
Gene Drive: Past, Present and Future Roads to Vertebrate Biocontrol

1
Elizabeth Macarthur Agricultural Institute, NSW Department of Primary Industries, Menangle, NSW 2568, Australia
2
The Roslin Institute and Royal (Dick) School of Veterinary Studies, University of Edinburgh, Midlothian EH25 9RG, UK
*
Author to whom correspondence should be addressed.
Appl. Biosci. 2023, 2(1), 52-70; https://doi.org/10.3390/applbiosci2010006
Submission received: 8 November 2022
/
Revised: 31 January 2023
/
Accepted: 1 February 2023
/
Published: 13 February 2023
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Scientists have long sought a technology to humanely control populations of damaging invasive pests in a species-specific manner. Gene drive technology could see this become a reality. This review charts the twists and turns on the road to developing gene drives in vertebrates. We focus on rodents, as these will likely be the first targets, and trace the journey from the early understanding of selfish genetic elements to engineering gene drives in mice; before discussing future research focuses and the crucial role that public perception and governance will play in the application of this technology. The realisation of robust gene drive strategies in vertebrate pests has the potential to revolutionise biocontrol.
1. Introduction
Invasive vertebrate species impact our environment, economy and society. Although their spread can have beneficial aspects, invasive species can disrupt ecosystems, reduce biodiversity and threaten livelihoods. Damaging invasive species are classified as invasive pests, and examples include: rabbits (Oryctolagus cuniculus) and feral cats (Felis catus) in Australia, American mink (Neovison vison) and grey squirrel (Sciurus carolinensis) in the United Kingdom, Burmese pythons (Python bivittatus) and wild boar (Sus scrofa) in the United States of America (US), and the omnipresent infestation of rats and mice around the globe [1,2,3]. The burden of invasive pests has been worsened by the increases in global trade, travel and climate change [4]. Current pest management tools for vertebrate pests are crude, costly, inhumane and often inadequate to cope with the scale of the problem. The ability to humanely control damaging invasive pests has long been a goal of scientists.
Of the numerous invasive vertebrate pests, mice and rats (Muridae) are recognised as some of the most prolific and will be the primary focus of this review, although the techniques described could be adapted to most vertebrate pests. At an economic level, the worldwide losses caused by mice and rats are enormous. It has been conservatively estimated that in Asia, each year, rats consume over 30 million tons of rice [5]. That is enough rice to feed 180 million people per year. In outbreak years, domestic damage to Australian wheat reduces the country’s total agricultural production from 3 to 4% [6]. In the US, it is estimated that introduced rats cost the economy more than $27 billion per year. In addition to agricultural impacts, in many countries and oceanic islands, invasive rodents are also a major cause of reduction in bird populations due to their predation of native chicks and bird eggs [7,8]. In response, mitigation and eradication strategies are required.
Current efforts to eradicate rats and mice include traps, poisons and biological controls, such as the introduction of predators or diseases. The application of rodenticides can be expensive due to costs associated with regulation compliance, dispersal methods, size of the treated area, and the toxicant itself [9]. Mechanical traps are more humane than rodenticides as they have less impact on non-target animals, humans or overall ecosystem health. However, placing traps and collecting the caught animals is labour-intensive and usually inadequate to eradicate a rodent population without additional methods [10]. Scientists may now be able to utilise the growing body of knowledge on selfish genetic elements and recent advances in genome engineering technologies to develop species-specific, humane and cost-effective genetic pest management tools.
2. Selfish Genetic Elements
First revealed in 1866 by a monk named Gregor Mendel, the conventional rules of inheritance, also known as Mendelian inheritance, determine that offspring have a 50% chance of inheriting an allele from one of their parents. As only half the offspring inherit the allele, the frequency of that allele in future generations will be comparable to the frequency of that allele in the parents’ generation. Selfish genetic elements are described as an exception to the conventional rules of inheritance. With selfish genetic elements, offspring have more than a 50% chance of inheriting a genetic element from a parent, and therefore a specific genotype will increase in the population over time. Such an element is said to ‘drive’ within a population [11,12].
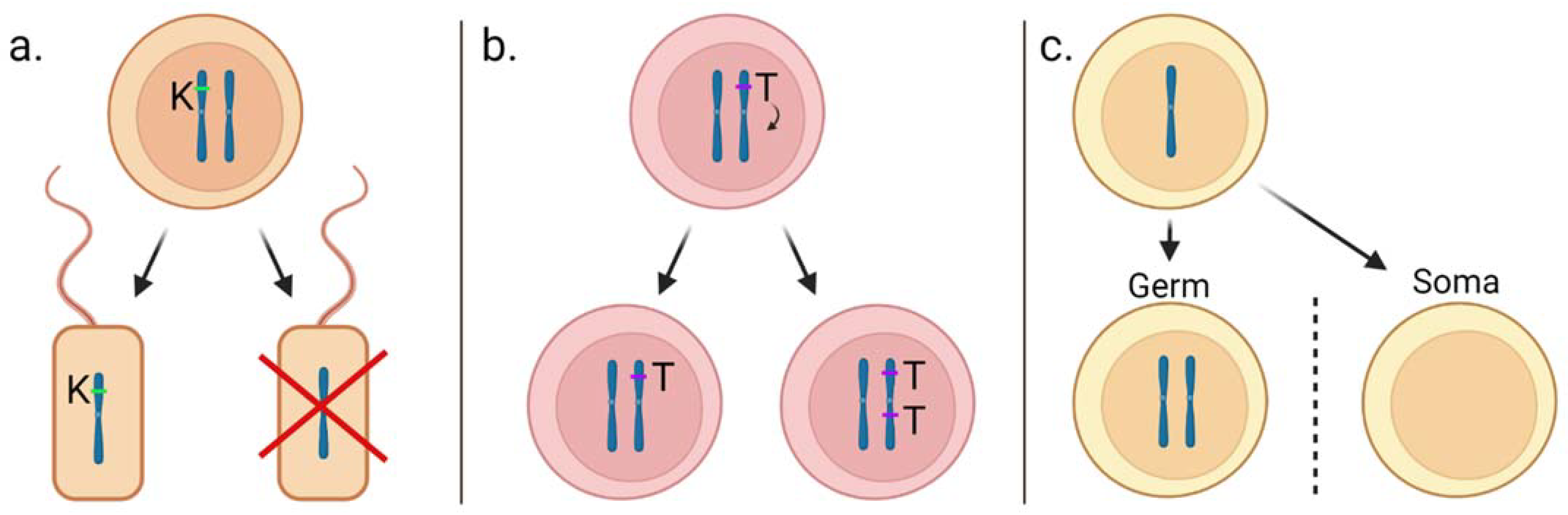
Scientists have known about selfish genetic elements that violate Mendel’s rules since the late 1880s. At a molecular level, they are sequences of DNA, such as genes or their fragments, for which inheritance is biased in their favour [11]. These elements achieve drive through one or more of three key mechanisms (Figure 1). 1. Interference: with this strategy, a gene gains an advantage by disrupting the transmission of an alternative gene. An example of a selfish element that operates by interference is the t-haplotype in mice, described in the meiotic drive section of this review. 2. Over-replication: these elements bias their transmission to the next generation by being replicated more than other genes in an animal. Transposable elements are an example of this mechanism. 3. Gonotaxis: where genetic elements bias movement towards the germline. Some B-chromosomes, which are accessory chromosomes that are not essential for the life of a species, act selfishly by replicating themselves and moving to the germline and avoiding the somatic cells [11].
