
Circadian Rhythms in NLRP3 Inflammasome Regulation: Possible Implications for the Nighttime Risk of Gout Flares


Abstract
:1. Introduction
2. Circadian Rhythms
3. The Transcription/Translation Feedback Loop (TTFL) Circadian Clock
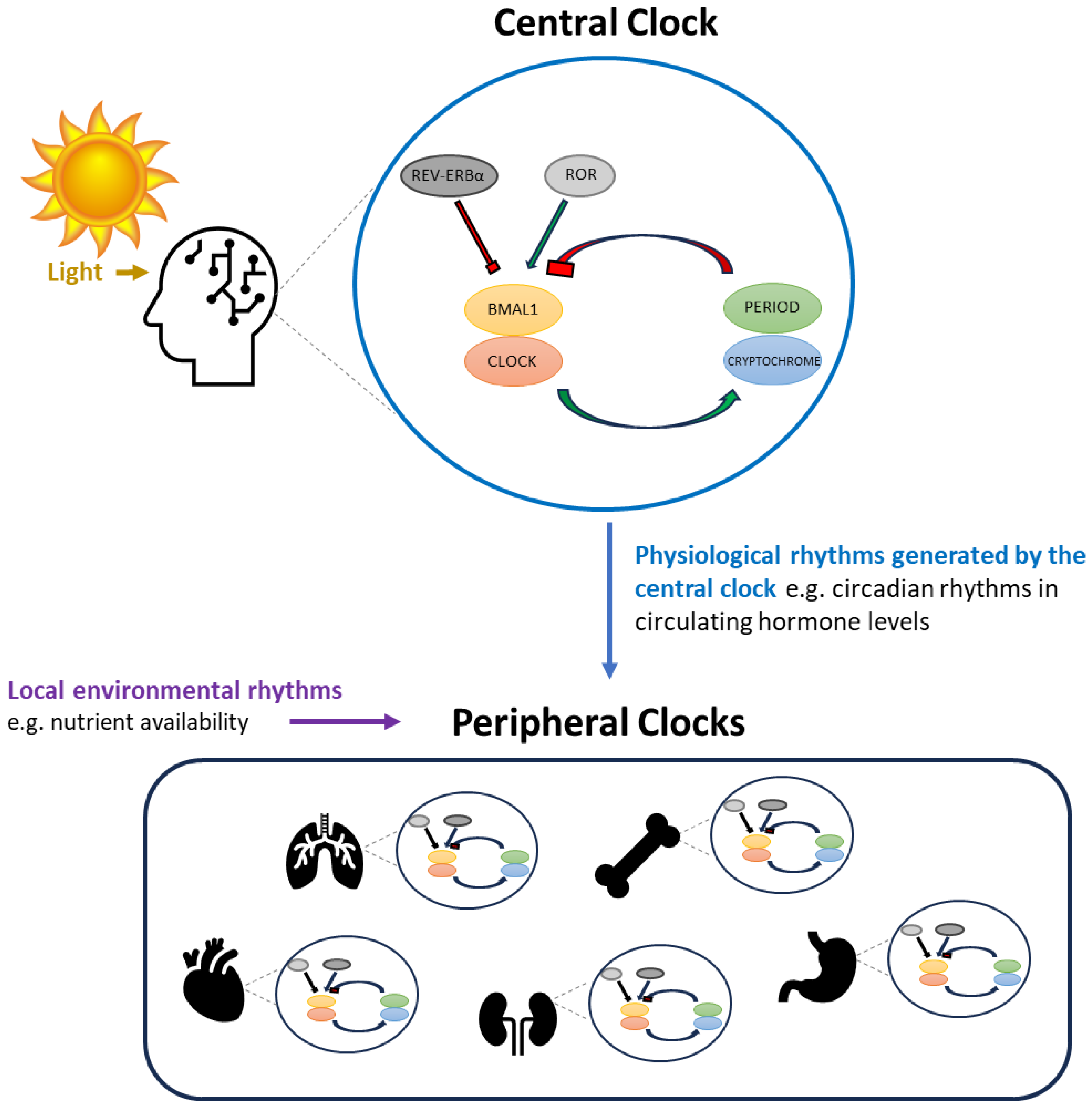
4. Central versus Peripheral Circadian Clocks
5. Circadian Rhythms in Immune Function and the NLRP3 Inflammasome
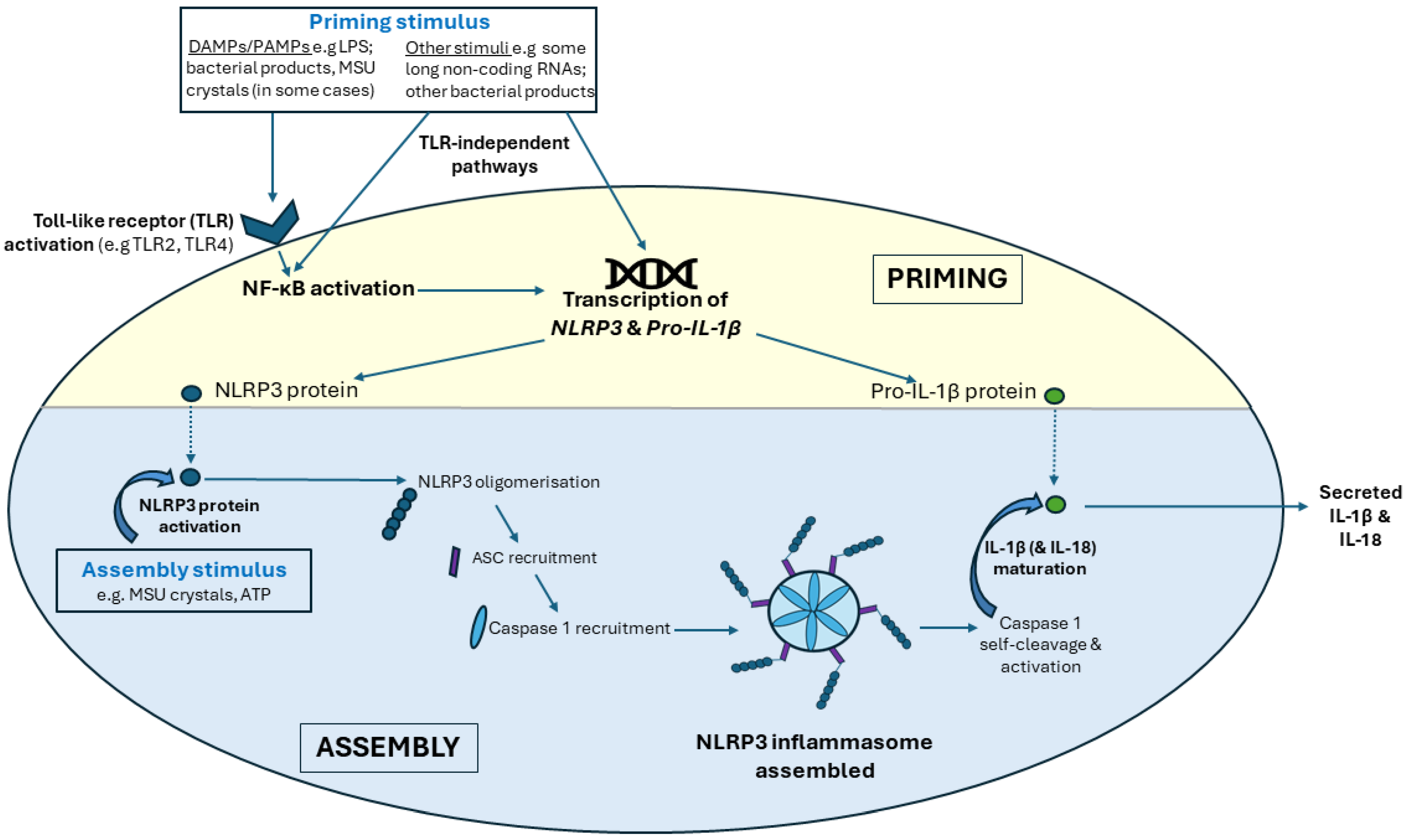
5.1. The NLRP3 Inflammasome
5.1.1. NLRP3 Inflammasome Priming
5.1.2. NLRP3 Inflammasome Assembly
5.1.3. NLRP3 Inflammasome Activation in Gout Flares
6. Mechanisms for Circadian Control of NLRP3 Inflammasome Activity: Role of the Central Clock
6.1. Melatonin
Melatonin, the NLRP3 Inflammasome and Gout
6.2. Glucocorticoids
Glucocorticoids, the NLRP3 Inflammasome and Gout
6.3. Melanocortins: ACTH and MSHs
ACTH, MSHs, the NLRP3 Inflammasome and Gout
6.4. Other Mechanisms by Which the Central Clock May Influence Gout Flares
7. Mechanisms for Circadian Control of NLRP3 Inflammasome Activity: Role of Peripheral Clocks
7.1. Peripheral Clock Control of NLRP3, TLRs and NF-κB
7.2. Emerging Evidence for Disruption of the Macrophage Peripheral Circadian Clock in Gout
8. Summary of the Effect of Circadian Clocks on the NLRP3 Inflammasome
Limitations of Existing Studies
9. Circadian Involvement in Gout Flares in the Broader Context of Circadian Involvement in Inflammatory Diseases
10. Current Gout Flare Treatments and Circadian Regulation of Immune Function
11. Conclusions and Future Direction
- determining whether circadian rhythms in circulating hormone levels contribute to time-of-day susceptibility to NLRP3 inflammasome-activating stimuli. At present, there are no data demonstrating that physiologically relevant concentrations of melatonin or MSHs/ACTH can regulate the NLRP3 inflammasome, although data from pre-clinical models suggest that both may have efficacy in reducing inflammation when used pharmacologically. Similarly, knowledge regarding the effects of circadian rhythms in relation to endogenous glucocorticoid use and susceptibility to gout flares is also limited. In particular, the potential effect of glucocorticoids in promoting NLRP3 inflammasome priming has been largely unexplored in the context of gout. However, given that other nuclear receptors involved in regulating NLRP3 priming are also regulated by the circadian clock, this may be relevant for understanding the overall time-of-day susceptibility to NLRP3 inflammasome priming.
- identifying whether there are time-of-day differences in the response of immune cells such as macrophages to stress-induced increases in endogenous levels of hormones, e.g., glucocorticoids. Given that there are circadian rhythms in GR expression on target cells and that the profile of GR-regulated genes involved in energy metabolism has been shown to differ depending on the time of day of glucocorticoid exposure, it seems highly likely that the magnitude of the effect of stress-induced upregulation of glucocorticoid production on NLRP3 inflammasome activation and inflammation differs depending on the time of day. This may be relevant for understanding the link between stress and risk of gout flares.
- confirming whether components of the NLRP3 inflammasome pathway are also expressed at higher levels in macrophages at night in humans, as has been observed in mice. There is compelling evidence that the NLRP3 inflammasome is under circadian control in both murine and human macrophages. However, it is unclear whether circadian control of the inflammasome is the same in both species and, in particular, whether the day/night patterning is the same in diurnal humans as in nocturnal mice. Understanding this is critical for understanding the potential consequences of circadian control of the NLRP3 inflammasome for time-of-day susceptibility to activating stimuli.
- establishing whether the changes in the expression of circadian clock components observed in MSU crystal-exposed THP-1 cells in vitro are also apparent in macrophages surrounding MSU crystal deposits within gout joints in vivo. Although essential for understanding the translatability of in vitro findings, there are a number of barriers to performing this type of study in human tissue. The timing of tissue collection would need to be carefully considered, as would the type of tissue used as a comparator. In this regard, a combination of both human studies and studies in animal models may be necessary. The circadian clock is highly conserved across species, and therefore it is reasonable to use animal models to study factors that regulate the clock. While murine models have been extensively utilized for circadian clock research in other disease contexts, in relation to studies of gout flares, where there is a specific question about the day/night association of effects, diurnal models, for instance, zebrafish, may have greater utility.
- determining how central and peripheral clock mechanisms interact to ultimately control NLRP3 inflammasome activity. Circulating hormone levels controlled by the central clock as well as immune cell-specific peripheral clocks can regulate NLRP3 inflammasome activity. In addition, immune cell numbers oscillate over the course of a day, and circadian rhythms in phagocytosis have also been observed, both of which may also alter the overall extent of NLRP3 inflammasome activity in a joint. Understanding the interaction between the different levels of circadian control of immune cell function and inflammation as well as the impact of diurnal rhythms of exposure to inflammasome-activating stimuli (e.g., alcohol) is critical for ultimately understanding nighttime susceptibility to gout flares.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sydenham, T. Tractatus de Podagra et Hydrope; G Kettilby: London, UK, 1683. [Google Scholar]
