

Reduction of Cancer Stem Cells and Invasiveness of Human Melanoma and Breast Cancer by Cucurbitacin B from Lagenaria siceraria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
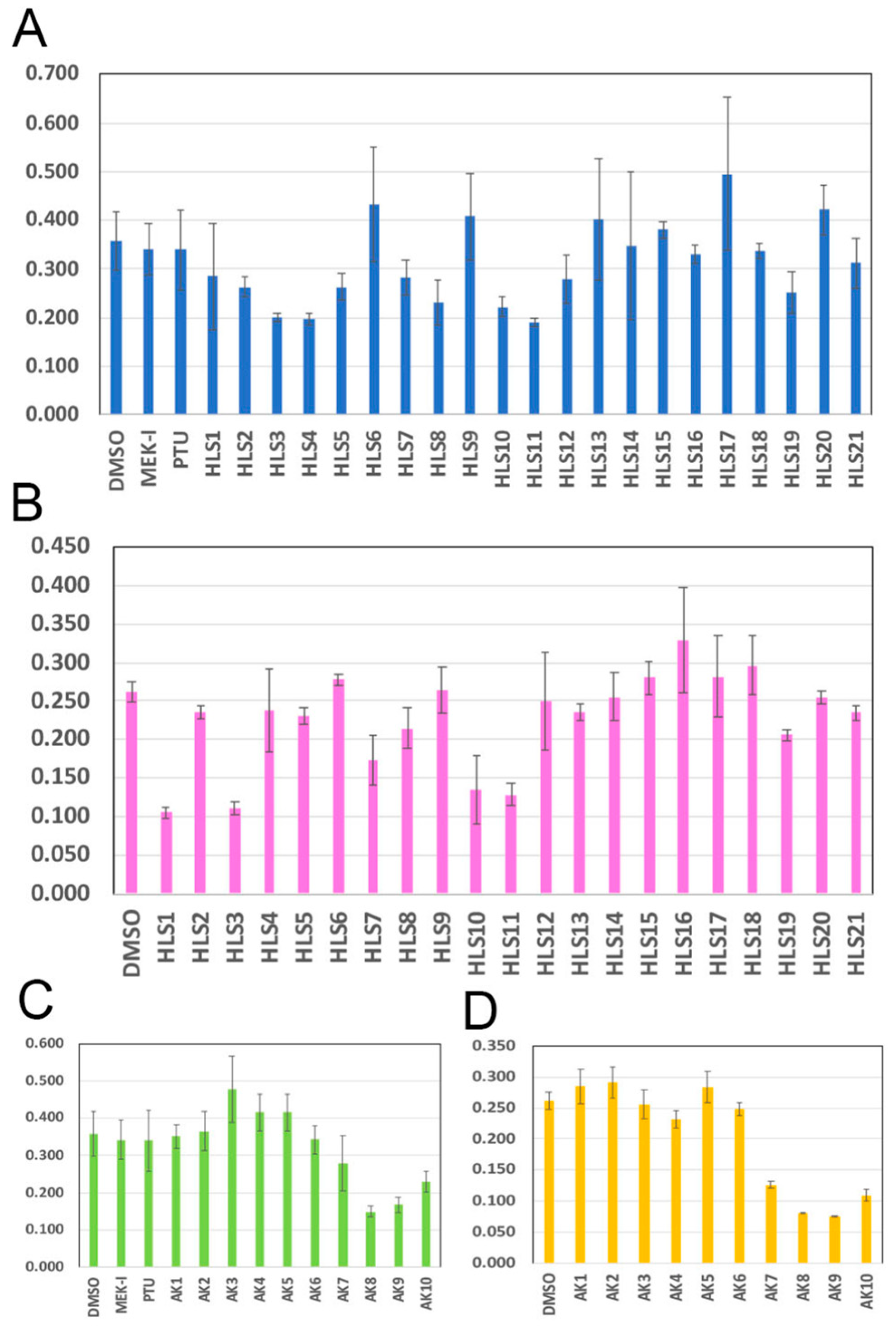
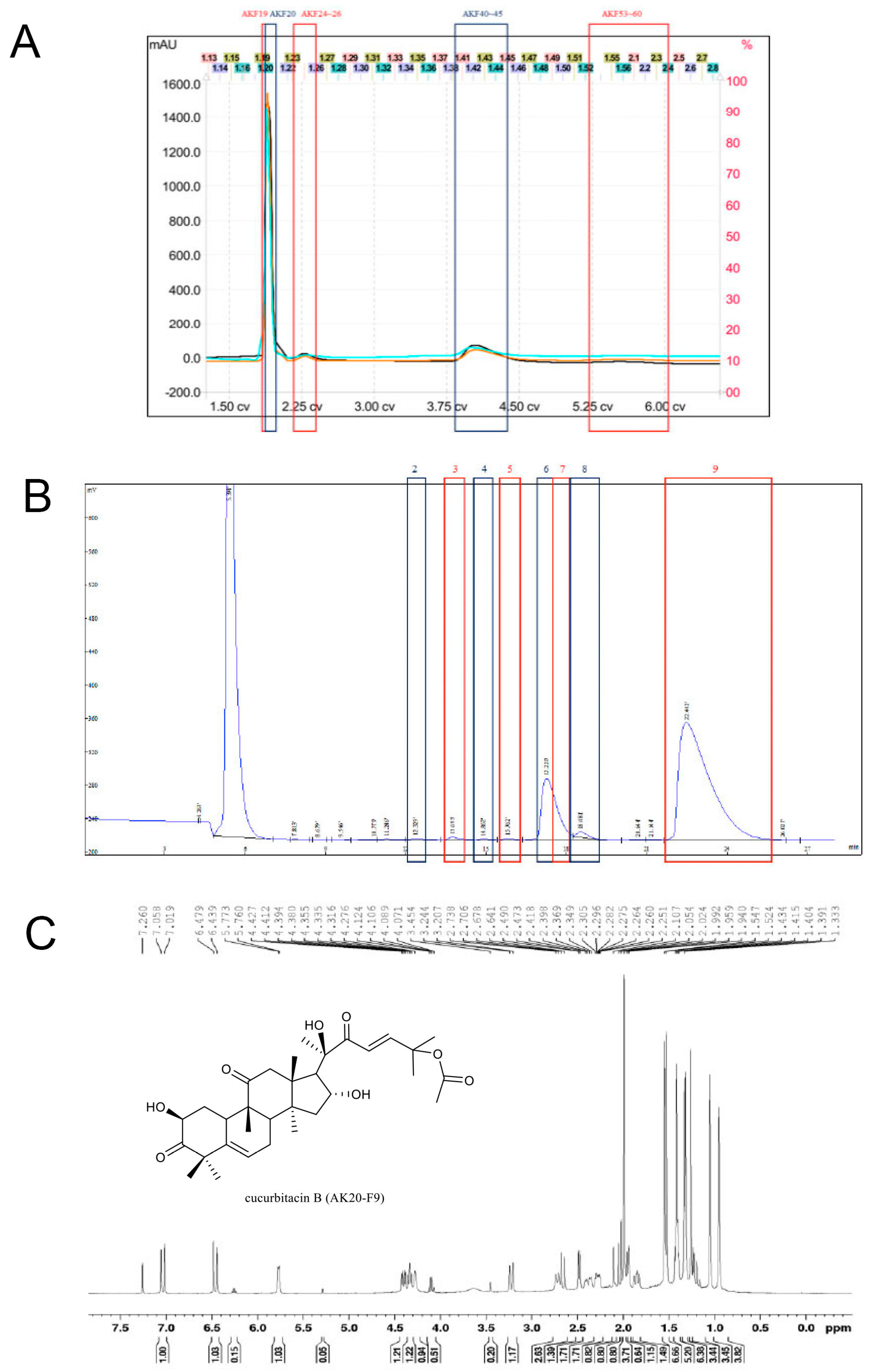
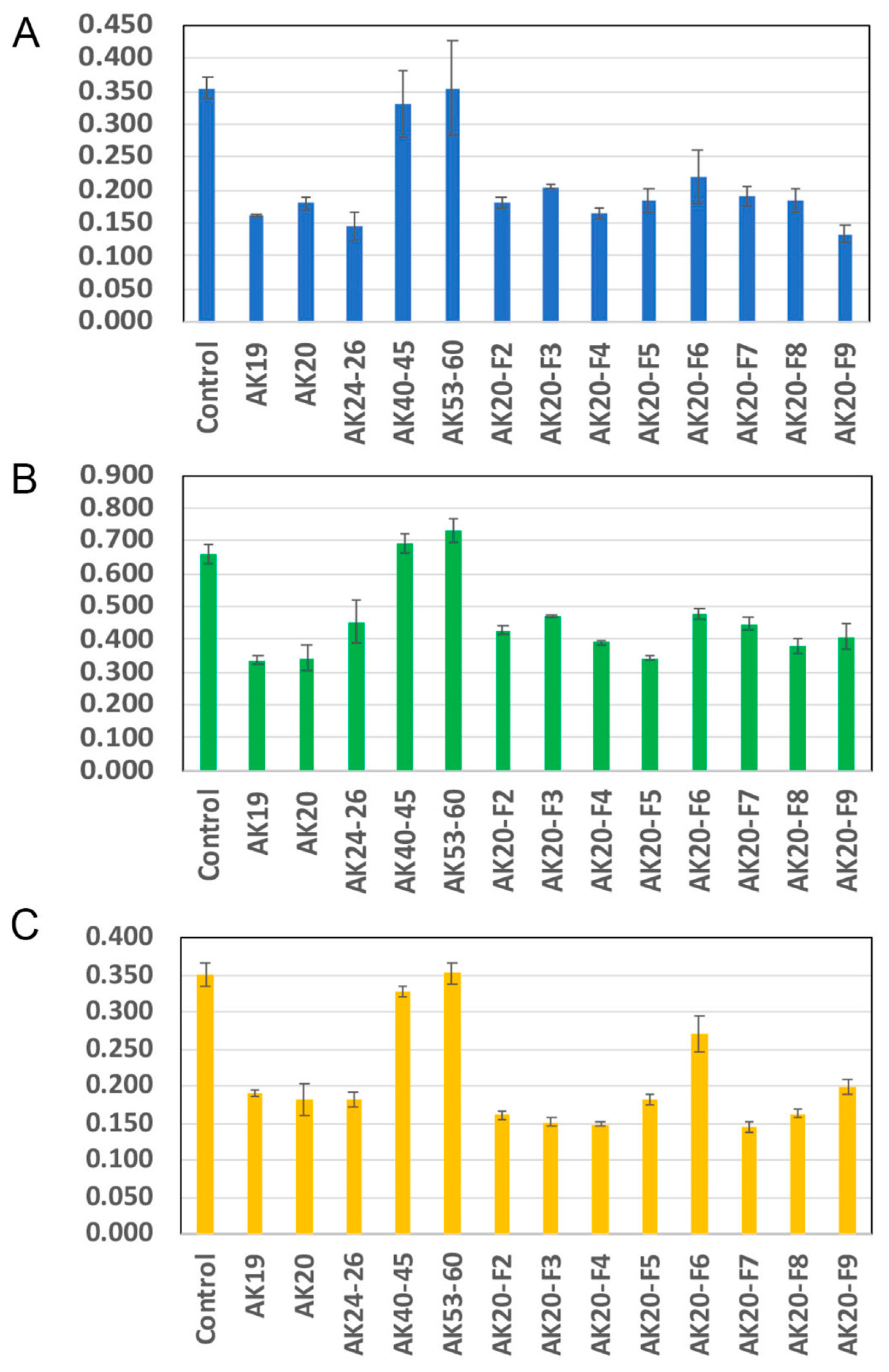
2.1. Anticancer Activity of Compounds in Bottle Gourd
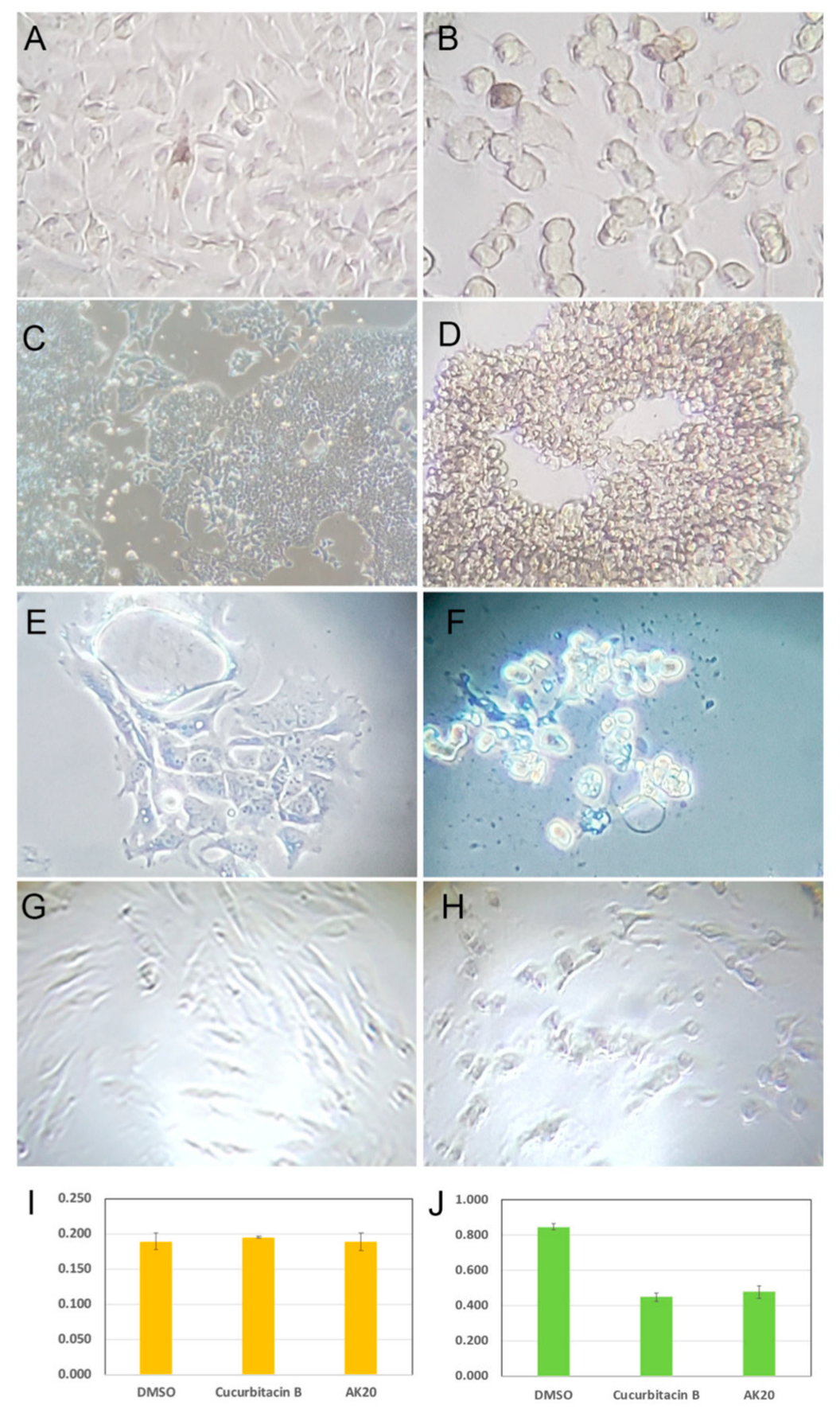
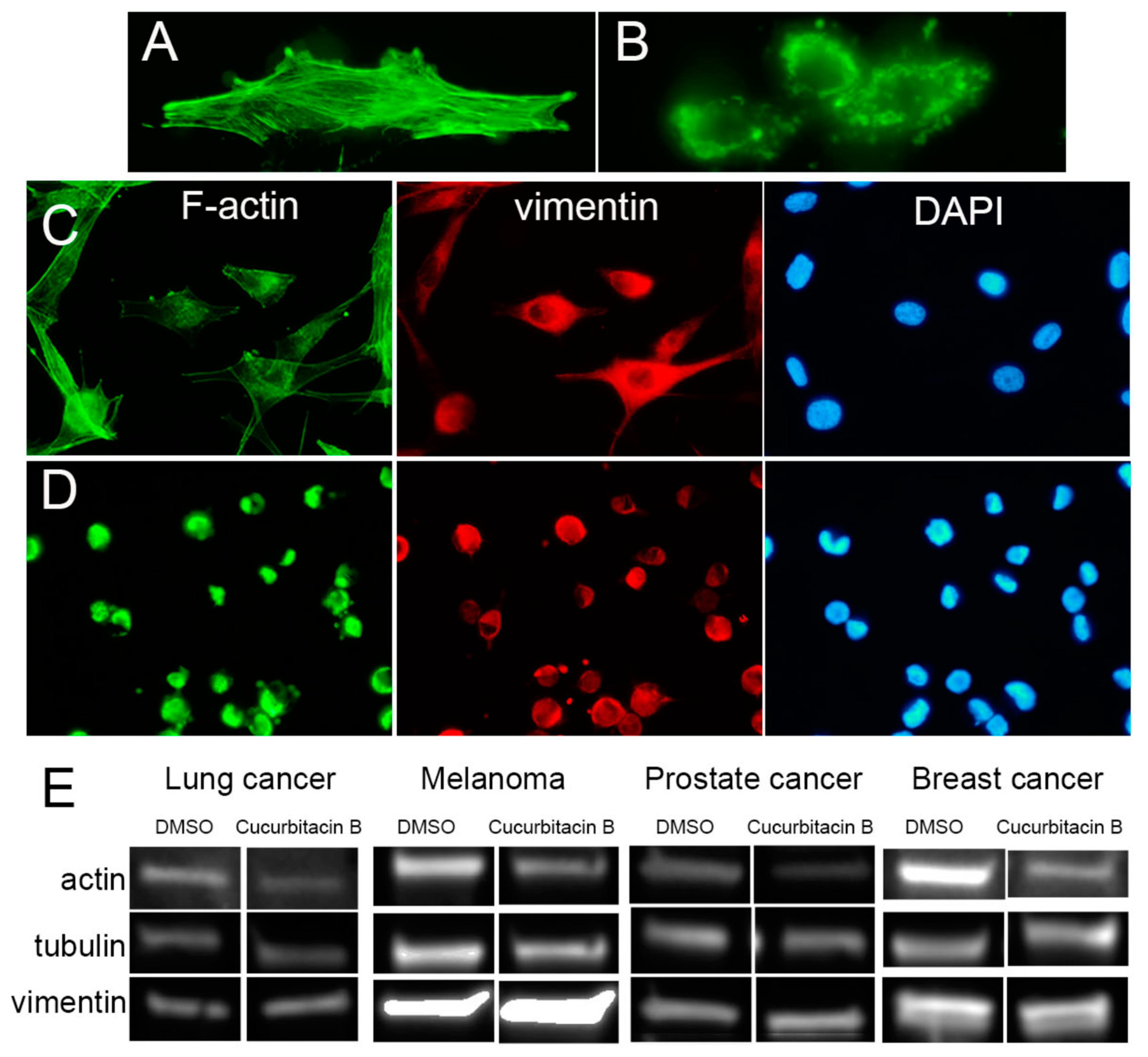
2.2. Cucurbitacin B Causes Cancer-Cell Retraction and Aggregation
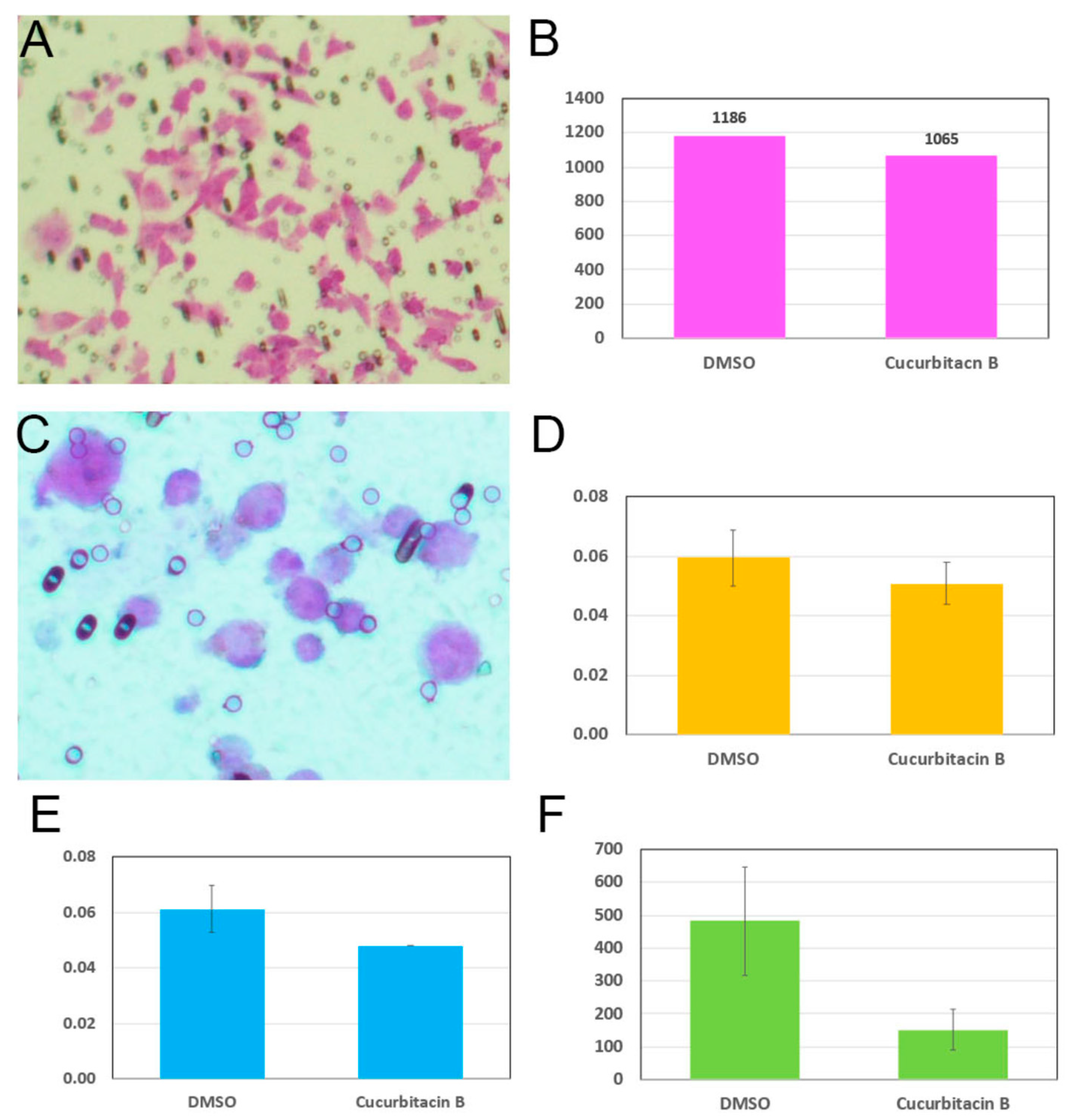
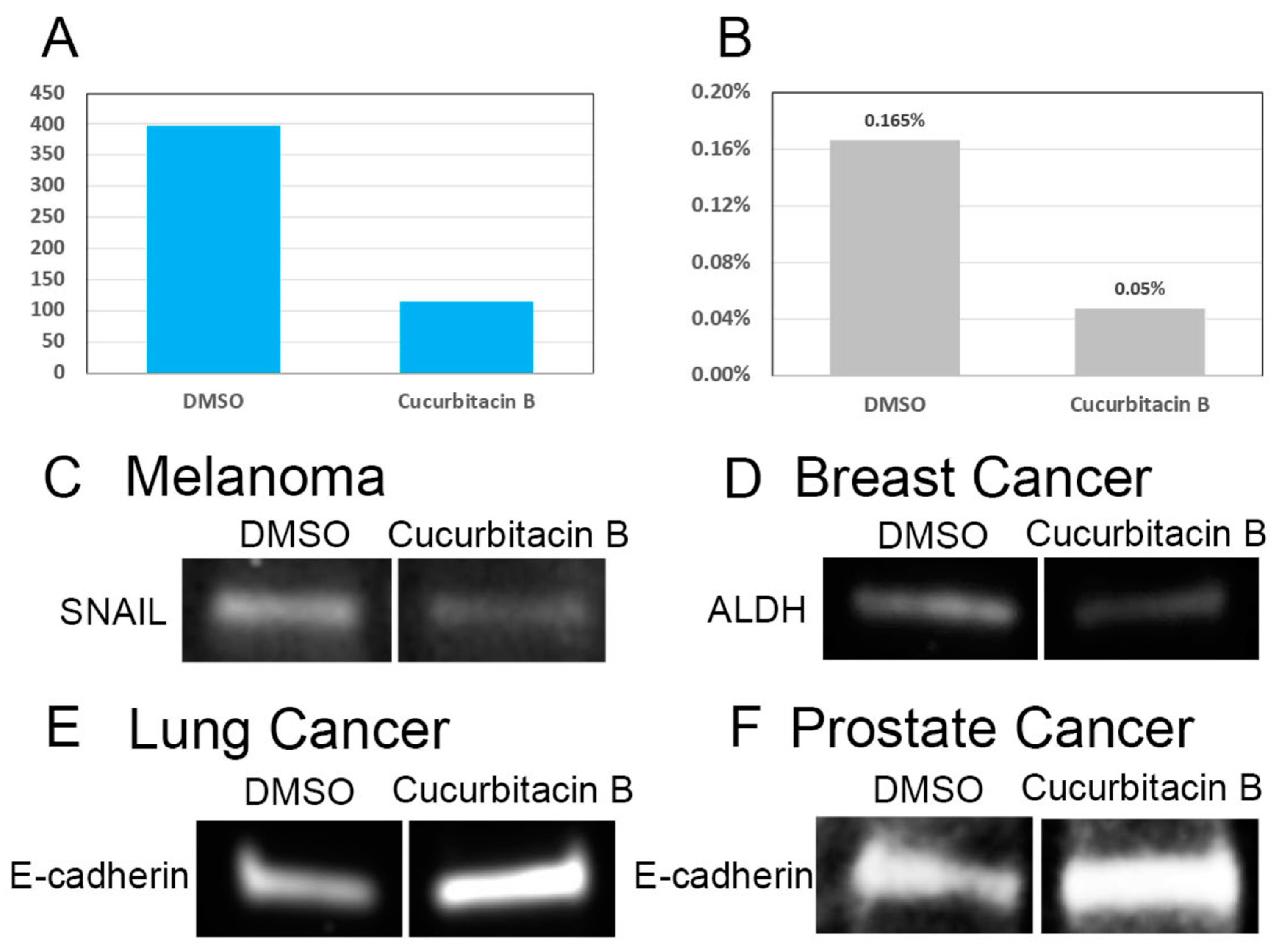
2.3. Cucurbitacin B Reduces Cancer-Cell Invasiveness and Cancer Stem Cells
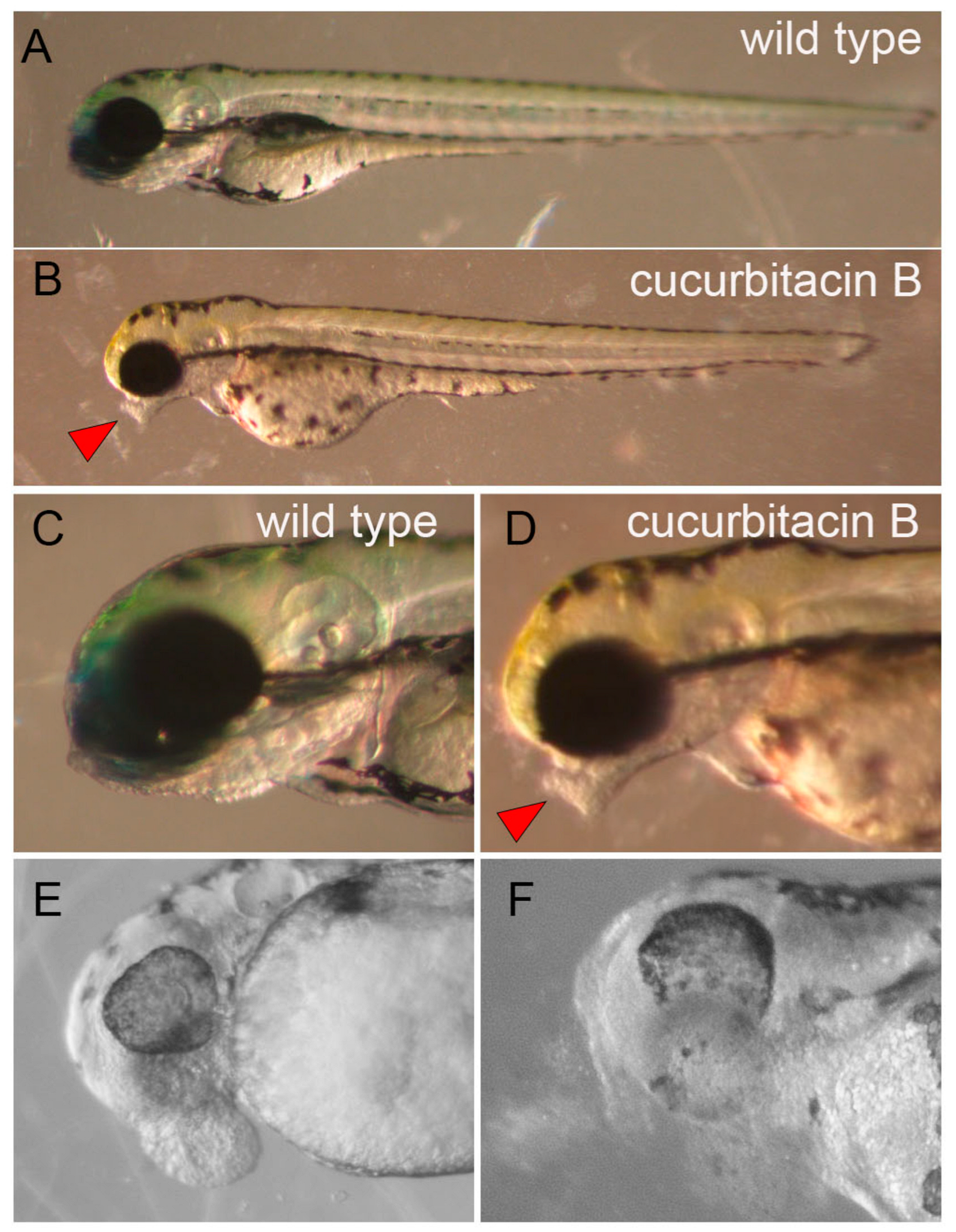
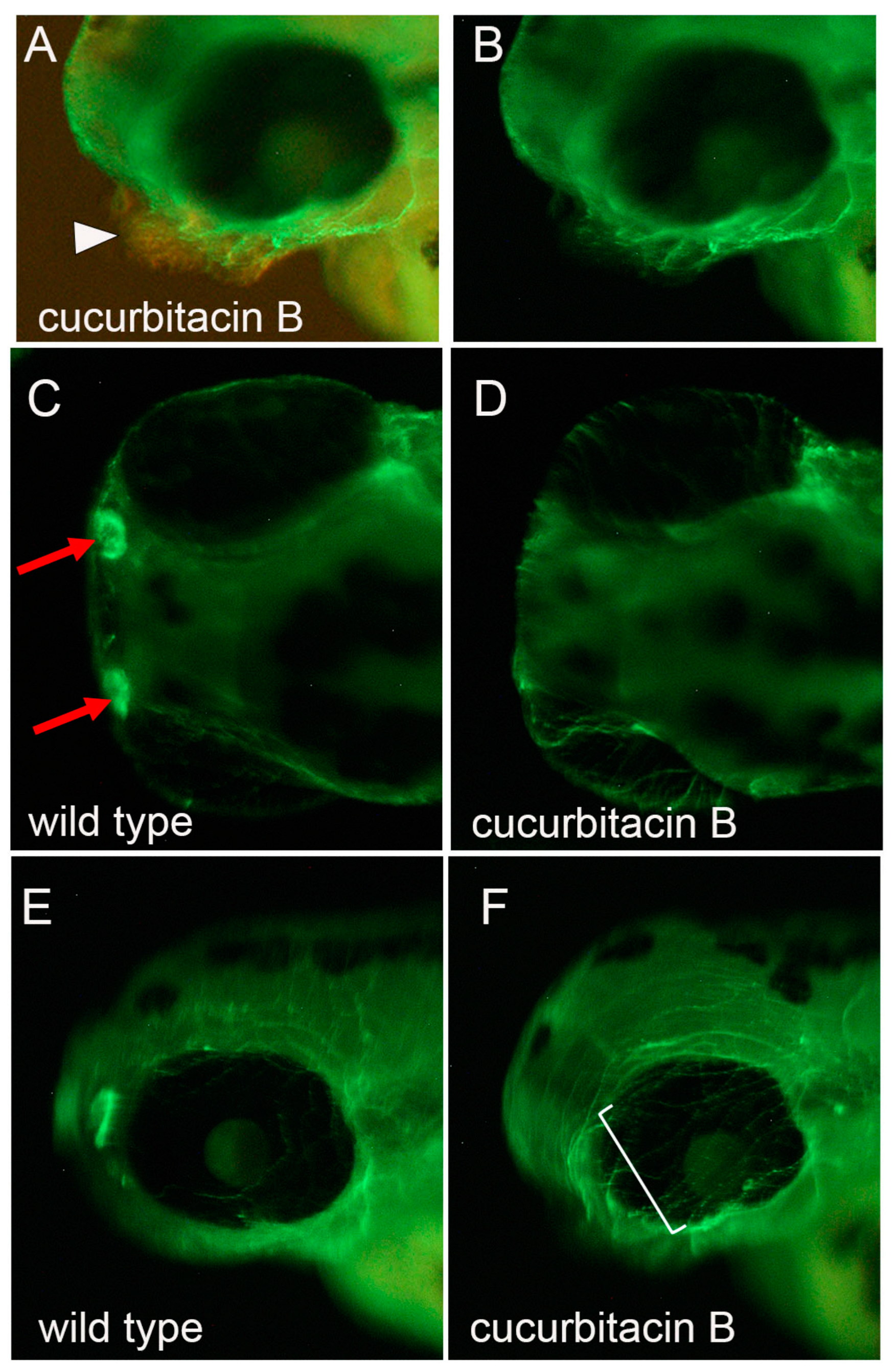
2.4. Cucurbitacin B Toxicity on Developing an Olfactory Organ in Zebrafish Embryo
3. Discussion
3.1. Cucurbitacin B for Cancer Metastasis Treatment
3.2. Cytoskeletal Restructuring and Stem-Cell Behaviors
3.3. Differences between Cucurbitacins and Other Cytoskeleton-Targeting Compounds
3.4. Cucurbitacin Toxicity in Developing Embryo
4. Materials and Methods
