
Seed Morpho-Anatomy and Germination Enhancement of the Australian Native Species Lomandra longifolia Labill. and L. hystrix (R.Br.) L.R. Fraser & Vickery
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Lots
2.2. Seed Fill, Weight and Morpho-Anatomy
2.3. Germination Stimulation Using Mechanical and Chemical Treatments
2.4. Data Collection and Analysis
3. Results
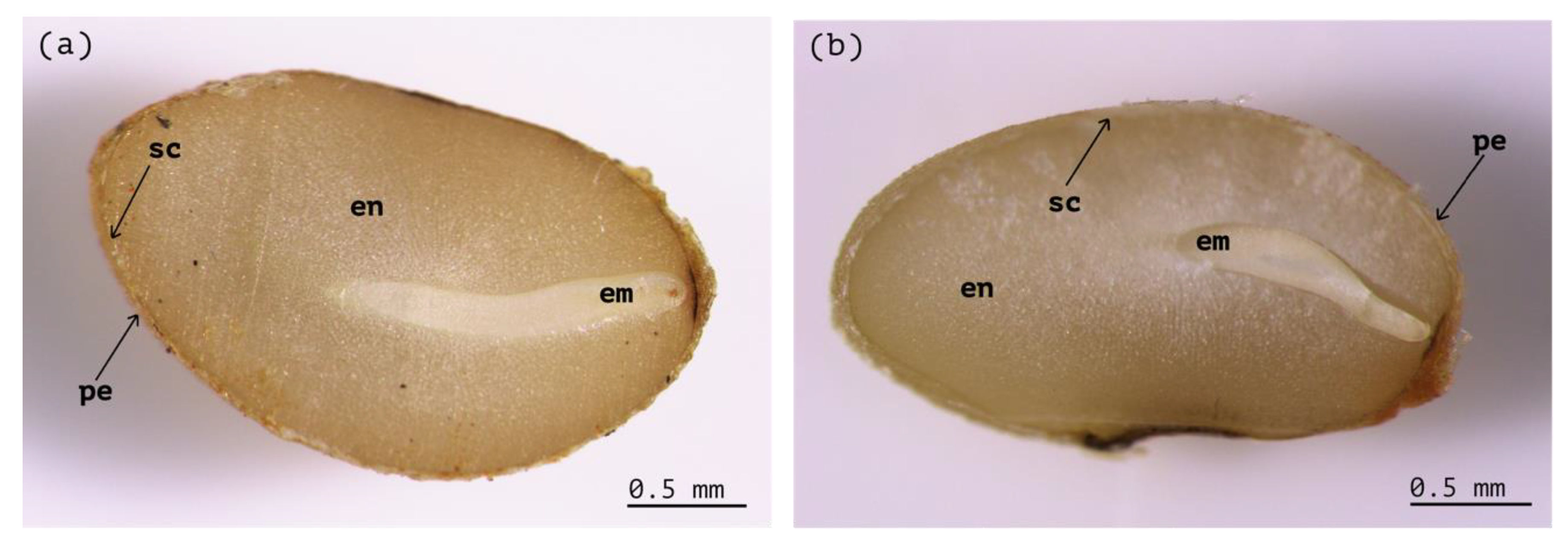
3.1. Seed Fill and Seed Morpho-Anatomy
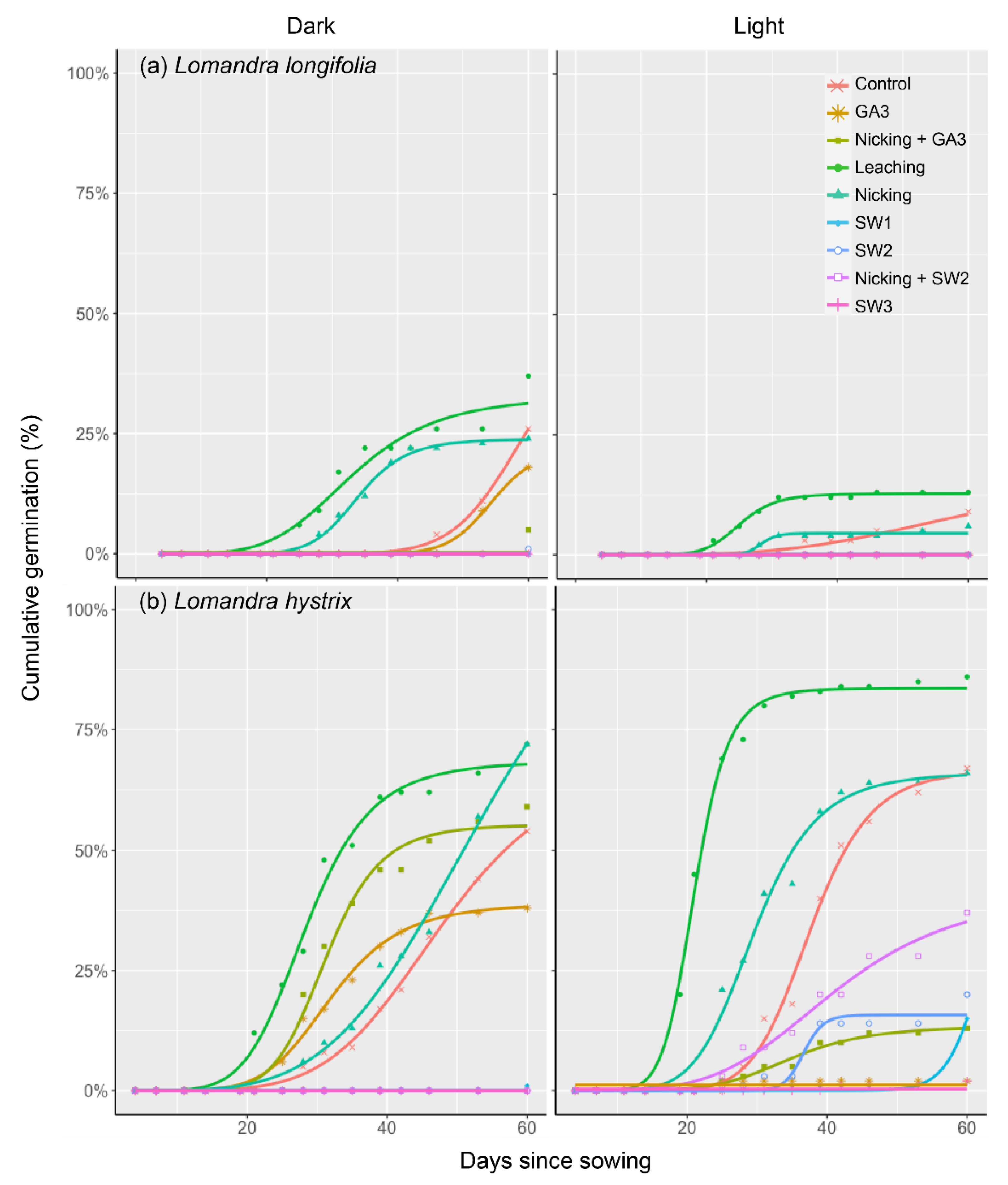
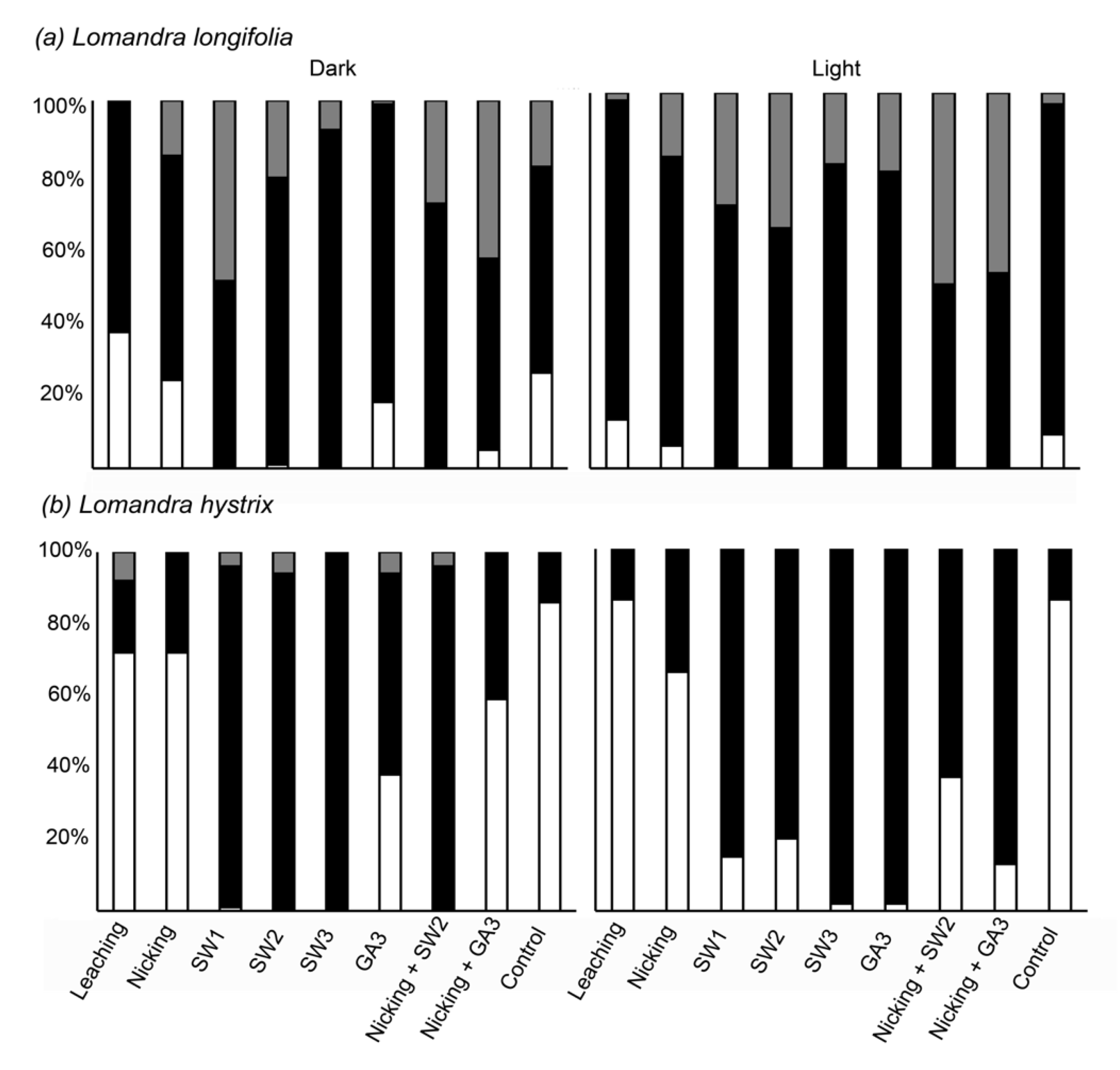
3.2. Germination Enhancement Using Mechanical and Chemical Treatments
4. Discussion
4.1. Seed Morpho-Anatomy
4.2. Seed Germination Ecology and Enhancement
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seed Information Database (SID), Royal Botanical Gardens Kew. Available online: http://data.kew.org/sid/ (accessed on 14 September 2018).
