

Biophotons: New Experimental Data and Analysis

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
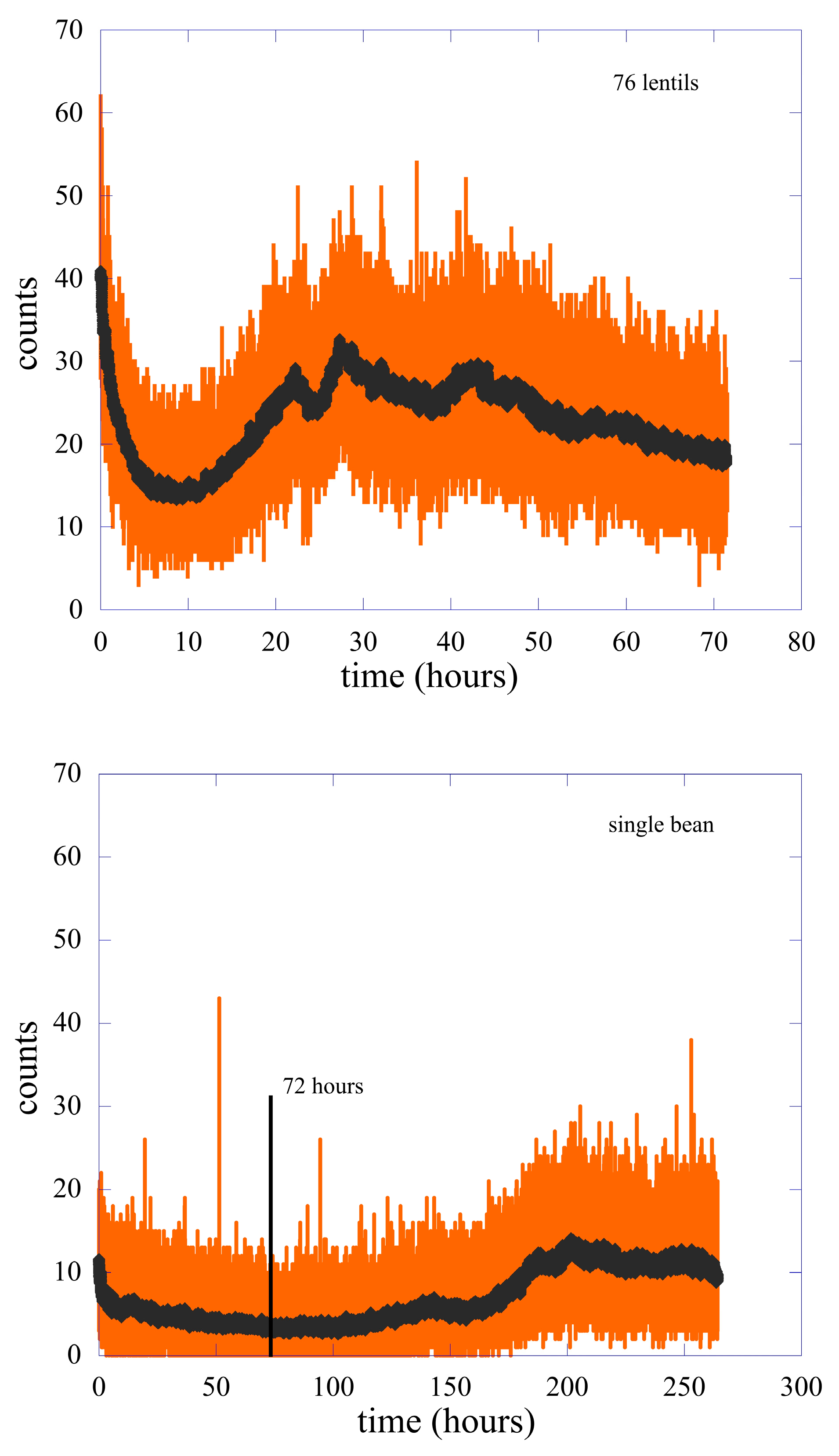
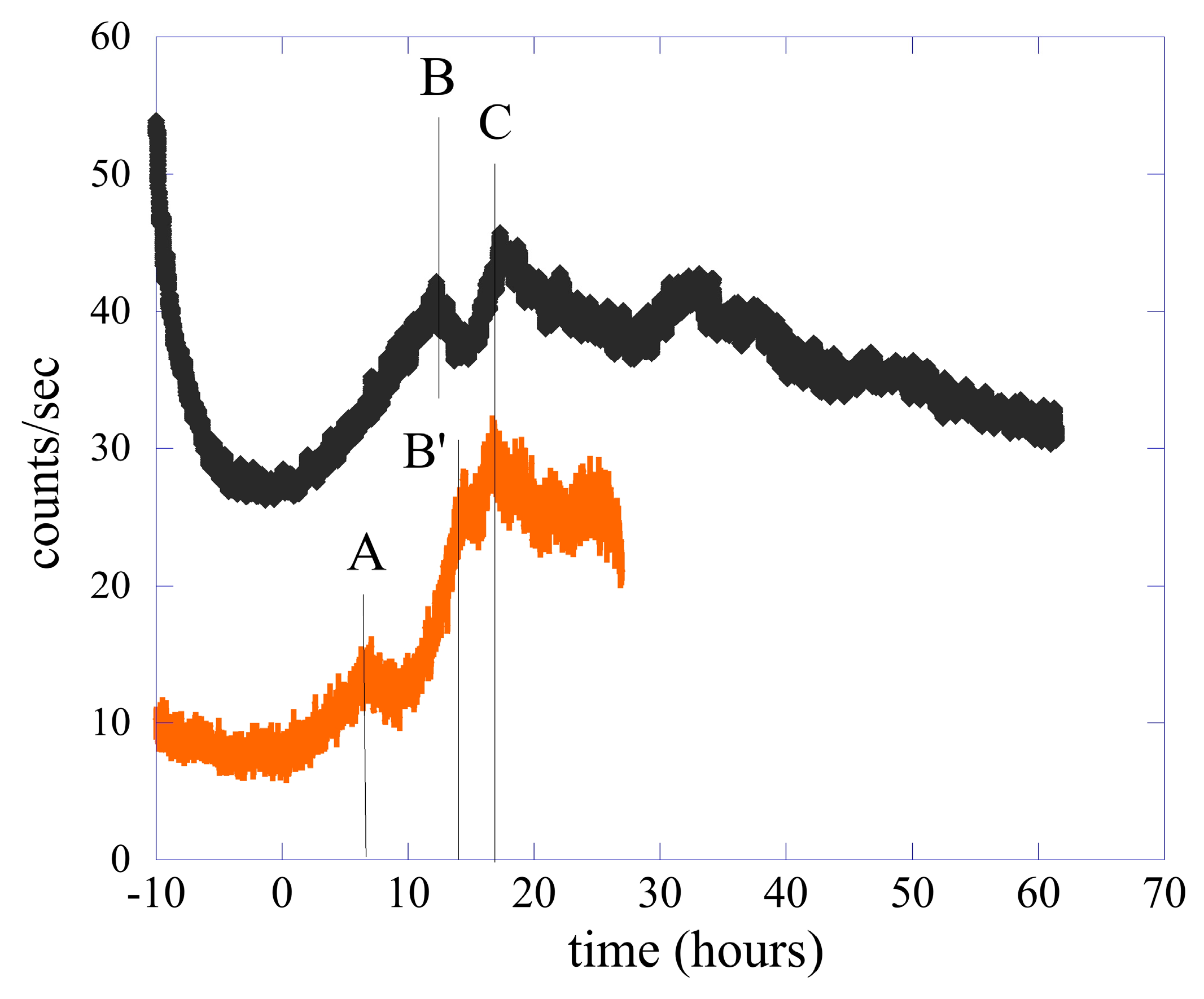
2. Methods and Experimental Data
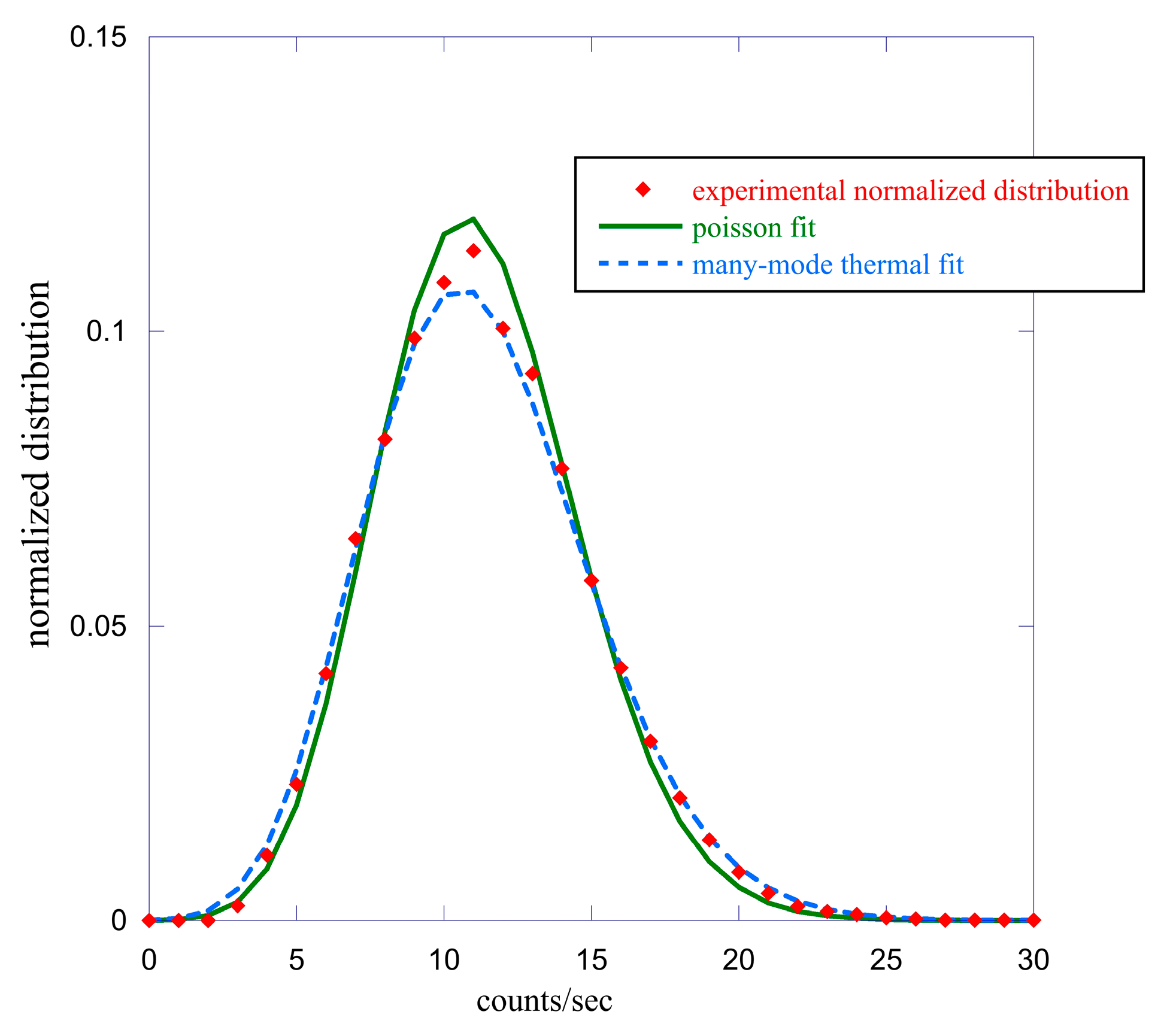
2.1. Data Analysis I—Probability Distribution Functions
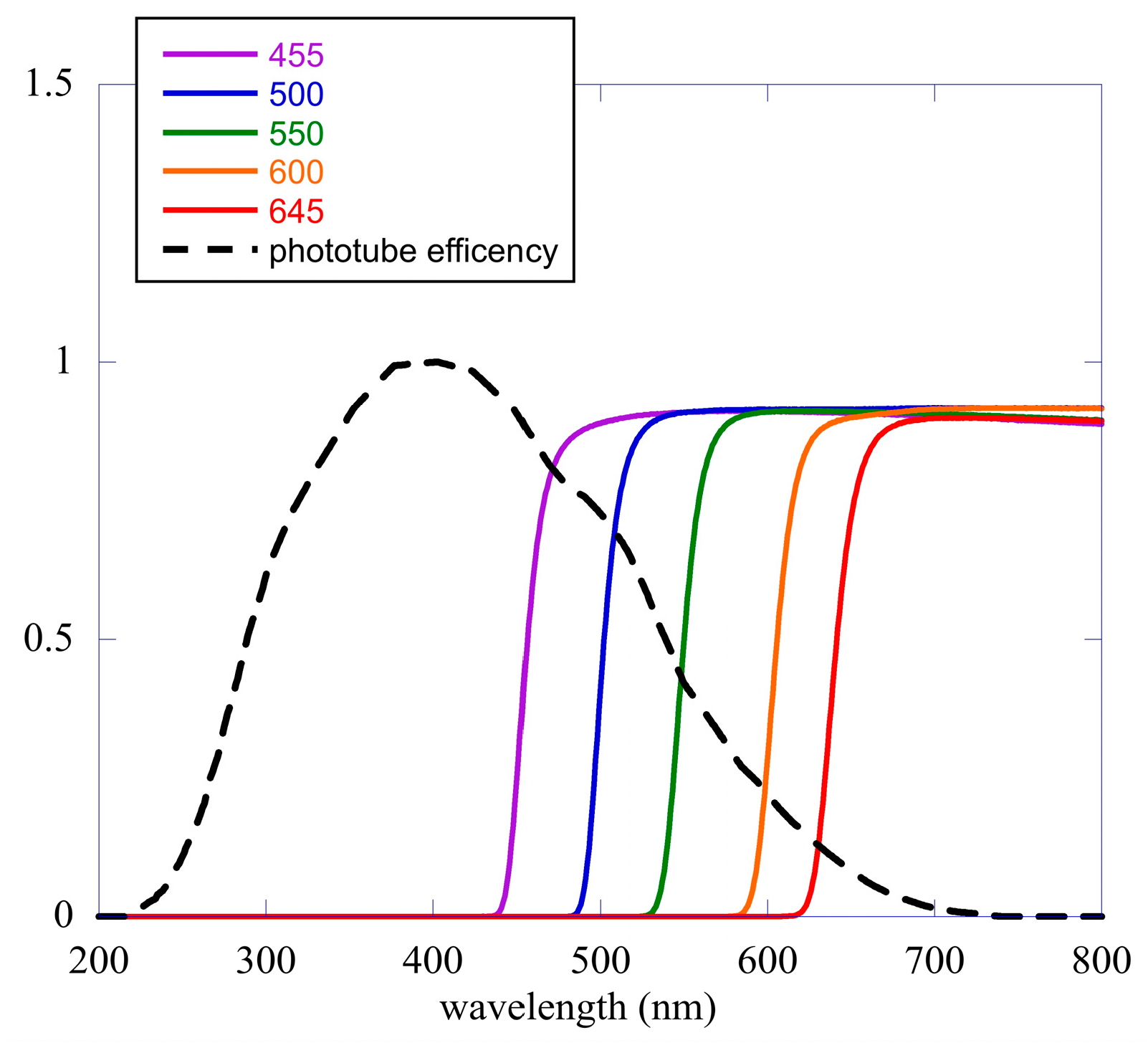
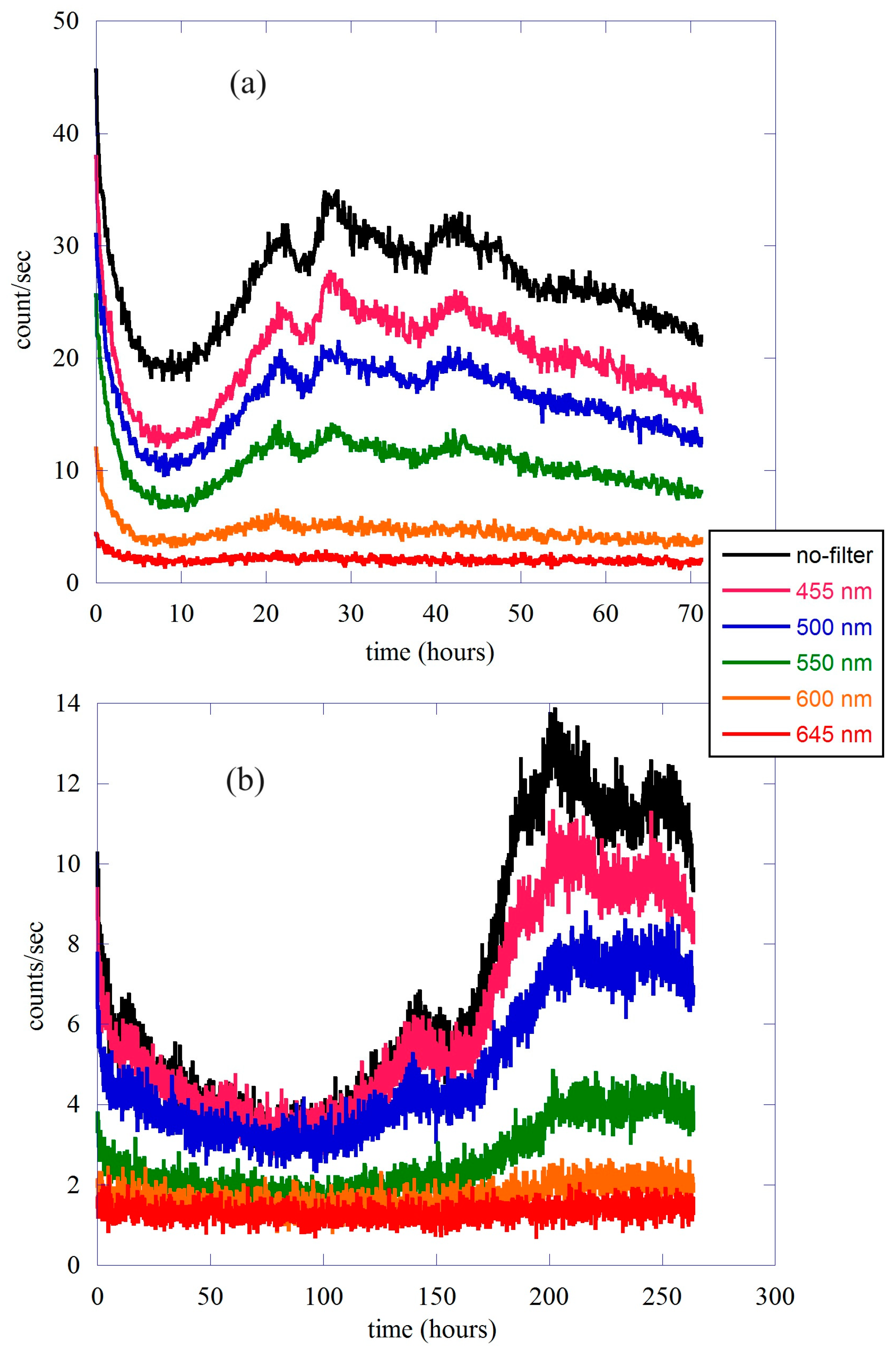
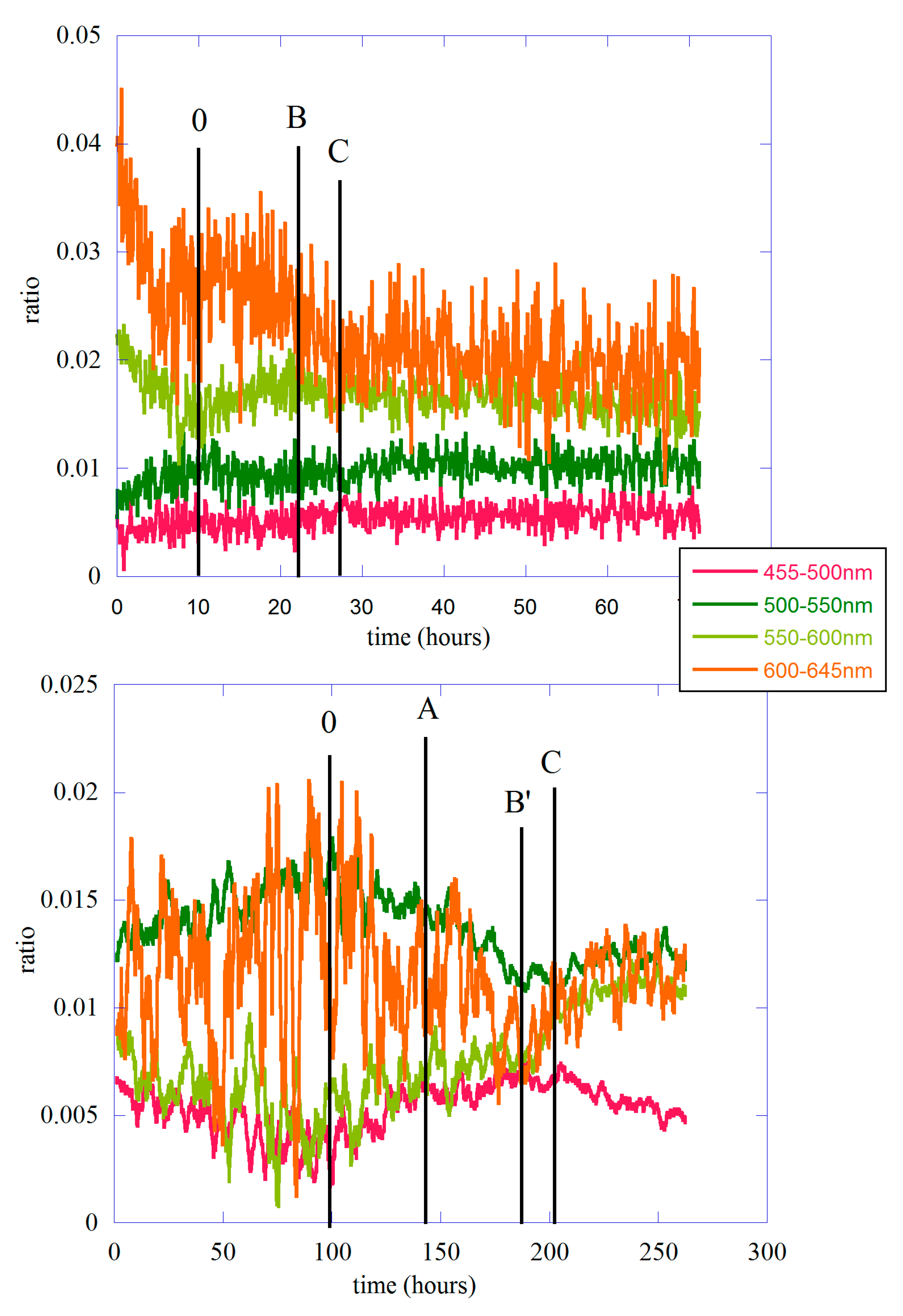
2.2. Data Analysis II—The Different Spectral Components
3. Conclusions and Suggestions for Future Works
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References and Notes
- Gurwitsch, A.G. Die Natur des spezifischen Erregers der Zellteilung. Arch. Entw. Mech. Org. 1923, 100, 11. [Google Scholar] [CrossRef]
