Communicability Characterization of Structural DWI Subcortical Networks in Alzheimer’s Disease


 ,
,
Abstract
:1. Introduction
2. Materials
3. Methods
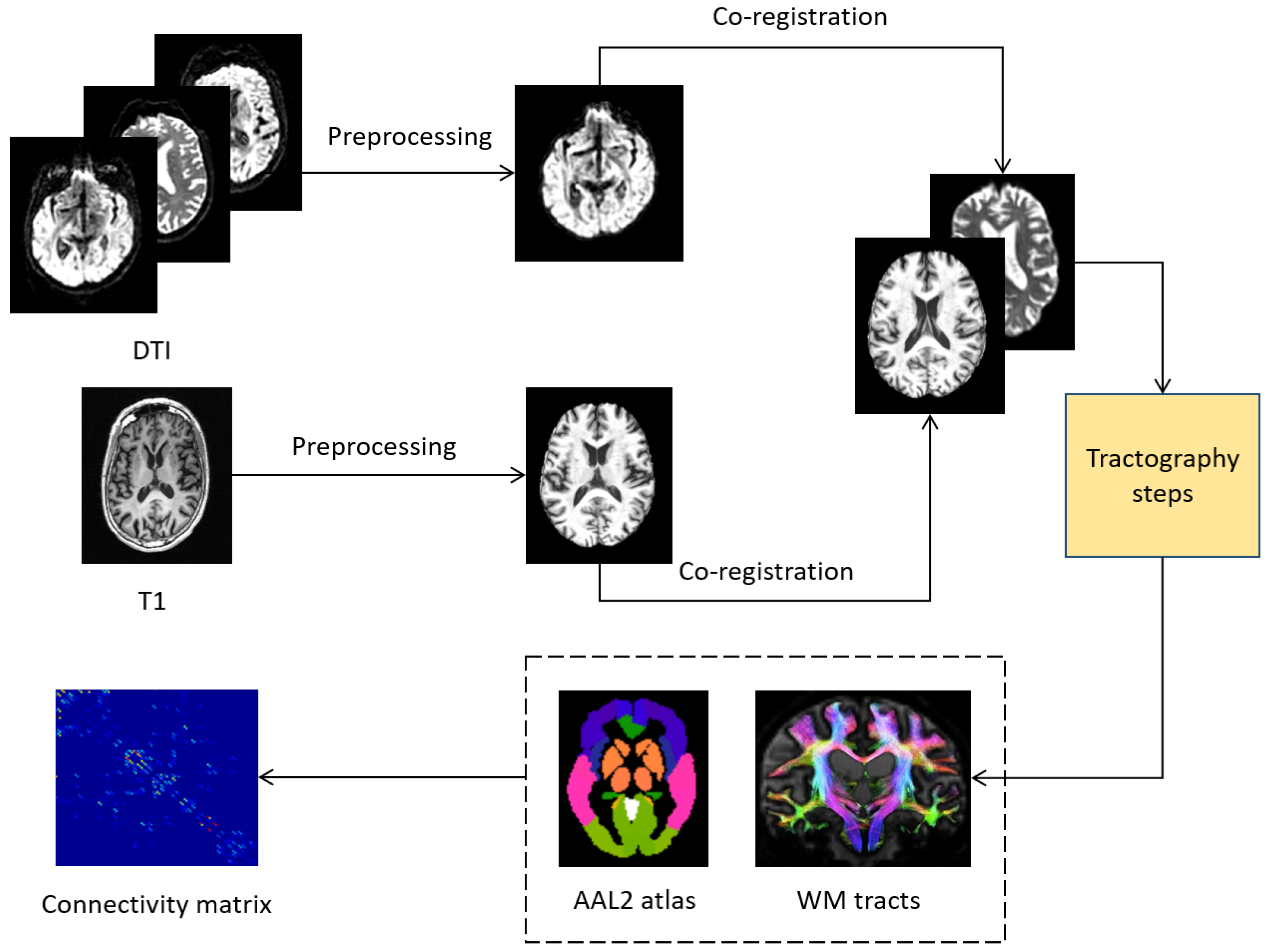
3.1. Image Processing
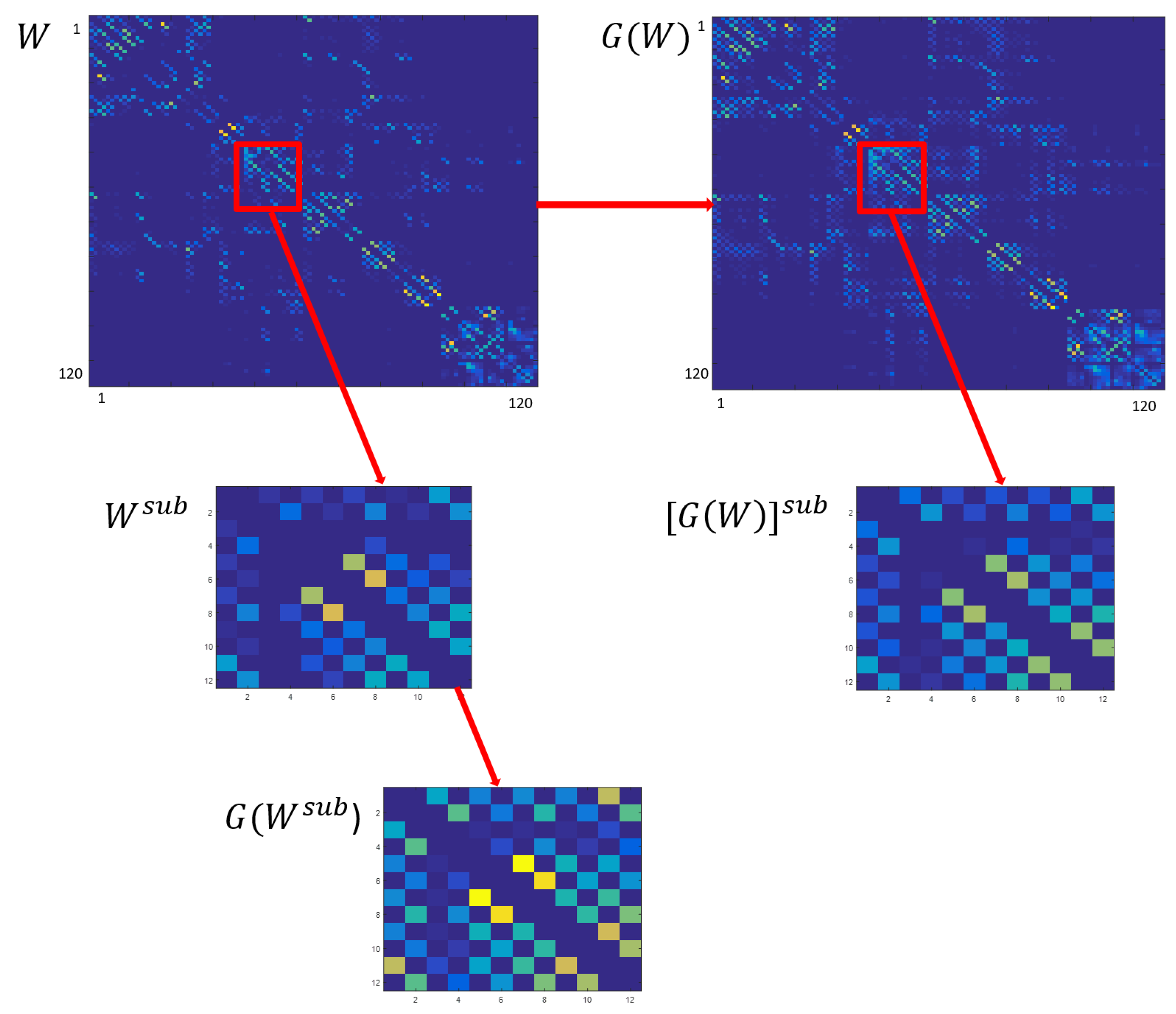
3.2. Subcortical Network Analysis
3.3. Group-Wise Statistical Analysis
3.4. Classification
