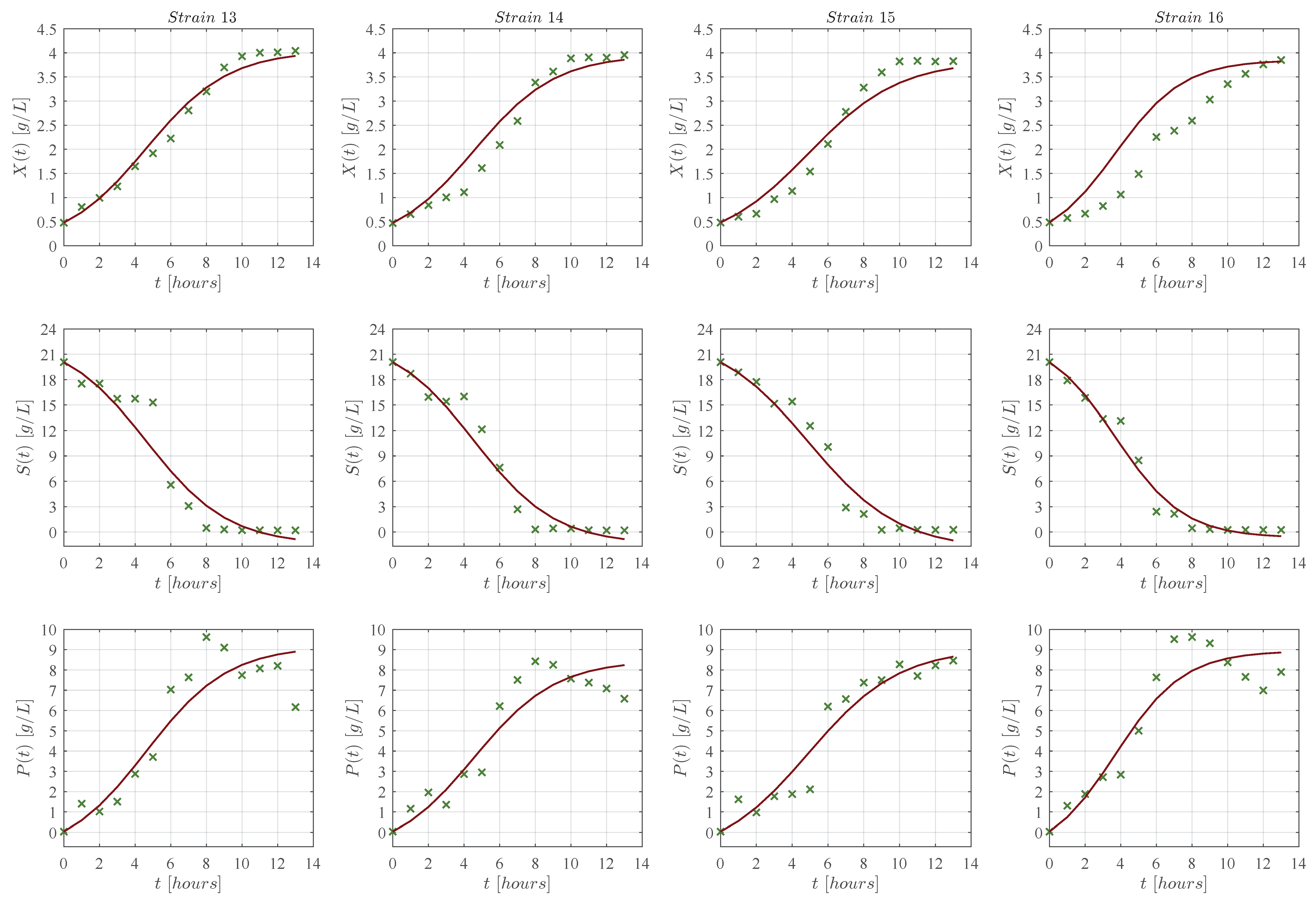
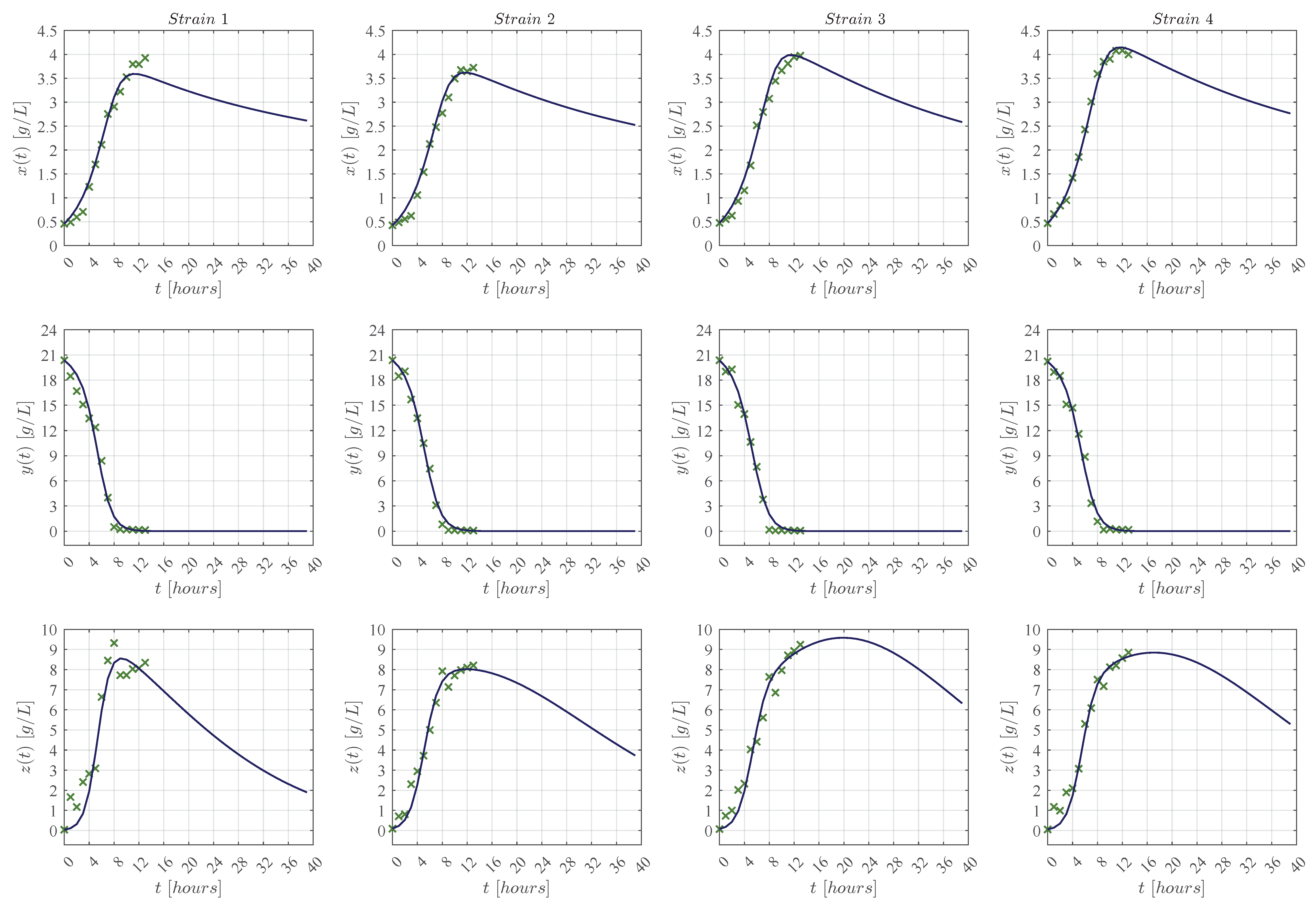
3. Results
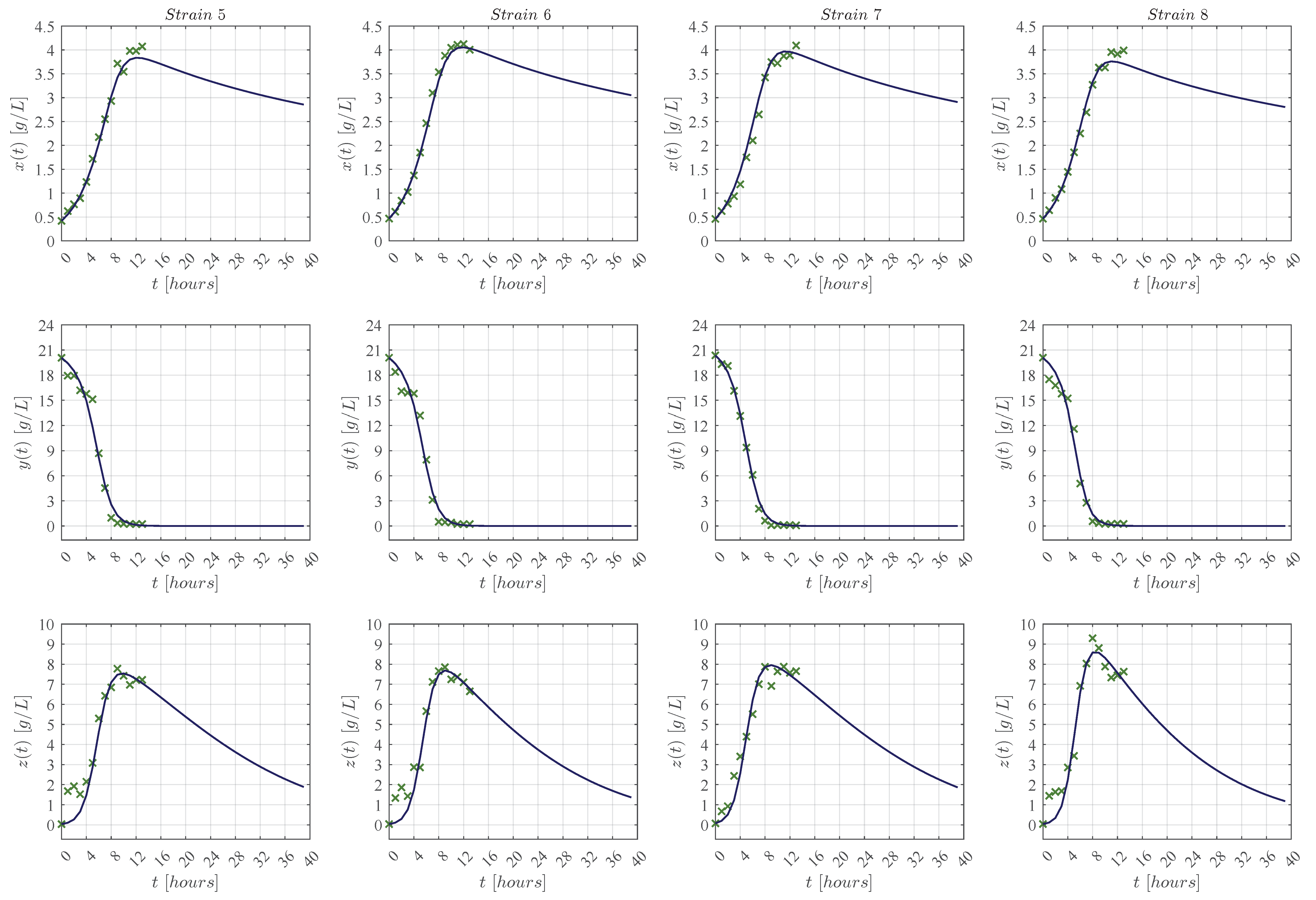
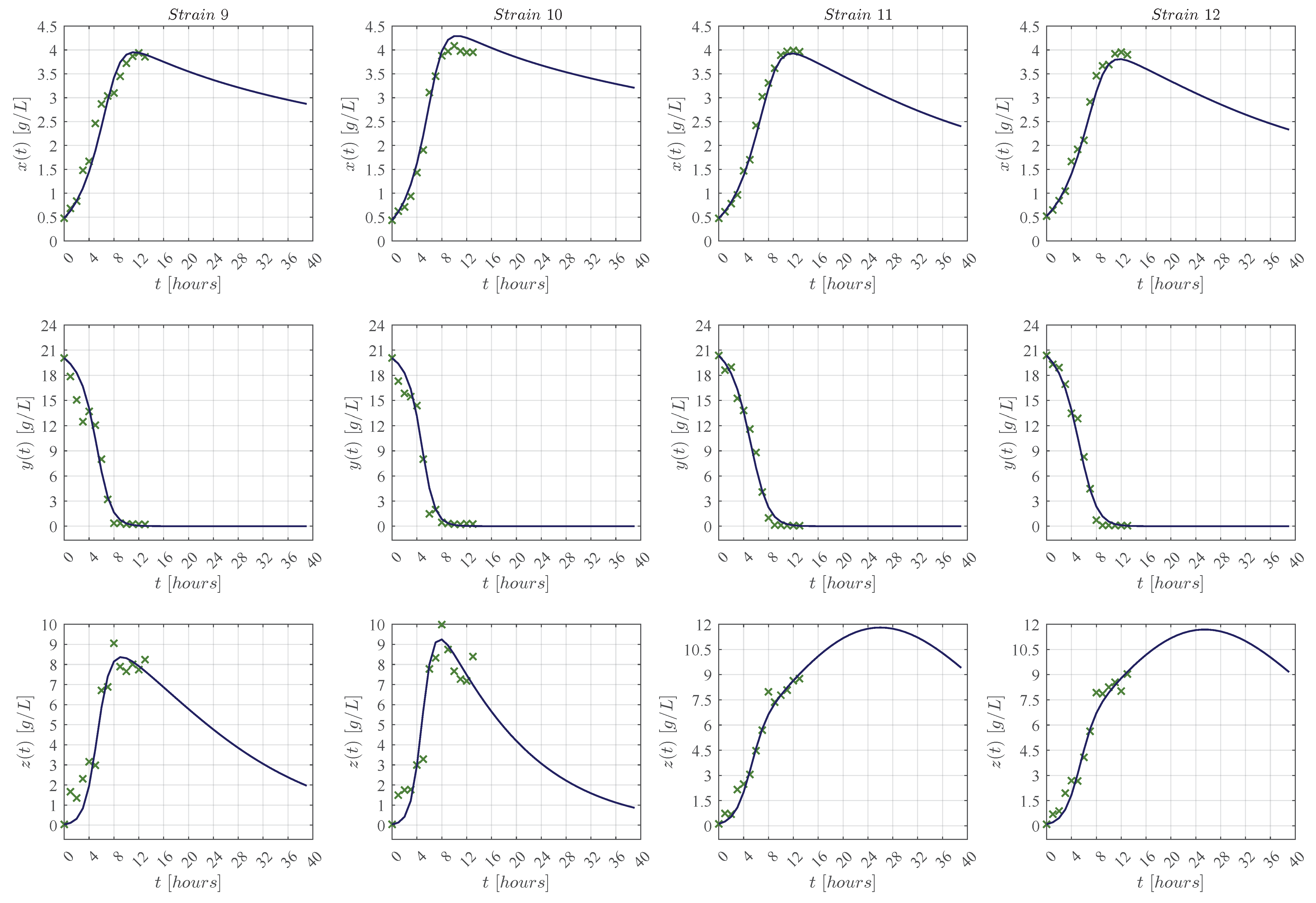
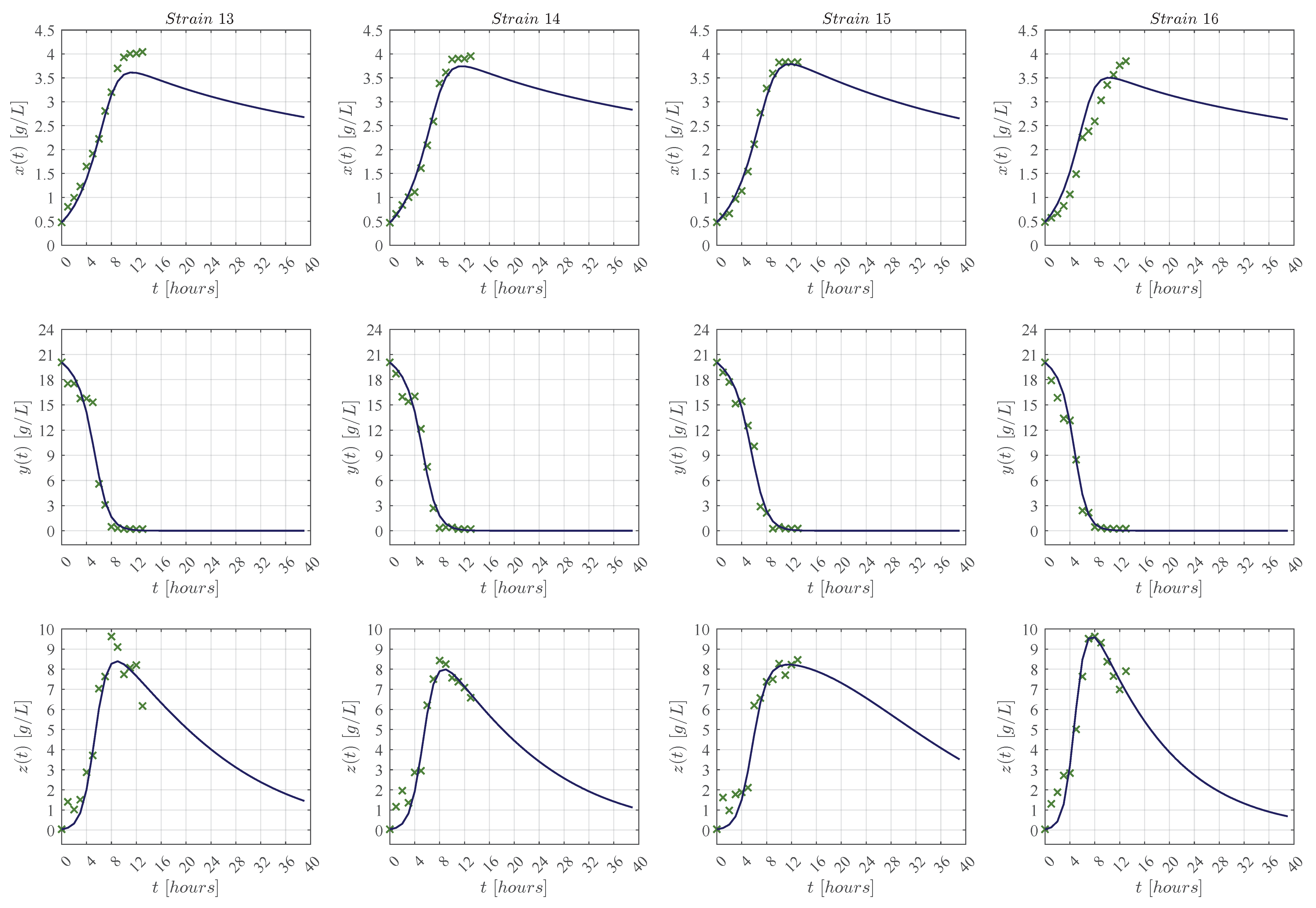
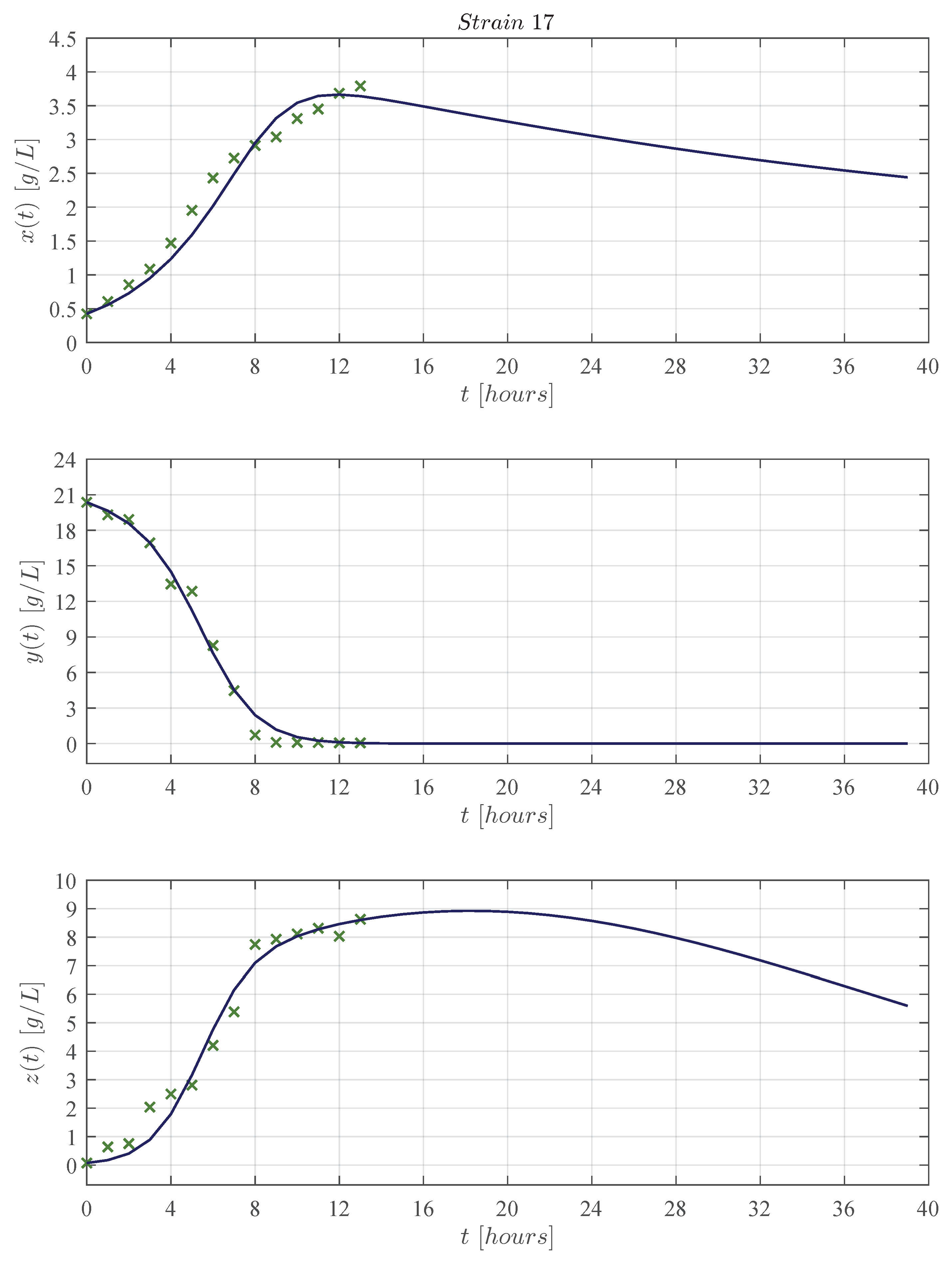
3.1. In Silico Experimentation and Goodness of Fit
3.2. Nonlinear Analysis: Localizing Domain, Asymptotic Stability, Existence and Uniqueness
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Akaike Infoirmation Criterion | |
| CI | Confidence Interval |
| K. marxianus | Kluyveromyces marxianus |
| LCIS | Localization of Compact Invariant Sets |
| Residual Sum of Squares | |
| Coefficient of Determination | |
| Standard Error |
Appendix A. Equilibria
Appendix B. Logistic, Pirt, and Luedeking–Piret Equations





Appendix C. Predictive Ability of the KM Mechanistic Model





References
- Zamora, F. Biochemistry of alcoholic fermentation. In Wine Chemistry and Biochemistry; Springer: Berlin/Heidelberg, Germany, 2009; pp. 3–26. [Google Scholar] [CrossRef]
- Dashko, S.; Zhou, N.; Compagno, C.; Piškur, J. Why, when, and how did yeast evolve alcoholic fermentation? FEMS Yeast Res. 2014, 14, 826–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourkoutas, Y.; Dimitropoulou, S.; Kanellaki, M.; Marchant, R.; Nigam, P.; Banat, I.; Koutinas, A. High-temperature alcoholic fermentation of whey using Kluyveromyces marxianus IMB3 yeast immobilized on delignified cellulosic material. Bioresour. Technol. 2002, 82, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, G.G.; Heinzle, E.; Wittmann, C.; Gombert, A.K. The yeast Kluyveromyces marxianus and its biotechnological potential. Appl. Microbiol. Biotechnol. 2008, 79, 339–354. [Google Scholar] [CrossRef]
- Morrissey, J.P.; Etschmann, M.M.; Schrader, J.; de Billerbeck, G.M. Cell factory applications of the yeast Kluyveromyces marxianus for the biotechnological production of natural flavour and fragrance molecules. Yeast 2015, 32, 3–16. [Google Scholar] [CrossRef]
