
Anatomical Differences in the Omasum of Weaning Calves Fed with Different Diets



Abstract
:1. Introduction
2. Materials and Methods
2.1. Euthanasia and Consideration of Ethical Aspects
2.2. Study Methods
2.3. Nomenclature
2.4. Statistics
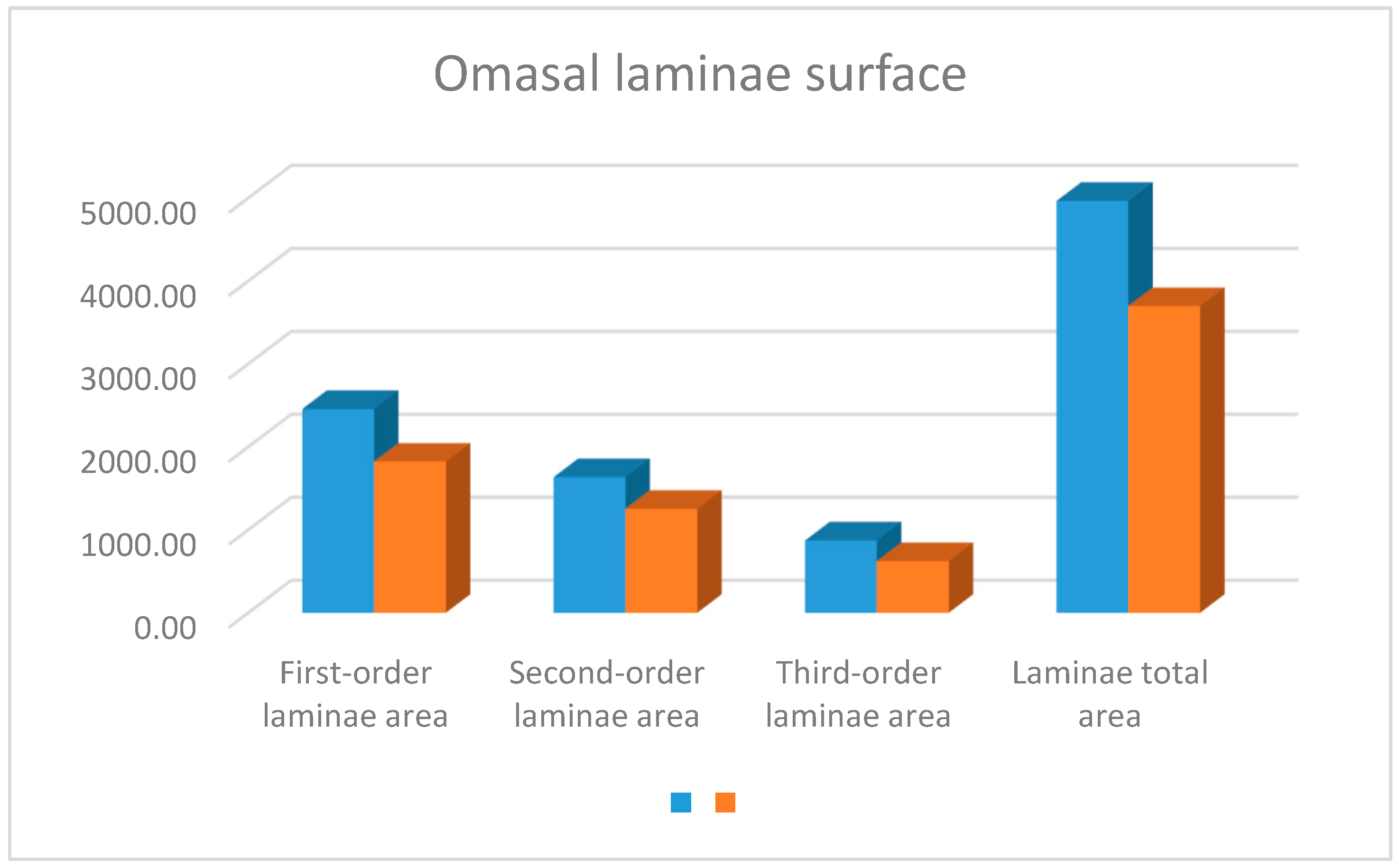
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drackley, J.K. Calf nutrition from birth to breeding. Vet. Clin. N. Am. Food Anim. Pract. 2008, 24, 55–86. [Google Scholar] [CrossRef] [PubMed]
- Guilloteau, P.; Zabielski, R.; Blum, J.W. Gastrointestinal tract and digestion in the young ruminant: Ontogenesis, adaptations, consequences and manipulations. J. Physiol. Pharm. 2009, 60 (Suppl. S1), 37–46. [Google Scholar]
- Khan, M.A.; Lee, H.J.; Lee, W.S.; Kim, H.S.; Kim, S.B.; Ki, K.S. Pre- and post-weaning behavior and performance of dairy calves fed milk through step-down and conventional methods. J. Dairy Sci. 2016, 99, 3735–3744. [Google Scholar]
- Repetto, J.L.; Mendoza, A.; Antúnez, G.; Cajarville, C. Nuevos paradigmas en la Cría y Recría de Hembras lecheras. In Jornadas Uruguayas de Buiatría; Centro Médico Veterinario de Paysandú/Sociedad Uruguaya de Buiatría: Paysandú, Uruguay, 2016; pp. 34–41. [Google Scholar]
- Stamey, J.A.; Wallace, R.L.; Grinstead, K.R.; Bremmer, D.R.; Drackley, J.K. Influence of plane of nutrition on growth of dairy calves. J. Dairy Sci. 2006, 89 (Suppl. S1), 50. [Google Scholar]
- Lagger, C. Protein nutrition in early life of calves. In Recent Advances in Animal Nutrition; Beynen, A.C., Ed.; Nottingham University Press: Nottingham, UK, 2010; pp. 81–106. [Google Scholar]
- Brown, R.E.; Mathews, K.H.; Patterson, H.H. Accelerated calf rearing program: Effects on growth and profitability. J. Dairy Sci. 2005, 88, 11–18. [Google Scholar]
- Brown, R.E.; Mathews, K.H.; Patterson, H.H. Accelerated calf rearing program: Effects on mammary development. J. Dairy Sci. 2005, 88, 19–27. [Google Scholar]
- de Trinidad, A.B. Effect of milk replacer volume on solid feed intake, growth, and weaning in dairy calves. J. Dairy Sci. 2014, 97, 3728–3734. [Google Scholar]
- Davis, C.L.; Drackley, J.K. The Development, Nutrition, and Management of the Young Calf; Iowa State University Press: Ames, IA, USA, 1998. [Google Scholar]
- Silper, B.; Lana, A.; Carvalho, A.; Ferreira, C.; Franzoni, A.; Lima, J.; Saturnino, H.; Reis, R.; Coelho, S. Effects of milk replacer feeding strategies on performance, ruminal development, and metabolism of dairy calves. J. Dairy Sci. 2014, 97, 1016–1025. [Google Scholar] [CrossRef] [Green Version]
