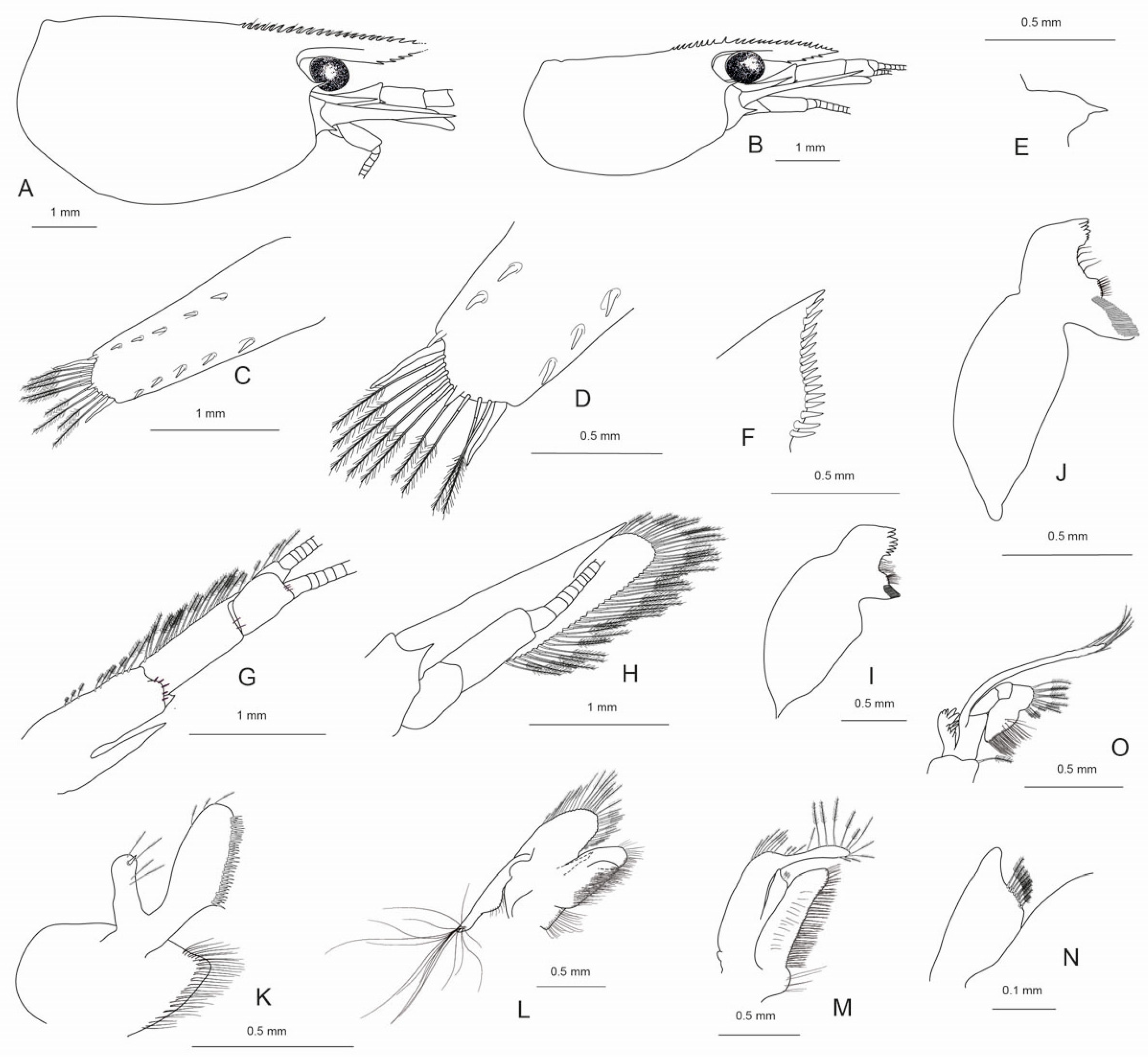
3.3. Description
Cephalothorax and cephalic appendages: Small shrimp with carapace lengths of 1.8–3.7 mm (median 2.3 mm, n = 7) in apparently adult specimens. Carapace (
Figure 1A,B) smooth, antennal spine clearly separated from suborbital lobe, pterygostomial angle broadly rounded. Rostrum (
Figure 1A,B) extending from midlength of second article of antennular peduncle to distal end of it, 0.70–0.85 (median 0.71, n = 5) times as long as carapace, nearly straight or slightly sinuous, terminating in acute tip; dorsal margin armed with 15–21 basally articulated teeth over entire length, including 4–5 postorbital, teeth above orbit larger than anterior and proximal teeth; ventral margin with 2–4 moderately large teeth located in distal half of the rostrum. Rostrum formula based on six specimens 4−5(4)+11−17/2−4. Eyes (
Figure 1A,B) well developed with pigmented globular cornea. Antennular peduncle (
Figure 1A,B,G), 0.86–0.93 (median 0.92, n = 4) times as long as carapace in females, 1.0 times as long as carapace in males, first segment 1.69–1.92 (median 1.81, n = 5) times as long as second segment, second segment 1.86–2.67 (median 1.88, n = 5) times longer as third segment; distolateral tooth on first segment well developed, acute, 0.15–0.38 (median 0.20, n = 5) times as longs as second segments; second and third segments unarmed, with few spiniform setae near distal margin. Stylocerite acuminate, reaching to 0.72–0.93 (median 0.80, n = 5) times the length of the basal segment of antennular peduncle. Antennal scaphocerite (
Figure 1H) with well-developed distolateral tooth, 3.86–4.0 (median 3.80, n = 3) times as long as wide.
Pleon, telson and uropods: The sixth pleomere 0.60–0.67 (median 0.63, n = 4) times the carapace length, 1.63–1.83 (median 1.72, n = 4) times as long as the fifth pleomere, and 0.87–1.0 (median 0.92, n = 4) times as long as the telson. Telson (
Figure 1C,D) slightly tapering distally, distal margin strongly convex to triangular, lacking median apex, with 3–4 (n = 3) pairs of dorsal short spiniform setae and one pair of short spiniform setae dorsolaterally; distal margin with 6–8 (n = 5) strong spiniform setae, where the lateral pair was the longest. Preanal carina (
Figure 1E) rounded, low. Uropodal diaeresis (
Figure 1F) with 10–13 movable spiniform setae, outermost one shorter than lateral angle.