- Plummer, J.; Crawford, A.; Taylor, S. Germination of Lomandra sonderi (Dasypogonaceae) promoted by pericarp removal and chemical-stimulation of the embryo. Aust. J. Bot. 1995, 43, 223–230. [Google Scholar] [CrossRef]
- Lee, A.; Macfarlane, T. Lomandra. FI Aust. 1986, 46, 100–141. [Google Scholar]
- Bonney, N.; Miles, A. What Seed Is That? N. Bonney: Beverley, Australia, 1994. [Google Scholar]
- Ahmad, N.M.; Martin, P.M.; Vella, J.M. Embryology of the dioecious Australian endemic Lomandra longifolia (Lomandraceae). Aust. J. Bot. 2009, 56, 651–665. [Google Scholar] [CrossRef]
- Ahmad, N.M.; Martin, P.M.; Vella, J.M. Clonal propagation of Lomandra longifolia by somatic embryogenesis. Sci. Hortic. 2014, 180, 102–110. [Google Scholar] [CrossRef]
- Merritt, D.; Turner, S.; Clarke, S.; Dixon, K. Seed dormancy and germination stimulation syndromes for Australian temperate species. Aust. J. Bot. 2007, 55, 336–344. [Google Scholar] [CrossRef]
- Erickson, T.; Barrett, R.; Merritt, D.; Dixon, K. Pilbara Seed Atlas and Field Guide: Plant Restoration in Australia’s Arid Northwest; CSIRO Publishing: Clayton, Australia, 2016. [Google Scholar]
- Baskin, C.; Baskin, J. Types of seeds and kinds of seed dormancy. In Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Elsevier: Amsterdam, The Netherlands, 2014; pp. 37–77. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: London, UK, 2014. [Google Scholar]
- Bonney, N. Understanding botanical pathlines for improved germination of native plant seed. In Proceedings of the Fourth Australian Workshop on Native Seed Biology for Revegetation, Mildura, Australia, 3–4 September 2001; pp. 105–111. [Google Scholar]
- Gibson-Roy, P.; Delpratt, J.; Moore, G. Restoring the Victorian Western (Basalt) Plains grassland. 1. Laboratory trials of viability and germination, and the implications for direct seeding. Ecol. Manag. Restor. 2007, 8, 114–122. [Google Scholar] [CrossRef]
- Grant, C.; Campbell, C.; Charnock, N. Selection of species suitable for derelict mine site rehabilitation in New South Wales, Australia. Water Air Soil Pollut. 2002, 139, 215–235. [Google Scholar] [CrossRef]
- Ruiz-Talonia, L.; Whalley, R.; Gross, C.; Carr, D.; Reid, N. Overcoming limitations to propagation from seed of 40 Australian species important for restoration. New For. 2022, 1–20. [Google Scholar] [CrossRef]
- Alvarado, V.; Nonogaki, H.; Bradford, K. Expression of endo-β-mannanase and SNF-related protein kinase genes in true potato seeds in relation to dormancy, gibberellin and abscisic acid. In Dormancy in Plants: From Whole Plant Behaviour to Cellular Control; Viemont, J., Crabbe, J., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 347–364. [Google Scholar]
- Roche, S. Smoke-a new process for germinating Australian plants. Aust. Hortic. 1994, 92, 46–47. [Google Scholar]
- Dixon, K.W.; Roche, S.; Pate, J.S. The promotive effect of smoke derived from burnt native vegetation on seed germination of Western Australian plants. Oecologia 1995, 101, 185–192. [Google Scholar] [CrossRef]
- Baskin, C.C.; Thompson, K.; Baskin, J.M. Mistakes in germination ecology and how to avoid them. Seed Sci. Res. 2006, 16, 165–168. [Google Scholar] [CrossRef]
- Turner, S.; Merritt, D. Seed germination and dormancy. In Plant Germplasm Conservation in Australia: Strategies and Guidelines for Developing, Managing and Utilising Ex Situ Collections; Offord, C.A., Meagher, P.F., Eds.; Australian Network for Plant Conservation Inc.: Canberra, Australia, 2009. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. A revision of Martin’s seed classification system, with particular reference to his dwarf-seed type. Seed Sci. Res. 2007, 17, 11–20. [Google Scholar] [CrossRef]