- Nocek, J.E.; Kesler, E.M. Growth and Rumen Characteristics of Holstein Steers Fed Pelleted or Conventional Diets1. J. Dairy Sci. 1980, 63, 249–254. [Google Scholar] [CrossRef]
- Hinders, R.G.; Owen, F.G. Relation of ruminal parakeratosis development to volatile fatty acid absorption. J. Dairy Sci. 1965, 48, 1069–1073. [Google Scholar] [CrossRef]
- von Engelhardt, W.; Hauffe, R. Funktionen des Blattermägens bei kleinen Hauswiederkäuern. IV. Resorption und Sekretion von Elektrolyten. J. Vet. Med. Ser. A 1975, 22, 363–375. [Google Scholar]
- Holtenius, K.; Björnhag, G. The significance of water absorption and fibre digestion in the omasum of sheep, goats and cattle. Comp. Biochem. Physiol. Part A 1989, 94, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Clauss, M.; Hofmann, R.R.; Hummel, J.; Adamczewski, J.; Nygren, K.; Pitra, C.; Streich, W.J.; Reese, S. Macroscopic anatomy of the omasum of free-ranging moose (Alces alces) and muskoxen (Ovibos moschatus) and a comparison of the omasal laminal surface area in 34 ruminant species. J. Zool. 2006, 270, 346–358. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kitamura, N.; Yamada, J.; Andren, A.; Yamashita, T. Morphological study of the surface structure of the omasal laminae in cattle, sheep and goats. Anat. Histol. Embryol. 1994, 23, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.F.; Kühnel, W.; Vives, P.; Wedel, T. Functional morphology of unguiculiform papillae of the reticular groove in the ruminant stomach. Ann. Anat. Anat. Anz. 2009, 191, 469–476. [Google Scholar] [CrossRef]
- Hofmann, R.R.; McKnight, M.H.; Skinner, J.D. On structural characteristics and morphophysiological adaptation of the springbok (Antidorcas marsupialis) digestive system. Trans. R. Soc. S. Afr. 1995, 50, 125–142. [Google Scholar] [CrossRef]
- Pérez, W.; Erdoğan, S.; Ungerfeld, R. Anatomical study of the gastrointestinal tract in free-living Axis deer (Axis axis). Anat. Histol. Embryol. 2015, 44, 43–49. [Google Scholar] [CrossRef]
- Sauer, C.; Bertelsen, M.F.; Lund, P.; Weisbjerg, M.R.; Clauss, M. Quantitative macroscopic anatomy of the giraffe (Giraffa camelopardalis) digestive tract. Anat. Histol. Embryol. 2016, 45, 338–349. [Google Scholar] [CrossRef] [Green Version]