4. Results
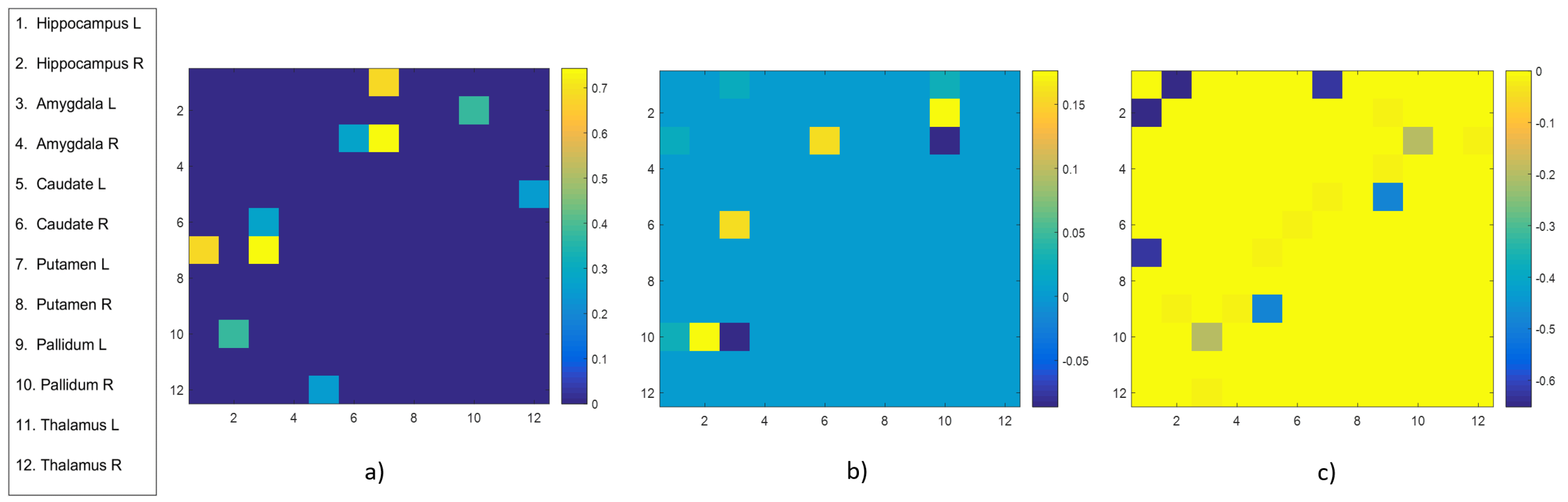
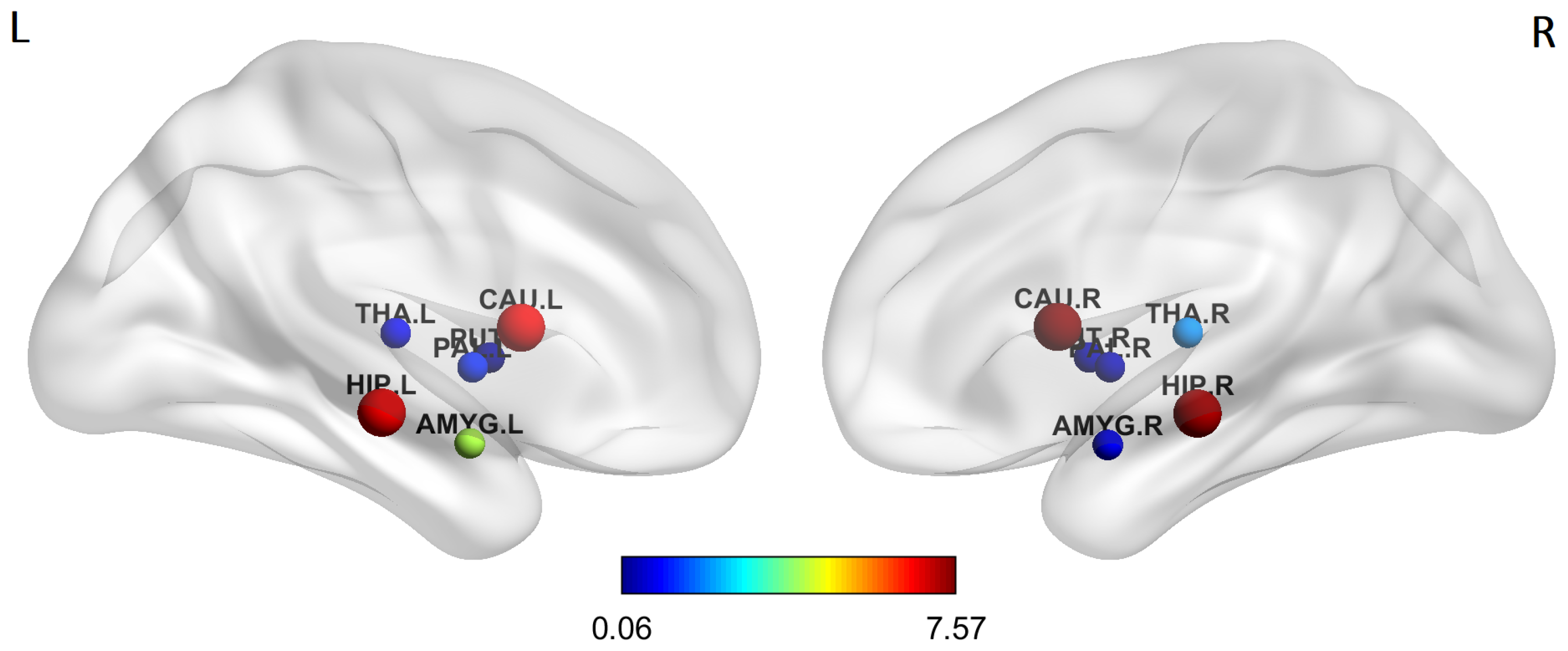
4.1. Group-Wise Statistical Analysis
4.2. Classification
5. Discussion and Conclusions
- (i)
- A group-wise statistical analysis has been performed to find subcortical brain region pairs with significantly different values of weights and communicability. The same analysis has been conducted to identify subcortical regions with different intra and inter-strength communicability, that are measures introduced to quantify the total intensity of subcortical nodes’ connectivity, in terms of communicability with the other subcortical nodes and with the rest of the whole network.