Mouthparts and branchiae: Mandibles (
Figure 1I) without palp, incisor process ending in irregular teeth, molar process truncated. Lower endite of maxillula (
Figure 1J) broadly rounded with long pappose setae and few rows of serrate setae near margin and scattered cuspidate setae proximal, upper endite elongate, with numerous distinct cuspidate setae flanked by a row of serrate setae on inner margin and few pappose setae subterminal, palp slender with few simple setae and one conical spiniform seta near tip. Upper endites of maxilla (
Figure 1K) subdivided, furnished with numerous pappose, simple and papposerrate setae arranged in rows, palp slender, scaphognathite tapering posteriorly, fringed with long, curved serrulate setae on truncate posterior margin and plumose setae inferiorly. Palp of first maxilliped (
Figure 1L,M) terminating in finger-like extension, caridean lobe and flagellum with plumose setae, base with numerous pappose, papposerrate and serrate setae arranged in rows and tufts. Podobranch on second maxilliped (
Figure 1N) well developed; exopod slender, protruding propodal segment, with long plumose setae distally; basis and merus with rows of pappose setae, distolateral margin of penultimate segment with numerous pappose and serrate setae; dactylus with numerous serrate setae flanked by a row of pappose setae. Third maxilliped (
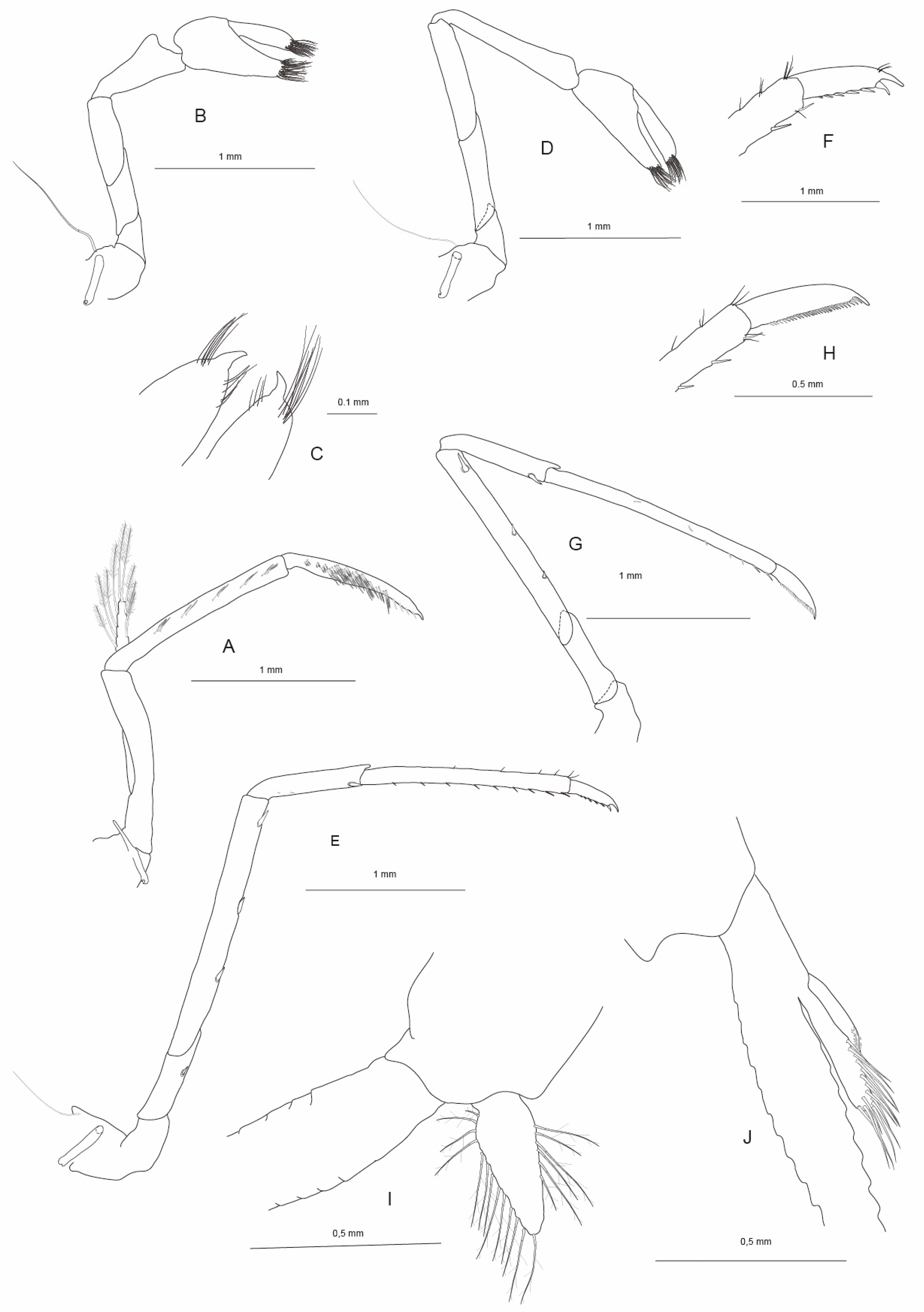
Figure 2A) slender, with two arthrobranchs; exopod slender, exceeding distal margin of antepenultimate segment, with few long plumose setae distally; antepenultimate segment of endopod slender, with few simple setae basally; penultimate segment slender, with straight lateral margins, with few simple setae distally, inner margin with rows of short serrate setae; ultimate segment slightly shorter than penultimate segment, tapering distally, ending in a claw-like curved spiniform seta (nail), proximal to nail with few cuspidate setae; inner margin densely covered with rows of serrate setae; epipod well developed, with distal hook. Branchial apparatus as shown in
Table 2; well-developed (with distal hooks) epipods present on third maxilliped and first pereiopod (
Figure 2B). Vestigial epipods (visible only after cleaning and staining of appendages) on second and third pereiopods.