4.1. Chemical Extraction and Isolation
4.2. Cell Culture
4.3. MTT Assay
4.4. Immunostaining with Cancer Cells
4.5. Cell-Invasion Assay
4.6. Western Blot
4.7. Zebrafish Husbandry and In Vitro Fertilization
4.8. Chemical Preparation for Zebrafish Experiments
4.9. Chemical Treatment of Zebrafish Embryos
4.10. Immunohistochemistry with Zebrafish Embryos
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Seyfried, T.N.; Huysentruyt, L.C. On the origin of cancer metastasis. Crit. Rev. Oncog. 2013, 18, 43–73. [Google Scholar] [CrossRef] [PubMed]
- Valent, P.; Bonnet, D.; De Maria, R.; Lapidot, T.; Copland, M.; Melo, J.V.; Chomienne, C.; Ishikawa, F.; Schuringa, J.J.; Stassi, G.; et al. Cancer stem cell definitions and terminology: The devil is in the details. Nat. Rev. Cancer 2012, 12, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Ishizawa, K.; Rasheed, Z.A.; Karisch, R.; Wang, Q.; Kowalski, J.; Susky, E.; Pereira, K.; Karamboulas, C.; Moghal, N.; Rajeshkumar, N.V.; et al. Tumor-initiating cells are rare in many human tumors. Cell Stem Cell 2010, 7, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Walcher, L.; Kistenmacher, A.