- Wittmann, C.; Hans, M.; Bluemke, W. Metabolic physiology of aroma-producing Kluyveromyces marxianus. Yeast 2002, 19, 1351–1363. [Google Scholar] [CrossRef]
- Zentou, H.; Zainal Abidin, Z.; Yunus, R.; Awang Biak, D.R.; Abdullah Issa, M.; Yahaya Pudza, M. A new model of alcoholic fermentation under a byproduct inhibitory effect. ACS Omega 2021, 6, 4137–4146. [Google Scholar] [CrossRef]
- Madigan, M.; Bender, K.; Buckley, D.; Sattley, W.; Stahl, D. Brock Biology of Microorganisms; Pearson: London, UK, 2018. [Google Scholar]
- Van Maris, A.J.; Abbott, D.A.; Bellissimi, E.; van den Brink, J.; Kuyper, M.; Luttik, M.A.; Wisselink, H.W.; Scheffers, W.A.; van Dijken, J.P.; Pronk, J.T. Alcoholic fermentation of carbon sources in biomass hydrolysates by Saccharomyces cerevisiae: Current status. Antonie Van Leeuwenhoek 2006, 90, 391–418. [Google Scholar] [CrossRef] [PubMed]
- Valdramidis, V. 1-Predictive Microbiology. In Modeling in Food Microbiology; Elsevier: Oxford, UK, 2016; pp. 1–15. [Google Scholar] [CrossRef]
- Perez-Rodriguez, F.; Valero, A. Predictive microbiology in foods. In Predictive Microbiology in Foods; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–10. [Google Scholar] [CrossRef]
- Ross, T.; McMeekin, T.; Baranyi, J. Predictive Microbiology and Food Safety. In Encyclopedia of Food Microbiology, 2nd ed.; Academic Press: Oxford, UK, 2014; pp. 59–68. [Google Scholar] [CrossRef]
- Zhi, N.N.; Zong, K.; Thakur, K.; Qu, J.; Shi, J.J.; Yang, J.L.; Yao, J.; Wei, Z.J. Development of a dynamic prediction model for shelf-life evaluation of yogurt by using physicochemical, microbiological and sensory parameters. CyTA-J. Food 2018, 16, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Ontiveros, M.F.A. Modelizado de la dinámica de la vida de anaquel de microorganismos en leche fermentada. Rev. Aristas 2022, 9, 219–225. [Google Scholar]