Pereiopods: Chela of the first pereiopod (
Figure 2C) was caridinoid in shape, slender, with well-developed palm, 2.44–2.90 (n = 2) times as long as wide, 1.30–1.32 (n = 2) times as long as carpus; fingertips rounded, without hooks but each with tuft of setae; dactylus 3.67–4.00 (n = 2) times as long as wide, 1.22–1.44 (n = 2) times as long as palm. Carpus slender, hardly excavated distally, 2.73–2.75 (n = 2) times as long as wide, 1.29–1.36 (n = 2) times as long as merus. Merus not inflated, 2.44–2.62 (n = 2) times as long as wide, shorter than ischium. Chela of second pereiopod (
Figure 2D) very slender, caridinoid in shape, with well-developed palm, 3.65–3.75 (n = 2) times as long as wide, 0.73–0.84 (n = 2) times as long as carpus; fingertips rounded, without hooks but each with duft of seta, dactylus 6.17–6.67 (n = 2) times as long as wide, 1.42–1.67 (n = 2) times as long as palm. Carpus slender, 6.17–6.63 (n = 2) times as long as wide, 1.64–1.67 (n = 2) times as long as merus. Merus not inflated, 3.82–4.55 (n = 2) times as long as wide, shorter than ischium. Third pereiopod (
Figure 2E,F) with dactylus (
Figure 2F) 5.00–5.33 (n = 2) times as long as wide (including terminal claw and spiniform setae on flexor margin), terminating in one claw, with five or six accessory spiniform setae on flexor margin; propodus not dilated or distally curved, 12.0 (n = 2) times as long as wide, 3.0 (n = 2) times as long as dactylus; carpus 4.33–4.71 (n = 2) times as long as wide, 0.54–0.55 (n = 2) times as long as propodus, 0.45–0.49 (n = 2) times as long as merus, with one large spiniform seta near distal margin and three small spiniform setae proximal; merus 7.07–9.73 (n = 2) times as long as wide, 2.04–2.21 (n = 2) times as long as carpus, with three or four strong movable spiniform setae on externo-inferior margin; ischium with one spiniform seta. Fourth pereiopod was similar to third but slightly smaller. Fifth pereiopod (
Figure 2G,H) slender; dactylus (
Figure 2H) 4.0–5.50 (n = 2) times as long as wide (including terminal claw and spiniform setae on flexor margin), terminating in one large claw, with 21–49 spiniform setae on flexor margin; propodus with small spiniform setae on ventral margin, distal pair of spiniform setae not enlarged, propodus 12.50–16.40 (n = 2) times as long as wide, 2.27–4.10 (n = 2) times as long as dactylus; carpus 5.0–7.83 (n = 2) times as long as wide, 0.50–0.57 (n = 2) times as long as propodus, 0.61–0.67 (n = 2) times as long as merus, with one strong spiniform seta near distal margin and 2–5 small spiniform setae proximal; merus 9.11–9.33 (n = 2) times as long as wide, 1.49–1.64 (n = 2) times as long as carpus, bearing three strong appressed movable spiniform setae on externo-inferior margin. Ischium with or without one spiniform seta.
Pleopods: Endopod of male first pleopod (
Figure 2I) elongated subtriangular, lacking appendix interna, 3.2 times as long as proximal width, 0.38 times as long as exopod, with long plumose setae on outer and distal margins, with few pappose setae on inner margin. Male second pleopod with appendix masculina rod-shaped (
Figure 2J), 8.3 (n = 1) times as long as wide, 0.63 (n = 1) times as long as endopod, armed with strong spiniform setae on inner margin and distal margin; appendix interna originating from about 0.4 times the length of appendix masculina, reaching to about 0.75 times the length of appendix masculina.
Reproductive biology: Ovigerous females with medium-sized embryos; size of undeveloped embryos (without eyes) 0.59–0.64 × 0.37–0.39 mm.
Size: Postorbital carapace length of apparently adult specimens 1.8–3.7 mm.
Colouration: Transparent to brownish with cream-colored transversal stripes on the pleon (Wilson 2008; Short, Page and Humphrey 2019).
Distribution: The species occurs in rivers around Darwin, Northern Territory, Australia.
Etymology: The species name darwin is derived from the Darwin river catchment, the natural habitat of the proposed new species, and is used as a noun in apposition.
Common name. Blackmore River Caridina.