One important particularity of selfish genetic elements is that they do not need to contribute to the reproductive success or fitness of the host organism to drive successfully [11]. In addition to the examples highlighted in Figure 1, homing endonuclease genes and meiotic drives represent two important examples of natural selfish genetic elements, having evolved several times in different taxa. Due to the significance of meiotic drives and homing endonuclease genes in the development of engineered selfish genetic elements, a brief description of these elements is provided below.
2.1. Homing Endonuclease Genes
Homing endonuclease genes (HEGs) achieve drive through an over-replication mechanism. These genes are situated on a chromosome within a specific DNA sequence that they recognise and cut. HEGs encode an endonuclease that cuts the recognition sequence on the chromosome that is homologous to the original chromosome containing the HEG. After the recognition sequence is cut, homologous recombination is used to copy the HEG into the cut in the homologous sequence. When this process occurs in the germ cells, the proportion of eggs or sperm that contain the HEG is greater than 50%, and therefore, the HEG can drive itself through the population [13]. HEGs are present in eukaryotes, archaea and bacteria, where their recognition sequences are found at low frequencies in the genome [14]. Well-characterised examples of HEGs include the fungal homing endonuclease I-PpoI from Physarum polycephaleum and I-SceI from Saccharomyces cerevisiae, which target 15 bp and 18 bp DNA motifs, respectively [15,16].
2.2. Meiotic Drive
The meiotic drive is a method by which a selfish genetic element can achieve drive through an interference mechanism [17]. The most studied natural meiotic drive in vertebrates is the t-haplotype of the house mouse (Mus musculus) [18,19]. The t-haplotype consists of a series of linked, independent T-complex distorter genes and a T-complex responder gene that are inherited together. When present in the heterozygous (Tt) state in males, the wild-type sperm show motility deficiencies and are functionally inactive, so more than 90% of the eggs are fertilised by t-haplotype harbouring sperm. This requires the combined action of the distorter genes, which attacks gametes, and a responder gene, which protects gametes carrying the t-haplotype. This action leads to morphological defects in spermatozoa that do not carry a t-haplotype due to excessive activation of the SMOK gene [20,21,22].
3. Harnessing Evolution
Although the scientific community has recognised selfish genetic elements since the late 19th century, the idea of using these elements as a means to control natural populations did not surface until the mid-20th century [11]. In 1960, Craig, Hickey & Vandehey suggested using a breeding program in which a “male-producing factor”, naturally present in some male mosquitoes, could be harnessed to control mosquito populations. Hickey and Craig [23] went on to identify the genomic region responsible for this phenomenon. Their logic behind this control strategy was that when male mosquitoes with this male-producing factor breed, most of their offspring then develop as males. It was, therefore, postulated that environmental releases of mosquitoes carrying this male-producing factor could potentially “reduce the number of females below the level required for efficient disease transmission” [24].
In the 1960s, Craig, Hickey and the other early pioneers did not yet have the molecular tools to engineer animals harbouring desirable genes. More than 30 years of biological research took place before the necessary genetic engineering tools became available. In 2003, Austin Burt proposed using HEGs to drive modified genes through a population [25]. Burt’s seminal work describing his idea, in combination with advanced knowledge about genetics and modern molecular tools, bolstered the field of inquiry into synthetic ‘gene drives’.
4. What Are Gene Drives?
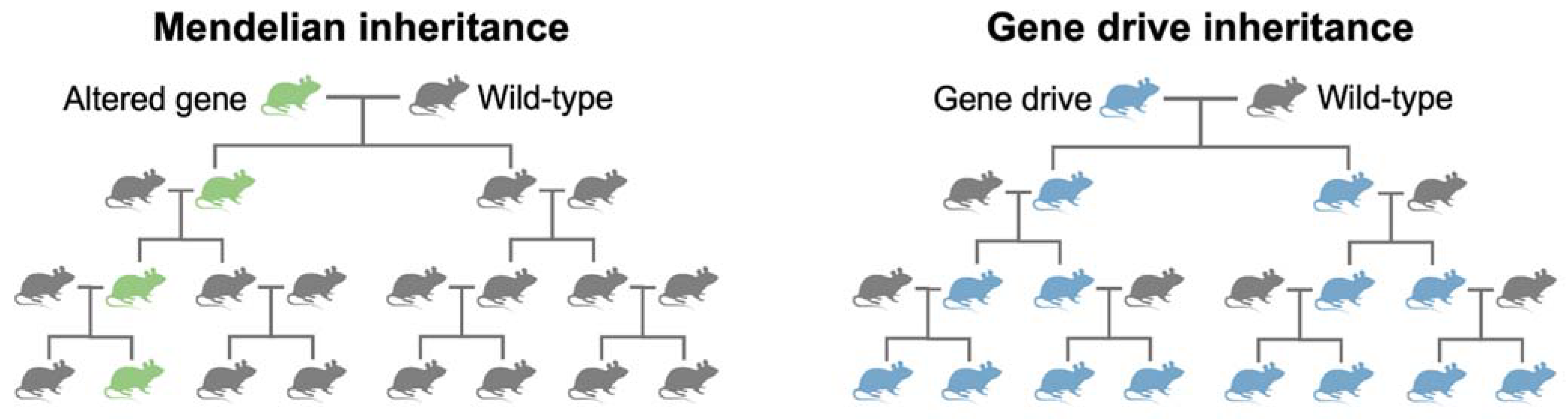
There is a growing acceptance that the term ‘gene drives’ simply refers to engineered selfish genetic elements. However, the terms ‘gene drive’ and ‘selfish genetic element’ are still used interchangeably. A 2016 report by the National Academies of Sciences [26] defined gene drives as “systems of biased inheritance in which the ability of a genetic element to pass from a parent to its offspring through sexual reproduction is enhanced”. Therefore, the result of a gene drive is the preferential increase in a specific genotype from one generation to the next, potentially spreading throughout the population. Figure 2 illustrates an idealised difference between Mendelian inheritance and inheritance through a gene drive.
Although this review focuses on gene drives for controlling populations of vertebrate pests, these elements have been proposed for numerous applications; perhaps the most prominent of these is to reduce populations of malaria-transmitting mosquitoes [28,29,30]. In this context, gene drives could provide a new means of tackling a disease that still infects 200 million people and causes 400,000 deaths each year [31]. Other potential applications outside pest control include genetically immunising populations of animals against disease and improving the sustainability of agriculture by reducing the demand for pesticides and herbicides [12,32]. Synthetic gene drives could have remarkable benefits for the environment, society and the economy.