- Petrie, K.J.; MacKrill, K.; Silvester, C.; Gamble, G.D.; Dalbeth, N.; Pennebaker, J.W. OK Computer? A Time Analysis of Google Searches About Symptoms. Clin. Psychol. Eur. 2019, 1, 32774. [Google Scholar] [CrossRef]
- Choi, H.K.; Niu, J.; Neogi, T.; Chen, C.A.; Chaisson, C.; Hunter, D.; Zhang, Y. Nocturnal risk of gout attacks. Arthritis Rheumatol. 2015, 67, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Aschoff, J. Freerunning and entrained circadian rhythms. In Biological Rhythms; Springer: Berlin/Heidelberg, Germany, 1981; pp. 81–93. [Google Scholar]
- De Mairan, J.J. d’Ortous Observation botanique. Hist. l’Academie R. Des Sci. 1729, 35–36. [Google Scholar]
- Czeisler, C.A.; Richardson, G.S.; Zimmerman, J.C.; Moore-Ede, M.C.; Weitzman, E.D. Entrainment of human circadian rhythms by light-DARK cycles: A reassessment. Photochem. Photobiol. 1981, 34, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, S.; Tsuji, K.; Kondo, K. Strain differences of the mouse’s free-running circadian rhythm in continuous darkness. Physiol. Behav. 1978, 20, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Hardin, P.E.; Hall, J.C.; Rosbash, M. Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature 1990, 343, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Scheiermann, C.; Kunisaki, Y.; Frenette, P.S. Circadian control of the immune system. Nat. Rev. Immunol. 2013, 13, 190–198. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J.S.; Reddy, A.B. Circadian clocks in human red blood cells. Nature 2011, 469, 498–503. [Google Scholar] [CrossRef]
- Yu, W.; Nomura, M.; Ikeda, M. Interactivating feedback loops within the mammalian clock: BMAL1 is negatively autoregulated and upregulated by CRY1, CRY2, and PER2. Biochem. Biophys. Res. Commun. 2002, 290, 933–941. [Google Scholar] [CrossRef]
- Guillaumond, F.; Dardente, H.; Giguere, V.; Cermakian, N. Differential control of Bmal1 circadian transcription by REV-ERB and ROR nuclear receptors. J. Biol. Rhythm. 2005, 20, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Stephan, F.K. The “other” circadian system: Food as a zeitgeber. J. Biol. Rhythm. 2002, 17, 284–292. [Google Scholar] [CrossRef]
- Cao, R.; Gkogkas, C.G.; de Zavalia, N.; Blum, I.D.; Yanagiya, A.; Tsukumo, Y.; Xu, H.; Lee, C.; Storch, K.F.; Liu, A.C.; et al. Light-regulated translational control of circadian behavior by eIF4E phosphorylation. Nat. Neurosci. 2015, 18, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Mavroudis, P.D.; Scheff, J.D.; Calvano, S.E.; Lowry, S.F.; Androulakis, I.P. Entrainment of peripheral clock genes by cortisol. Physiol. Genom. 2012, 44, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Golombek, D.A.; Rosenstein, R.E. Physiology of Circadian Entrainment. Physiol. Rev. 2010, 90, 1063–1102. [Google Scholar] [CrossRef] [PubMed]
- Honma, S. The mammalian circadian system: A hierarchical multi-oscillator structure for generating circadian rhythm. J. Physiol. Sci. 2018, 68, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.L.; Silver, R. Resetting the brain clock: Time course and localization of mPER1 and mPER2 protein expression in suprachiasmatic nuclei during phase shifts. Eur. J. Neurosci. 2004, 19, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef] [PubMed]
- Hattar, S.; Liao, H.W.; Takao, M.; Berson, D.M.; Yau, K.W. Melanopsin-containing retinal. ganglion cells: Architecture, projections, and intrinsic photosensitivity. Science 2002, 295, 1065–1070. [Google Scholar] [CrossRef]
- Cuesta, M.; Cermakian, N.; Boivin, D.B. Glucocorticoids entrain molecular clock components in human peripheral cells. FASEB J. 2015, 29, 1360–1370. [Google Scholar] [CrossRef]
- Damiola, F.; Le Minh, N.; Preitner, N.; Kornmann, B.; Fleury-Olela, F.; Schibler, U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes. Dev. 2000, 14, 2950–2961. [Google Scholar] [CrossRef] [PubMed]
- Stokkan, K.-A.; Yamazaki, S.; Tei, H.; Sakaki, Y.; Menaker, M. Entrainment of the Circadian Clock in the Liver by Feeding. Science 2001, 291, 490. [Google Scholar] [CrossRef]
- Lowrey, P.L.; Takahashi, J.S. Mammalian circadian biology: Elucidating genomewide levels of temporal organization. Ann. Rev. Genom. Hum. Genet. 2004, 5, 407–441. [Google Scholar] [CrossRef] [PubMed]
- Peek, C.B.; Ramsey, K.M.; Levine, D.C.; Marcheva, B.; Perelis, M.; Bass, J. Circadian regulation of cellular physiology. Methods Enzymol. 2015, 552, 165–184. [Google Scholar] [CrossRef]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and Peripheral Circadian Clocks in Mammals. Ann. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Ripperger, J.A.; Schibler, U. Rhythmic CLOCK-BMAL1 binding to multiple E-box motifs drives circadian Dbp transcription and chromatin transitions. Nat. Genet. 2006, 38, 369–374. [Google Scholar] [CrossRef]
- Takahata, S.; Sogawa, K.; Kobayashi, A.; Ema, M.; Mimura, J.; Ozaki, N.; Fujii-Kuriyama, Y. Transcriptionally Active Heterodimer Formation of an Arnt-like PAS Protein, Arnt3, with HIF-1a, HLF, and Clock. Biochem. Biophys. Res. Commun. 1998, 248, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2016, 18, 164. [Google Scholar] [CrossRef] [PubMed]
- Khapre, R.V.; Kondratova, A.A.; Patel, S.; Dubrovsky, Y.; Wrobel, M.; Antoch, M.P.; Kondratov, R.V. BMAL1-dependent regulation of the mTOR signaling pathway delays aging. Aging 2014, 6, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Dang, F.B.; Li, P.; Wang, P.F.; Xu, Q.; Liu, Z.S.; Li, Y.; Wu, Y.T.; Chen, Y.Q.; Liu, Y. The Circadian Protein Period2 Suppresses mTORC1 Activity via Recruiting Tsc1 to mTORC1 Complex. Cell Metab. 2019, 29, 653–667. [Google Scholar] [CrossRef]