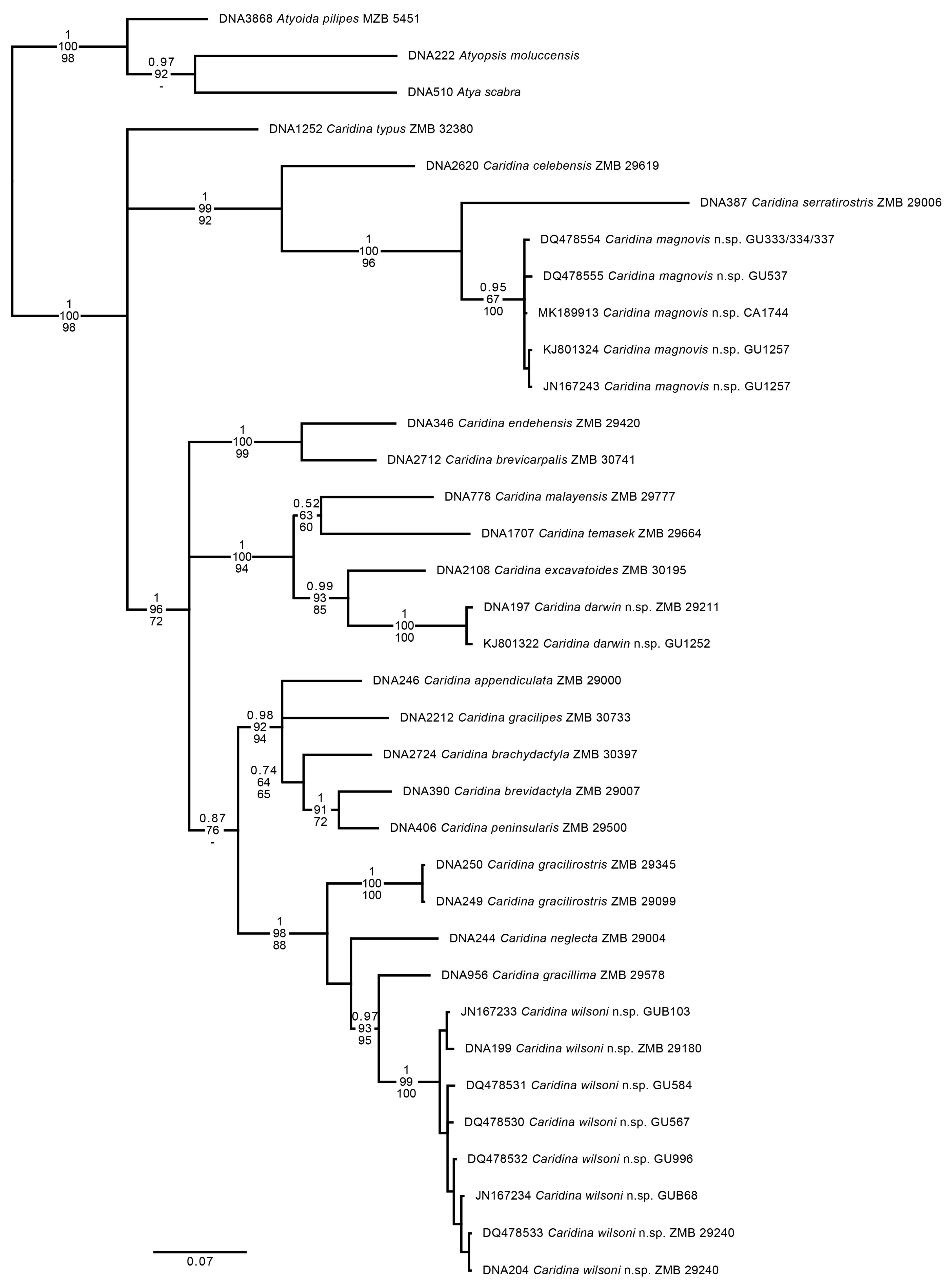
Remarks: Based on morphology,
Caridina darwin n. sp. is related to a group of species that includes
Caridina propinqua de Man, 1908 and
Caridina laevis Heller, 1862 as the best known members. Within this group of species related to
C. laevis, two species with large embryos—
Caridina temasek Choy and Ng, 1991 and
Caridina excavatoides Johnson, 1961, both occurring in freshwaters of the Malayan peninsula—are most closely related to the proposed new species.
Caridina darwin n. sp. differs from
C. temasek by a higher number of teeth on the dorsal margin of the rostrum (15–21 vs. 14–16 in
C. temasek), the development of epipods (vestigial epipods (only visible after clearing and staining of the appendages) on the second and third pereiopods, the lack of an epipod on the fourth pereiopod vs. a well-developed (with distal hooks) epipod on the second pereiopod, a reduced epipod on the third and a vestigial epipod on the fourth pereiopod in
C. temasek). Furthermore, the proposed new species differs from
C. temasek by lacking an appendix interna on the first pleopod of males (vs. with well-developed appendix interna in
C. temasek) and smaller embryo size (0.59–0.64 × 0.37–0.39 mm vs. 0.70–0.80 × 0.44–0.54 mm in
C. temasek according to the original description). There is still some confusion about the shape of the preanal carina in
C. temasek. Choy and Ng [
23] did not give a drawing of this part, but in the verbal description, they wrote “Pre-anal carina with about 10 setae”, suggesting that they did not observe a distinct hook-like spine. In their paper on Atyidae from peninsular Malaysia and Singapore [
24], Cai et al. wrote that the preanal carina in
C. temasek is “triangular, no spine or with a very tiny spine”. Four of five paratypes of
C. temasek observed for the present study show a prominent hook-like spine on the preanal carina, as do all four topotypical specimens from Singapore listed above. The holotype of
C. temasek lacks a spine on the preanal carina (Y. Cai, pers. comm.). It seems likely that specimens assigned to
C. temasek with and without a hook-like spine on the preanal carina might belong to different species. All specimens of
Caridina darwin n. sp. lack a spine on the preanal carina.
Caridina darwin n. sp. is also similar to Caridina excavatoides Johnson, 1961. Both species lack an appendix interna on the endopod of the first pleopod in males. The proposed new species differs from C. excavatoides by a longer sixth pleomere (0.60–0.67 times the length of the carapace vs. less than 0.6 times the length of the carapace in C. excavatoides). Furthermore, Caridina darwin n. sp. differs from this species in the shape of the palp of the first maxilliped (with a distinct finger-like extension vs. ending in a triangular extension in C. excavatoides). Caridina darwin n. sp. differs from C. excavatoides in the development of the epipods on the third maxilliped and pereiopods in the same way as C. temasek. The dactylus of the fifth pereiopod has a smaller number of spiniform setae on the flexor margin (21–49 vs. 50–60 in C. excavatoides).
Caridina magnovis n. sp.
Caridina sp. WA 4 Page, von Rintelen and Hughes 2007;
Caridina sp. ‘WA 4’ Wilson, 2008;
Caridina sp. WA 4 Cook, Page and Hughes 2011;
Caridina ‘sp. WA4’ Short, Page and Humphrey, 2019;
3.4. Material Examined
Holotype: ♂ cl 2.9 mm, Australia, Western Australia, Fitzroy River catchment, Calder Yard, FIT12-6, leg. M. Scanlon 29.04.1998, WAM C81392.