- Martin, A. The comparative internal morphology of seeds. Am. Midl. Nat. 1946, 36, 513–660. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Underdeveloped embryos in dwarf seeds and implications for assignment to dormancy class. Seed Sci. Res. 2005, 15, 357. [Google Scholar] [CrossRef]
- Commander, L.; Merritt, D.; Rokich, D.; Dixon, K. Seed biology of Australian arid zone species: Germination of 18 species used for rehabilitation. J. Arid Environ. 2009, 73, 617–625. [Google Scholar] [CrossRef]
- Ooi, M.K. Dormancy classification and potential dormancy-breaking cues for shrub species from fire-prone south-eastern Australia. In Seeds: Biology, Development and Ecology; Adkins, S., Ashmore, S., Navie, S.C., Eds.; CAB International: Wallingford, UK, 2007; pp. 205–216. [Google Scholar]
- Merrit, D. Seed storage and testing. In Australian Seeds: A Guide to Their Collection, Identification and Biology; Sweedman, L., Merritt, D., Eds.; CSIRO Publishing: Collingwood, Australia, 2006; pp. 53–60. [Google Scholar]
- Cochrane, A.; Kelly, A.; Brown, K.; Cunneen, S. Relationships between seed germination requirements and ecophysiological characteristics aid the recovery of threatened native plant species in Western Australia. Ecol. Manag. Restor. 2002, 3, 47–60. [Google Scholar] [CrossRef]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Pedrini, S.; Lewandrowski, W.; Stevens, J.C.; Dixon, K.W. Optimising seed processing techniques to improve germination and sowability of native grasses for ecological restoration. Plant Biol. 2018, 21, 415–424. [Google Scholar] [CrossRef]
- Maguire, J.D. Speed of Germination—Aid in selection and evaluation for seedling emergence and vigor 1. Crop Sci. 1962, 2, 176–177. [Google Scholar] [CrossRef]
- Shaw, N.; Barak, R.S.; Campbell, R.E.; Kirmer, A.; Pedrini, S.; Dixon, K.; Frischie, S. Seed use in the field: Delivering seeds for restoration success. Restor. Ecol. 2020, 28, S276–S285. [Google Scholar] [CrossRef]
- Erickson, T.E.; Muñoz-Rojas, M.; Kildisheva, O.A.; Stokes, B.A.; White, S.A.; Heyes, J.L.; Dalziell, E.L.; Lewandrowski, W.; James, J.J.; Madsen, M.D. Benefits of adopting seed-based technologies for rehabilitation in the mining sector: A Pilbara perspective. Aust. J. Bot. 2017, 65, 646–660. [Google Scholar] [CrossRef]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. The control of seed dormancy and germination by temperature, light and nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Turner, S.R.; Lewandrowski, W.; Elliott, C.P.; Merino-Martín, L.; Miller, B.P.; Stevens, J.C.; Erickson, T.E.; Merritt, D.J. Seed ecology informs restoration approaches for threatened species in water-limited environments: A case study on the short-range Banded Ironstone endemic Ricinocarpos brevis (Euphorbiaceae). Aust. J. Bot. 2017, 65, 661–677. [Google Scholar] [CrossRef]
- Adkins, S.W.; Bellairs, S.M.; Preston, C.; Thompson, L.; Farley, G. Identifying dormancy mechanisms of Australian native plant species. In Proceedings of the Fourth Australian Workshop in Native Seed Biology for Revegetation, Mildura, Australia, 3–4 September 2002; pp. 61–70. [Google Scholar]
- Kildisheva, O.A.; Dixon, K.W.; Silveira, F.A.; Chapman, T.; Di Sacco, A.; Mondoni, A.; Turner, S.R.; Cross, A.T. Dormancy and germination: Making every seed count in restoration. Restor. Ecol. 2020, 28, S256–S265. [Google Scholar] [CrossRef]
- Turner, S.; Merritt, D.; Ridley, E.; Commander, L.; Baskin, J.; Baskin, C.; Dixon, K. Ecophysiology of seed dormancy in the Australian endemic species Acanthocarpus preissii (Dasypogonaceae). Ann. Bot. 2006, 98, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, G.; Daws, M.; Steadman, K.; Adkins, S. Mimicking a semi-arid tropical environment achieves dormancy alleviation for seeds of Australian native Goodeniaceae and Asteraceae. Ann. Bot. 2008, 101, 701–708. [Google Scholar] [CrossRef]