- Collins, E.J.; Cervantes-Silva, M.P.; Timmons, G.A.; O’Siorain, J.R.; Curtis, A.M.; Hurley, J.M. Post-transcriptional circadian regulation in macrophages organizes temporally distinct immunometabolic states. Genome Res. 2021, 31, 171–185. [Google Scholar] [CrossRef]
- Halberg, F.; Johnson, E.A.; Brown, B.W.; Bittner, J.J. Susceptibility rhythm to E. coli endotoxin and bioassay. Proc. Soc. Exp. Biol. Med. 1960, 103, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Ella, K.; Csépányi-Kömi, R.; Káldi, K. Circadian regulation of human peripheral neutrophils. Brain. Behav. Immun. 2016, 57, 209–221. [Google Scholar] [CrossRef] [PubMed]
- De Bree, L.C.J.; Mourits, V.P.; Koeken, V.A.C.M.; Moorlag, S.J.; Janssen, R.; Folkman, L.; Barreca, D.; Krausgruber, T.; Fife-Gernedl, V.; Novakovic, B. Circadian rhythm influences induction of trained immunity by BCG vaccination. J. Clin. Investig. 2020, 130, 5603–5617. [Google Scholar] [CrossRef] [PubMed]
- Long, J.E.; Drayson, M.T.; Taylor, A.E.; Toellner, K.M.; Lord, J.M.; Phillips, A.C. Morning vaccination enhances antibody response over afternoon vaccination: A cluster-randomised trial. Vaccine 2016, 34, 2679–2685. [Google Scholar] [CrossRef]
- Dimitrov, S.; Benedict, C.; Heutling, D.; Westermann, J.; Born, J.; Lange, T. Cortisol and epinephrine control opposing circadian rhythms in T cell subsets. Blood, J. Am. Soc. Hematol. 2009, 113, 5134–5143. [Google Scholar] [CrossRef]
- Scheiermann, C.; Kunisaki, Y.; Lucas, D.; Chow, A.; Jang, J.-E.; Zhang, D.; Hashimoto, D.; Merad, M.; Frenette, P.S. Adrenergic nerves govern circadian leukocyte recruitment to tissues. Immunity 2012, 37, 290–301. [Google Scholar] [CrossRef]
- Martinon, F.; Pétrilli, V.; Mayor, A.; Tardivel, A.; Tschopp, J. Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature 2006, 440, 237–241. [Google Scholar] [CrossRef]
- Popov, D.; Jain, L.; Alhilali, M.; Dalbeth, N.; Poulsen, R.C. Monosodium urate crystals alter the circadian clock in macrophages leading to loss of NLRP3 inflammasome repression: Implications for timing of the gout flare. FASEB J. 2023, 37, e22940. [Google Scholar] [CrossRef]
- Busillo, J.M.; Azzam, K.M.; Cidlowski, J.A. Glucocorticoids sensitize the innate immune system through regulation of the NLRP3 inflammasome. J. Biol. Chem. 2011, 286, 38703–38713. [Google Scholar] [CrossRef]
- Liu, G.; Zhao, Y. Toll-like receptors and immune regulation: Their direct and indirect modulation on regulatory CD4+ CD25+ T cells. Immunology 2007, 122, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Kerur, N.; Hirano, Y.; Tarallo, V.; Fowler, B.J.; Bastos-Carvalho, A.; Yasuma, T.; Yasuma, R.; Kim, Y.; Hinton, D.R.; Kirschning, C.J.; et al. TLR-independent and P2X7-dependent signaling mediate Alu RNA-induced NLRP3 inflammasome activation in geographic atrophy. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7395–7401. [Google Scholar] [CrossRef] [PubMed]
- Cassel, S.L.; Eisenbarth, S.C.; Iyer, S.S.; Sadler, J.J.; Colegio, O.R.; Tephly, L.A.; Carter, A.B.; Rothman, P.B.; Flavell, R.A.; Sutterwala, F.S. The Nalp3 inflammasome is essential for the development of silicosis. Proc. Natl. Acad. Sci. USA 2008, 105, 9035–9040. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Guarda, G.; Romero, J.F.; Menu, P.; Gross, O.; Tardivel, A.; Suva, M.-L.; Stehle, J.-C.; Kopf, M.; Stamenkovic, I.; et al. Malarial hemozoin is a Nalp3 inflammasome activating danger signal. PLoS ONE 2009, 4, e6510. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Pétrilli, V.; Van Bruggen, R.; Steele, C.; Mossman, B.T.; Tschopp, J. Innate immune activation through Nalp3 inflammasome sensing of asbestos and silica. Science 2008, 320, 674–677. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Magupalli, V.G.; Ruan, J.; Yin, Q.; Atianand, M.K.; Vos, M.R.; Schröder, G.F.; Fitzgerald, K.A.; Wu, H.; Egelman, E.H. Unified polymerization mechanism for the assembly of ASC-dependent inflammasomes. Cell 2014, 156, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Boucher, D.; Monteleone, M.; Coll, R.C.; Chen, K.W.; Ross, C.M.; Teo, J.L.; Gomez, G.A.; Holley, C.L.; Bierschenk, D.; Stacey, K.J.; et al. Caspase-1 self-cleavage is an intrinsic mechanism to terminate inflammasome activity. J. Exp. Med. 2018, 215, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Liu-Bryan, R.; Scott, P.; Sydlaske, A.; Rose, D.M.; Terkeltaub, R. Innate immunity conferred by Toll-like receptors 2 and 4 and myeloid differentiation factor 88 expression is pivotal to monosodium urate monohydrate crystal-induced inflammation. Arthritis Rheum. 2005, 52, 2936–2946. [Google Scholar] [CrossRef]
- Yagnik, D.R.; Hillyer, P.; Marshall, D.; Smythe, C.D.; Krausz, T.; Haskard, D.O.; Landis, R.C. Noninflammatory phagocytosis of monosodium urate monohydrate crystals by mouse macrophages. Implications for the control of joint inflammation in gout. Arthritis Rheum. 2000, 43, 1779–1789. [Google Scholar] [CrossRef]
- Landis, R.C.; Yagnik, D.R.; Florey, O.; Philippidis, P.; Emons, V.; Mason, J.C.; Haskard, D.O. Safe disposal of inflammatory monosodium urate monohydrate crystals by differentiated macrophages. Arthritis Rheum. 2002, 46, 3026–3033. [Google Scholar] [CrossRef] [PubMed]
- Helget, L.N.; Mikuls, T.R. Environmental Triggers of Hyperuricemia and Gout. Rheum. Dis. Clin. N. Am. 2022, 48, 891–906. [Google Scholar] [CrossRef] [PubMed]