Paratypes: One ♂ cl 2.6 mm, one ♀ cl 3.7 mm, same data as holotype WAM C81393; one ♂ cl 2.7 mm, Australia, Western Australia, Ord River catchment, Mantinea Flats, leg. M. Scanlon 18.09.1998, WAM C81394; two ov. ♀♀ cl 4.0 and 4.3 mm, one ♀ cl 3.7 mm, Darwin Area, Howard River, Gunn Pt. Rd Xing, cultured specimens provided by D. Wilson NTM Cr019573; three ♀♀ cl 1.8–2.2 mm, two ♂♂ cl 2.3 and 2.4 mm, Fitzroy River catchment, Dimond Gorge, leg. M. Scanlon 14.04.1998 ZMB 33101; one ♂ cl 3.0 mm, Darwin Area, Blackmore River, cultured specimens provided by D. Wilson ZMB 33112; one ♀ cl 2.1 mm, one ov. ♀ cl. 3.1 mm, four juv. cl 1.6–1.8 mm, Australia, Northern Territory, Top End, Coomalie Creek, S 13°0.884′, E 131°7.371′, 85-04, leg. N. Brinkmann und T. von Rintelen 28.06.2004, ZMB 29152; two ♀♀ cl 2.5 und 2.7 mm, two ov. ♀♀ cl 2.8 und 2.9 mm, Australia, Northern Territory, Howard Springs, S 12°27.345′ E 131°3.146′, 79-04, leg. M. Glaubrecht, N. Brinkmann, T. von Rintelen 27.06.2004, ZMB 32120.
3.6. Description
Cephalothorax and cephalic appendages: Small shrimp with carapace lengths of 2.1–4.3 mm (median 2.9 mm, n = 16) in apparently adult specimens. Carapace (
Figure 3A,B) smooth, antennal spine almost fused with suborbital lobe, pterygostomial angle rounded. Rostrum (
Figure 3A,B) extending to end of second segment of antennular peduncle or to distal end of antennular peduncle, 0.53–0.76 (median 0.64, n = 8) times as long as carapace, nearly straight or slightly convex, ending in acute tip; dorsal margin armed with 17–29 teeth over entire length, including 5–8 postorbital; teeth above orbit larger than anterior and posterior teeth, ventral margin with 3–6 teeth located in distal half of rostrum. Rostrum formula based on 12 specimens 5 − 8 + 12 − 24/3 − 6. Eyes (
Figure 3A,B) well developed with pigmented globular cornea. Antennular peduncle (
Figure 3A,B,G) 0.60–0.71 (median 0.67, n = 9) times as long as carapace in females, 0.91 times as long as carapace in males, first segment 1.69–2.10 (median 2.0, n = 9) times as long as second segment, second segment 2.20–2.75 (median 2.31, n = 8) times as long as third segment; distolateral tooth on first segment short (0.14–0.23 times as long as second segment), well developed; second and third segments unarmed, with few spiniform setae near distal margin. Stylocerite acuminate, reaching to 0.89–1.05 (median 1.00, n = 9) times the length of basal segment of antennular peduncle. Antennal scaphocerite (
Figure 3H) with well-developed distolateral tooth, 2.92–3.7 (median 3.14, n = 5) times as long as wide.
Pleon, telson and uropods: Sixth pleomere was 0.44–0.54 (median 0.46, n = 7) times the carapace length, 1.23–1.69 (median 1.55, n = 6) times as long as fifth somite, 0.80–0.89 (median 0.87, n = 5) times as long as telson. Telson (
Figure 3C,D) slightly tapering distally, distal margin convex, with a prominent median point, with 4–5 (n = 7) pairs of dorsal short spiniform setae and one pair of short spiniform setae dorsolaterally; distal margin with one pair of strong spiniform setae and 7–9 plumose setae clearly exceeding lateral pair of spiniform setae. Preanal carina (
Figure 3E) rounded, high, with a small tooth in 2 of 7 specimens. Uropodal diaeresis (
Figure 3F) with 12–15 movable spiniform setae, the outermost shorter than the lateral angle.
Mouthparts and branchiae: Shown in
Figure 3I–O and
Figure 4A. Similar to
Caridina darwin n. sp. except for the incisor process of mandible (
Figure 3I,J), comparably well-developed and numerous irregular teeth and palp of first maxilliped (
Figure 3M,N) terminates in a triangular extension. Third maxilliped (
Figure 4A) was slender, with the last segment of maxilliped slightly shorter than penultimate segment. Branchial apparatus as shown in
Table 3, with one well developed and one reduced arthrobranch on third maxilliped and one arthrobranch on first pereiopod.