- The International Committee on Veterinary Gross Anatomical Nomenclature, 6th ed.; Editorial Committee: Hannover, Germany; Columbia, MO, USA; Ghent, Belgium; Sapporo, Japan, 2017; Available online: https://www.studocu.com/en-gb/document/edinburgh-napier-university/veterinary-practice-management-for-veterinary-nurses/nav-6th-edition-2017-complete-version/24021516 (accessed on 8 May 2023).
- Clauss, M.; Hofmann, R.R. The Digestive System of Ruminants, and Peculiarities of (Wild) Cattle. Ecology, Evolution and Behaviour of Wild Cattle: Implications for Conservation; Cambridge University Press: Cambridge, UK, 2014; pp. 57–62. [Google Scholar]
- Clauss, M.; Frei, S.; Ortmann, S.; Hatt, J.M. Omasum function in ruminants—What we know and what we need to know. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1179–1194. [Google Scholar]
- Agungpriyono, S.; Yamamoto, Y.; Kitamura, N.; Yamada, J.; Sigit, K.; Yamashita, T. Morphological study on the stomach of the lesser mouse deer (Tragulus javanicus) with special reference to the internal surface. J. Vet. Med. Sci. 1992, 54, 1063–1069. [Google Scholar] [CrossRef] [Green Version]
- Clauss, M.; Rössner, G.E. Old world ruminant morphophysiology, life history, and fossil record: Exploring key innovations of a diversification sequence. Ann. Zool. Fenn. 2014, 51, 80–95. [Google Scholar] [CrossRef] [Green Version]
- Garrod, A.H. Notes on the visceral anatomy and osteology of the ruminants, with a suggestion regarding a method of expressing the relations of species by means of formulae. Proc. Zool. Soc. Lond. 1877, 1877, 2–18. [Google Scholar]
- Langer, P. Vergleichend-anatomische Untersuchungen am Magen der Artiodactyla (Owen, 1848). II. Untersuchungen am Magen der Tylopoda und Ruminantia. Gegenbaurs Morphol. Jahrb. 1973, 119, 633–695. [Google Scholar] [PubMed]
- Hofmann, R.R. Zur Topographie und Morphologie des Wiederkäuermagens im Hinblick auf seine Funktion (nach vergleichenden Untersuchungen an Material ostafrikanischer Wildarten). Zent. Für Eterinärmedizin 1969, 10, 1–180. [Google Scholar]
- Hofmann, R.R. The Ruminant Stomach; East African Literature Bureau: Nairobi, Kenya, 1973. [Google Scholar]
- Hofmann, R.R. Morphophysiological evolutionary adaptations of the ruminant digestive system. In Aspects of Digestive Physiology in Ruminants; Dobson, A., Dobson, M.J., Eds.; Cornell University Press: Ithaca, NY, USA, 1988; pp. 1–20. [Google Scholar]
- Langer, P. The Mammalian Herbivore Stomach; Gustav Fischer Verlag: Stuttgart, Germany, 1988. [Google Scholar]