- Yue, T.F.; Gutman, A.B. Effect of allopurinol (4-hydroxypyrazolo-(3,4-D)pyrimidine) on serum and urinary uric acid in primary and secondary gout. Am. J. Med. 1964, 37, 885–898. [Google Scholar] [CrossRef] [PubMed]
- Borstad, G.C.; Bryant, L.R.; Abel, M.P.; Scroggie, D.A.; Harris, M.D.; Alloway, J.A. Colchicine for prophylaxis of acute flares when initiating allopurinol for chronic gouty arthritis. J. Rheumatol. 2004, 31, 2429–2432. [Google Scholar] [PubMed]
- Lang, F.; Greger, R.; Oberleithner, H.; Griss, E.; Lang, K.; Pastner, D.; Dittrich, P.; Deetjen, P. Renal handling of urate in healthy man in hyperuricaemia and renal insufficiency: Circadian fluctuation, effect of water diuresis and of uricosuric agents. Eur. J. Clin. Investig. 1980, 10, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Minors, D.S.; Waterhouse, J.M. Circadian rhythms of urinary excretion: The relationship between the amount excreted and the circadian changes. J. Physiol. 1982, 327, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Sennels, H.P.; Jørgensen, H.L.; Goetze, J.P.; Fahrenkrug, J. Rhythmic 24-h variations of frequently used clinical biochemical parameters in healthy young males—The Bispebjerg study of diurnal variations. Scand. J. Clin. Lab. Investig. 2012, 72, 287–295. [Google Scholar] [CrossRef]
- Devgun, M.S.; Dhillon, H.S. Importance of diurnal variations on clinical value and interpretation of serum urate measurements. J. Clin. Pathol. 1992, 45, 110–113. [Google Scholar] [CrossRef]
- Shimizu, M.; Naito, R.; Sato, A.; Ishiwata, S.; Yatsu, S.; Shitara, J.; Matsumoto, H.; Murata, A.; Kato, T.; Suda, S.; et al. Diurnal Variations in Serum Uric Acid, Xanthine, and Xanthine Oxidoreductase Activity in Male Patients with Coronary Artery Disease. Nutrients 2023, 15, 4480. [Google Scholar] [CrossRef]
- Kanemitsu, T.; Tsurudome, Y.; Kusunose, N.; Oda, M.; Matsunaga, N.; Koyanagi, S.; Ohdo, S. Periodic variation in bile acids controls circadian changes in uric acid via regulation of xanthine oxidase by the orphan nuclear receptor PPARα. J. Biol. Chem. 2017, 292, 21397–21406. [Google Scholar] [CrossRef]
- Cutolo, M.; Seriolo, B.; Craviotto, C.; Pizzorni, C.; Sulli, A. Circadian rhythms in RA. Ann. Rheum. Dis. 2003, 62, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Arvidson, N.G.; Gudbjörnsson, B.; Elfman, L.; Rydén, A.C.; Tötterman, T.H.; Hällgren, R. Circadian rhythm of serum interleukin-6 in rheumatoid arthritis. Ann. Rheum. Dis. 1994, 53, 521–524. [Google Scholar] [CrossRef]
- Voisey, J.; Carroll, L.; van Daal, A. Melanocortins and their receptors and antagonists. Curr. Drug Targets 2003, 4, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Tosini, G.; Davidson, A.J.; Fukuhara, C.; Kasamatsu, M.; Castanon-Cervantes, O. Localization of a circadian clock in mammalian photoreceptors. FASEB J. 2007, 21, 3866–3871. [Google Scholar] [CrossRef]
- Cecon, E.; Markus, R.P. Relevance of the chronobiological and non-chronobiological actions of melatonin for enhancing therapeutic efficacy in neurodegenerative disorders. Recent. Pat. Endocr. Metab. Immune Drug Discov. 2011, 5, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Muxel, S.M.; Pires-Lapa, M.A.; Monteiro, A.W.A.; Cecon, E.; Tamura, E.K.; Floeter-Winter, L.M.; Markus, R.P. NF-κB drives the synthesis of melatonin in RAW 264.7 macrophages by inducing the transcription of the arylalkylamine-N-acetyltransferase (AA-NAT) gene. PLoS ONE 2012, 7, e52010. [Google Scholar] [CrossRef]
- Tan, D.-X.; Chen, L.-D.; Poeggeler, B.; Manchester, L.; Reiter, R. Melatonin: A potent, endogenous hydroxyl radical scavenger. Endocr. J. 1993, 1, 57–60. [Google Scholar]
- Becker-André, M.; Wiesenberg, I.; Schaeren-Wiemers, N.; André, E.; Missbach, M.; Saurat, J.H.; Carlberg, C. Pineal gland hormone melatonin binds and activates an orphan of the nuclear receptor superfamily. J. Biol. Chem. 1994, 269, 28531–28534. [Google Scholar] [CrossRef]
- Ma, H.; Kang, J.; Fan, W.; He, H.; Huang, F. ROR: Nuclear Receptor for Melatonin or Not? Molecules 2021, 26, 2693. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Jetten, A.M. RORα is not a receptor for melatonin. BioEssays 2016, 38, 1193–1194. [Google Scholar] [CrossRef]
- Masana, M.I.; Benloucif, S.; Dubocovich, M.L. Circadian rhythm of mt1 melatonin receptor expression in the suprachiasmatic nucleus of the C3H/HeN mouse. J. Pineal Res. 2000, 28, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Pozo, D.; García-Mauriño, S.; Guerrero, J.M.; Calvo, J.R. mRNA expression of nuclear receptor RZR/RORalpha, melatonin membrane receptor MT, and hydroxindole-O-methyltransferase in different populations of human immune cells. J. Pineal Res. 2004, 37, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Calvo, J.R.; González-Yanes, C.; Maldonado, M.D. The role of melatonin in the cells of the innate immunity: A review. J. Pineal Res. 2013, 55, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Nejati Moharrami, N.; Bjørkøy Tande, E.; Ryan, L.; Espevik, T.; Boyartchuk, V. RORα controls inflammatory state of human macrophages. PLoS ONE 2018, 13, e0207374. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, F.; Acuña-Castroviejo, D.; Doerrier, C.; Dayoub, J.C.; López, L.C.; Venegas, C.; García, J.A.; López, A.; Volt, H.; Luna-Sánchez, M.; et al. Melatonin blunts the mitochondrial/NLRP3 connection and protects against radiation-induced oral mucositis. J. Pineal Res. 2015, 58, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Gil, B.; Moneim, A.E.A.; Ortiz, F.; Shen, Y.-Q.; Soto-Mercado, V.; Mendivil-Perez, M.; Guerra-Librero, A.; Acuña-Castroviejo, D.; Molina-Navarro, M.M.; García-Verdugo, J.M.; et al. Melatonin protects rats from radiotherapy-induced small intestine toxicity. PLoS ONE 2017, 12, e0174474. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, L.; Wang, Z.; Li, C.; Li, S.; Li, L.; Fan, Q.; Zheng, L. Melatonin Suppresses Estrogen Deficiency-Induced Osteoporosis and Promotes Osteoblastogenesis by Inactivating the NLRP3 Inflammasome. Calcif. Tissue Int. 2018, 103, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.-Z.; Zhao, L.; Cao, L.; Zhu, X.-X.; Zou, H.-J. Melatonin Alleviates Acute Gouty Inflammation In Vivo and In Vitro. Curr. Med. Sci. 2021, 41, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Arendt, J.; Aulinas, A. Physiology of the Pineal Gland and Melatonin. [Updated 2022 Oct 30]. In Endotext [Internet]; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., Dhatariya, K., Dungan, K., Hofland, H., Kaira, S., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books (accessed on 15 January 2024).
- Yao, Z.; DuBois, D.C.; Almon, R.R.; Jusko, W.J. Modeling circadian rhythms of glucocorticoid receptor and glutamine synthetase expression in rat skeletal muscle. Pharm. Res. 2006, 23, 670–679. [Google Scholar] [CrossRef]
- Caratti, G.; Iqbal, M.; Hunter, L.; Kim, D.; Wang, P.; Vonslow, R.M.; Begley, N.; Tetley, A.J.; Woodburn, J.L.; Pariollaud, M.; et al. REVERBa couples the circadian clock to hepatic glucocorticoid action. J. Clin. Investig. 2018, 128, 4454–4471. [Google Scholar] [CrossRef]
- Nader, N.; Chrousos, G.P.; Kino, T. Circadian rhythm transcription factor CLOCK regulates the transcriptional activity of the glucocorticoid receptor by acetylating its hinge region lysine cluster: Potential physiological implications. FASEB J. 2009, 23, 1572–1583. [Google Scholar] [CrossRef] [PubMed]
- Lamia, K.A.; Papp, S.J.; Yu, R.T.; Barish, G.D.; Uhlenhaut, N.H.; Jonker, J.W.; Downes, M.; Evans, R.M. Cryptochromes mediate rhythmic repression of the glucocorticoid receptor. Nature 2011, 480, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Charmandari, E.; Chrousos, G.P.; Lambrou, G.I.; Pavlaki, A.; Koide, H.; Ng, S.S.M.; Kino, T. Peripheral CLOCK regulates target-tissue glucocorticoid receptor transcriptional activity in a circadian fashion in man. PLoS ONE 2011, 6, e25612. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, J.D.; Dalbeth, N.; Mikuls, T.; Brignardello-Petersen, R.; Guyatt, G.; Abeles, A.M.; Gelber, A.C.; Harrold, L.R.; Khanna, D.; King, C.; et al. 2020 American College of Rheumatology Guideline for the Management of Gout. Arthritis Care Res. 2020, 72, 744–760. [Google Scholar] [CrossRef] [PubMed]
- Mittelstadt, P.R.; Ashwell, J.D. Inhibition of AP-1 by the Glucocorticoid-inducible Protein GILZ*. J. Biol. Chem. 2001, 276, 29603–29610. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Prefontaine, K.E. Physical association and functional antagonism between the p65 subunit of transcription factor NF-kappa B and the glucocorticoid receptor. Proc. Natl. Acad. Sci. USA 1994, 91, 752–756. [Google Scholar] [CrossRef]
- Galon, J.; Franchimont, D.; Hiroi, N.; Frey, G.; Boettner, A.; Ehrhart-Bornstein, M.; O’Shea, J.J.; Chrousos, G.P.; Bornstein, S.R. Gene profiling reveals unknown enhancing and suppressive actions of glucocorticoids on immune cells. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2002, 16, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Hermoso, M.A.; Matsuguchi, T.; Smoak, K.; Cidlowski, J.A. Glucocorticoids and tumor necrosis factor alpha cooperatively regulate toll-like receptor 2 gene expression. Mol. Cell. Biol. 2004, 24, 4743–4756. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.K.; Adya, R.; Lewandowski, K.C.; O’Hare, J.P.; Randeva, H.S. Diurnal variation and effect of insulin on circulating high molecular weight (HMW) adiponectin and NF-κB activity in human endothelial cells. Atherosclerosis 2011, 214, 174–177. [Google Scholar] [CrossRef]
- Richardson, V.M.; Santostefano, M.J.; Birnbaum, L.S. Daily Cycle of bHLH-PAS Proteins, Ah Receptor and Arnt, in Multiple Tissues of Female Sprague–Dawley Rats. Biochem. Biophys. Res. Commun. 1998, 252, 225–231. [Google Scholar] [CrossRef]
- Dhabhar, F.S. Stress-induced augmentation of immune function—The role of stress hormones, leukocyte trafficking, and cytokines. Brain. Behav. Immun. 2002, 16, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Harno, E.; Gali Ramamoorthy, T.; Coll, A.P.; White, A. POMC: The Physiological Power of Hormone Processing. Physiol. Rev. 2018, 98, 2381–2430. [Google Scholar] [CrossRef] [PubMed]
- Altmeyer, P.; Stöhr, L.; Holzmann, H. Seasonal Rhythm of the Plasma Level of Alpha-Melanocyte Stimulating Hormone. J. Investig. Dermatol. 1986, 86, 454–456. [Google Scholar] [CrossRef] [PubMed]
- Gianoulakis, C.; Dai, X.; Thavundayil, J.; Brown, T. Levels and circadian rhythmicity of plasma ACTH, cortisol, and β-endorphin as a function of family history of alcoholism. Psychopharmacology 2005, 181, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Waltervancauter, E.; Virasoro, E.; Leclercq, R.; Copinschi, G. Seasonal, Circadian and episodic variations of human immunoreactive beta-msh, acth and cortisol. Int. J. Pept. Protein Res. 1981, 17, 3–13. [Google Scholar]
- Hale, A.C.; Ratter, S.J.; Tomlin, S.J.; Lytras, N.; Besser, G.M.; Rees, L.H. Measurement of immunoreactive gamma-MSH in human plasma. Clin. Endocrinol. 1984, 21, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Moon, K.W.; Kim, H.O.; Lee, Y.A.; Hong, S.J.; Jung, J.Y.; Kim, H.A.; Suh, C.H.; Ha, Y.J.; Kim, I.J.; et al. Seasonal Variations and Associated Factors of Gout Attacks: A Prospective Multicenter Study in Korea. J. Korean Med. Sci. 2020, 35, e133. [Google Scholar] [CrossRef]