- (ii)
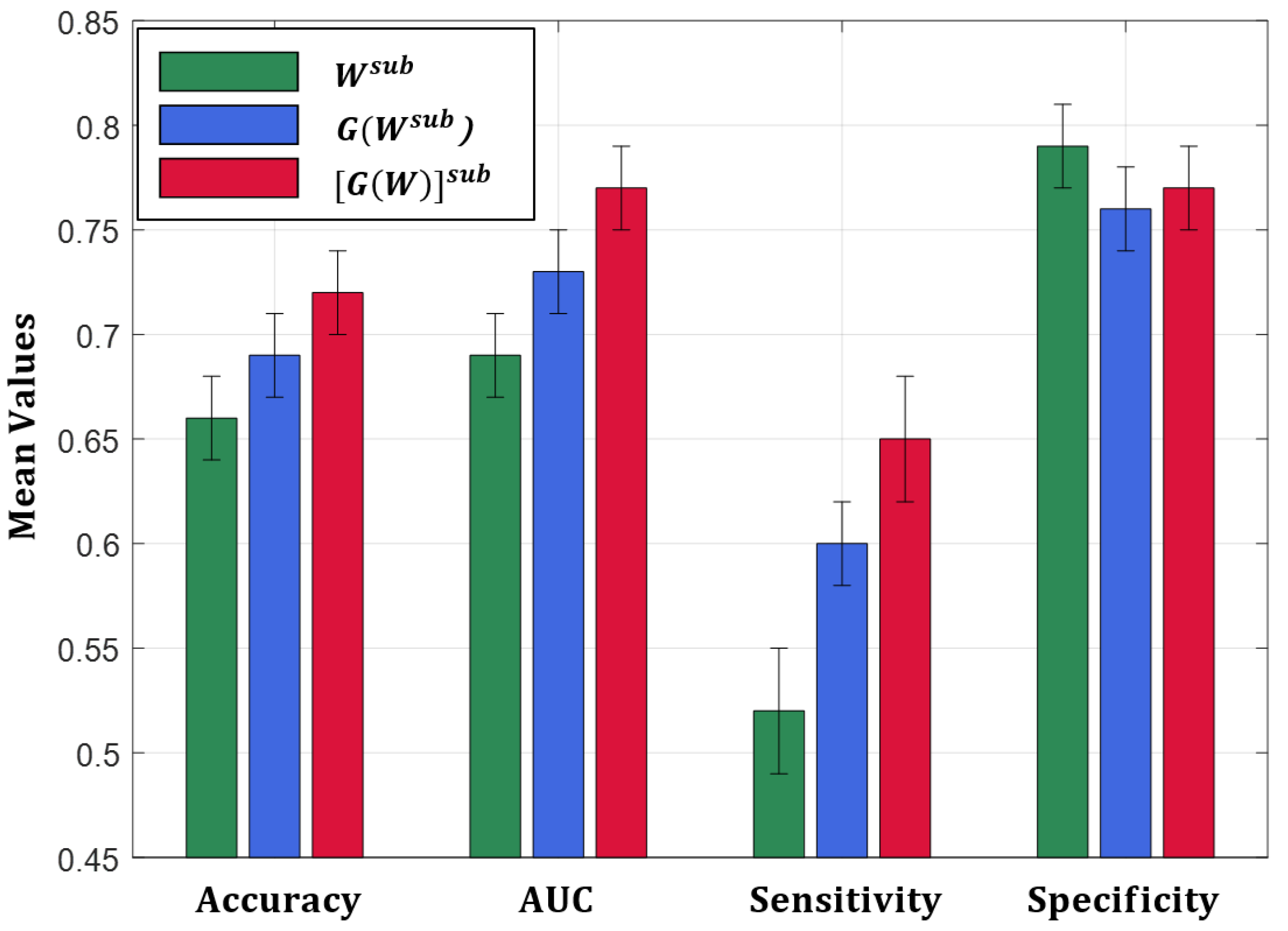
- A classification procedure has been adopted to investigate to which extent the sub-network communicability values and the extracted sub-network communicability between the subcortical regions are able to automatically discriminate between HC subjects and AD patients. The performance were also compared to the ones obtained using the subcortical edge weights as features for training the classification models.