-K.; Suo, H.; Kitte, R.; Dluczek, S.; Straub, A.; Blaudszun, A.-R.; Yevsa, T.; Fricke, S.; Kossatz-Boehlert, U. Cancer stem cells- origins and biomarkers: Perspectives for targeted personalized therapies. Front. Immunol. 2020, 11, 1280. [Google Scholar] [CrossRef]
- Dalerba, P.; Clarke, M.F. Cancer stem cells and tumor metastasis: First steps into uncharted territory. Cell Stem Cell 2007, 1, 241–242. [Google Scholar] [CrossRef]
- Vasaikar, S.V.; Deshmukh, A.P.; Hollander, P.; Addanki, S.; Kuburich, N.A.; Kudaravalli, S.; Joseph, R.; Chang, J.T.; Soundararajan, R.; Mani, S.A. EMTome: A resource for pan-cancer analysis of epithelial-mesenchymal transition genes and signatures. Br. J. Cancer 2021, 124, 259–269. [Google Scholar] [CrossRef]
- Tania, M.; Khan, M.A.; Fu, J. Epithelial to mesenchymal transition inducing transcription factors and metastatic cancer. Tumor Biol. 2014, 35, 7335–7342. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Yu, T.-S.; McKay, R.M.; Burns, D.K.; Kernie, S.G.; Parada, L.F. A restricted cell population propagates glioblastoma growth after chemotherapy. Nature 2012, 488, 522–526. [Google Scholar] [CrossRef]
