

Unveiling the Role of Concanavalin A in a Rodent Model of Chemical-Induced Hepatocellular Carcinoma: A Promising Guide in Understanding Liver Cancer Development
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Histopathological Analysis
2.4. Chemical and Reagents
2.5. Lectin Histochemistry of the Liver
2.6. Confocal Scanning Laser Microscopy (CSLM) Analysis
2.7. Concanavalin A Signal Quantification by CLSM
2.8. Statistics
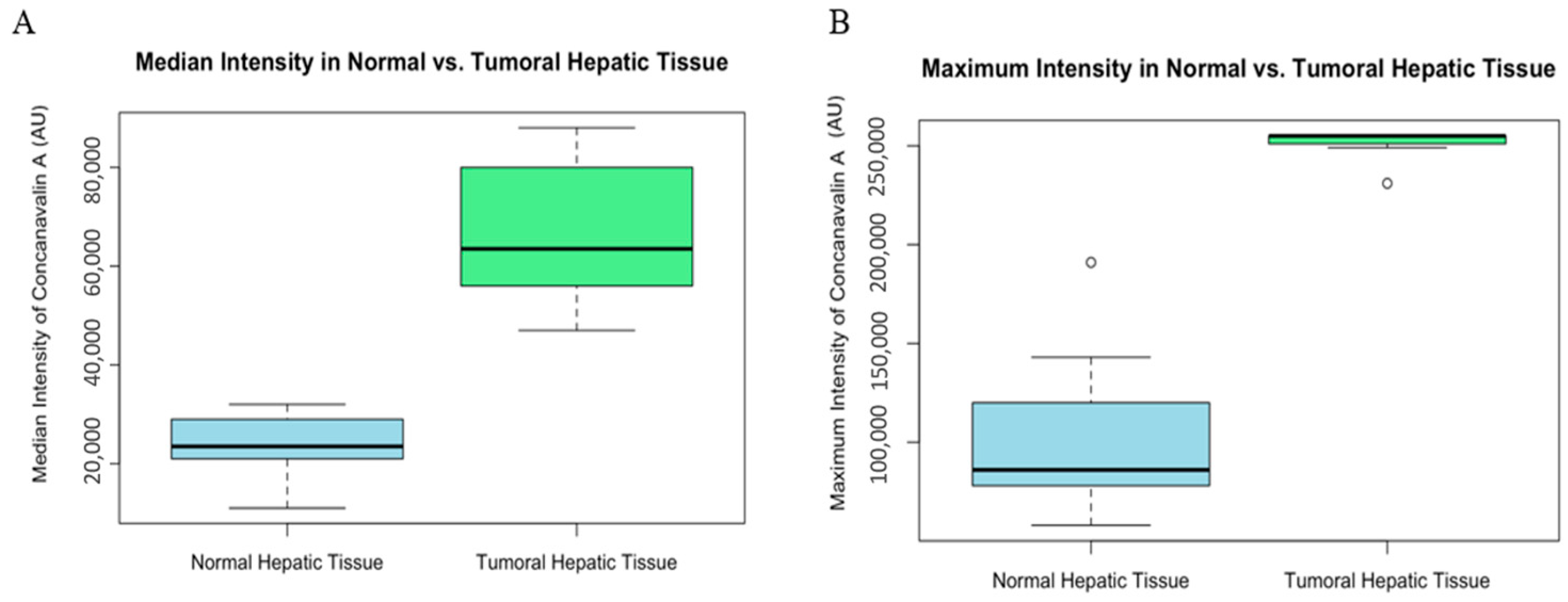
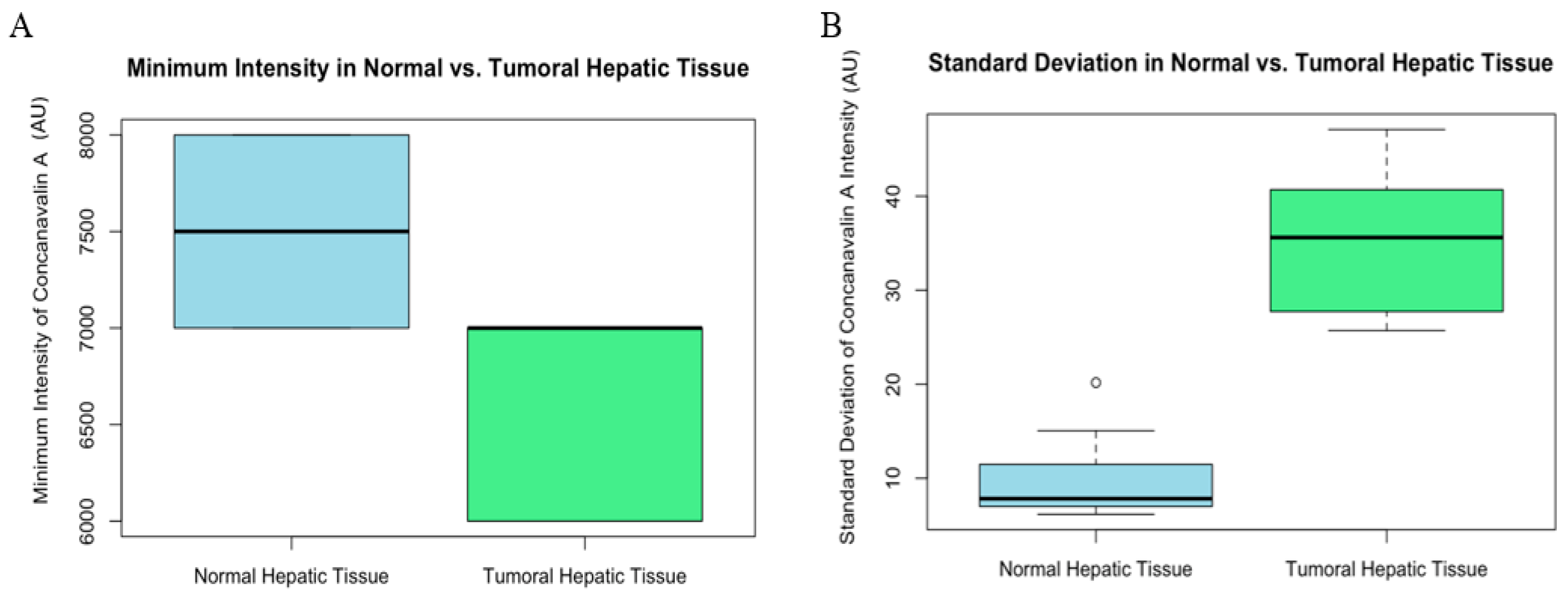
3. Results
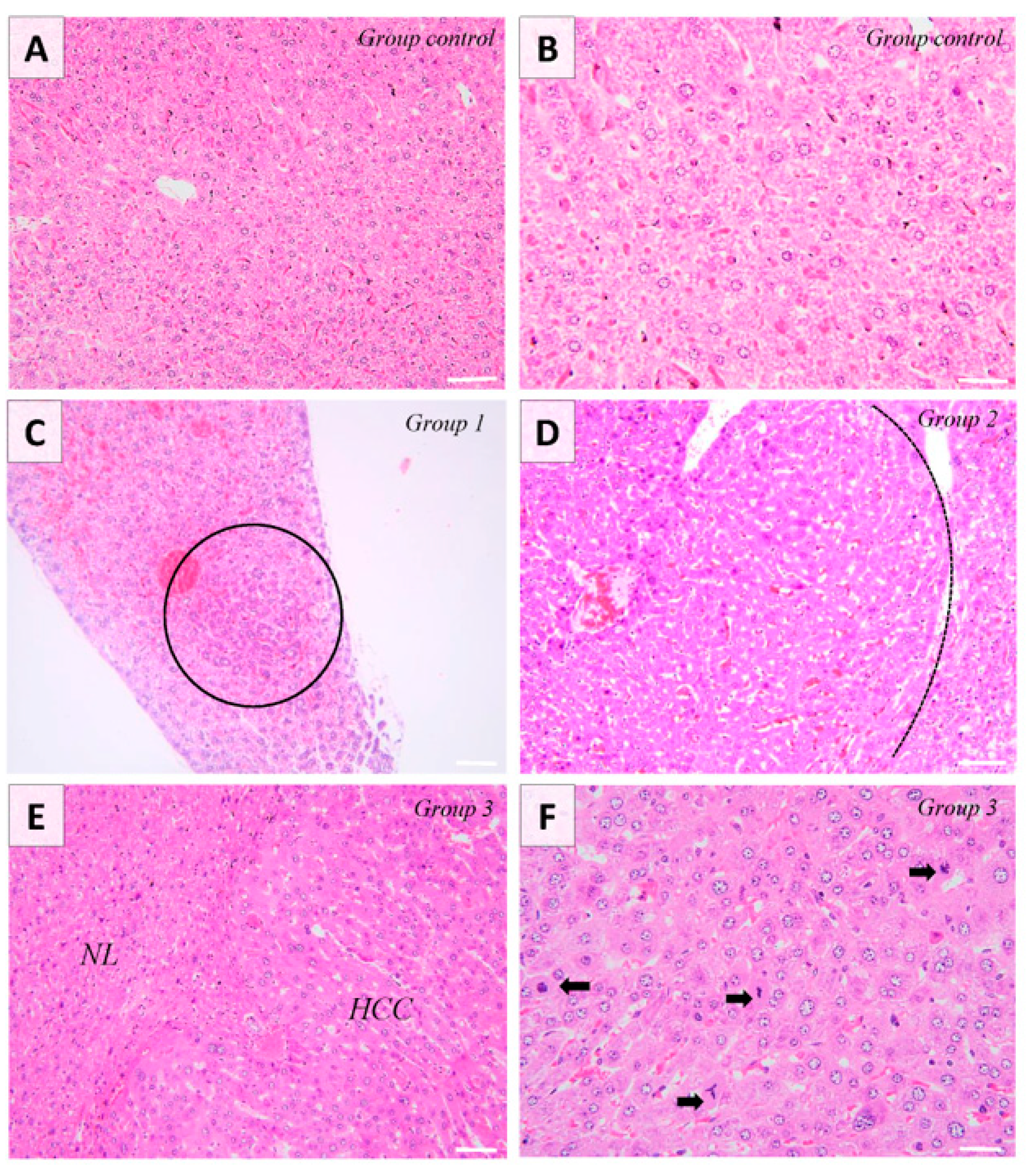
3.1. Gross and Histological Findings
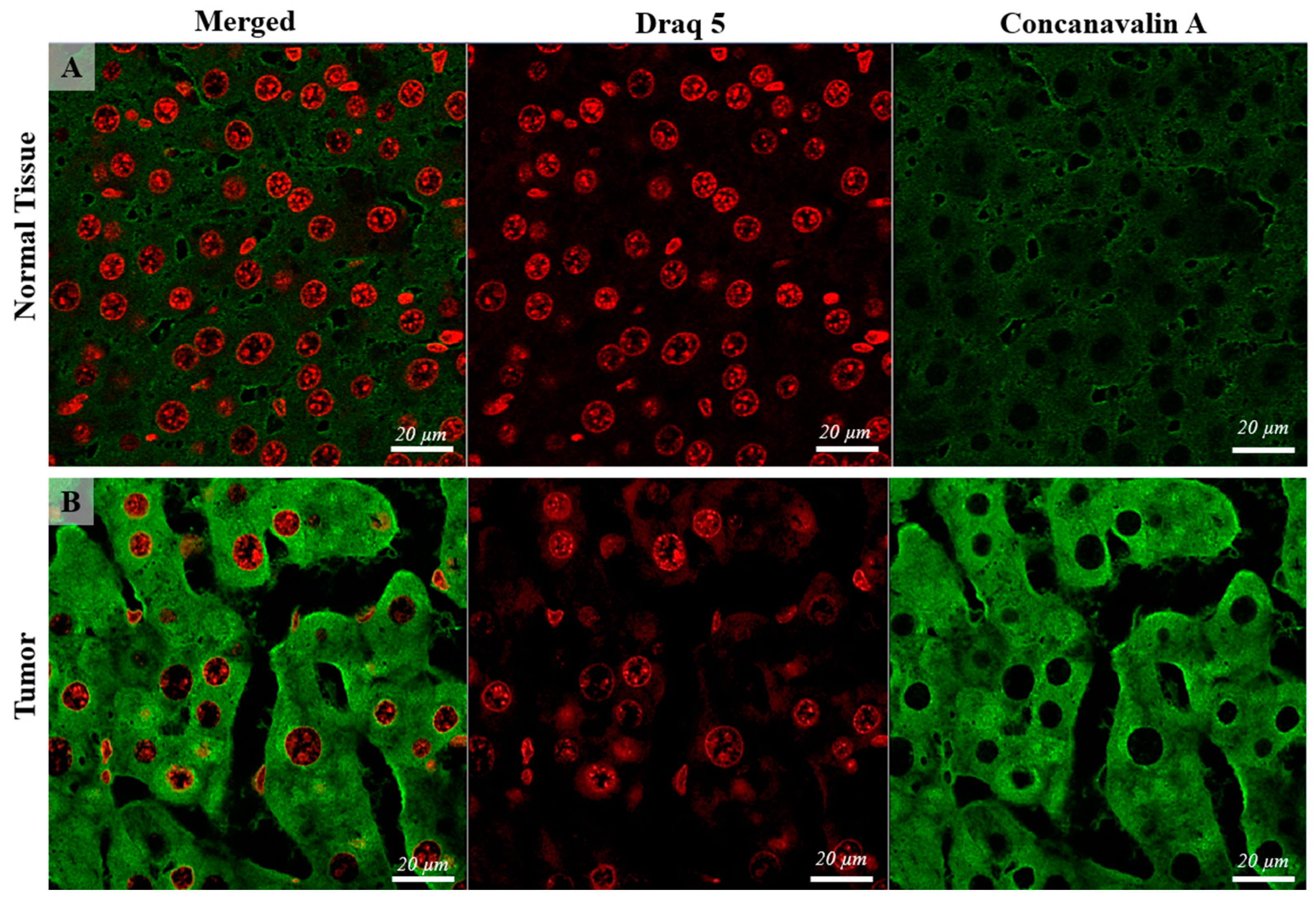
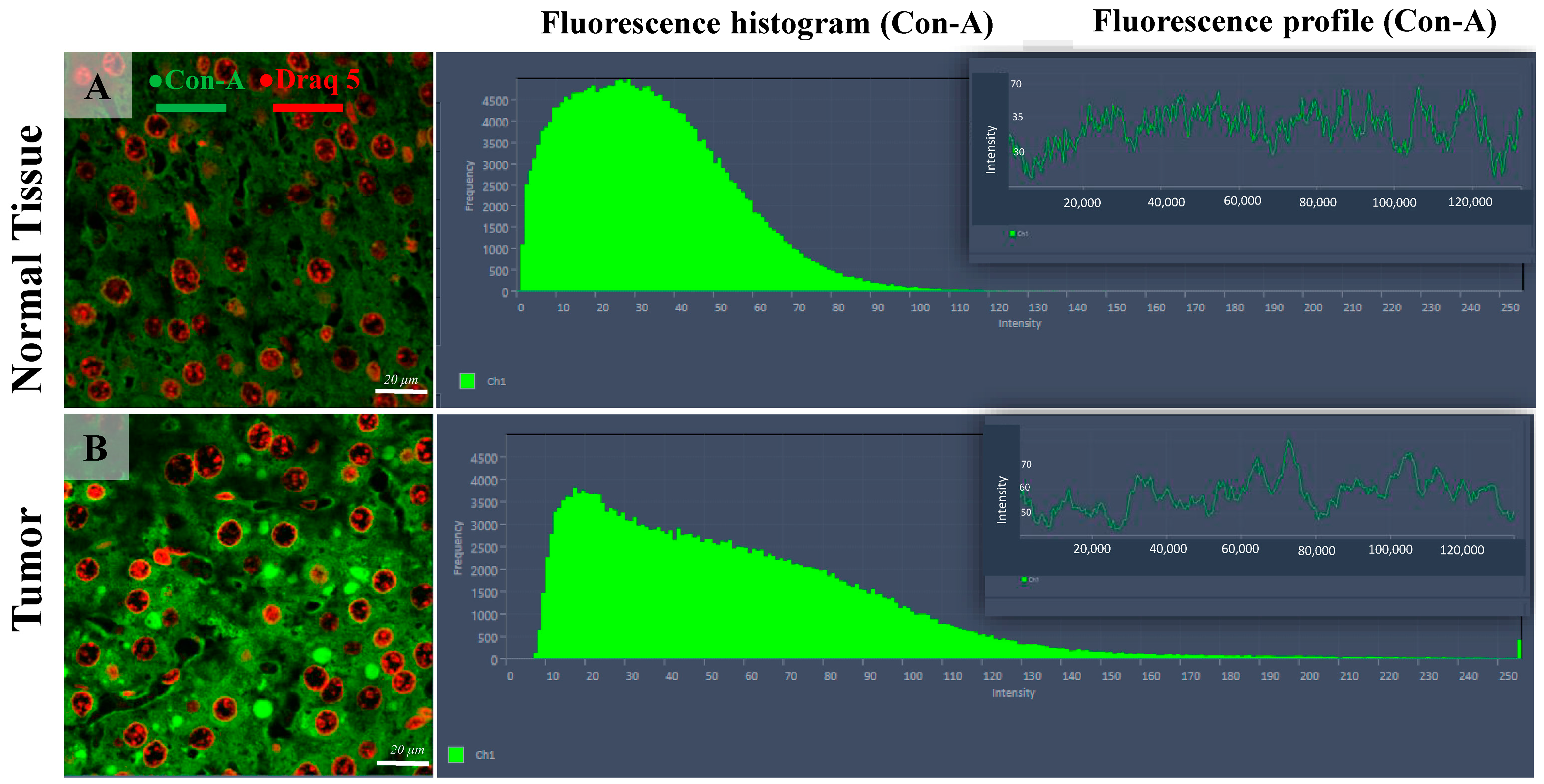
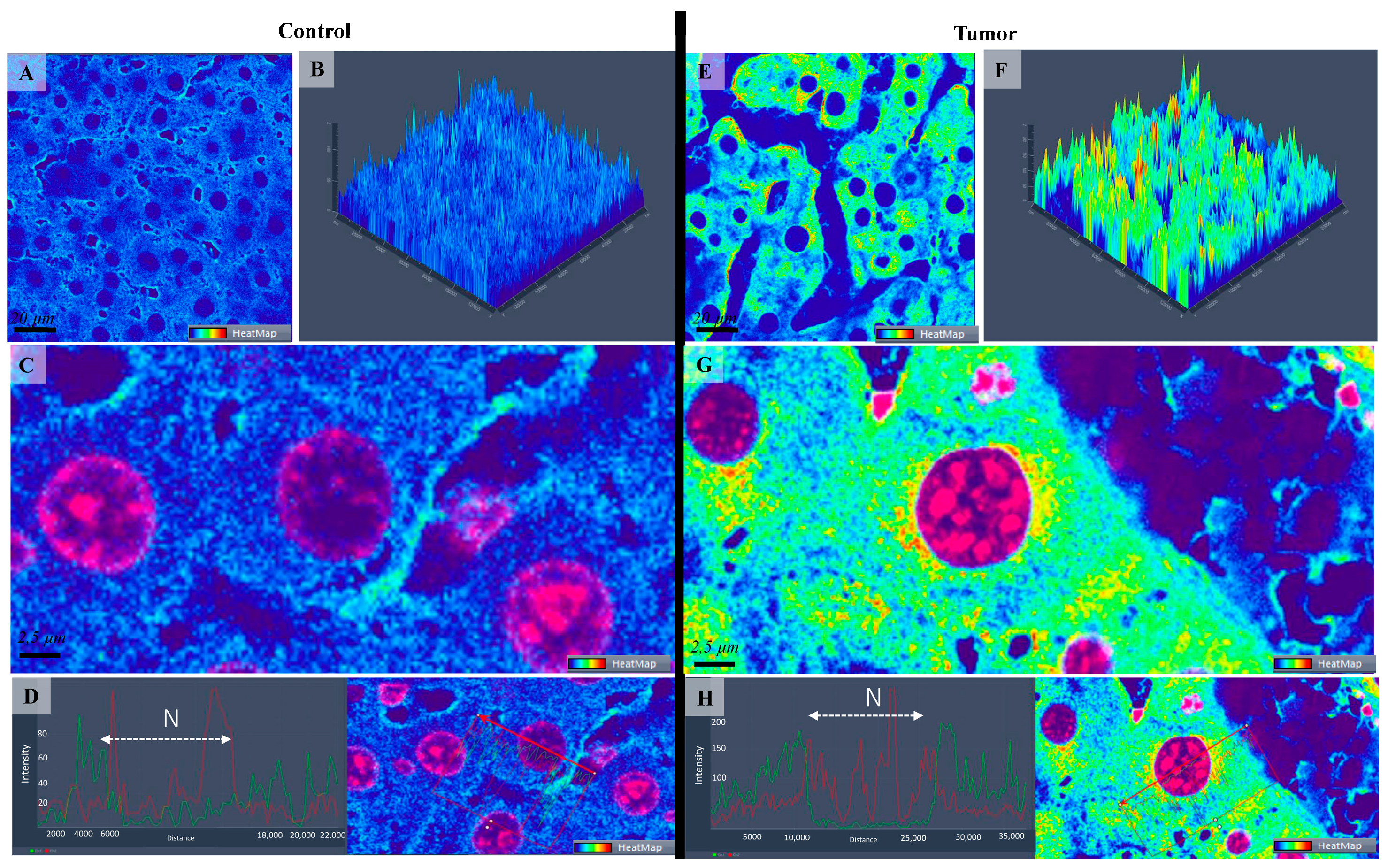
3.2. Confocal Scanning Laser Microscopy (CSLM)
3.3. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liptak, J.M.; Dernell, W.S.; Monnet, E.; Powers, B.E.; Bachand, A.M.; Kenney, J.G.; Withrow, S.J. Massive hepatocellular carcinoma in dogs: 48 cases (1992–2002). J. Am. Vet. Med. Assoc. 2004, 225, 1225–1230. [Google Scholar] [CrossRef]
- Goussev, S.A.; Center, S.A.; Randolph, J.F.; Kathrani, A.; Butler, B.P.; McDonough, S.P. Clinical characteristics of hepatocellular carcinoma in 19 cats from a single institution (1980–2013). J. Am. Anim. Hosp. Assoc. 2016, 52, 36–41. [Google Scholar] [CrossRef]
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef]