- Renquist, B.J.; Murphy, J.G.; Larson, E.A.; Olsen, D.; Klein, R.F.; Ellacott, K.L.J.; Cone, R.D. Melanocortin-3 receptor regulates the normal fasting response. Proc. Natl. Acad. Sci. USA 2012, 109, E1489–E1498. [Google Scholar] [CrossRef] [PubMed]
- Sutton, G.M.; Begriche, K.; Kumar, K.G.; Gimble, J.M.; Perez-Tilve, D.; Nogueiras, R.; McMillan, R.P.; Hulver, M.W.; Tschoep, M.H.; Butler, A.A. Central nervous system melanocortin-3 receptors are required for synchronizing metabolism during entrainment to restricted feeding during the light cycle. FASEB J. 2010, 24, 862–872. [Google Scholar] [CrossRef]
- Getting, S.J.; Lam, C.W.; Chen, A.S.; Grieco, P.; Perretti, M. Melanocortin 3 receptors control crystal-induced inflammation. FASEB J. 2006, 20, 2234–2241. [Google Scholar] [CrossRef]
- Getting, S.J.; Christian, H.C.; Flower, R.J.; Perretti, M. Activation of melanocortin type 3 receptor as a molecular mechanism for adrenocorticotropic hormone efficacy in gouty arthritis. Arthritis Rheum. 2002, 46, 2765–2775. [Google Scholar] [CrossRef]
- Wolfson, W.Q.; Cohn, C.; Levine, R. Rapid treatment of acute gouty arthritis by concurrent administration of pituitary adrenocorticotropic hormone (ACTH) and colchicine. J. Lab. Clin. Med. 1949, 34, 1766–1768. [Google Scholar]
- Spies, T.D.; Stone, R.E. Relief of the symptoms of acute gout and rheumatoid arthritis by means of pituitary adrenocorticotropic hormone. South. Med. J. 1949, 42, 720–722. [Google Scholar] [CrossRef] [PubMed]
- Margolis, H.M.; Caplan, P.S. Treatment of acute gouty arthritis with pituitary adrenocorticotropic hormone (ACTH). J. Am. Med. Assoc. 1950, 142, 256–258. [Google Scholar] [CrossRef] [PubMed]
- Daoussis, D.; Kordas, P.; Varelas, G.; Michalaki, M.; Onoufriou, A.; Mamali, I.; Iliopoulos, G.; Melissaropoulos, K.; Ntelis, K.; Velissaris, D.; et al. ACTH vs steroids for the treatment of acute gout in hospitalized patients: A randomized, open label, comparative study. Rheumatol. Int. 2022, 42, 949–958. [Google Scholar] [CrossRef]
- Capsoni, F.; Ongari, A.M.; Reali, E.; Catania, A. Melanocortin peptides inhibit urate crystal-induced activation of phagocytic cells. Arthritis Res. Ther. 2009, 11, R151. [Google Scholar] [CrossRef]
- Shishioh-Ikejima, N.; Ogawa, T.; Yamaguti, K.; Watanabe, Y.; Kuratsune, H.; Kiyama, H. The increase of alpha-melanocyte-stimulating hormone in the plasma of chronic fatigue syndrome patients. BMC Neurol. 2010, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.; Lee, G.; Lee, G.-S. Lower Temperatures Exacerbate NLRP3 Inflammasome Activation by Promoting Monosodium Urate Crystallization, Causing Gout. Cells 2021, 10, 1919. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, M.; Chan, X.Y.; Tan, S.Y.; Subramaniam, S.; Fan, Y.; Loh, E.; Chang, K.T.E.; Tan, T.C.; Chen, Q. Uncovering the mystery of opposite circadian rhythms between mouse and human leukocytes in humanized mice. Blood J. Am. Soc. Hematol. 2017, 130, 1995–2005. [Google Scholar] [CrossRef]
- Hayashi, M.; Shimba, S.; Tezuka, M. Characterization of the molecular clock in mouse peritoneal macrophages. Biol. Pharm. Bull. 2007, 30, 621–626. [Google Scholar] [CrossRef]
- Neogi, T.; Chen, C.; Chaisson, C.; Hunter, D.J.; Zhang, Y. Drinking water can reduce the risk of recurrent gout attacks. In Proceedings of the ACR Annual Scientific Meeting, Philadelphia, PA, USA, 16–21 October 2009; Volume 2009, pp. 16–21. [Google Scholar]
- Roddy, E.; Muller, S.; Hayward, R.; Mallen, C.D. The association of gout with sleep disorders: A cross-sectional study in primary care. BMC Musculoskelet. Disord. 2013, 14, 119. [Google Scholar] [CrossRef]
- Cobo, I.; Cheng, A.; Murillo-Saich, J.; Coras, R.; Torres, A.; Abe, Y.; Lana, A.J.; Schlachetzki, J.; Liu-Bryan, R.; Terkeltaub, R.; et al. Monosodium urate crystals regulate a unique JNK-dependent macrophage metabolic and inflammatory response. Cell Rep. 2022, 38, 110489. [Google Scholar] [CrossRef] [PubMed]
- Silver, A.C.; Buckley, S.M.; Hughes, M.E.; Hastings, A.K.; Nitabach, M.N.; Fikrig, E. Daily oscillations in expression and responsiveness of Toll-like receptors in splenic immune cells. Heliyon 2018, 4, e00579. [Google Scholar] [CrossRef]
- Wang, S.; Lin, Y.; Yuan, X.; Li, F.; Guo, L.; Wu, B. REV-ERBα integrates colon clock with experimental colitis through regulation of NF-κB/NLRP3 axis. Nat. Commun. 2018, 9, 4246. [Google Scholar] [CrossRef] [PubMed]
- Pourcet, B.; Zecchin, M.; Ferri, L.; Beauchamp, J.; Sitaula, S.; Billon, C.; Delhaye, S.; Vanhoutte, J.; Mayeuf-Louchart, A.; Thorel, Q.; et al. Nuclear Receptor Subfamily 1 Group D Member 1 Regulates Circadian Activity of NLRP3 Inflammasome to Reduce the Severity of Fulminant Hepatitis in Mice. Gastroenterology 2018, 154, 1449–1464.e20. [Google Scholar] [CrossRef]
- Billon, C.; Murray, M.H.; Avdagic, A.; Burris, T.P. RORγ regulates the NLRP3 inflammasome. J. Biol. Chem. 2019, 294, 10–19. [Google Scholar] [CrossRef]
- Spengler, M.L.; Kuropatwinski, K.K.; Comas, M.; Gasparian, A.V.; Fedtsova, N.; Gleiberman, A.S.; Gitlin, I.I.; Artemicheva, N.M.; Deluca, K.A.; Gudkov, A.V.; et al. Core circadian protein CLOCK is a positive regulator of NF-κB-mediated transcription. Proc. Nat. Acad. Sci. USA 2012, 109, E2457–E2465. [Google Scholar] [CrossRef]
- Hughey, J.J.; Butte, A.J. Differential Phasing between Circadian Clocks in the Brain and Peripheral Organs in Humans. J. Biol. Rhythm. 2016, 31, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Challet, E. Minireview: Entrainment of the Suprachiasmatic Clockwork in Diurnal and Nocturnal Mammals. Endocrinology 2007, 148, 5648–5655. [Google Scholar] [CrossRef]