5. Genetic Engineering
To develop synthetic gene drives, scientists required precision genetic engineering technologies, that facilitated site-specific modifications to the genome. Initial successes in the mid-1980s with precision genetic engineering were accomplished by microinjecting DNA sequences with high homology to the targeted genomic sequence into cells [33]. These experiments utilised a homologous recombination-dependent approach, which took advantage of the cell’s own DNA repair machinery to replace a targeted genetic locus with an introduced DNA sequence. However, the targeted engineering events occurred at very low rates (around 1 in 3 × 104 cells), even with the benefit of antibiotic selection [34].
6. Nuclease-Assisted Genetic Engineering
A key breakthrough in the field of genetic engineering was the realisation that double-stranded DNA breaks (DSBs) greatly stimulated cellular DNA repair mechanisms [35,36]. All eukaryotic cells efficiently repair DSBs using one of three primary pathways: non-homologous end-joining (NHEJ), microhomology-mediated end-joining (MMEJ) or via a form of homologous recombination, known as homology-directed repair (HDR; Figure 3). NHEJ simply joins the broken ends of the DNA, this can create small insertion or deletion mutations (indels); MMEJ can occur when short microhomologies exist, both upstream and downstream of the DSB, the two microhomologies can be annealed, often resulting in deletions of the intervening sequence; HDR uses a homologous DNA template to replace the broken region with high fidelity [37,38,39]. Thus, the induction of directed DSBs at a genomic locus of interest can stimulate HDR-based genetic engineering efficiency (Figure 3). As targeted DSBs in the genome could be introduced by nucleases, this method has become known as nuclease-assisted genetic engineering. It is the same premise that HEGs have naturally evolved to use and the logic behind Austin Burt’s idea of applying HEGs for developing synthetic gene drives.
Early nuclease-assisted genetic engineering experiments using HEGs demonstrated that DSBs increased the frequency of template integration [35]. In 2011, Windbichler et al. [40] collaborated with Austin Burt to describe the use of a HEG to create a gene drive in mosquitoes. In this instance, a mosquito was engineered with a HEG recognition site near a fluorescence gene, and on the expression of the HEG from a donor DNA plasmid, the recognition site was cut, allowing the HEG to be copied into the target site via HDR. The increased transmission of the HEG demonstrated these nucleases have the potential for developing gene drives. However, the practical utility of HEGs to target any genomic loci at will was limited by the long and highly specific DNA recognition sequences (14 to 40 bp) of these enzymes. Furthermore, the engineering of HEGs was extremely challenging for researchers because the DNA recognition sites and cleavage functions of these enzymes are intertwined in a single domain [41,42,43]
Fortunately, the possibility of manipulating genomes at will captivated scientists’ attention from all corners of biology. The initial challenges posed by early genetic engineering tools were soon overcome by developing a new genre of truly programmable nucleases.
7. ZFNs and TALENs
Programmable nucleases have transformed the field of genetic engineering. This class of nucleases can be engineered to bind to a specific nucleotide sequence in the genome and subsequently generate a DSB at a user-defined site. Zinc finger nucleases (ZFNs) and transcription activator-like effector nucleases (TALENs) were the first classes of programmable nuclease to be developed. These nucleases are engineered DNA binding proteins that facilitate targeted cutting of the genome in a similar way to HEGs [44]. ZFNs combine a nuclease domain derived from a restriction enzyme (typically FokI) with a DNA binding domain mediated by zinc fingers. As with ZFNs, TALENs typically utilise the same nuclease domain from FokI. In place of the zinc fingers, TALENs use an alternative DNA-binding domain called a Transcription Activator-Like effector (TALE), derived from the plant pathogenic bacterium Xanthomonas [45].
Although both ZFNs and TALENs are highly specific genome editors that have successfully modified the genome of many species, including mice and rats, their creation can be time-consuming and labour-intensive; requiring a new protein pair to be created for every DNA sequence to be edited [46,47]. Simoni et al. [48] demonstrated that ZFNs and TALENs could be used to develop gene drives much like HEGs, with homing frequencies of 34% and 49% to available target loci, respectively, in Drosophila melanogaster. Although, in many instances these systems are not transmitted reliably due to the number of repetitive elements within their design and their subsequent tendency to recombine, leading to their loss of function. Unfortunately, both ZFNs and TALENs were not optimal gene drive systems.
8. CRISPR-Cas
The arrival of the RNA-guided CRISPR-Cas (Clustered regularly interspaced short palindromic repeats-CRISPR-associated protein) system has been game-changing. The system was evolved as an adaptive immune system in bacteria and archaea, and was first observed in Escherichia coli in 1987 by its striking genomic structure [52]. The system uses a set of Cas genes to incorporate exogenous DNA sequences into the CRISPR locus, and subsequently transcribes them as RNA templates that guide the Cas proteins to cleave homologous invasive DNA or RNA [53]. However, the system was not repurposed into a genome editing tool until 2012 when Jinek et al. published their seminal paper. In the proceeding years, six types of CRISPR-Cas systems had been identified. Of these, the type II CRISPR-SpCas9 derived from Streptococcus pyogenes is, presently, the most widely used as a genome editing tool [54,55].