- Buchanan, R.L. Predictive food microbiology. Trends Food Sci. Technol. 1993, 4, 6–11. [Google Scholar] [CrossRef]
- Sharma, V.; Mishra, H.N. Unstructured kinetic modeling of growth and lactic acid production by Lactobacillus plantarum NCDC 414 during fermentation of vegetable juices. LWT-Food Sci. Technol. 2014, 59, 1123–1128. [Google Scholar] [CrossRef]
- Gompertz, B., XXIV. On the nature of the function expressive of the law of human mortality, and on a new mode of determining the value of life contingencies. In a letter to Francis Baily, Esq. FRS &c. Philos. Trans. R. Soc. Lond. 1825, 513–583. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Murado, M.A. Unstructured mathematical model for biomass, lactic acid and bacteriocin production by lactic acid bacteria in batch fermentation. J. Chem. Technol. Biotechnol. Int. Res. Process. Environ. Clean Technol. 2008, 83, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Baranyi, J.; Roberts, T.; McClure, P. A non-autonomous differential equation to model bacterial growth. Food Microbiol. 1993, 10, 43–59. [Google Scholar] [CrossRef]
- Garcia, B.E.; Rodriguez, E.; Salazar, Y.; Valle, P.A.; Flores-Gallegos, A.C.; Rutiaga-Quiñones, O.M.; Rodriguez-Herrera, R. Primary Model for Biomass Growth Prediction in Batch Fermentation. Symmetry 2021, 13, 1468. [Google Scholar] [CrossRef]
- Monod, J. The growth of bacterial cultures. Annu. Rev. Microbiol. 1949, 3, 371–394. [Google Scholar] [CrossRef] [Green Version]
- Teissier, G. Growth of bacterial populations and the available substrate concentration. Rev. Sci. Instrum. 1942, 3208, 209–214. [Google Scholar]
- Haldane, J. Enzymes Longmans; Green and Co.: Oxford, UK, 1930. [Google Scholar]
- Moser, H. The Dynamics of Bacterial Populations Maintained in the Chemostat; Carnegie Institution of Washington: Washington, DC, USA, 1958. [Google Scholar]