- Obeid, M.; Khabbaz, R.C.; Garcia, D.K.; Schachtschneider, M.K.; Gaba, R.C. Translational animal models for liver cancer. Am. J. Interv. Radiol. 2018, 2, 1–7. [Google Scholar] [CrossRef]
- Brown, Z.J.; Heinrich, B.; Greten, T.F. Mouse models of hepatocellular carcinoma: An overview and highlights for immunotherapy research. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 536–554. [Google Scholar] [CrossRef]
- Tolba, R.; Kraus, T.; Liedtke, C.; Schwarz, M.; Weiskirchen, R. Diethylnitrosamine (DEN)-induced carcinogenic liver injury in mice. Lab. Anim. 2015, 49 (Suppl. 1), 59–69. [Google Scholar] [CrossRef]
- Santos, N.P.; Pereira, I.C.; Pires, M.J.; Lopes, C.; Andrade, R.; Oliveira, M.M.; Colaço, A.; Peixoto, F.; Oliveira, P.A. Histology, bioenergetics and oxidative stress in mouse liver exposed to N-diethylnitrosamine. In Vivo 2012, 26, 921–929. [Google Scholar]
- De Matteis, S.; Ragusa, A.; Marisi, G.; De Domenico, S.; Casadei Gardini, A.; Bonafè, M.; Giudetti, A.M. Aberrant metabolism in hepatocellular carcinoma provides diagnostic and therapeutic opportunities. Oxidative Med. Cell. Longev. 2018, 2018, 7512159. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Li, J.; Wu, L.; Yu, Q.; Ji, J.; Wu, J.; Dai, W.; Guo, C. Emerging roles and the regulation of aerobic glycolysis in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2020, 39, 126. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.U.; Shariff, M.I.; Crossey, M.M.; Gomez-Romero, M.; Holmes, E.; Cox, I.J.; Rye, H.K.S.; Njie, R.; Taylor-Robinson, S.D. Hepatocellular carcinoma: Review of disease and tumor biomarkers. World J. Hepatol. 2016, 8, 471. [Google Scholar] [CrossRef] [PubMed]
- Torizuka, T.; Tamaki, N.; Inokuma, T.; Magata, Y.; Sasayama, S.; Yonekura, Y.; Tanaka, A.; Yamaoka, Y.; Yamamoto, K.; Konishi, J. In Vivo assessment of glucose metabolism in hepatocellular carcinoma with FDG-PET. J. Nucl. Med. 1995, 36, 1811–1817. [Google Scholar] [PubMed]
- Ye, Y.; Yu, B.; Wang, H.; Yi, F. Glutamine metabolic reprogramming in hepatocellular carcinoma. Front. Mol. Biosci. 2023, 10, 1242059. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Han, J.; Xing, H.; Zhang, H.; Li, Z.; Liang, L.; Li, C.; Dai, S.; Wu, M.; Shen, F.; et al. Dysregulated fatty acid metabolism in hepato-cellular carcinoma. Hepatic Oncol. 2016, 3, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Facciorusso, A.; Villani, R.; Bellanti, F.; Mitarotonda, D.; Vendemiale, G.; Serviddio, G. Mitochondrial signaling and hepatocellular carcinoma: Molecular mechanisms and therapeutic implications. Curr. Pharm. Des. 2016, 22, 2689–2696. [Google Scholar] [CrossRef] [PubMed]
- Mody, R.; Antaram Joshi, S.H.; Chaney, W. Use of lectins as diagnostic and therapeutic tools for cancer. J. Pharmacol. Toxicol. Methods 1995, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A. Emerging applications of lectins in cancer detection and biomedicine. Mater. Today Proc. 2020, 31, 651–661. [Google Scholar] [CrossRef]