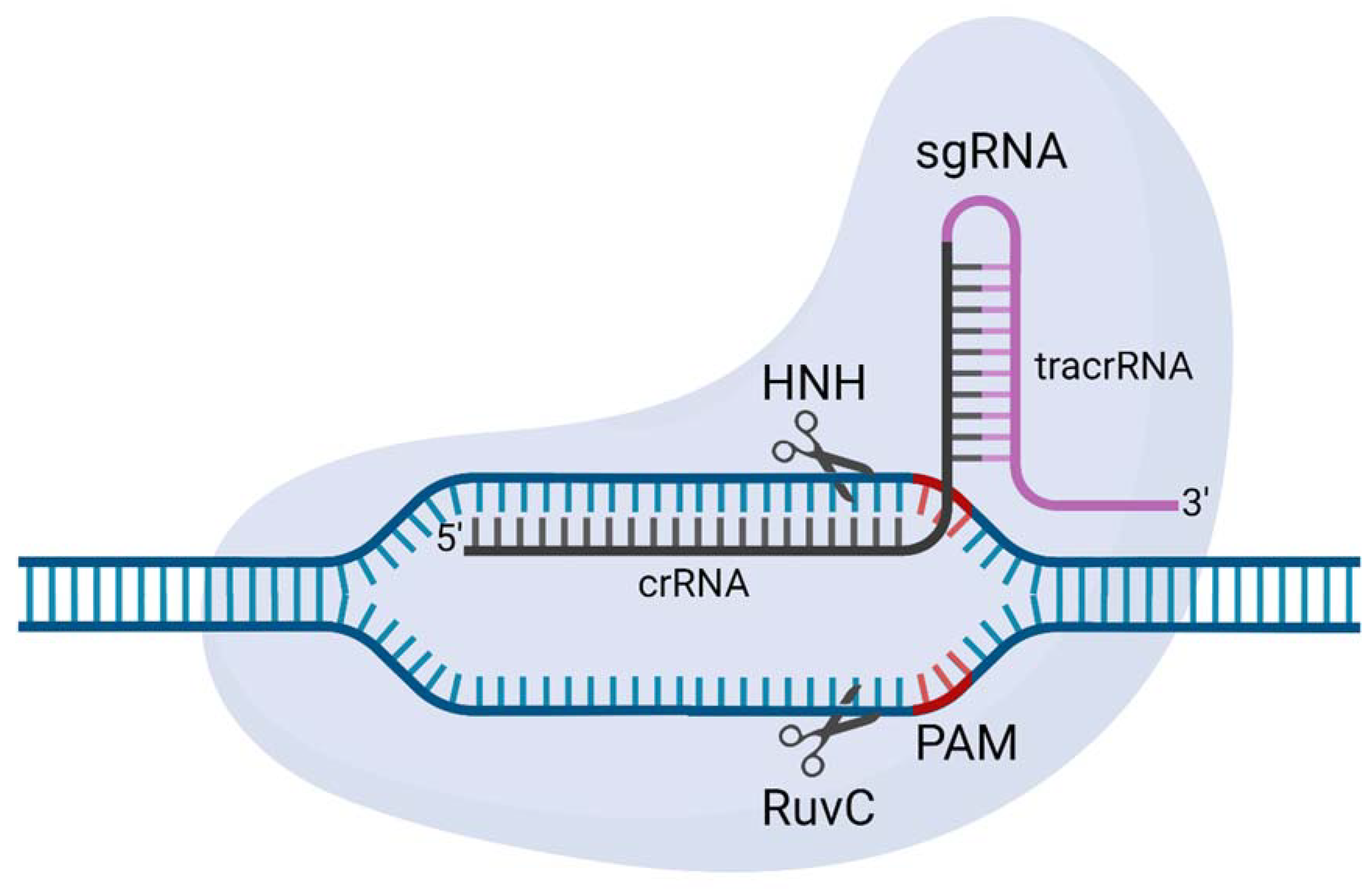
The S. pyogenes type II CRISPR-Cas system consists of a SpCas9 nuclease and two RNAs; CRISPR RNA (crRNA) and trans-activating CRISPR RNA (tracrRNA). The crRNA and tracrRNA hybridise to form a guide RNA (gRNA) before complexing with SpCas9 [56]. SpCas9 is directed to the target site by a 20 nt sequence of the crRNA that Watson–Crick base-pairs with the target DNA sequence [49]. However, crRNA and tracrRNA can be fused to form a 102 nt single-guide RNA [50], which is referred to as a single-guide RNA (sgRNA). Once the gRNA guides the SpCas9 to the target site, the cleavage of the target site only occurs if a protospacer adjacent motif (PAM) and a defined 3 nt sequence at the 3’ end of the crRNA sequence is present. For SpCas9, the PAM sequence is 5’-NGG, although it can recognise other noncanonical 5’-NAG and 5’-NGA PAMs to a lesser extent [57]. At an overall structural level, SpCas9 contains two nuclease domains, HNH and RuvC, each of which cleaves one strand of the target DNA (Figure 4). In SpCas9, these domains cleave 3 bp upstream from the PAM sequence, leaving blunt ends [57,58,59]
In contrast to ZFNs and TALENs, which require the design of proteins that are encoded by large repetitive DNA segments (500 to 1500 bp) for each new target site, CRISPR-SpCas9 can be easily adapted to target almost any genomic sequence by simply exchanging the crRNA sequence. The Cas protein component remains unchanged, alleviating the need for complex protein engineering. Therefore, the reagents are simple, cheap and quick to design and generate [60].
9. CRISPR-Based Gene Drives
CRISPR-Cas allows scientists to engineer synthetic gene drives in a standard molecular laboratory. SpCas9-based gene drives have been developed in yeast [61], D. melanogaster [62], three species of mosquitoes [30,63,64] and most recently, in mice [65,66,67]. These studies have demonstrated mechanisms of how gene drives could be used to achieve two potential outcomes:
- Population suppression: the spread of a genetic element that causes the number of individuals in a population to decrease.
- Population replacement: the spread of a genetic element that causes a population’s genotype to change.
To date, gene drive systems with the intention of population suppression have been engineered in proof-of-concept studies in mosquitoes, drosophila, and mice [30,64,65,68,69,70]. However, gene drives with the alternative aim of population replacement, rather than suppression, have been developed in the same three species, as well as yeast [61,62,63,66].
For this review, we will concentrate on strategies for population suppression in vertebrates, with a specific focus on mice, to establish gene drives in vertebrates. In addition to being invasive pests, we have more information on mice at the population, genetic and ecological levels than for most other potential targets of genetic pest management [71,72]. Importantly, we also have highly developed tools for manipulating the genomes of mice. For these reasons, almost all vertebrate gene drive research has been undertaken in mice.
10. Driving Population Suppression
Gene drives could enable humane population suppression by distorting the sex-ratio of a damaging pest population in a species-specific manner. By distorting the sex ratio of the breeding population away from the favoured Fisherian ratio of 1:1 male to female, these strategies could manipulate the reproductive performance of a population. In most pest species, including mice and rats, female procreative capacity is responsible for maintaining the overall population size [73,74]. Therefore, an effective means of population suppression is to bias the sex ratio of the breeding population in favour of males. This could be accomplished by either spreading female infertility through a population or by ensuring all or most offspring born are male [30,68]. A largely male breeding population will result in population suppression, while an all-male breeding population will lead to eradication.
One possibility that has been suggested and is currently under investigation is to engineer the mouse t-haplotype to carry the sex-determining Sry gene in genetic females so they develop as males but are sterile because they lack essential Y-linked genes for sperm development [75,76,77]. However, this approach may face challenges as two laboratory studies on independent mouse populations have found that male mice carrying the t-haplotype fertilise only around 20% of offspring when competing against wild-type males [78,79]. This can be attributed to the relative sperm competitiveness of t-haplotype males compared to wild-type males and may explain why the frequency of the t-haplotype in wild populations is relatively low, typically ranging between 5 and 30% [18,80].
10.1. tCRISPR
An extension of the concept of using the t-haplotype to spread a population-suppressing genotype is the tCRISPR approach [65]. This method involves integrating a tCRISPR cassette harbouring a Cas nuclease and one or more gRNAs into a neutral intergenic region within the t-haplotype. The Cas and gRNAs are expressed in the male germline to cut and generate loss-of-function copies of a haplosufficient female fertility gene through the NHEJ or MMEJ error-prone repair pathways. The idea is that the t-haplotype segregation distortion in males will increase the frequency of the tCRISPR cassette within the population, which in turn, will continuously generate loss-of-function copies of the targeted haplosufficient female fertility gene. With each generation, the frequency of infertile homozygous females carrying faulty copies of the targeted haplosufficient female fertility gene will increase, ultimately reducing population size. The team has successfully engineered a SpCas9 tCRISPR mouse, targeting the prolactin gene (Prl). The modified t-haplotype exhibited biased transmission (95%), and loss-of-function alleles in the Prl gene were at a frequency of 80% [65,81]. The team’s models predict that these levels are sufficient for eradication. Although promising, the formation of functionally resistant alleles in the haplosufficient female fertility gene and unpredicted fitness costs remain to be explored in more detail.