- Muloiwa, M.; Nyende-Byakika, S.; Dinka, M. Comparison of unstructured kinetic bacterial growth models. S. Afr. J. Chem. Eng. 2020, 33, 141–150. [Google Scholar] [CrossRef]
- Li, H.; Xie, G.; Edmondson, A.S. Review of secondary mathematical models of predictive microbiology. J. Food Prod. Mark. 2008, 14, 57–74. [Google Scholar] [CrossRef]
- Fakruddin, M.; Mazumder, R.M.; Mannan, K.S.B. Predictive microbiology: Modeling microbial responses in food. Ceylon J. Sci. 2011, 40, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Zentou, H.; Zainal Abidin, Z.; Yunus, R.; Awang Biak, D.R.; Zouanti, M.; Hassani, A. Modelling of molasses fermentation for bioethanol production: A comparative investigation of Monod and Andrews models accuracy assessment. Biomolecules 2019, 9, 308. [Google Scholar] [CrossRef] [Green Version]
- Sansonetti, S.; Hobley, T.J.; Calabrò, V.; Villadsen, J.; Sin, G. A biochemically structured model for ethanol fermentation by Kluyveromyces marxianus: A batch fermentation and kinetic study. Bioresour. Technol. 2011, 102, 7513–7520. [Google Scholar] [CrossRef]
- Lei, F.; Rotbøll, M.; Jørgensen, S.B. A biochemically structured model for Saccharomyces cerevisiae. J. Biotechnol. 2001, 88, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Steinmeyer, D.; Shuler, M. Structured model for Saccharomyces cerevisiae. Chem. Eng. Sci. 1989, 44, 2017–2030. [Google Scholar] [CrossRef]
- Reyes-Sánchez, F.J.; Páez-Lerma, J.B.; Rojas-Contreras, J.A.; López-Miranda, J.; Soto-Cruz, N.Ó.; Reinhart-Kirchmayr, M. Study of the Enzymatic Capacity of Kluyveromyces marxianus for the Synthesis of Esters. Microb. Physiol. 2019, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Valle, P.A.; Coria, L.N.; Plata, C.; Salazar, Y. CAR-T Cell Therapy for the Treatment of ALL: Eradication Conditions and In Silico Experimentation. Hemato 2021, 2, 28. [Google Scholar] [CrossRef]