- Schreiber, S.; Gocht, A.; Wegwitz, F.; Deppert, W.; Schumacher, U. Lectin histochemistry of murine WAP-T mammary cancer reveals similar glycoconjugate changes to those in human breast cancer. Anticancer Res. 2014, 34, 7045–7053. [Google Scholar] [PubMed]
- Indramanee, S.; Silsirivanit, A.; Pairojkul, C.; Wongkham, C.; Wongkham, S. Aberrant glycosylation in cholangiocarcinoma demonstrated by lectin-histochemistry. Asian Pac. J. Cancer Prev. 2012, 13, 119–124. [Google Scholar]
- Jauregui, H.O.; McMillan, P.N.; Hevey, K.; Naik, S. A quantitative analysis of lectin binding to adult rat hepatocyte cell surfaces. In Vitr. Cell. Dev. Biol. 1988, 24, 401–412. [Google Scholar] [CrossRef]
- Sumner, J.B. The globulins of the jack bean, Canavalia ensiformis: Preliminary paper. J. Biol. Chem. 1919, 37, 137–142. [Google Scholar] [CrossRef]
- Li, W.W.; Yu, J.Y.; Xu, H.L.; Bao, J.K. Concanavalin A: A potential anti-neoplastic agent targeting apoptosis, autophagy and anti-angiogenesis for cancer therapeutics. Biochem. Biophys. Res. Commun. 2011, 414, 282–286. [Google Scholar] [CrossRef]
- Murakami, I.; Sarker, A.B.; Hayashi, K.; Akagi, T. Lectin Binding Patterns in Normal Liver, Chronic Active Hepatitis, Liver Cirrhosis, and Hepatocellular Carcinoma an Immunohistochemical and Immunoelectron Microscopic Study. Pathol. Int. 1992, 42, 566–572. [Google Scholar] [CrossRef]