10.2. X-Shredder
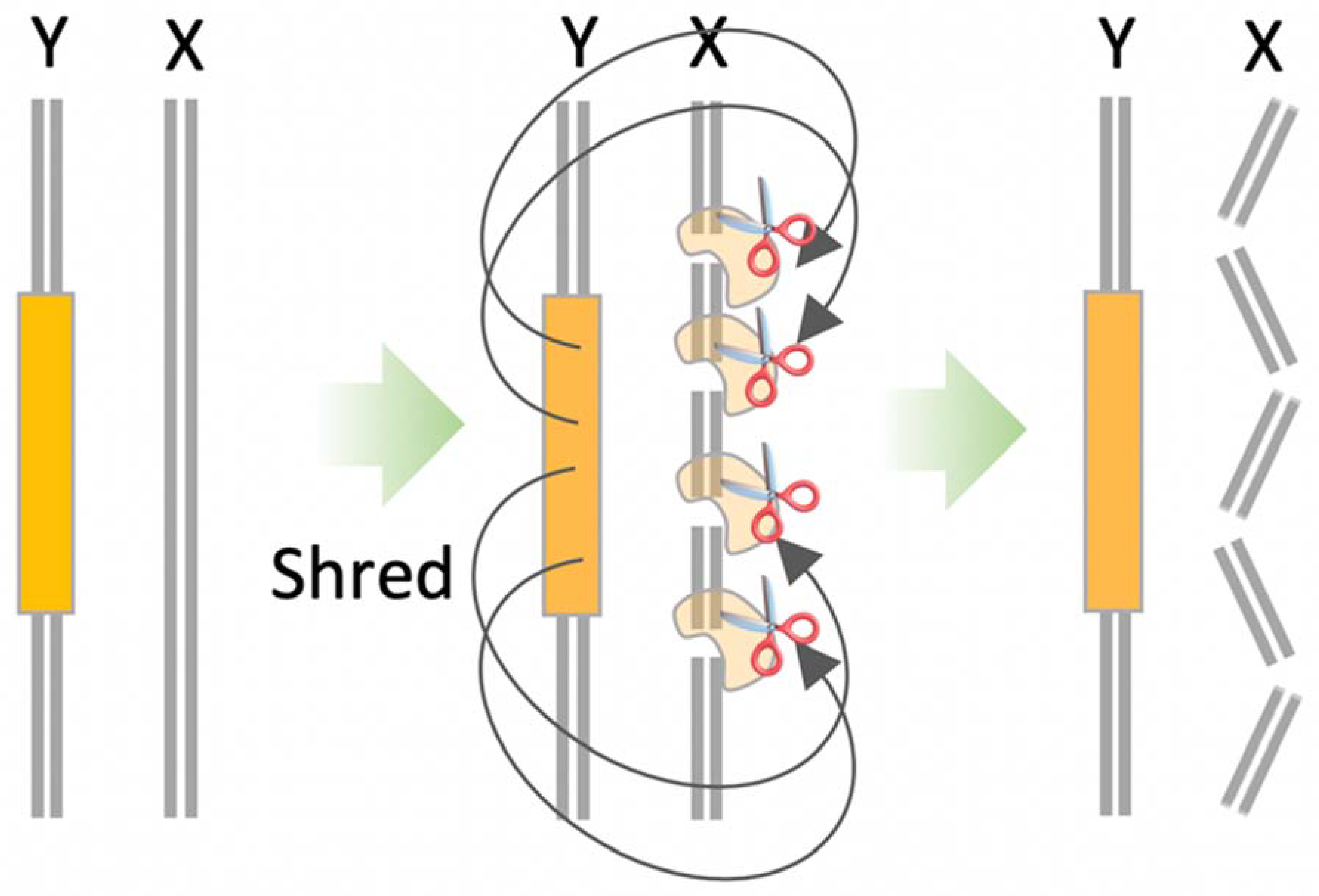
An alternative approach is the X-shredder (XS) strategy—a type of synthetic meiotic drive that employs a driving Y-chromosome. In XY heterogametic species, such as rodents, an XS is a type of sex-ratio distorting gene drive that cuts the X-chromosome at multiple sites during spermatogenesis—shredding the X-chromosome beyond repair [73]. To engineer an XS using CRISPR, an XS cassette is inserted within a neutral intergenic region of the Y-chromosome. The cassette encodes a Cas nuclease, which is expressed under the control of a spermatogenesis-specific promoter, and one or more gRNAs that target conserved repetitive sequences unique to the X-chromosome (Figure 5). Given that the X-chromosomes will be destroyed during spermatogenesis, most sperm that mature and reach the oocyte for fertilisation are Y-bearing, resulting in bias towards male offspring. By placing the XS cassette on the Y-chromosome, all male offspring will inherit the XS cassette and continue transmitting the XS to subsequent generations [27].
Significant progress has been made in developing this concept in insects. A CRISPR-based XS system has been engineered in mosquitoes, resulting in a male bias among offspring that ranges from 86.1% to 94.8% in laboratory-contained populations [68]. Although successful in mosquitos, there are technical challenges that face the adaption of an XS into rodents. These include identifying appropriate spermatogenesis-specific promoters and the transcriptional silencing of mammalian sex chromosomes during meiosis. The latter may hinder the expression of the Cas nuclease from the Y-chromosome and the accessibility of the X-chromosome for shredding [27].
Y-CHOPE (Y-CHromosome deletion using Orthogonal Programmable Endonucleases) is a proposed alternative to the X-shredder strategy, which aims to shred the Y-chromosome instead of the X, converting XY males into fertile XO females [82]. For Y-CHOPE to effectively result in population decline, it requires the use of two different Cas endonucleases systems: one for homing the Y-CHOPE cassette to an autosome, and another for shredding the Y-chromosome. While Prowse et al. [82] successfully demonstrated the elimination of the Y-chromosome in mouse cells using a transiently expressed Cas nuclease and gRNAs, Y-CHOPE still faces significant challenges. These include the reliance on efficient homing of the Y-CHOPE cassette and the possibility of temporarily increasing the population size in polygynous species until males become limited.