- Páez-Lerma, J.B.; Arias-García, A.; Rutiaga-Quiñones, O.M.; Barrio, E.; Soto-Cruz, N.O. Yeasts isolated from the alcoholic fermentation of Agave duranguensis during mezcal production. Food Biotechnol. 2013, 27, 342–356. [Google Scholar] [CrossRef]
- Paredes-Ortíz, A.; Olvera-Martínez, T.; Páez-Lerma, J.; Rojas-Contreras, J.; Moreno-Jiménez, M.; Aguilar, C.; Soto-Cruz, N. Isoamyl acetate production during continuous culture of Pichia fermentans. Rev. Mex. Ing. Química 2022, 21, Bio2654. [Google Scholar] [CrossRef]
- da Silva, B.; Gonzaga, L.V.; Fett, R.; Costa, A.C.O. Simplex-centroid design and Derringer’s desirability function approach for simultaneous separation of phenolic compounds from Mimosa scabrella Bentham honeydew honeys by HPLC/DAD. J. Chromatogr. A 2019, 1585, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Almeida, I.C.; Pacheco, T.F.; Machado, F.; Gonçalves, S.B. Evaluation of different strains of Saccharomyces cerevisiae for ethanol production from high-amylopectin BRS AG rice (Oryza sativa L.). Sci. Rep. 2022, 12, 1–15. [Google Scholar] [CrossRef]
- De Leenheer, P.; Aeyels, D. Stability properties of equilibria of classes of cooperative systems. IEEE Trans. Autom. Control. 2001, 46, 1996–2001. [Google Scholar] [CrossRef] [Green Version]
- Britton, N.F.; Britton, N. Essential Mathematical Biology; Springer: Berlin/Heidelberg, Germany, 2003; Volume 453. [Google Scholar] [CrossRef]
- Ding, J.; Huang, X.; Zhang, L.; Zhao, N.; Yang, D.; Zhang, K. Tolerance and stress response to ethanol in the yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2009, 85, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Kubota, S.; Takeo, I.; Kume, K.; Kanai, M.; Shitamukai, A.; Mizunuma, M.; Miyakawa, T.; Shimoi, H.; Iefuji, H.; Hirata, D. Effect of ethanol on cell growth of budding yeast: Genes that are important for cell growth in the presence of ethanol. Biosci. Biotechnol. Biochem. 2004, 68, 968–972. [Google Scholar] [CrossRef]