- Snelling, S.J.B.; Forster, A.; Mukherjee, S.; Price, A.J.; Poulsen, R.C. The chondrocyte-intrinsic circadian clock is disrupted in human osteoarthritis. Chronobiol. Int. 2016, 33, 574–579. [Google Scholar] [CrossRef]
- Padmanabhan, K.; Billaud, M. Desynchronization of Circadian Clocks in Cancer: A Metabolic and Epigenetic Connection. Front. Endocrinol. 2017, 8, 278091. [Google Scholar] [CrossRef] [PubMed]
- Morton, A.J.; Wood, N.I.; Hastings, M.H.; Hurelbrink, C.; Barker, R.A.; Maywood, E.S. Disintegration of the sleep-wake cycle and circadian timing in Huntington’s disease. J. Neurosci. 2005, 25, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, J.E.; Ray, D.W. The role of the circadian clock in rheumatoid arthritis. Arthritis Res. Ther. 2013, 15, 205. [Google Scholar] [CrossRef]
- Kornhauser, J.M.; Nelson, D.E.; Mayo, K.E.; Takahashi, J.S. Regulation of jun-B Messenger RNA and AP-1 Activity by Light and a Circadian Clock. Science 1992, 255, 1581–1584. [Google Scholar] [CrossRef] [PubMed]
- Early, J.O.; Menon, D.; Wyse, C.A.; Cervantes-Silva, M.P.; Zaslona, Z.; Carroll, R.G.; Palsson-McDermott, E.M.; Angiari, S.; Ryan, D.G.; Corcoran, S.E.; et al. Circadian clock protein BMAL1 regulates IL-1β in macrophages via NRF2. Proc. Natl. Acad. Sci. USA 2018, 115, E8460–E8468. [Google Scholar] [CrossRef] [PubMed]
- Daigneault, M.; Preston, J.A.; Marriott, H.M.; Whyte, M.K.B.; Dockrell, D.H. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PLoS ONE 2010, 5, e8668. [Google Scholar] [CrossRef] [PubMed]
- De Juan, A.; Lavin Plaza, B. Isolation and Culturing of Primary Mouse and Human Macrophages BT—Atherosclerosis: Methods and Protocols; Ramji, D., Ed.; Springer: New York, NY, USA, 2022; pp. 113–124. ISBN 978-1-0716-1924-7. [Google Scholar]
- Nagoshi, E.; Saini, C.; Bauer, C.; Laroche, T.; Naef, F.; Schibler, U. Circadian Gene Expression in Individual Fibroblasts: Cell-Autonomous and Self-Sustained Oscillators Pass Time to Daughter Cells. Cell 2004, 119, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Balsalobre, A.; Damiola, F.; Schibler, U. A Serum Shock Induces Circadian Gene Expression in Mammalian Tissue Culture Cells. Cell 1998, 93, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Balsalobre, A.; Brown, S.A.; Marcacci, L.; Tronche, F.; Kellendonk, C.; Reichardt, H.M.; Schutz, G.; Schibler, U. Resetting of circadian time peripheral tissues by glucocorticoid signaling. Science 2000, 289, 2344–2347. [Google Scholar] [CrossRef]
- Yagita, K.; Okamura, H. Forskolin induces circadian gene expression of rPer1, rPer2 and dbp in mammalian rat-1 fibroblasts. FEBS Lett. 2000, 465, 79–82. [Google Scholar] [CrossRef]
- Du, X.; Shi, R.; Wang, Y.; Wu, W.; Sun, S.; Dai, Z.; Chen, C.; Weng, Z.; Li, X.; Liu, Q.; et al. Isoforskolin and forskolin attenuate lipopolysaccharide-induced inflammation through TLR4/MyD88/NF-κB cascades in human mononuclear leukocytes. Phytother. Res. 2019, 33, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N.; Khalili, H.; Konijeti, G.G.; Higuchi, L.M.; de Silva, P.; Fuchs, C.S.; Richter, J.M.; Schernhammer, E.S.; Chan, A.T. Sleep duration affects risk for ulcerative colitis: A prospective cohort study. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2014, 12, 1879–1886. [Google Scholar] [CrossRef] [PubMed]
- Swanson, G.R.; Kochman, N.; Amin, J.; Chouhan, V.; Yim, W.; Engen, P.A.; Shaikh, M.; Naqib, A.; Tran, L.; Voigt, R.M.; et al. Disrupted Circadian Rest-Activity Cycles in Inflammatory Bowel Disease Are Associated with Aggressive Disease Phenotype, Subclinical Inflammation, and Dysbiosis. Front. Med. 2022, 8, 770491. [Google Scholar] [CrossRef]
- Scheer, F.A.J.L.; Hilton, M.F.; Evoniuk, H.L.; Shiels, S.A.; Malhotra, A.; Sugarbaker, R.; Ayers, R.T.; Israel, E.; Massaro, A.F.; Shea, S.A. The endogenous circadian system worsens asthma at night independent of sleep and other daily behavioral or environmental cycles. Proc. Natl. Acad. Sci. USA 2021, 118, e2018486118. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Tyrrell, D.; Coyle, K.; Higgins, P.; Willman, J. Diurnal Variation in the Symptoms of Colds and Influenza. Chronobiol. Int. 1988, 5, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Orange, D.E.; Blachere, N.E.; DiCarlo, E.F.; Mirza, S.; Pannellini, T.; Jiang, C.S.; Frank, M.O.; Parveen, S.; Figgie, M.P.; Gravallese, E.M.; et al. Rheumatoid Arthritis Morning Stiffness Is Associated with Synovial Fibrin and Neutrophils. Arthritis Rheumatol. 2020, 72, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Perry, M.G.; Kirwan, J.R.; Jessop, D.S.; Hunt, L.P. Overnight variations in cortisol, interleukin 6, tumour necrosis factor alpha and other cytokines in people with rheumatoid arthritis. Ann. Rheum. Dis. 2009, 68, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Malibary, N.H.; Ezzat, M.A.; Mogharbel, A.M.; Kouzaba, K.A.; Alkadi, A.A.; Malki, U.H.; Gharib, S.M.; Altowairqi, F.M.; Saadah, O.I.; Mosli, M.H. Factors Affecting Ulcerative Colitis Flare-Ups: Associations with Smoking Habits and Other Patient Characteristics. Cureus 2021, 13, e19834. [Google Scholar] [CrossRef]
- De Silva, M.; Binder, A.; Hazleman, B.L. The timing of prednisolone dosage and its effect on morning stiffness in rheumatoid arthritis. Ann. Rheum. Dis. 1984, 43, 790–793. [Google Scholar] [CrossRef]
- Arvidson, N.G.; Gudbjörnsson, B.; Larsson, A.; Hällgren, R. The timing of glucocorticoid administration in rheumatoid arthritis. Ann. Rheum. Dis. 1997, 56, 27–31. [Google Scholar] [CrossRef]
- Nichols, T.; Nugent, C.A.; Tyler, F.H. Diurnal variation in suppression of adrenal function by glucocorticoids. J. Clin. Endocrinol. Metab. 1965, 25, 343–349. [Google Scholar] [CrossRef]