10.3. Homing-Based Gene Drives
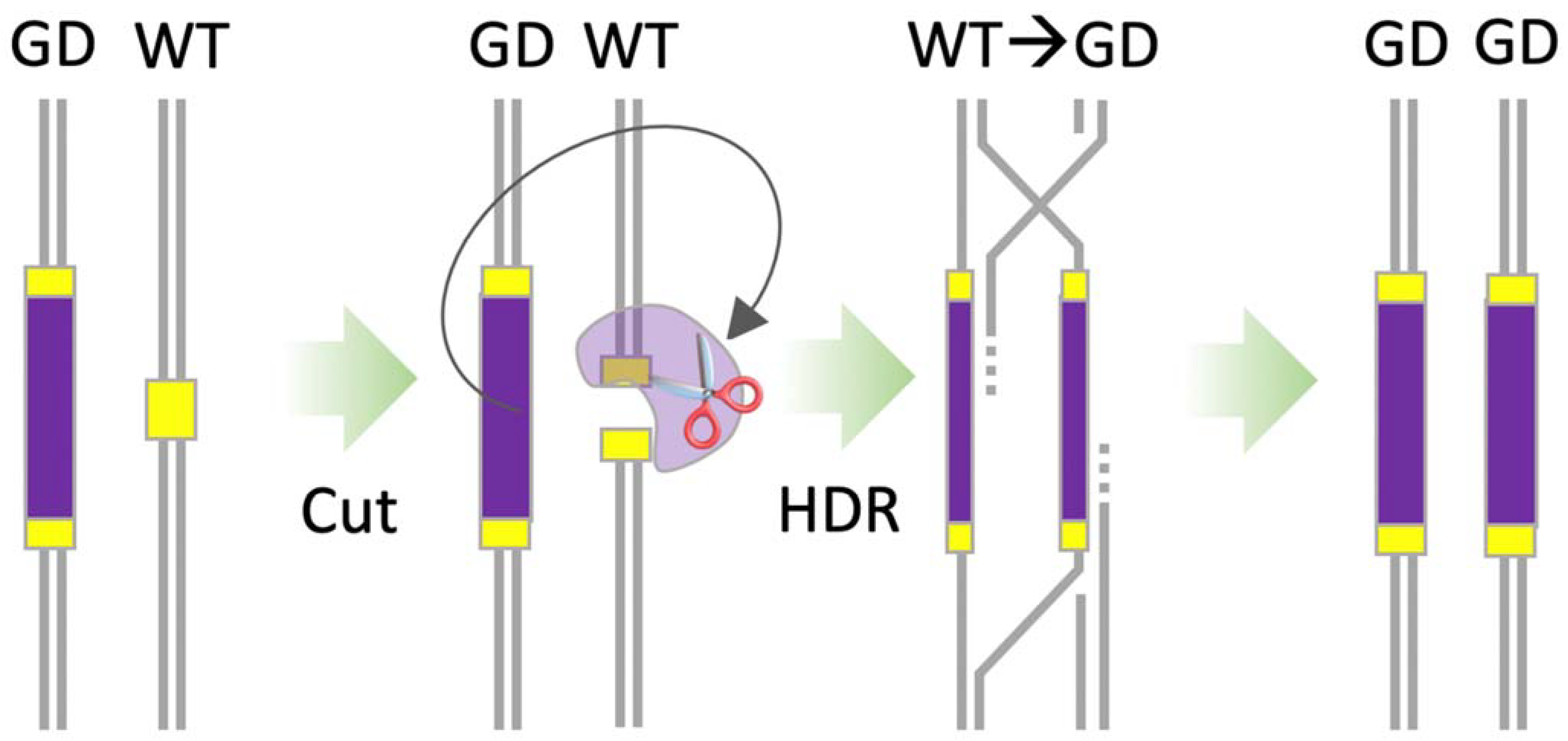
CRISPR-based homing gene drives were theorised and named after Austin Burt’s homing endonuclease gene drive approach, which drives through an over-replication mechanism. A homing gene drive works by copying or ‘homing’ itself into a specified target site in the genome. To build a CRISPR-based homing gene drive, an animal is engineered to harbour a gene drive cassette that expresses a Cas endonuclease, such as SpCas9, and one or more gRNAs. The Cas nuclease and gRNAs are expressed from one allele to cut at a target site on the sister allele on the homologous chromosome (Figure 6). After the CRISPR-mediated cleavage, HDR results in the CRISPR machinery and any additional genetic payload (trait of interest) included in the gene drive cassette being copied onto the homologous chromosome. This process ensures the cell is homozygous for the gene drive cassette. When this process occurs in the germ cells, the proportion of mature gametes in an animal that contains the gene drive cassette is greater than 50%, and therefore the cassette could drive itself through the population, spreading the trait of interest [25,27,73].
In 2019, a team led by Dr Kimberly Cooper at the University of California San Diego, published a successful homing-based gene drive in mice, capable of driving a fluorescent genetic element embedded in the mouse tyrosinase (Tyr) gene, through the female germline. Tyr affects mouse coat colour, which enabled the frequency of the gene drive to be tracked over generations by monitoring coat colour and confirmed by PCR-based genotyping. Their successful gene drive design achieved up to 72% transmission in the female germline, but no homing was observed in males harbouring the same gene drive system [66]. The team hypothesised that sex-specific differences were due to the timing of SpCas9 nuclease expression during meiosis. To investigate this further, the team developed a new gene drive strategy with adjusted the timing of SpCas9 expression during meiosis. With this approach, they achieved homing in both females and males [67]. Although the transmission levels were low, importantly, the team demonstrated that CRISPR-based homing gene drives can function in mammals.
In parallel, Dr Paul Thomas and his colleagues were working on their own mouse gene drive systems. They developed and tested two systems that were able to generate indels at target sites but observed no homing of the gene drive allele. The team published an in-depth analysis of the mice [83]. The insights from their analysis align with previous reports, highlighting the importance of robust and specific SpCas9 expression during meiosis for efficient CRISPR-based homing gene drives. These studies have revealed the nuanced challenges homing gene drives in mammals face compared to insect counterparts. Ongoing efforts developing homing gene drives in rodents are focused on identifying promoters that provide a sufficient level of SpCas9 expression during meiosis I.
Once a suitable promoter is identified, a homing gene drive could be used for population suppression. The system could be targeted to a haplosufficient female-fertility gene to disrupt the coding sequence of the gene, rendering homozygous female offspring infertile. In contrast, males and heterozygous females will retain normal fertility and continue transmitting the drive. With every generation, the sex-ratio of the breeding population will become more biased toward males, eventually resulting in a population decline due to the lack of fertile females available as breeding partners [30,84]. Hammond et al. [30] developed this system in mosquitos and achieved transmission rates of 91.4 to 99.6% in caged populations. Further investment and research is needed if similar transmission levels are to be achieved in vertebrates.
11. Containment Strategies
The forms of gene drive described above may only require the release of a small number of animals to spread through a population. The extent of the spread would be limited by naturally arising resistant alleles that prevent CRISPR from recognising its target site. Resistant alleles could exist in the population before release or originate from indels generated when CRISPR-mediated cleavage is repaired by the error-prone NHEJ pathway and alters the 20nt crRNA recognition sequence [85,86]. The rate of NHEJ-mediated repair will depend on the species, target site and timing of SpCas9 expression. Given that natural selection tends to favour equal sex ratios, resistant alleles that restore function would spread rapidly through the population to equalise the sex ratio [27,87].
If enough genetic information is known about the target pest population, scientists can leverage naturally present genetic differences between populations of the same species to restrict the spread of a gene drive. These genetic differences between subpopulations are known as ‘private alleles’. Targeting CRISPR to a specific private allele, which could be a single nucleotide alteration, would restrict the spread of the drive to only the population of animals carrying that private allele [88,89]. The caveat with containing a gene drive using private alleles is that widespread DNA sequencing of the population needs to occur prior to developing a gene drive strategy.