- Malladi, S.; Macalinao, D.G.; Jin, X.; He, L.; Basnet, H.; Zou, Y.; de Stanchina, E.; Massagué, J. Metastatic Latency and Immune Evasion through Autocrine Inhibition of WNT. Cell 2016, 165, 45–60. [Google Scholar] [CrossRef]
- Kaushik, U.; Aeri, V.; Mir, S.R. Cucurbitacins—An insight into medicinal leads from nature. Pharmacogn. Rev. 2015, 9, 12–18. [Google Scholar]
- Garg, S.; Kaul, S.C.; Wadhwa, R. Cucurbitacin B and cancer intervention: Chemistry, biology and mechanisms (Review). Int. J. Oncol. 2018, 52, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Bao, J.; Guo, J.; Ding, Q.; Lu, J.; Huang, M.; Wang, Y. Biological activities and potential molecular targets of cucurbitacins: A focus on cancer. Anti-Cancer Drugs 2012, 23, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.L.; Duncan, M.D.; Alley, M.C.; Sausville, E.A. Cucurbitacin E-induced disruption of the actin and vimentin cytoskeleton in prostate carcinoma cells. Biochem. Pharmacol. 1996, 52, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tanaka, M.; Peixoto, H.S.; Wink, M. Cucurbitacins: Elucidation of their interactions with the cytoskeleton. PeerJ 2017, 5, e3357. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Zhang, B.; Zhang, S.; Duan, C.; Cao, Y.; Kang, W.; Yan, H.; Ding, X.; Zhou, F.; Wu, L.; et al. Low nanomolar concentrations of Cucurbitacin-I induces G2/M phase arrest and apoptosis by perturbing redox homeostasis in gastric cancer cells in vitro and in vivo. Cell Death Dis. 2016, 7, e2106. [Google Scholar] [CrossRef] [PubMed]
- Litaudon, M.; Gaspard, C.; Sevenet, T. Morierinin: A New Cytotoxic Cucurbitacin from the Leaves of Morierina Montana Vieill. Nat. Prod. Res. 2003, 17, 229–233. [Google Scholar] [CrossRef]
- Boiko, A.D.; Razorenova, O.V.; Rijn, M.V.D.; Swetter, S.M.; Johnson, D.L.; Ly, D.P.; Butler, P.D.; Yang, G.P.; Joshua, B.; Kaplan, M.J.; et al. Human melanoma-initiating cells express neural crest nerve growth factor receptor CD271. Nature 2010, 466, 133–137. [Google Scholar] [CrossRef]
- Yin, Q.; Shi, X.; Lan, S.; Jin, H.; Wu, D. Effect of melanoma stem cells on melanoma metastasis. Oncol. Lett. 2021, 22, 566. [Google Scholar] [CrossRef]