- Misawa, T.; Takahama, M.; Kozaki, T.; Lee, H.; Zou, J.; Saitoh, T.; Akira, S. Microtubule-driven spatial arrangement of mitochondria promotes activation of the NLRP3 inflammasome. Nat. Immunol. 2013, 14, 454–460. [Google Scholar] [CrossRef]
- Niel, E.; Scherrmann, J.-M. Colchicine today. Jt. Bone Spine 2006, 73, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Froy, O. Cytochrome P450 and the biological clock in mammals. Curr. Drug Metab. 2009, 10, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-D.; Yeh, J.-K.; Peng, M.-T.; Shie, S.-S.; Lin, S.-L.; Yang, C.-H.; Chen, T.-H.; Hung, K.-C.; Wang, C.-C.; Hsieh, I.-C.; et al. Circadian CLOCK Mediates Activation of Transforming Growth Factor-β Signaling and Renal Fibrosis through Cyclooxygenase 2. Am. J. Pathol. 2015, 185, 3152–3163. [Google Scholar] [CrossRef]
- Kato, K.; Murai, I.; Asai, S.; Takahashi, Y.; Nagata, T.; Komuro, S.; Mizuno, S.; Iwasaki, A.; Ishikawa, K.; Arakawa, Y. Circadian rhythm of melatonin and prostaglandin in modulation of stress-induced gastric mucosal lesions in rats. Aliment. Pharmacol. Ther. 2002, 16 (Suppl. S2), 29–34. [Google Scholar] [CrossRef]
- Al-Waeli, H.; Nicolau, B.; Stone, L.; Abu Nada, L.; Gao, Q.; Abdallah, M.N.; Abdulkader, E.; Suzuki, M.; Mansour, A.; Al Subaie, A.; et al. Chronotherapy of Non-Steroidal Anti-Inflammatory Drugs May Enhance Postoperative Recovery. Sci. Rep. 2020, 10, 468. [Google Scholar] [CrossRef]
- Tamimi, Z.; Abusamak, M.; Al-Waeli, H.; Al-Tamimi, M.; Al Habashneh, R.; Ghanim, M.; Al-Nusair, M.; Gao, Q.; Nicolau, B.; Tamimi, F. NSAID chronotherapy after impacted third molar extraction: A randomized controlled trial. Oral. Maxillofac. Surg. 2022, 26, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Yang, N.; Borysiewicz, E.; Dudek, M.; Williams, J.L.; Li, J.; Maywood, E.S.; Adamson, A.; Hastings, M.H.; Bateman, J.F.; et al. Catabolic cytokines disrupt the circadian clock and the expression of clock-controlled genes in cartilage via an NFkB-dependent pathway. Osteoarthr. Cartil. 2015, 23, 1981–1988. [Google Scholar] [CrossRef]
- Xu, H.; Huang, L.; Zhao, J.; Chen, S.; Liu, J.; Li, G. The circadian clock and inflammation: A new insight. Clin. Chim. Acta 2021, 512, 12–17. [Google Scholar] [CrossRef]
- Cermakian, N.; Westfall, S.; Kiessling, S. Circadian Clocks and Inflammation: Reciprocal Regulation and Shared Mediators. Arch. Immunol. Ther. Exp. 2014, 62, 303–318. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Mice | Effect on Inflammation/NLRP3 Inflammasome | Day/Night Association | Possible Time-of-Day Effect on NLRP3 Inflammasome Activity | |
|---|---|---|---|---|
| Day | Night | |||
| Circadian rhythms in hormonal regulators of inflammation | At pharmacological concentrations, all are anti-inflammatory. Glucocorticoids can promote inflammasome priming | Higher circulating levels of glucocorticoids, melatonin and melanocortins at night | Higher inflammatory cytokine production from active inflammasome * | Reduced inflammatory cytokine production from active inflammasome * Increased susceptibility to inflammasome activation |
| Time-of-day differences in expression of inflammasome components and responsiveness to TLR2 and TLR4 agonists | Higher basal levels of inflammasome components may reduce threshold for inflammasome priming | High expression of TLRs, NLRP3 and RelA and higher NF-κB activity at night | Increased susceptibility to inflammasome activation | |
| Humans | Effect on Inflammation/NLRP3 Inflammasome | Day/Night Association | Possible Time-of-Day Effect on NLRP3 Inflammasome Activity | |
|---|---|---|---|---|
| Day | Night | |||
| Circadian rhythms in hormonal regulators of inflammation | At pharmacological concentrations, all are anti-inflammatory. Glucocorticoids can promote inflammasome priming | Glucocorticoids and melanocortins high in morning; melatonin high at night | Reduced inflammatory cytokine production from active inflammasomes throughout the day and night * | |
| Increased susceptibility to inflammasome-activating stimuli * | ||||
| Time-of-day differences in responsiveness of macrophages to MSU crystals | Levels of caspase 1 activity and IL-1β production depend on time-of-day of MSU crystal exposure | Higher inflammatory response to MSU crystals at night? | Greater extent of inflammation generated by MSU crystal-induced NLRP3 inflammasome activation | |
| Disruption of the circadian clock in macrophages by MSU crystal exposure | Loss of circadian repression of NLRP3 expression and NLRP3 inflammasome activation due to reduced levels of REV-ERBα and BMAL1 | Greater repression of REV-ERBα at night? BMAL1 naturally more abundant during the day? | Increased susceptibility to inflammasome-activating stimuli Greater extent of inflammation generated by NLRP3 inflammasome | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poulsen, R.C.; Dalbeth, N. Circadian Rhythms in NLRP3 Inflammasome Regulation: Possible Implications for the Nighttime Risk of Gout Flares. Gout Urate Cryst. Depos. Dis. 2024, 2, 108-132. https://doi.org/10.3390/gucdd2020011
Poulsen RC, Dalbeth N. Circadian Rhythms in NLRP3 Inflammasome Regulation: Possible Implications for the Nighttime Risk of Gout Flares. Gout, Urate, and Crystal Deposition Disease. 2024; 2(2):108-132. https://doi.org/10.3390/gucdd2020011
Chicago/Turabian StylePoulsen, Raewyn C., and Nicola Dalbeth. 2024. "Circadian Rhythms in NLRP3 Inflammasome Regulation: Possible Implications for the Nighttime Risk of Gout Flares" Gout, Urate, and Crystal Deposition Disease 2, no. 2: 108-132. https://doi.org/10.3390/gucdd2020011