- Erickson, T.E.; Shackelford, N.; Dixon, K.W.; Turner, S.R.; Merritt, D.J. Overcoming physiological dormancy in seeds of Triodia (Poaceae) to improve restoration in the arid zone. Restor. Ecol. 2016, 24, S64–S76. [Google Scholar] [CrossRef]
- Cochrane, A.; Probert, R. Temperature and dormancy-breaking treatments: Germination of endemic and geographically restricted herbaceous perennials. Aust. J. Bot. 2006, 54, 349–356. [Google Scholar] [CrossRef]
- Merritt, D.; Rokich, D. Seed biology and ecology. In Australian Seeds: A Guide to Their Collection, Identification and Biology; Sweedman, L., Merritt, D., Eds.; CSIRO Publishing: Collingwood, Australia, 2006; pp. 19–24. [Google Scholar]
- Vening, G.S.; Guja, L.K.; Spooner, P.G.; Price, J.N. Seed dormancy and germination of three grassy woodland forbs required for diverse restoration. Aust. J. Bot. 2017, 65, 625–637. [Google Scholar] [CrossRef]
- Adkins, S.; Peters, N. Smoke derived from burnt vegetation stimulates germination of arable weeds. Seed Sci. Res. 2001, 11, 213–222. [Google Scholar]
- Light, M.; Gardner, M.; Jäger, A.; Van Staden, J. Dual regulation of seed germination by smoke solutions. Plant Growth Regul. 2002, 37, 135–141. [Google Scholar] [CrossRef]
- Thompson, K.; Ooi, M.K. To germinate or not to germinate: More than just a question of dormancy. Seed Sci. Res. 2010, 20, 209–211. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Type |
|---|---|
| Leaching | Mechanical |
| Nicking | Mechanical |
| SW1: Smoke water (50 mL L−1) | Chemical |
| SW2: Smoke water (100 mL L−1) | Chemical |
| SW3: Smoke water (200 mL L−1) | Chemical |
| GA3: gibberellic acid (289 μM) | Chemical |
| Nicking + SW2 | Combination |
| Nicking + GA3 | Combination |
| Tissue | Lomandra longifolia | Lomandra hystrix |
|---|---|---|
| E:S ratio | 0.4 ± 0.03 | 0.4 ± 0.04 |
| Seed length (mm) | 3.9 ± 0.1 | 4.9 ± 0.8 |
| Seed width (mm) | 2.3 ± 0.2 | 2.5 ± 0.4 |
| Seed area (mm2) | 7.5 ± 0.6 | 11.8 ± 5.2 |
| Embryo length (mm) | 1.4 ± 0.1 | 2.2 ± 0.5 |
| Embryo area (mm2) | 0.3 ± 0.0 | 0.6 ± 0.1 |
| 100 seed weight (mg) | 860 ± 20 | 900 ± 20 |
| Treatment | Lomandra longifolia | Lomandra hystrix | ||
|---|---|---|---|---|
| Light | Dark | Light | Dark | |
| Leaching | 0.5 ± 0.1 | 1.0 ± 0.1 | 3.7 ± 0.1 | 2.4 ± 0.2 |
| Nicking | 0.2 ± 0.1 | 0.7 ± 0.0 | 2.2 ± 0.3 | 1.6 ± 0.4 |
| SW1 | - | - | 0.3 ± 0.0 | - |
| SW2 | - | - | 0.5 ± 0.1 | - |
| SW3 | - | - | - | - |
| GA3 | - | 0.3 ± 0.1 | - | 1.2 ± 0.2 |
| SW2 + nicking | - | - | 0.9 ± 0.3 | - |
| GA3 + nicking | - | 0.1 ± 0.0 | 0.4 ± 0.1 | 1.7 ± 0.1 |
| Control | 0.2 ± 0.1 | 0.5 ± 0.1 | 1.7 ± 0.2 | 1.2 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beveridge, F.C.; Williams, A.; Cave, R.; Kalaipandian, S.; Adkins, S.W. Seed Morpho-Anatomy and Germination Enhancement of the Australian Native Species Lomandra longifolia Labill. and L. hystrix (R.Br.) L.R. Fraser & Vickery. Appl. Biosci. 2023, 2, 222-235. https://doi.org/10.3390/applbiosci2020016
Beveridge FC, Williams A, Cave R, Kalaipandian S, Adkins SW. Seed Morpho-Anatomy and Germination Enhancement of the Australian Native Species Lomandra longifolia Labill. and L. hystrix (R.Br.) L.R. Fraser & Vickery. Applied Biosciences. 2023; 2(2):222-235. https://doi.org/10.3390/applbiosci2020016
Chicago/Turabian StyleBeveridge, Fernanda Caro, Alwyn Williams, Robyn Cave, Sundaravelpandian Kalaipandian, and Steve W. Adkins. 2023. "Seed Morpho-Anatomy and Germination Enhancement of the Australian Native Species Lomandra longifolia Labill. and L. hystrix (R.Br.) L.R. Fraser & Vickery" Applied Biosciences 2, no. 2: 222-235. https://doi.org/10.3390/applbiosci2020016