Pereiopods: Chela of first pereiopod (
Figure 4B) was caridinoid in shape, slender, with well-developed palm, 1.75–2.84 (median 2.21, n = 5) times as long as wide, 0.93–1.17 (median 1.12, n = 5) times as long as carpus; fingertips rounded, without hooks but each with a tuft of setae; dactylus 2.73–4.25 (median 3.57, n = 5) times as long as wide, 1.00–1.64 (median 1.25, n = 5) times as long as palm. Carpus slender, hardly excavated distally, 2.12–3.22 (median 2.78, n = 5) times as long as wide, 1.09–1.25 (median 1.17, n = 5) times as long as merus. Merus not inflated, 2.75–4.33 (median 3.00, n = 0) times as long as wide, 0.95–1.18 (median 1.0, n = 5) times as long as ischium. Chela of second pereiopod (
Figure 4C) very slender, caridinoid in shape, with well-developed palm, 2.75–4.40 (median 3.30, n = 5) times as long as wide, 0.66–0.71 (median 0.69, n = 5) times as long as carpus; fingertips rounded, without hooks; dactylus 4.44–5.45 (median 5.00, n = 5) times as long as wide, 1.25–1.36 (median 1.33, n = 5) times as long as palm, with tufts of setae distally; carpus slender, 6.36–8.17 (median 7.14, n = 5) times as long as wide, 1.04–1.40 (median 1.35, n = 5) times as long as merus. Merus not inflated, 5.83–7.83 (median 6.24, n = 5) times as long as wide, 1.06–1.17 (median1.13, n = 4) times as long as ischium. Third pereiopod (
Figure 4D–G) with dactylus (
Figure 4E–G) slightly sexually dimorphic, spiniform setae on flexor margin more curved, 3.78 times as long as wide (including terminal claw and spiniform setae on flexor margin) in males (n = 1), 4.00–5.00 (median 4.56, n = 4) times as long as wide in females, terminating in one claw, with 6–8 accessory spiniform setae on flexor margin, first spiniform seta on flexor margin strong and curved in some specimens; propodus not enlarged or curved distally, distal spiniform seta not enlarged, 10.6–14.29 (median 12.50, n = 5) times as long as wide, 3.24–3.73 (median 3.33, n = 5) times as long as dactylus; carpus 5.67–6.80 (median 5.86, n = 4) times as long as wide, 0.63–0.70 (median 0.66, n = 4) times as long as propodus, 0.50–0.59 (median 0.57, n = 4) times as long as merus, with one large spiniform seta near distal margin and 3–6 small spiniform setae proximally; merus 6.84–9.08 (median 8.47, n = 4) times as long as wide, 1.69–2.00 (median 1.74, n =4) times as long as carpus, with 3–5 strong movable spiniform setae on externo-inferior margin; ischium without or with one spiniform seta. Fourth pereiopod similar to third but slightly smaller. Fifth pereiopod (
Figure 4H,I) was slender; dactylus (
Figure 4I) 3.6–5.00 (n = 2) times as long as wide (including terminal claw and spiniform setae on flexor margin), terminating in one large claw, with 16–18 spiniform setae on flexor margin; propodus with small spiniform setae on ventral margin, distal pair of spiniform setae not enlarged, propodus 12.40–13.80 (median 12.50, n = 3) times as long as wide, 3.44–3.75 (n = 2) times as long as dactylus; carpus 5.83–7.09 (median 6.62, n = 3) times as long as wide, 0.56–0.57 (median 0.57, n = 3) times as long as propodus, 0.66–0.68 (median 0.67, n = 3) times as long as merus, with one strong spiniform seta near distal margin and five small spiniform setae proximally; merus 7.43–8.43 (median 8.40, n = 3) times as long as wide, 1.47–1.51 (median 1.49, n = 3) times as long as carpus, with 2–4 strong appressed movable spiniform setae on externo-inferior margin. Ischium with one or without spiniform seta.
Pleopods: Endopod of male first pleopod (
Figure 4J) elongated triangular, lacking appendix interna, 2.0 (n = 1) times as long as proximal width, 0.40 times as long as exopod, with long plumose setae on distal margin, and few shorter plumose setae on inner and outer margins. Male second pleopod with appendix masculina rod-shaped (
Figure 4K), 10.0 (n = 1) times as long as wide, 0.83 (n = 1) times as long as endopod, armed with strong spiniform setae on inner margin and distal margin; appendix interna originating from about 0.4 times the length of appendix masculina, reaching to about 0.65 times the length of appendix masculina.
Reproductive biology: Ovigerous females with medium-sized embryos; embryo size 0.66–0.76 × 0.39–0.49 mm (n = 6). Twenty-five and thirty embryos counted in two ovigerous females.
Size: Postorbital carapace length of apparently adult specimens 2.1–4.3 mm.
Colouration: Variable. Some specimens reddish with a pale median dorsal line extending from the rostrum to the sixth pleomere, some specimens mottled with reddish and creamy chromatophores, smaller specimens mostly transparent [
4,
25].
Distribution: The species is known from rivers draining into the Timor Sea. From the Fitzroy River in Western Australia eastwards to the Howard River in the Northern Territory.
Etymology: The species name magnovis is derived from the rather large size of the embryos of the proposed new species compared to C. serratirostris.
Common name: North Australian Chameleon Shrimp.
Remarks: The general morphology of Caridina magnovis n. sp., with a straight rostrum, armed throughout close to the tip and a high number of postorbital teeth (5–8), a long stylocerite, slender chelipeds and distal margin of the telson armed with long plumose setae clearly exceeding a lateral pair of strong spiniform setae and the variable coloration of living specimens resembles that of Caridina serratirostris de Man, 1892. The proposed new species differs from C. serratirostris in the armature of the rostrum, i.e., ventral teeth located in the distal half of the rostrum vs. ventral teeth placed at midlength of the rostrum in C. serratirostris. The stylocerite of Caridina magnovis n. sp. is shorter, reaching to 0.89–1.05 of the first segment of antennular peduncle vs. clearly exceeding this segment (1.09–1.47 times the length of the first segment). Carpus of first pereiopod is 2.12–3.22 times as long as wide in Caridina magnovis n. sp. vs. 2.63–4.67 times in C. serratirostris. Dactylus of third pereiopod with 6–8 spiniform setae vs. with 4–5 in C. serratirostris. Dactylus of fifth pereiopod with 16–18 spiniform setae on flexor margin, first seta not markedly enlarged vs. with 11–17 spiniform setae, first seta enlarged in C. serratirostris. Uropodal diaeresis with 12–15 movable spiniform setae vs. with 16–21 movable spiniform setae in C. serratirostris. Preanal carina rounded, without prominent tooth in most specimens (vs. preanal carina with a prominent hook-like tooth in all specimens of C. serratirostris). Embryos 0.66–0.76 × 0.39–0.49 mm vs. 0.37–0.44 × 0.20–0.26 mm in C. serratirostris.
Caridina wilsoni n. sp.
Caridina sp. Gulf 1 Page, von Rintelen and Hughes, 2007;
Caridina sp. ‘Gulf 1’ Wilson, 2008;
Caridina sp. Gulf 1 Cook, Page and Hughes, 2011;
Caridina ‘sp. Gulf1’ Short, Page and Humphrey, 2019;
3.7. Material Examined
Holotype.:_ ♂ cl 3.4 mm, Australia, Queensland, Archer River catchment, Archer River at telegraph line, FNARH5/216, leg. DERM, 10.04.2000, QM W29816.