- Stobo, I.J.F.; Roy, J.H.B.; Gaston, H.J. Rumen development in the calf. 2. The effect of diets containing different proportions of concentrates to hay on digestive efficiency. Br. J. Nutr. 1966, 20, 189–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauwers, H. Morfologische bijdrage tot de kennis von her resorberend vermogen van rundercoormagen. Meded. Van de Fac. Diergeneeskd. Rijksuniv. Gent 1973, 17, 1–261. [Google Scholar]
- Hamada, T.; Maeda, S.; Kameoka, K. Factors influencing growth of rumen, liver, and other organs in kids weaned from milk replacers to solid foods. J. Dairy Sci. 1975, 59, 1110–1118. [Google Scholar] [CrossRef]
- Bailey, C.B. Growth of digestive organs and their contents in holstein steers: Relation to body weight and diet. Can. J. Anim. Sci. 1986, 66, 653–661. [Google Scholar] [CrossRef]
- Johnson, C.L.; Johnson, D.E.; Rumpler, W.V. Source and level of alimentation effects on visceral organ mass of fat steers. In Energy Metabolism of Farm Animals; Moe, P.W., Tyrrell, H.F., Reynolds, P.J., Eds.; Rowman and Littlefiled: Totowa, NJ, USA, 1987; pp. 50–53. [Google Scholar]
- McLeod, K.R.; Baldwin, R.L. Effects of diet forage: Concentrate ratio and metabolizable energy intake on visceral organ growth and in vitro oxidative capacity of gut tissues in sheep. J. Anim. Sci. 2000, 78, 760–770. [Google Scholar] [CrossRef]
- Becker, R.B.; Marshall, S.P.; Arnold, P.D. Anatomy, development, and functions of the bovine omasum. J. Dairy Sci. 1963, 46, 835–839. [Google Scholar] [CrossRef]
- Tahas, S.A.; Martin Jurado, O.; Hammer, S.; Arif, A.; Reese, S.; Hatt, J.M.; Clauss, M. Gross measurements of the digestive tract and visceral organs of addax antelope (Addax nasomaculatus) following a concentrate or forage feeding regime. Anat. Histol. Embryol. 2017, 46, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Fluharty, F.L.; McClure, K.E.; Solomon, M.B.; Clevenger, D.D.; Lowe, G.D. Energy source and ionophore supplementation effects on lamb growth, carcass characteristics, visceral organ mass, diet digestibility, and nitrogen metabolism. J. Anim. Sci. 1999, 77, 816–823. [Google Scholar] [CrossRef] [PubMed]
- McClure, K.E.; Solomont, M.B.; Loerch, S.C. Body weight and tissue gain in lambs fed an all-concentrate diet and implanted with trenbolone acetate or grazed on alfalfa. J. Anim. Sci. 2000, 78, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Mathiesen, S.D.; Haga, Ø.E.; Kaino, T.; Tyler, N.J.C. Diet composition, rumen papillation and maintenance of carcass mass in female Norwegian reindeer (Rangifer tarandus tarandus) in winter. J. Zool. 2000, 251, 129–138. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | |
|---|---|
| Crude protein (%) | 25.0 |
| Fat (%) | 20.0 |
| ME (Kcal) | 1.6 |
| Crude fiber (%) | 0.3 |
| Ether extracts (%) | 20.0 |
| Lactose (%) | 44.0 |
| Ash (%) | 4.5 |
| Calcium (%) | 1.3 |
| Phosphorous (%) | 0.6 |
| Sodium (%) | 0.4 |
| Chlorine (%) | 0.5 |
| Inorg. copper (ppm) | 11.0 |
| Inorg. zinc (ppm) | 44.0 |
| Ferrus (ppm) | 111.0 |
| Vit A (IU/Kg) | 27,000.0 |
| Vit D3 (IU/Kg) | 5300.0 |
| Vit E (IU/Kg) | 50,000.0 |
| Ionophore (ppm) | 100,000.0 |
| Total lysine (%DM) | 2.7 |
| Total methionine (%DM) | 0.9 |
| Parameters | |
|---|---|
| Humidity (%) | 12.1 |
| Protein (%) | 18.1 |
| Crude fiber (%) | 3.3 |
| ADF (%) | 4.0 |
| NDF (%) | 15.0 |
| Ether extracts (%) | 3.4 |
| Ash (%) | 4.9 |
| NEl (Mcal/Kg DM) | 1.9 |
| Aflatoxins (B1, B2, G1, G2) (ppb) | <5 |
| DON (ppb) | <500 |
| Zearalenone (ppb) | <50 |
| Forage Feed Group | Concentrated Feed Group | p (t-Test) | |
|---|---|---|---|
| Body weight (Kg) | 87.7 ± 7.6 | 91.9 ± 8.7 | Ns |
| Full OM weight (g) | 1072.0 ± 236.1 | 954.8 ± 255.3 | Ns |
| Empty OM weight (g) | 572.8 ± 106.2 | 542.3 ± 120.2 | Ns |
| Omasal wall weight without laminae | 225.5 ± 34.3 | 207.1 ± 32.0 | Ns |
| OM curvature (mm) | 32.2 ± 6.3 | 32.3 ± 3.3 | Ns |
| OM height (mm) | 12.9 ± 2.4 | 12.7 ± 2.5 | Ns |
| OM length (mm) | 14.1 ± 2.6 | 13.3 ± 1.4 | Ns |
| Number of 1st-order laminae | 22.2 ± 3.0 | 21.4 ± 3.0 | Ns |
| Number of 2nd-order laminae | 26.0 ± 3.7 | 23.8 ± 4.0 | Ns |
| Number of 3rd-order laminae | 37.8 ± 3.6 | 37.0 ± 4.1 | 0.04 |
| Total number of laminae | 86.0 ± 5.4 | 77.0 ± 7.4 | Ns |
| Forage Feed Group | Concentrated Feed Group | p (t-Test) | |
|---|---|---|---|
| ROO | 1073.2 ± 255.0 | 797.8 ± 214.7 | 0.05 |
| Middle part | 458.8 ± 128.9 | 528.5 ± 38.0 | Ns |
| OABO | 239.6 ± 41.2 | 343.0 ± 49.6 | 0.00142 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez, W.; Duro, S.; Gündemir, O. Anatomical Differences in the Omasum of Weaning Calves Fed with Different Diets. Anatomia 2023, 2, 176-188. https://doi.org/10.3390/anatomia2020016
Pérez W, Duro S, Gündemir O. Anatomical Differences in the Omasum of Weaning Calves Fed with Different Diets. Anatomia. 2023; 2(2):176-188. https://doi.org/10.3390/anatomia2020016
Chicago/Turabian StylePérez, William, Sokol Duro, and Ozan Gündemir. 2023. "Anatomical Differences in the Omasum of Weaning Calves Fed with Different Diets" Anatomia 2, no. 2: 176-188. https://doi.org/10.3390/anatomia2020016