- Yosuoka, A.; Hirose, Y.; Yoda, H.; Aihara, Y.; Suwa, H.; Niwa, K.; Sasado, T.; Morinaga, C.; Deguchi, T.; Henrich, T.; et al. Mutations affecting the formation of posterior lateral line system in Medaka, Oryzias latipes. Mech. Dev. 2004, 121, 729–738. [Google Scholar] [CrossRef]
- Schatton, T.; Murphy, G.F.; Frank, N.Y.; Yamaura, K.; Waaga-Gasser, A.M.; Gasser, M.; Zhan, Q.; Jordan, S.; Duncan, L.M.; Weishaupt, C.; et al. Identification of cells initiating human melanoma. Nature 2008, 451, 345–349. [Google Scholar] [CrossRef]
- Quintana, E.; Shackleton, M.; Sabel, M.S.; Fullen, D.R.; Johnson, T.M.; Morrison, S.J. Efficient tumor formation by single human melanoma cells. Nature 2008, 456, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Farahani, E.; Patra, H.K.; Jangamreddy, J.R.; Rashedi, I.; Kawalec, M.; Rao Pariti, R.K.; Batakis, P.; Wiechec, E. Cell adhesion molecules and their relation to (cancer) cell stemness. Carcinogenesis 2014, 35, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Bonneau, B.; Popgeorgiev, N.; Prudent, J.; Gillet, G. Cytoskeleton