- Reiter, T.; Gabor, D. Ultraviolette Strahlung und Zellteilung. In Wissenschaftliche Verffentlichungen aus dem Siemens-Konzern; Springer: Berlin/Heidelberg, Germany, 1928. [Google Scholar]
- Colli, L.; Facchini, U. Light Emission by Germinating Plants. Il Nuovo C. 1954, 12, 150. [Google Scholar] [CrossRef]
- Colli, L.; Facchini, U.; Guidotti, G.; Dugnani Lonati, R.; Orsenigo, M.; Sommariva, O. Further Measurements on the Bioluminescence of the Seedlings. Experientia 1955, 11, 479. [Google Scholar] [CrossRef]
- Popp, F.A.; Gu, Q.; Li, K.H. Biophoton Emission: Experimental Background and Theoretical Approaches. Mod. Phys. Lett. B 1994, 8, 1269. [Google Scholar] [CrossRef]
- Van Wijk, R. Light in Shaping Life: Biophotons in Biology and Medicine; Boeken Service: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Slawinski, J.; Ezzahir, A.; Godlewski, M.; Kwiecinska, T.; Rajfur, Z.; Sitko, D.; Wierzuchowska, D. Stress-induced photon emission from perturbed organisms. Experientia 1992, 48, 1041. [Google Scholar] [CrossRef] [PubMed]
- Brizhik, L.; Scordino, A.; Triglia, A.; Musumeci, F. Delayed luminescence of biological systems arising from correlated many-soliton states. Phys. Rev. E 2001, 64, 031902. [Google Scholar] [CrossRef]
- Scordino, A.; Grasso, R.; Gulino, M.; Lanzanò, L.; Musumeci, F.; Privitera, G.; Tedesco, M.; Triglia, A.; Brizhik, L. Delayed luminescence from collagen as arising from soliton and small polaron states. Int. J. Quantum Chem. 2010, 110, 221. [Google Scholar] [CrossRef]