- (i)
- The weights of brain networks, which have widespread use in literature to describe the brain connectivity, could not be informative enough, taken alone, to discriminate between HC and AD when relying on the subcortical regions’ connectivity.
- (ii)
- If the whole brain network communicability matrix is calculated and a sub-network communicability is extracted including the 12 subcortical regions (which are well-known AD related-brain regions), these features describe AD connectivity changes better than subcortical edge weights, and lead to better classification performance.
- (iii)
- Using the communicability metric gives a different viewpoint to describe the subcortical brain connectivity and allowed us to point out a sort of resilience mechanism of subcortical regions that tends to increase their communication (mainly through cortical nodes) in order to compensate the physical structural disconnection occurring between them because of AD.
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Sporns, O.; Tononi, G.; Kötter, R. The human connectome: A structural description of the human brain. PLoS Comput. Biol. 2005, 1, e42. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186. [Google Scholar] [CrossRef]
- Van den Heuvel, M.P.; Sporns, O. Network hubs in the human brain. Trends Cogn. Sci. 2013, 17, 683–696. [Google Scholar] [PubMed]
- Lombardi, A.; Tangaro, S.; Bellotti, R.; Bertolino, A.; Blasi, G.; Pergola, G.; Taurisano, P.; Guaragnella, C. A novel synchronization-based approach for functional connectivity analysis. Complexity 2017. [Google Scholar] [CrossRef]
- Lombardi, A.; Guaragnella, C.; Amoroso, N.; Monaco, A.; Fazio, L.; Taurisano, P.; Pergola, G.; Blasi, G.; Bertolino, A.; Bellotti, R.; et al. Modelling cognitive loads in schizophrenia by means of new functional dynamic indexes. NeuroImage 2019, 195, 150–164. [Google Scholar] [CrossRef]
- Tangaro, S.; Amoroso, N.; Brescia, M.; Cavuoti, S.; Chincarini, A.; Errico, R.; Inglese, P.; Longo, G.; Maglietta, R.; Tateo, A.; et al. Feature selection based on machine learning in MRIs for hippocampal segmentation. Comput. Math. Methods Med. 2015. [Google Scholar] [CrossRef]
- Fornito, A.; Zalesky, A.; Breakspear, M. The connectomics of brain disorders. Nat. Rev. Neurosci. 2015, 16, 159. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Association. 2017 Alzheimer’s disease facts and figures. Alzheimers Dement. 2017, 13, 325–373. [Google Scholar] [CrossRef]
- Delbeuck, X.; Van der Linden, M.; Collette, F. Alzheimer’s disease as a disconnection syndrome? Neuropsychol. Rev. 2003, 13, 79–92. [Google Scholar] [PubMed]
- Rose, S.E.; Chen, F.; Chalk, J.B.; Zelaya, F.O.; Strugnell, W.E.; Benson, M.; Doddrell, D.M. Loss of connectivity in Alzheimer’s disease: An evaluation of white matter tract integrity with colour coded MR diffusion tensor imaging. J. Neurol. Neurosurg. Psychiatry 2000, 69, 528–530. [Google Scholar]
- Le Bihan, D.; Mangin, J.F.; Poupon, C.; Clark, C.A.; Pappata, S.; Molko, N.; Chabriat, H. Diffusion tensor imaging: Concepts and applications. J. Magn. Reson. Imaging Off. J. Int. Soc. Magn. Reson. Med. 2001, 13, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Tournier, J.D.; Calamante, F.; Gadian, D.G.; Connelly, A. Direct estimation of the fiber orientation density function from diffusion-weighted MRI data using spherical deconvolution. NeuroImage 2004, 23, 1176–1185. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.Y.; Wang, P.N.; Chou, K.H.; Wang, J.; He, Y.; Lin, C.P. Diffusion tensor tractography reveals abnormal topological organization in structural cortical networks in Alzheimer’s disease. J. Neurosci. 2010, 30, 16876–16885. [Google Scholar] [CrossRef]