Compared to a homing gene drive, XS is less likely to be contained using private alleles and less prone to inactivation by resistant alleles because the strategy targets multiple sites and, therefore, would require an animal to simultaneously acquire multiple resistant alleles to halt the drive. It has been suggested that the impact of resistant alleles generated by homing gene drives could be reduced by using multiple gRNAs closely spaced along the target region [12]. Several in silico and in vivo studies have now demonstrated that multiple gRNAs can assist homing gene drives to evade drive resistance and successfully spread through pest populations [70,84,90,91]. Even if drive resistance were to prevail, it would be possible to release a second gene drive targeting a different sequence, such as the newly acquired resistant allele, to continue suppressing the population.
If a gene drive were not limited by private or resistant alleles, it could potentially spread through the entire species. Therefore, it is essential to have strategies in place that can deliberately inactivate a gene drive that was causing unforeseen impacts. Fortunately, both homing gene drives and XS systems could be inactivated by the releasing animals that have engineered resistant alleles or a reversal gene drive that immunises the animal against the original drive [12]. However, these reactive stop buttons are not ideal as timely deployment requires that appropriate animals have been generated, gained regulatory approval, and are in sufficient a number at the required location.
More proactive approaches are needed, and scientists are working to develop gene drives that are inherently self-limiting—that is, they stay in a population transiently and could, therefore, be localised to targeted populations. Numerous self-limiting gene drive concepts have been proposed, and there is a growing body of evidence for the development and application [92,93,94,95,96,97]. Of these, daisy drives have been the focus of several in silico population modelling studies and have been well-publicised; daisy drives include: ‘daisy-chain’ and ‘daisyfield’ drives. These systems are complex and have yet to be engineered for population suppression in any species; however, there is value in describing them here as future development would put the power in the hands of communities to locally deploy a gene drive, avoiding the need for complex geopolitical agreements that will be required for the release of a self-perpetuating gene drive.
11.1. Daisy-Chain System
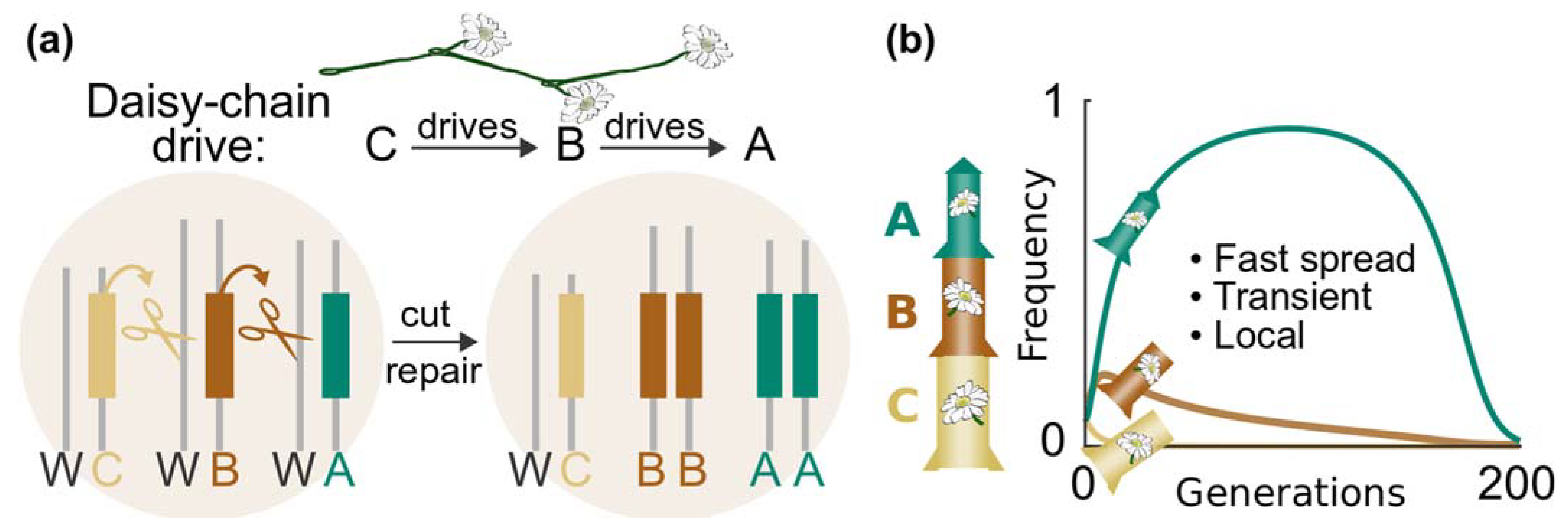
In daisy-chain gene drive systems, the CRISPR components are scattered throughout the genome so that none of them can be copied on its own. Each element is reliant on the presence of another element to spread. Although each element is physically separated in the genome, they are functionally arranged in a daisy chain. As can be seen in Figure 7a, element C causes element B to drive, and element B causes element A to drive. Element C cannot drive, so its abundance is limited by the number of daisy-drive animals released. That means B will initially increase in frequency, then decline as C elements become scarce. In time, A will increase even more rapidly, but will eventually run out of B and disappear [96].
With a sufficient understanding of the target population, daisy chains provide a genetic clock that could be programmed to function for an approximate number of generations before running out of genetic fuel. The elements of a daisy-chain system are analogous to the booster stages of a genetic rocket: those at the base of the daisy chain help lift off until they run out of fuel and are successively lost (Figure 7b). In silico modelling shows that releasing a daisy-chain drive with a five-element chain is hundreds of times more efficient than releasing one with only element A [96].
In theory, daisy-chain systems can do anything a standard homing gene-drive system can achieve, although the complexity of engineering such a system and having it stably inherited poses a significant hurdle. Daisy-chain drives were theorised in 2016; at present, a daisy chain of two or more homing elements has not been reported.