- Bajpai, R.P. Quantum coherence of biophotons and living systems. Indian J. Exp. Biol. 2003, 41, 514. [Google Scholar]
- Pospíšil, P.; Prasad, A.; Rác, M. Role of reactive oxygen species in ultra-weak photon emission in biological systems. J. Photochem. Photobiol. B 2014, 139, 11. [Google Scholar] [CrossRef]
- Mauburov, S.N. Photonic Communications in Biological Systems. J. Samara State Tech. Univ. Ser. Phys. Math. Sci. 2011, 15, 260. [Google Scholar]
- Kucera, O.; Cifra, M. Cell-to-cell signaling through light: Just a ghost of chance? Cell Comm. Signal. 2013, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Fels, D. Cellular Communication through light. PLoS ONE 2009, 4, e5086. [Google Scholar] [CrossRef]
- Beloussov, L.V.; Burlakov, A.B.; Louchinskaia, N.N. Biophotonic Pattern of optical interaction between fish eggs and embryos. Indian J. Exp. Biol. 2003, 41, 424–430. [Google Scholar] [PubMed]
- Hunt von Herbing, I.; Tonello, L.; Benfatto, M.; Pease, A.; Grigolini, P. Crucial Development: Criticality Is Important to Cell-to-Cell Communication and Information Transfer in Living Systems. Entropy 2021, 23, 1141. [Google Scholar] [CrossRef]
- Benfatto, M.; Pace, E.; Curceanu, C.; Scordo, A.; Clozza, A.; Davoli, I.; Lucci, M.; Francini, R.; De Matteis, F.; Grandi, M.; et al. Biophotons and Emergence of Quantum Coherence—A Diffusion Entropy Analysis. Entropy 2021, 23, 554. [Google Scholar] [CrossRef]