- Fischer, F.U.; Wolf, D.; Scheurich, A.; Fellgiebel, A. Altered whole-brain white matter networks in preclinical Alzheimer’s disease. NeuroImage Clin. 2015, 8, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Estrada, E.; Hatano, N. Communicability in complex networks. Phys. Rev. E 2008, 77, 036111. [Google Scholar] [CrossRef]
- Lella, E.; Amoroso, N.; Lombardi, A.; Maggipinto, T.; Tangaro, S.; Bellotti, R. Communicability disruption in Alzheimer’s disease connectivity networks. J. Complex Netw. 2018, 7, 83–100. [Google Scholar] [CrossRef]
- Seo, E.H.; Lee, D.Y.; Lee, J.M.; Park, J.S.; Sohn, B.K.; Lee, D.S.; Choe, Y.M.; Woo, J.I. Whole-brain functional networks in cognitively normal, mild cognitive impairment, and Alzheimer’s disease. PLoS ONE 2013, 8, e53922. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Chen, Z.; Evans, A. Structural insights into aberrant topological patterns of large-scale cortical networks in Alzheimer’s disease. J. Neurosci. 2008, 28, 4756–4766. [Google Scholar] [CrossRef]
- Tang, X.; Holland, D.; Dale, A.M.; Younes, L.; Miller, M.I.; Alzheimer’s Disease Neuroimaging Initiative. Shape abnormalities of subcortical and ventricular structures in mild cognitive impairment and Alzheimer’s disease: Detecting, quantifying, and predicting. Hum. Brain Mapp. 2014, 35, 3701–3725. [Google Scholar] [CrossRef]
- Son, S.J.; Kim, J.; Park, H. Structural and functional connectional fingerprints in mild cognitive impairment and Alzheimer’s disease patients. PLoS ONE 2017, 12, e0173426. [Google Scholar] [CrossRef]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-mental state”: A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Rosen, W.G.; Mohs, R.C.; Davis, K.L. A new rating scale for Alzheimer’s disease. Am. J. Psychiatry 1984, 141, 1356–1364. [Google Scholar]
- Mohs, R.C.; Knopman, D.; Petersen, R.C.; Ferris, S.H.; Ernesto, C.; Grundman, M.; Thal, L.J. Development of cognitive instruments for use in clinical trials of antidementia drugs: Additions to the Alzheimer’s Disease Assessment Scale that broaden its scope. Alzheimer Dis. Assoc. Disord. 1997, 11, S13–S21. [Google Scholar] [CrossRef]
- Tournier, J.D.; Calamante, F.; Connelly, A. MRtrix: Diffusion tractography in crossing fiber regions. Int. J. Imaging Syst. Technol. 2012, 22, 53–66. [Google Scholar] [CrossRef]
- Tournier, J.D.; Smith, R.E.; Raffelt, D.A.; Tabbara, R.; Dhollander, T.; Pietsch, M.; Christiaens, D.; Jeurissen, B.; Yeh, C.H.; Connelly, A. MRtrix3: A fast, flexible and open software framework for medical image processing and visualisation. bioRxiv 2019, 551739. [Google Scholar]
- Jenkinson, M.; Beckmann, C.F.; Behrens, T.E.; Woolrich, M.W.; Smith, S.M. FSL. NeuroImage 2012, 62, 782–790. [Google Scholar] [CrossRef]
- Veraart, J.; Novikov, D.S.; Christiaens, D.; Ades-Aron, B.; Sijbers, J.; Fieremans, E. Denoising of diffusion MRI using random matrix theory. NeuroImage 2016, 142, 394–406. [Google Scholar] [CrossRef]
- Smith, S.M. Fast robust automated brain extraction. Hum. Brain Mapp. 2002, 17, 143–155. [Google Scholar] [CrossRef]
- Zhang, Y.; Brady, M.; Smith, S. Segmentation of brain MR images through a hidden Markov random field model and the expectation-maximization algorithm. IEEE Trans. Med. Imaging 2001, 20, 45–57. [Google Scholar] [CrossRef]
- Jeurissen, B.; Tournier, J.D.; Dhollander, T.; Connelly, A.; Sijbers, J. Multi-tissue constrained spherical deconvolution for improved analysis of multi-shell diffusion MRI data. NeuroImage 2014, 103, 411–426. [Google Scholar] [CrossRef]