dynamics in early zebrafish development. BioArchitecture 2011, 1, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Villamayor, P.R.; Arana, A.J.; Coppel, C.; Ortiz-Leal, I.; Torres, M.V.; Sanches-Quinteiro, P.; Sanches, L. A comprehensive structural, lectin and immunohistochemical characterization of the zebrafish olfactory system. Sci. Rep. 2021, 11, 8865. [Google Scholar] [CrossRef]
- Reiten, I.; Uslu, F.E.; Fore, S.; Pelgrims, R.; Ringers, C.; Verdugo, C.D.; Hoffman, M.; Lal, P.; Kawakami, K.; Pekkan, K.; et al. Motile-cilia-mediated flow improves sensitivity and temporal resolution of olfactory computations. Curr. Biol. 2017, 27, 166–174. [Google Scholar] [CrossRef]
- Schlaeger, N.M.; Berkan, C.; Albu, I.; Monte, A.P.; Huang, C.-C. Synthetic compounds that inhibit melanoma growth and invasiveness by reducing cancer stem cell population. J. Cancer Biol. Res. 2020, 8, 1126. [Google Scholar]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio); University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- Haege, E.R.; Huang, H.-C.; Huang, C.-C. Identification of lactate as a cardiac protectant by inhibiting inflammation and cardiac hypertrophy using a zebrafish acute heart failure model. Pharmaceuticals 2021, 14, 261. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.-C.; Balding, K.K.; Zimmerman, S.J.; Chang, C.-Y.; Lu, S.-M.; Huang, H.-C. Reduction of Cancer Stem Cells and Invasiveness of Human Melanoma and Breast Cancer by Cucurbitacin B from Lagenaria siceraria. Drugs Drug Candidates 2023, 2, 358-376. https://doi.org/10.3390/ddc2020019
Huang C-C, Balding KK, Zimmerman SJ, Chang C-Y, Lu S-M, Huang H-C. Reduction of Cancer Stem Cells and Invasiveness of Human Melanoma and Breast Cancer by Cucurbitacin B from Lagenaria siceraria. Drugs and Drug Candidates. 2023; 2(2):358-376. https://doi.org/10.3390/ddc2020019
Chicago/Turabian StyleHuang, Cheng-Chen, Kiera K. Balding, Sydney J. Zimmerman, Che-Yuan Chang, Si-Min Lu, and Hui-Chi Huang. 2023. "Reduction of Cancer Stem Cells and Invasiveness of Human Melanoma and Breast Cancer by Cucurbitacin B from Lagenaria siceraria" Drugs and Drug Candidates 2, no. 2: 358-376. https://doi.org/10.3390/ddc2020019