- Fonseca, G.G.; Gombert, A.K.; Heinzle, E.; Wittmann, C. Physiology of the yeast Kluyveromyces marxianus during batch and chemostat cultures with glucose as the sole carbon source. FEMS Yeast Res. 2007, 7, 422–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfenden, R.; Yuan, Y. Rates of spontaneous cleavage of glucose, fructose, sucrose, and trehalose in water, and the catalytic proficiencies of invertase and trehalas. J. Am. Chem. Soc. 2008, 130, 7548–7549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrussamee, N.; Lertwattanasakul, N.; Hirata, K.; Limtong, S.; Kosaka, T.; Yamada, M. Growth and ethanol fermentation ability on hexose and pentose sugars and glucose effect under various conditions in thermotolerant yeast Kluyveromyces marxianus. Appl. Microbiol. Biotechnol. 2011, 90, 1573–1586. [Google Scholar] [CrossRef]
- Wu, Z.; Song, L.; Liu, S.Q.; Huang, D. Independent and additive effects of glutamic acid and methionine on yeast longevity. PLoS ONE 2013, 8, e79319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, G.; Standaert, P.; Verachtert, H. Effects of metabolic inhibitors on the alcoholic fermentation by several yeasts in batch or in immobilized cell systems. Appl. Microbiol. Biotechnol. 1984, 19, 91–99. [Google Scholar] [CrossRef]
- Willey, J. Prescott’s Microbiology; McGraw-Hill Education: New York, NY, USA, 2019. [Google Scholar]
- Byers, J.P.; Sarver, J.G. Pharmacokinetic modeling. In Pharmacology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 201–277. [Google Scholar] [CrossRef]
- Garfinkel, A.; Shevtsov, J.; Guo, Y. Modeling Life: The Mathematics of Biological Systems; Springer: Cham, Switzerland, 2017. [Google Scholar]
- MathWorks. lsqcurvefit. 2022. Available online: https://www.mathworks.com/help/optim/ug/lsqcurvefit.html (accessed on 7 December 2022).