- Tournier, J.-D.; Calamante, F.; Connelly, A. Improved probabilistic streamlines tractography by 2nd order integration over fibre orientation distributions. Proc. Int. Soc. Magn. Reson. Med. 2010, 18, 1670. [Google Scholar]
- Smith, R.E.; Tournier, J.D.; Calamante, F.; Connelly, A. SIFT2: Enabling dense quantitative assessment of brain white matter connectivity using streamlines tractography. NeuroImage 2015, 119, 338–351. [Google Scholar] [CrossRef]
- Smith, R.E.; Tournier, J.-D.; Calamante, F.; Connelly, A. Anatomically-constrained tractography: Improved diffusion MRI streamlines tractography through effective use of anatomical information. NeuroImage 2012, 62, 1924–1938. [Google Scholar] [CrossRef]
- Rolls, E.T.; Joliot, M.; Tzourio-Mazoyer, N. Implementation of a new parcellation of the orbitofrontal cortex in the automated anatomical labeling atlas. NeuroImage 2015, 122, 1–5. [Google Scholar] [CrossRef]
- Smith, R.E.; Tournier, J.D.; Calamante, F.; Connelly, A. The effects of SIFT on the reproducibility and biological accuracy of the structural connectome. NeuroImage 2015, 104, 253–265. [Google Scholar] [CrossRef]
- Amico, E.; Goñi, J. Mapping hybrid functional-structural connectivity traits in the human connectome. Netw. Neurosci. 2018, 2, 306–322. [Google Scholar] [CrossRef]
- Tipnis, U.; Amico, E.; Ventresca, M.; Goni, J. Modeling communication processes in the human connectome through cooperative learning. IEEE Trans. Netw. Sci. Eng. 2018. [Google Scholar] [CrossRef]
- Crofts, J.J.; Higham, D.J. A weighted communicability measure applied to complex brain networks. J. R. Soc. Interface 2009, 6, 411–414. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Lella, E.; Amoroso, N.; Bellotti, R.; Diacono, D.; La Rocca, M.; Maggipinto, T.; Tangaro, S. Machine learning for the assessment of Alzheimer’s disease through DTI. In Applications of Digital Image Processing XL; International Society for Optics and Photonics: Bellingham, WA, USA, 2017; Volume 10396, p. 1039619. [Google Scholar]
- Maggipinto, T.; Bellotti, R.; Amoroso, N.; Diacono, D.; Donvito, G.; Lella, E.; Monaco, A.; Scelsi, M.A.; Tangaro, S. DTI measurements for Alzheimer’s classification. Phys. Med. Biol. 2017, 62, 2361. [Google Scholar] [CrossRef]
- Vapnik, V. The Nature of Statistical Learning Theory; Springer Science and Business Media: Berlin, Germany, 2013. [Google Scholar]
- Yan, K.; Zhang, D. Feature selection and analysis on correlated gas sensor data with recursive feature elimination. Sens. Actuators B Chem. 2015, 212, 353–363. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Allen, G.; Barnard, H.; McColl, R.; Hester, A.L.; Fields, J.A.; Weiner, M.F.; Ringe, W.K.; Lipton, A.M.; Brooker, M.; McDonald, E.; et al. Reduced hippocampal functional connectivity in Alzheimer disease. Arch. Neurol. 2007, 64, 1482–1487. [Google Scholar] [CrossRef]
- Cuénod, C.A.; Denys, A.; Michot, J.L.; Jehenson, P.; Forette, F.; Kaplan, D.; Syrota, A.; Boller, F. Amygdala atrophy in Alzheimer’s disease: An in vivo magnetic resonance imaging study. Arch. Neurol. 1993, 50, 941–945. [Google Scholar] [CrossRef]
- De Jong, L.W.; Van der Hiele, K.; Veer, I.M.; Houwing, J.J.; Westendorp, R.G.J.; Bollen, E.L.E.M.; Van Der Grond, J. Strongly reduced volumes of putamen and thalamus in Alzheimer’s disease: An MRI study. Brain 2008, 131, 3277–3285. [Google Scholar] [CrossRef]