- Allegrini, P.; Grigolini, P.; Hamilton, P.; Palatella, L.; Raffaelli, G. Memory beyond memory in heart heating, a sign of a healthy physiological condition. Phys. Rev. E 2002, 65, 041926. [Google Scholar] [CrossRef]
- Allegrini, P.; Benci, V.; Grigolini, P.; Hamilton, P.; Ignaccolo, M.; Menconi, G.; Palatella, L.; Raffaelli, G.; Scafetta, N.; Virgilio, M.; et al. Compression and diffusion: A joint approach to detect complexity. Chaos Solitons Fractals 2003, 15, 517–535. [Google Scholar] [CrossRef]
- Lukovic, M.; Grigolini, P. Power spectra for both interrupted and perennial aging process. J. Chem. Phys. 2008, 129, 184102. [Google Scholar] [CrossRef]
- Jelinek, H.F.; Tuladhar, R.; Culbreth, G.; Bohara, G.; Cornforth, D.; West, B.J.; Grigolini, P. Diffusion Entropy versus Multiscale and Renyi Entropy to detect progression of Autonomic Neuropathy. Front. Physiol. 2020, 11, 607324. [Google Scholar] [CrossRef]
- Mancuso, S. The Revolutionary Genius of Plants: A New Understanding of Plant Intelligence and Behaviour; Simon and Shuster: New York, NY, USA, 2018. [Google Scholar]
- Mancuso, S.; Viola, A. Brilliant Green: The Surprising History and Science of Plant Intelligence; Island Press: Washington, DC, USA, 2015. [Google Scholar]
- All Fits Are Done Using the KeleidaGraph Version 5.02—Synergy Software—For Details See the User Guide. Available online: https://www.synergy.com/ (accessed on 20 February 2021).