- Koutsoyiannis, A. Theory of Econometrics. An Introductory Exposition of Econometric Methods; The Macmillan Press LTD: New York, NY, USA, 1977. [Google Scholar]
- Motulsky, H. Intuitive Biostatistics: A Nonmathematical Guide to Statistical Thinking; Oxford University Press: New York, NY, USA, 2018. [Google Scholar]
- Akaike, H. Canonical correlation analysis of time series and the use of an information criterion. Math. Sci. Eng. 1976, 126, 27–96. [Google Scholar] [CrossRef]
- Hu, S. Akaike information criterion. Cent. Res. Sci. Comput. 2007, 93. Available online: https://www.researchgate.net/publication/267201163 (accessed on 11 January 2023).
- Slavkova, K.P.; Patel, S.H.; Cacini, Z.; Kazerouni, A.S.; Gardner, A.; Yankeelov, T.E.; Hormuth, D.A. Mathematical modelling of the dynamics of image-informed tumor habitats in a murine model of glioma. Sci. Rep. 2023, 13, 2916. [Google Scholar] [CrossRef] [PubMed]
- Krishchenko, A.P. Localization of invariant compact sets of dynamical systems. Differ. Equations 2005, 41, 1669–1676. [Google Scholar] [CrossRef]
- Valle, P.A.; Coria, L.N.; Gamboa, D.; Plata, C. Bounding the Dynamics of a Chaotic-Cancer Mathematical Model. Math. Probl. Eng. 2018, 2018, 14. [Google Scholar] [CrossRef]
- Krishchenko, A.P.; Starkov, K.E. Localization of compact invariant sets of the Lorenz system. Phys. Lett. A 2006, 353, 383–388. [Google Scholar] [CrossRef]
- Khalil, H.K. Nonlinear Systems, 3rd ed.; Prentice-Hall: Hoboken, NJ, USA, 2002. [Google Scholar]
- Hahn, W.; Hosenthien, H.H.; Lehnigk, S.H. Theory and Application of Liapunov’s Direct Method; Dover Publications, Inc.: Mineola, NY, USA, 2019. [Google Scholar]
- Perko, L. Differential Equations and Dynamical Systems; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; Volume 7. [Google Scholar]
- Jiménez-Islas, D.; Páez-Lerma, J.; Soto-Cruz, N.O.; Gracida, J. Modelling of ethanol production from red beet juice by Saccharomyces cerevisiae under thermal and acid stress conditions. Food Technol. Biotechnol. 2014, 52, 93–100. Available online: https://hrcak.srce.hr/118561 (accessed on 11 March 2023).
- Pirt, S.J. Principles of Microbe and Cell Cultivation; Blackwell Scientific Publications: Hoboken, NJ, USA, 1975. [Google Scholar]
- Luedeking, R.; Piret, E.L. A kinetic study of the lactic acid fermentation. Batch process at controlled pH. J. Biochem. Microbiol. Technol. Eng. 1959, 1, 393–412. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
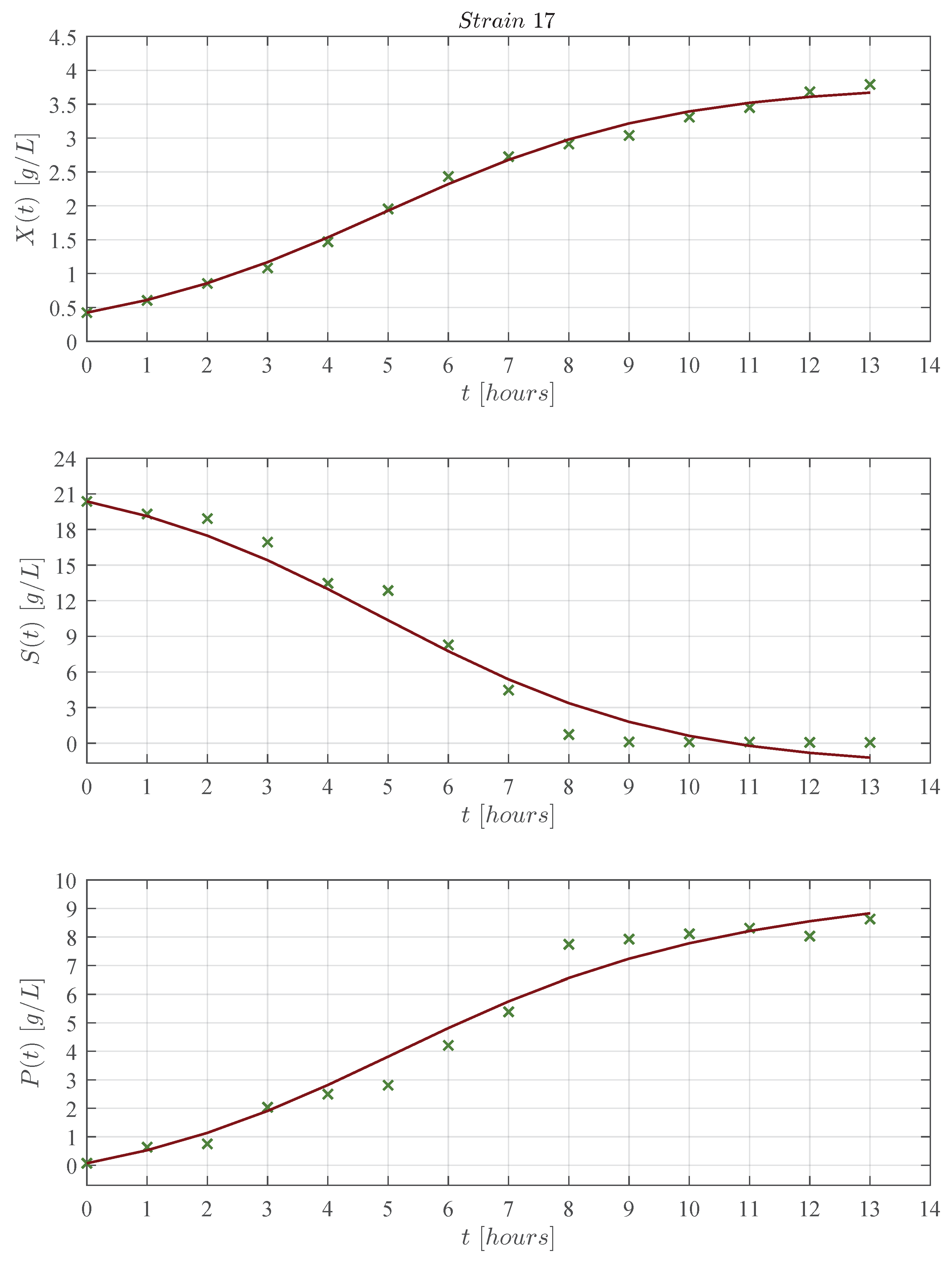{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables/ Parameters | Description | Values | Units |
|---|---|---|---|