- Grady, C.L.; Furey, M.L.; Pietrini, P.; Horwitz, B.; Rapoport, S.I. Altered brain functional connectivity and impaired short-term memory in Alzheimer’s disease. Brain 2001, 124, 739–756. [Google Scholar] [CrossRef]
- Rombouts, S.A.; Barkhof, F.; Witter, M.P.; Scheltens, P. Unbiased whole-brain analysis of gray matter loss in Alzheimer’s disease. Neurosci. Lett. 2000, 285, 231–233. [Google Scholar] [CrossRef]
- Ryan, N.S.; Keihaninejad, S.; Shakespeare, T.J.; Lehmann, M.; Crutch, S.J.; Malone, I.B.; Thornton, J.S.; Mancini, L.; Hyare, H.; Yousry, T.; et al. Magnetic resonance imaging evidence for presymptomatic change in thalamus and caudate in familial Alzheimer’s disease. Brain 2013, 136, 1399–1414. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, L.; Thompson, P.M.; Jack, C.R., Jr.; Dodge, H.; Zhan, L.; Alzheimer’s Disease Neuroimaging Initiative. The added value of diffusion-weighted MRI-derived structural connectome in evaluating mild cognitive impairment: A multi-cohort validation. J. Alzheimers Disease 2018, 64, 149–169. [Google Scholar] [CrossRef]
- Crofts, J.J.; Higham, D.J.; Bosnell, R.; Jbabdi, S.; Matthews, P.M.; Behrens, T.E.J.; Johansen-Berg, H. Network analysis detects changes in the contralesional hemisphere following stroke. NeuroImage 2011, 54, 161–169. [Google Scholar] [CrossRef]
- Wang, L.; Zang, Y.; He, Y.; Liang, M.; Zhang, X.; Tian, L.; Li, K. Changes in hippocampal connectivity in the early stages of Alzheimer’s disease: Evidence from resting state fMRI. NeuroImage 2006, 31, 496–504. [Google Scholar] [CrossRef]
- Zhan, L.; Liu, Y.; Wang, Y.; Zhou, J.; Jahanshad, N.; Ye, J.; Thompson, P.M. Boosting brain connectome classification accuracy in Alzheimer’s disease using higher-order singular value decomposition. Front. Neurosci. 2015, 9, 257. [Google Scholar] [CrossRef]
- Prasad, G.; Joshi, S.H.; Nir, T.M.; Toga, A.W.; Thompson, P.M. Alzheimer’s Disease Neuroimaging Initiative (ADNI) Brain connectivity and novel network measures for Alzheimer’s disease classification. Neurobiol. Aging 2015, 36, S121–S131. [Google Scholar] [CrossRef]
- Beekly, D.L.; Ramos, E.M.; Lee, W.W.; Deitrich, W.D.; Jacka, M.E.; Wu, J.; Hubbard, J.L.; Koepsell, T.D.; Morris, J.C.; Kukull, W.A. The National Alzheimer’s Coordinating Center (NACC) database: The uniform data set. Alzheimer Dis. Assoc. Disord. 2007, 21, 249–258. [Google Scholar] [CrossRef]
- Impedovo, D.; Pirlo, G.; Vessio, G. Dynamic Handwriting Analysis for Supporting Earlier Parkinson’s Disease Diagnosis. Information 2018, 9, 247. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC (46) | AD (40) | p-Value | |
|---|---|---|---|
| Age | 0.31 | ||
| Gender | 21 M/25 F | 25 M/15 F | 0.11 |
| MMSE | <0.0001 | ||
| ADAS 11 | <0.0001 | ||
| ADAS 13 | <0.0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lella, E.; Amoroso, N.; Diacono, D.; Lombardi, A.; Maggipinto, T.; Monaco, A.; Bellotti, R.; Tangaro, S. Communicability Characterization of Structural DWI Subcortical Networks in Alzheimer’s Disease. Entropy 2019, 21, 475. https://doi.org/10.3390/e21050475
Lella E, Amoroso N, Diacono D, Lombardi A, Maggipinto T, Monaco A, Bellotti R, Tangaro S. Communicability Characterization of Structural DWI Subcortical Networks in Alzheimer’s Disease. Entropy. 2019; 21(5):475. https://doi.org/10.3390/e21050475
Chicago/Turabian StyleLella, Eufemia, Nicola Amoroso, Domenico Diacono, Angela Lombardi, Tommaso Maggipinto, Alfonso Monaco, Roberto Bellotti, and Sabina Tangaro. 2019. "Communicability Characterization of Structural DWI Subcortical Networks in Alzheimer’s Disease" Entropy 21, no. 5: 475. https://doi.org/10.3390/e21050475