Paratypes: One ov. ♀ cl 4.3 mm, same data as holotype, QM W29817; ♂ cl 3.0 mm, Australia, Queensland, Wenlock River catchment, Wenlock River at Stones Xing, leg. DERM, 10.04.2000, QM W29818; one ♀ cl 3.4 mm, commercial production ponds (most likely Blackmore R., could be Darwin R.), Darwin Area, cultured specimens provided by D. Wilson NTM Cr019572; seven ov. ♀♀ cl 3.2–4.0 mm, three ♂♂ cl 2.4–2.5 mm, Australia, Northern Territory, Top End, Darvin River crossing; S 12°49.089′, E 130°57.634′, 83-04, coll. N. Brinkmann and T. von Rintelen 27.6.2004, ZMB 29180; two ♂♂ cl 1.9 and 2.2 mm, four ♀♀ cl 2.3–2.7 mm, Australia, Northern Territory, Top End, Coomalie Creek, S 13°0.884′, E 131°7.371′; coll. N. Brinkmann and T. von Rintelen 28.06.2003, ZMB 29240.
3.8. Comparative Material Examined
Syntypes. Caridina gracilirostris De Man, 1892. Two ov. ♀♀ cl 5.2 and 5.3 mm, Indonesia, Celebes (=Sulawesi): river near Balangnipa 1888–1889, leg. M. Weber: RMNH.CRUS.D.2557; one ov. ♀ cl 5.3 mm, two ♂♂ cl 4.0 and 4.5 mm, Indonesia, Celebes (=Sulawesi), river near Balangnipa 1888–1889, leg. M. Weber, RMNH.CRUS.D.1076.
Other material. Caridina gracillima Lanchester, 1901. Two ♀♀ cl 5.2 and 5.3 mm, Malaysia, Peninsula, Terengganu, Kg. Dawi, Lake Kenyir, 05°08.232′ N, 102°48.211′ E (loc 56-10); leg. B. Stelbrink, 02. 06. 2010, ZMB 29578.
Three specimens, Malaysia, Sabah, Kinabatangan River at Sukau, N 5°31′37.67″ E 118°17′39.23″, leg. M. de Bruyn 01.01.2009; two specimens ZMB 29579. Thailand, Phitsanulok, Tributary of Kaek River, c. 28 km E of Phitsanulok, N 16°50.866′ E 100°30.709′, leg. F. Köhler 27.03.2006, ZMB 29513.
3.9. Description
Cephalothorax and cephalic appendages: Small shrimp with carapace lengths of 1.9–4.3 mm (median 2.8 mm, n = 22) in apparently adult specimens. Carapace (
Figure 5A,B) smooth, antennal spine clearly separated from the suborbital lobe, pterygostomial angle broadly rounded. Rostrum (
Figure 5A,B) long, reaching well beyond distal margin of scaphocerite, 1.04–2.31 (median 1.97, n = 8) times as long as carapace, curved upward; dorsal margin armed with 7–9 widely spaced teeth on proximal part of the rostrum, no teeth or occasionally with one tooth behind orbital margin, one subapical tooth near tip, ventral margin with 35 teeth. Rostrum formula based on 18 specimens 0 – 1 + 6 – 9 + 1/19 − 35. Eyes (
Figure 5A,B) well developed with pigmented globular cornea. Antennular peduncle (
Figure 5A,B,G), 0.86–1.02 (median 0.91, n = 7) times as long as carapace in females, 0.89–1.08 (median 1.00, n = 6) times as long as carapace in males, first segment 1.85–2.33 (median 2.16, n = 12) times as long as second segment, second segment 1.61–2.75 (median 2.05, n = 12) times longer than third segment; distolateral tooth on first segment acute, well developed, 0.23–0.39 (median 0.33, n = 11) times as long as second segment; second and third segments unarmed, with few spiniform setae near distal margin. Stylocerite acuminate, reaching to 0.70–1.04 (median 0.77, n = 11) times the length of basal segment of antennular peduncle. Antennal scaphocerite (
Figure 5H) with well-developed distolateral tooth, 3.87–4.95 (median 4.66, n = 5) times as long as wide.
Pleon, telson and uropods: Sixth pleomere was 0.86–1.02 (median 0.92, n = 17) times the length of carapace, 1.81–2.41 (median 2.18, n = 17) times as long as the fifth pleomere, 1.08–1.30 (median 1.13, n = 15) times as long as telson. Telson (
Figure 5C,D) slightly tapering distally, distal margin broadly convex, lacking a median point with three or four (n = 10) pairs of dorsal short spiniform setae and one pair of short spiniform setae dorsolaterally; distal margin with two pairs of spiniform setae, lateral pair stronger and longer compared to inner setae. Preanal carina (
Figure 5E) with a prominent hook-like tooth. Uropodal diaeresis (
Figure 5F) with 5–9 movable spiniform setae, outermost seta clearly shorter than lateral angle.
Mouthparts and branchiae: Shown in
Figure 5I–N and
Figure 6A. Similar to
Caridina darwin n. sp. except that palp of first maxilliped (
Figure 5L,M) terminates in a short blunt extension. Third maxilliped (
Figure 6A) with last segment shorter than penultimate segment. Branchial apparatus as shown in
Table 4.