| Biomass concentration | − | g/L | |
| Glucose concentration | − | g/L | |
| Ethanol concentration | − | g/L | |
| Biomass maximum growth rate | h | ||
| Affinity with substrate constant | g/L | ||
| Inhibition rate of biomass growth due to product accumulation | L/(g × h) | ||
| Biomass death rate | h | ||
| Consumption rate for biomass growth | L/(g × h) | ||
| Consumption rate for ethanol production | L/(g × h) | ||
| Glucose spontaneous decomposition rate | h | ||
| Ethanol production associated with the biomass growth rate | L/(g × h) | ||
| Glucose converted in ethanol | L/(g × h) | ||
| Ethanol degradation rate | h |
| Strain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | |||||||||
| 2 | |||||||||
| 3 | |||||||||
| 4 | |||||||||
| 5 | |||||||||
| 6 | |||||||||
| 7 | |||||||||
| 8 | |||||||||
| 9 | |||||||||
| 10 | |||||||||
| 11 | |||||||||
| 12 | |||||||||
| 13 | |||||||||
| 14 | |||||||||
| 15 | |||||||||
| 16 | |||||||||
| 17 |
| Strain | Biomass | Glucose | Ethanol |
|---|---|---|---|
| 1 | |||
| 2 | |||
| 3 | |||
| 4 | |||
| 5 | |||
| 6 | |||
| 7 | |||
| 8 | |||
| 9 | |||
| 10 | |||
| 11 | |||
| 12 | |||
| 13 | |||
| 14 | |||
| 15 | |||
| 16 | |||
| 17 |
| Strain | |||
|---|---|---|---|
| 1 | |||
| 2 | |||
| 3 | |||
| 4 | |||
| 5 | |||
| 6 | |||
| 7 | |||
| 8 | |||
| 9 | |||
| 10 | |||
| 11 | |||
| 12 | |||
| 13 | |||
| 14 | |||
| 15 | |||
| 16 | |||
| 17 |
| Strain | ||||||
|---|---|---|---|---|---|---|
| 1 | ||||||
| 2 | ||||||
| 3 | ||||||
| 4 | ||||||
| 5 | ||||||
| 6 | ||||||
| 7 | ||||||
| 8 | ||||||
| 9 | ||||||
| 10 | ||||||
| 11 | ||||||
| 12 | ||||||
| 13 | ||||||
| 14 | ||||||
| 15 | ||||||
| 16 | ||||||
| 17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar, Y.; Valle, P.A.; Rodríguez, E.; Soto-Cruz, N.O.; Páez-Lerma, J.B.; Reyes-Sánchez, F.J. Mechanistic Modelling of Biomass Growth, Glucose Consumption and Ethanol Production by Kluyveromyces marxianus in Batch Fermentation. Entropy 2023, 25, 497. https://doi.org/10.3390/e25030497
Salazar Y, Valle PA, Rodríguez E, Soto-Cruz NO, Páez-Lerma JB, Reyes-Sánchez FJ. Mechanistic Modelling of Biomass Growth, Glucose Consumption and Ethanol Production by Kluyveromyces marxianus in Batch Fermentation. Entropy. 2023; 25(3):497. https://doi.org/10.3390/e25030497
Chicago/Turabian StyleSalazar, Yolocuauhtli, Paul A. Valle, Emmanuel Rodríguez, Nicolás O. Soto-Cruz, Jesús B. Páez-Lerma, and Francisco J. Reyes-Sánchez. 2023. "Mechanistic Modelling of Biomass Growth, Glucose Consumption and Ethanol Production by Kluyveromyces marxianus in Batch Fermentation" Entropy 25, no. 3: 497. https://doi.org/10.3390/e25030497