- Sherwani, A.F.; Mohmood, S.; Khan, F.; Khan, R.H.; Azfer, M.A. Characterization of lectins and their specificity in carcinomas—An appraisal. Indian J. Clin. Biochem. 2003, 18, 169–180. [Google Scholar] [CrossRef]
- Thoolen, B.; Maronpot, R.R.; Harada, T.; Nyska, A.; Rousseaux, C.; Nolte, T.; Malarkey, D.E.; Kaufmann, W.; Küttler, K.; Deschl, U.; et al. Proliferative and nonproliferative lesions of the rat and mouse hepatobiliary system. Toxicol. Pathol. 2010, 38 (Suppl. 7), 5S–81S. [Google Scholar] [CrossRef] [PubMed]
- Rebelo, A.L.; Contessotto, P.; Joyce, K.; Kilcoyne, M.; Pandit, A. An optimized protocol for combined fluorescent lectin/immunohistochemistry to characterize tissue-specific glycan distribution in human or rodent tissues. STAR Protoc. 2021, 2, 100237. [Google Scholar] [CrossRef] [PubMed]
- Clichici, S.; Biris, A.R.; Tabaran, F.; Filip, A. Transient oxidative stress and inflammation after intraperitoneal administration of multiwalled carbon nanotubes functionalized with single strand DNA in rats. Toxicol. Appl. Pharmacol. 2012, 259, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Zucker, R.M.; Price, O.T. Practical confocal microscopy and the evaluation of system performance. Methods 1999, 18, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Clichici, S.; Biris, A.R.; Catoi, C.; Filip, A.; Tabaran, F. Short-term splenic impact of single-strand DNA functionalized multi-walled carbon nanotubes intraperitoneally injected in rats. J. Appl. Toxicol. 