Pereiopods: Chela of first pereiopod (
Figure 6B) was caridinoid in shape, with well-developed palm, 1.75–2.32 (median 2.10, n = 9) times as long as wide, 1.16–1.54 (median 1.41, n = 9) times as long as carpus; fingertips rounded, with prominent hooks hidden by tufts of setae (
Figure 6C); dactylus 3.00–3.80 (median 3.29, n = 7) times as long as wide, 0.80–1.36 (median 1.13, n = 9) times as long as palm. Carpus slender, hardly excavated distally, 1.73–2.27 (median 2.00, n = 9) times as long as wide, 0.65–1.27 (median 1.14, n = 9) times as long as merus. Merus not inflated, 2.33–4.58 (median 2.48, n = 9) times as long as wide, 0.96–1.96 (median 1.06, n = 8) times as long as ischium. Chela of second pereiopod (
Figure 6D) slender, caridinoid in shape, with well-developed palm, 2.21–2.75 (median 2.47, n = 9) times as long as wide, 0.80–0.94 (median 0.88, n = 9) times as long as carpus; fingertips rounded, with prominent hooks hidden by tufts of setae, dactylus 3.80–4.44 (median 4.17, n = 7) times as long as wide, 1.25–1.73 (median 1.33, n = 9) times as long as palm. Carpus slender, 3.73–4.94 (median 4.31, n = 9) times as long as wide, 1.17–1.41 (median 1.32, n = 9) times as long as merus. Merus not inflated, 3.50–5.00 (median 4.40, n = 9) times as long as wide, 0.91–1.06 (median 0.97, n = 8) times as long as ischium. Third pereiopod (
Figure 6E–F) slender, dactylus (
Figure 6F) not sexually dimorphic, 2.71–3.75 (median 3.75, n = 3) times as long as wide (including terminal claw and spiniform setae on flexor margin), terminating in one claw, with 6–8 accessory spiniform setae on flexor margin; propodus not dilated or curved distally, distal spiniform seta not enlarged, 7.88–10.60 (median 10.00, n = 3) times as long as wide, 3.00–3.53 (median 3.32, n = 3) times as long as dactylus; carpus 5.38–6.67 (median 5.65, n = 4) times as long as wide, 0.62–0.64 (n = 2) times as long as propodus, 0.50–0.56 (median 0.52, n = 4) times as long as merus, with one large spiniform seta near distal margin and one small spiniform setae proximal; merus 8.56–9.67 (median 8.85, n = 4) times as long as wide, 1.79–2.00 (median 1.93, n =4) times as long as carpus, with two or three strong movable spiniform setae on externo-inferior margin; ischium with one spiniform seta (absent in one specimen examined). Fourth pereiopod similar to third one but slightly smaller. Fifth pereiopod (
Figure 6G,H) slender; dactylus (
Figure 6H) 3.00–3.80 (median 3.63, n = 6) times as long as wide (including terminal claw and spiniform setae on flexor margin), terminating in one large claw, with 21–35 spiniform setae on flexor margin; propodus with small spiniform setae on ventral margin, distal pair of spiniform setae not markedly enlarged, propodus 10.75–12.14 (median 11.54, n = 5) times as long as wide, 3.07–3.54 (median 3.33, n = 5) times as long as dactylus; carpus 5.17–5.75 (median 5.43, n = 3) times as long as wide, 0.51–0.55 (median 0.53, n = 4) times as long as propodus, 0.53–0.61 (median 0.56, n = 4) times as long as merus, with one strong spiniform seta near distal margin; merus 8.33–10.73 (median 9.57, n = 3) times as long as wide, 1.63–1.90 (median 1.79, n = 4) times as long as carpus, with two or three strong, appressed, movable spiniform setae on the externo-inferior margin. Ischium without spiniform setae.
Pleopods: Endopod of male first pleopod (
Figure 6I) elongated triangular, lacking appendix interna, 2.00—2.57 (median 2.45, n = 4) times as long as proximal width, 0.23–0.26 (median 0.24, n = 3) times as long as exopod, with long pappose setae on distal, inner and outer margins. Male second pleopod with appendix masculina rod-shaped (
Figure 6J), 9.33–11.50 (n = 2) times as long as wide, 0.68–0.72 (n = 2) times as long as endopod, armed with strong spiniform setae on inner and distal margins; appendix interna originating from about 0.4 times the length of appendix masculina, reaching to 0.74–0.81 (n = 2) times the length of appendix masculina.
Reproductive biology: Ovigerous females with medium-sized embryos (0.59 − 0.66 × 0.34 − 0.43 mm (n = 8)). A total of 25–108 embryos counted in four ovigerous specimens.
Size. Postorbital carapace length of apparently adult specimens 1.9–4.3 mm.
Colouration: Body transparent. Rostrum red, red chromatophores also arranged in a narrow line running along the sternites of the pleon, on the distal part of the exopod of the uropod and a red transverse line at the dorsal margin of the third pleomere. Embryos are greenish (Wilson 2008; Short, Page and Humphrey 2019).
Distribution: The species is known from rivers flowing into the Gulf of Carpentaria (Queensland) and to the Timor Sea near Darwin (Northern Territory).
Etymology: The species is named after the biologist and aquarist Dave Wilson, collector of some of the type material and an enthusiast on Australian fish and shrimp fauna.
Common name: Darwin Red Nose Shrimp.
Remarks:
Caridina wilsoni n. sp. Resembles
Caridina gracilirostris De Man, 1892, both in coloration and in general morphology (body very slender; rostrum strongly curved upwards with few widely spaced teeth on proximal part of dorsal margin, no postorbital teeth; preanal carina with one hook-like tooth; distal margin of telson with few spiniform setae). The proposed new species differs from this species in the larger size of the embryos (0.59–0.66 × 0.34–0.43 mm vs. 0.34–0.39 × 0.21–0.25 mm in
C. gracilirostris), a slightly stouter carpus of the second cheliped (3.73–4.94 times as long as wide vs. 4.69–5.26 times as long as wide in
C. gracilirostris), merus of third pereiopod 8.56–9.67 times as long as wide vs. 9.55–10.22 in
C. gracilirostris, propodus of fifth pereiopod 3.07–3.54 times as long as dactylus vs. 4.0–4.25 in
C. gracilirostris. Some of these morphological characters overlap slightly (length-to-width ratio of carpus of second cheliped, length-to-width ratio of merus of third pereiopod), but by combining the morphological characters, these two species can be distinguished. The proposed new species also resembles
C. gracillima Lanchester, 1901, known from the Malayan peninsula, especially in the size of the embryos (0.59 –0.66 × 0.34–0.43 mm vs. 0.55– 0.66 × 0.35–0.40 mm in
C. gracillima) [
26], but differs from this species in its preanal carina, bearing a hook-like spine vs. preanal carina rounded without a spine in
C. gracillima.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}