- The logistic equation is an equation with the following form . this form has been known since the early 1800s and was used by P.F. Verhulst for studies on population growth. It has the characteristic that it has a solution that reaches a saturation limit.
- Gallep, C.M.; Dos Santos, S.R. Photon-count during germination of wheat (Triticum aestivum) in waste water sediment solution correlated with seedling growth. Seed Sci. Technol. 2007, 35, 607–614. [Google Scholar] [CrossRef]
- Saeidfirozeh, H.; Shafiekhani, A.; Cifra, M.; Masoudi, A.A. Endogenous Chemiluminescence from Germinating Arabidopsis thaliana Seeds. Sci. Rep. 2018, 8, 16231. [Google Scholar] [CrossRef] [PubMed]
- Supplemento Gazzetta Ufficiale—serie generale n.2 Allegato 1 A (4-1-1993).
- Loudon, R. The Quantum Theory of Light; Oxford University Press: Oxford, UK, 2000; ISBN 978-0-19-850176-3. [Google Scholar]
- Cifra, M.; Brouder, C.; Nerudova, M.; Kucera, O. Biophotons, coherence and photocount statistics: A critical review. J. Lumin. 2015, 164, 38. [Google Scholar] [CrossRef]
- Test Sheet Hamamatsu for the Phototube H12386-210, Serial Number 30050260. Available online: https://www.hamamatsu.com/resources/pdf/etd/H12386_TPMO1073E.pdf (accessed on 20 February 2021).
- Edmund Optics Long Pass Filters with These Names: RG-645, R-60, OG-550, Y-50, GG-455. Data Sheets Can Be Found on the Website of Edmund Optics. Available online: https://www.edmundoptics.com/ (accessed on 20 February 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Single Bean | Lentil Seeds | ||
|---|---|---|---|
| Time Interval | S Index | Time Interval | S Index |
| 82–265 | 0.50 | 10–70 | 0.23 |
| 82–150 | 0.81 | 20–70 | 0.24 |
| 150–200 | 0.30 | 50–70 | 0.28 |
| 200–265 | 0.43 | 35–36 | 0.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benfatto, M.; Pace, E.; Davoli, I.; Francini, R.; De Matteis, F.; Scordo, A.; Clozza, A.; De Paolis, L.; Curceanu, C.; Grigolini, P. Biophotons: New Experimental Data and Analysis. Entropy 2023, 25, 1431. https://doi.org/10.3390/e25101431
Benfatto M, Pace E, Davoli I, Francini R, De Matteis F, Scordo A, Clozza A, De Paolis L, Curceanu C, Grigolini P. Biophotons: New Experimental Data and Analysis. Entropy. 2023; 25(10):1431. https://doi.org/10.3390/e25101431
Chicago/Turabian StyleBenfatto, Maurizio, Elisabetta Pace, Ivan Davoli, Roberto Francini, Fabio De Matteis, Alessandro Scordo, Alberto Clozza, Luca De Paolis, Catalina Curceanu, and Paolo Grigolini. 2023. "Biophotons: New Experimental Data and Analysis" Entropy 25, no. 10: 1431. https://doi.org/10.3390/e25101431