2014, 34, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, D.E.; Weiss, D.; Carnell, P.H.; Vito, R.P.; Vega, D.; Gao, X.; Nie, S.; Taylor, W.R. Quantitative 3D fluorescence technique for the analysis of en face preparations of arterial walls using quantum dot nanocrystals and two-photon excitation laser scanning microscopy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R114–R123. [Google Scholar] [CrossRef] [PubMed]
- Pawley, J. The 39 steps: A cautionary tale of quantitative 3-D fluorescence microscopy. Biotechniques 2000, 28, 884–887. [Google Scholar] [CrossRef]
- Bartolini, D.; Dallaglio, K.; Torquato, P.; Piroddi, M.; Galli, F. Nrf2-p62 autophagy pathway and its response to oxidative stress in hepatocellular carcinoma. Transl. Res. 2018, 193, 54–71. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; Teufel, A.; Yevsa, T.; Staib, F.; Hohmeyer, A.; Walenda, G.; Zimmerman, H.W.; Vucur, M.; Huss, S.; Gassler, N.; et al. Adaptive immunity suppresses formation and progression of diethylnitrosamine-induced liver cancer. Gut 2012, 61, 1733–1743. [Google Scholar] [CrossRef]
- Aravalli, R.N.; Steer, C.J.; Cressman, E.N. Molecular mechanisms of hepatocellular carcinoma. Hepatology 2008, 48, 2047–2063. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Maronpot, R.R.; Boorman, G.A.; Morris, R.W.; Stitzel, K.A. Foci of cellular alteration in the rat liver: A review. J. Toxicol. Pathol. 1990, 3, 161–188. [Google Scholar] [CrossRef]
- Feo, F.; Frau, M.; Tomasi, M.L.; Brozzetti, S.; Pascale, R.M. Genetic and epigenetic control of molecular alterations in hepatocellular carcinoma. Exp. Biol. Med. 2009, 234, 726–736. [Google Scholar] [CrossRef]
- Jiang, Y.; Han, Q.-J.; Zhang, J. Hepatocellular carcinoma: Mechanisms of progression and immunotherapy. World J. Gastroenterol. 2019, 25, 3151. [Google Scholar] [CrossRef]
- Guillouzo, A.; Feldmann, G. Surface and intracellular localization of concanavalin A binding sites in rat liver cells. J. Histochem. Cytochem. 1977, 25, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Pattanaik, A.K.; Das, J.; Bhutia, S.K.; Behera, B.; Singh, P.; Maiti, T.K. Role of PI3K/Akt/mTOR and MEK/ERK pathway in Concanavalin A induced autophagy in HeLa cells. Chem. Biol. Interact. 2014, 210, 96–102. [Google Scholar] [CrossRef]
- Lei, H.-Y.; Chang, C.-P. Induction of autophagy by concanavalin A and its application in anti-tumor therapy. Autophagy 2007, 3, 402–404. [Google Scholar] [CrossRef]
- Leathem, A.J.; Brooks, S.A. Light microscopy: Overview and basic methods. In Lectin Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 1998; pp. 3–20. [Google Scholar]
- Brooks, S.A.; Hall, D.M.S. Lectin histochemistry to detect altered glycosylation in cells and tissues. In Metastasis Research Protocols; Springer: Berlin/Heidelberg, Germany, 2012; pp. 31–50. [Google Scholar]
- Díaz, M.C.; González, N.V.; Zanuzzi, C.N.; Najle, R.; Barbeito, C.G. Lectin histochemistry for detecting cadmium-induced changes in the glycosylation pattern of rat placenta. Biotech. Histochem. 2017, 92, 36–45. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pop, R.; Hodor, D.; Cătoi, C.; Mocan, T.; Mocan, L.; Tăbăran, A.-F. Unveiling the Role of Concanavalin A in a Rodent Model of Chemical-Induced Hepatocellular Carcinoma: A Promising Guide in Understanding Liver Cancer Development. Targets 2024, 2, 52-63. https://doi.org/10.3390/targets2010003
Pop R, Hodor D, Cătoi C, Mocan T, Mocan L, Tăbăran A-F. Unveiling the Role of Concanavalin A in a Rodent Model of Chemical-Induced Hepatocellular Carcinoma: A Promising Guide in Understanding Liver Cancer Development. Targets. 2024; 2(1):52-63. https://doi.org/10.3390/targets2010003
Chicago/Turabian StylePop, Romelia, Dragoș Hodor, Cornel Cătoi, Teodora Mocan, Lucian Mocan, and Alexandru-Flaviu Tăbăran. 2024. "Unveiling the Role of Concanavalin A in a Rodent Model of Chemical-Induced Hepatocellular Carcinoma: A Promising Guide in Understanding Liver Cancer Development" Targets 2, no. 1: 52-63. https://doi.org/10.3390/targets2010003