11.2. Daisyfield System
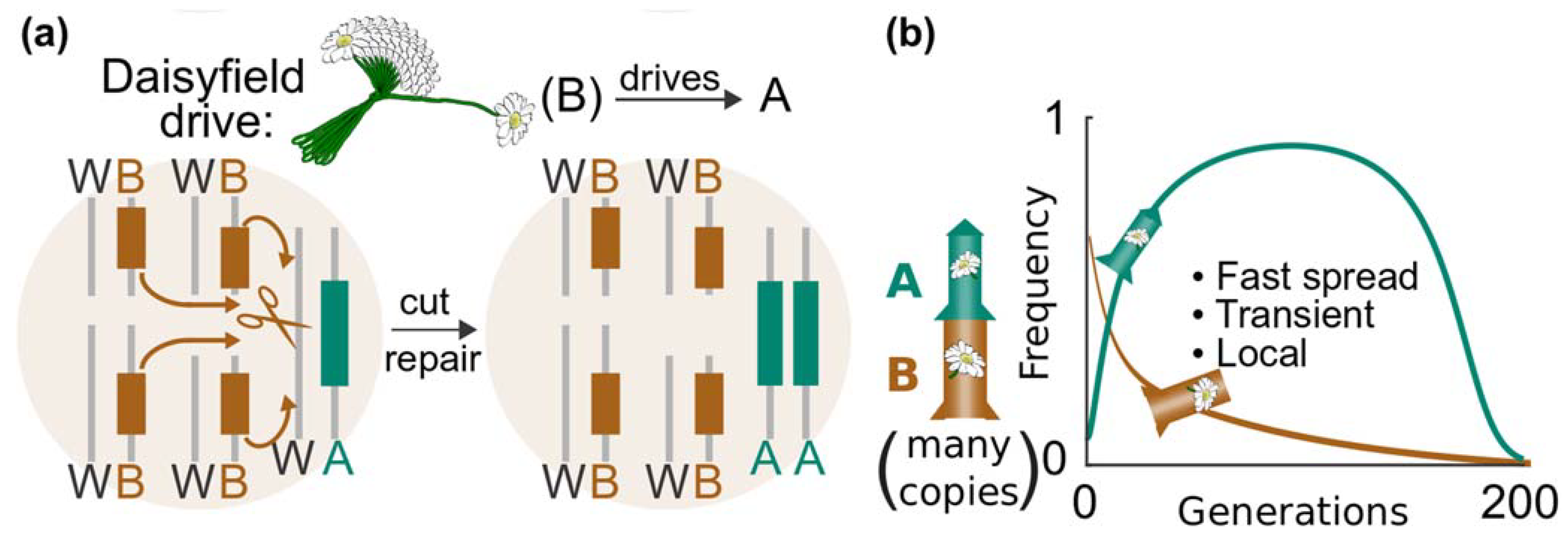
Another option is to use a form of daisy drive that does not require each of the booster elements to home via HDR. This significantly reduces engineering complexity. Instead of a daisy chain of linked elements (C→B→A), a daisyfield system scatters B elements (gRNA expression cassettes), throughout neutral loci in the genome (Figure 8a). If at least one of these is present, element A (Cas nuclease) will be copied, propagating the trait of interest. Depending on the location of the Cas nuclease, this could knock out a gene of interest by disrupting the coding sequence or propagate a genetic cargo that contains a trait of interest. With every generation of mating with wild-type animals, it halves the number of daisy B elements (gRNAs), eventually burning out as the generational clock winds down (Figure 8b) [95,99].
Compared to a daisy-chain drive, a daisyfield system should be much simpler to engineer. For example, the element A (Cas nuclease) could be targeted to the site of interest using CRISPR-assisted genome engineering and the B elements (gRNA expression cassettes) scattered through the genome using lentiviral integration to generate a daisyfield animal. Daisyfield drives consisting of one B element have been successfully constructed as a ‘split drive’ confinement strategies [61], and the engineering technicalities of increasing the number B elements should not be inhibitory.
12. Public Perception
Genetically engineered animals typically pose minimal risk to the environment. Most engineered traits are for human benefit and will not be favoured by natural selection. By contrast, gene drives could spread through a population even if they reduce the fitness of each carrier animal. This gives gene drives the potential to unintentionally affect extraneous ecosystems [12,27]. Although technical and regulatory hurdles exist for the practical use of gene-drive-based control of invasive species, perhaps the most significant hurdle to be overcome will be public acceptance of the technology.
Public acceptance starts with open, trustworthy research. Researchers involved in the field have been keen to highlight the importance of transparency when undertaking research and have stated this commitment by disclosing research plans before commencing [101,102]. With previous emerging biotechnologies, such as the first genetically modified foods, the authority of scientists centred on their knowledge and expertise; with gene drives, a “moral authority” has become increasingly valued [103]. To help gene-drive scientists make morally correct decisions, lay society is involved in the development and application process. Dr Kevin Esvelt and his team have been strong proponents of this approach. As part of their Mice Against Ticks project, society members in the proposed release zone are regularly engaged to share their suggestions and concerns, and scientists adjust their research accordingly [104].
In addition to the importance of open and trustworthy science, the public acceptance of gene drives will also be swayed by whether its use is sufficiently warranted to solve the problems being addressed. A Pew Research Centre study by Funk and Hefferon [105] indicated that public attitudes toward the use of genetic engineering on animals, at least in the US, are often supportive if the technology is being applied to a serious human health issue, such as mosquitoes transmitting disease. The US public was less supportive of other uses involving the environment, such as reviving extinct species to restore biodiversity. Whether the public considers the damage from invasive vertebrates a problem that warrants the use of gene drives is yet to be determined.
The acceptance of gene drives among the general public will continue to be complicated by the complex science that underlies the technology, making it difficult for scientists to communicate to the public and challenging for the public to grasp without knowledge of genetics and reproduction [106]. In most cases, the target audience will not achieve a detailed understanding of gene drive technology. Still, the public should be aware of the risks and potential benefits of using gene drives. If used wisely and the risks managed appropriately, gene drives could transform public health, agriculture and ecosystem conservation; but for these benefits to be realised, the public must be supportive. As seen with previous biotechnologies, public support is less likely to be determined by scientific debate but will hinge on the trustworthiness of research, the problem the technology addresses and the outcomes of initial trials.
13. Governance and Regulations
With gene-drive research progressing, adequate regulations and governance for the environmental release of gene-drive organisms must be in place before a release can occur. There are regulations in place in many countries to guard against the unintended effects of releasing genetically engineered organisms into the environment. However, these regulations were developed for animals with conventional genetic modifications that are typically for welfare or human benefit and reduce the animal’s fitness outside a controlled environment. Current regulations call for a step-by-step process where the first releases are in small numbers in isolated areas [107]. With gene drives, the intention is for the engineered animal genotypes to spread, to some extent, in the environment, so there is a need to amend established regulations.
Although there are many cases of genetically modified organisms being legally released into the wild, at present, no nation has regulations in place specifically for gene drives and the release of an organism with an engineered gene drive has not been recorded. The most broad-ranging and widely accepted international governance system for genetic resources and biosafety is the United Nations Convention on Biological Diversity (UNCBD), as implemented through the Cartagena and Nagoya Protocols [26,108]. The regulatory systems that many countries have developed in response to the Cartagena Protocol, are predicated on a strong precautionary, nearly preventative approach. To cope with the unique aspects of gene drive, existing approaches to governance need to be adapted. Integrating new policies and laws for gene drives into existing international governance poses significant challenges, but it is necessary, and this need was highlighted in 2020 by a UNCBD Ad Hoc Technical Expert Group (AHTEG) Report on Risk Assessment [109].
14. Conclusions
- The road towards gene drive technology has traced centuries of scientific investigation and technological breakthroughs. The study and understanding of naturally occurring selfish genetic elements, such as homing endonuclease genes and meiotic drive, has been essential to conceptualising synthetic gene drives.
- To engineer synthetic gene drives, scientists required precision genetic engineering technologies. Due to their complex protein designs, initial attempts using early genome engineering nucleases struggled. The repurposing of CRISPR-SpCas9 into a genome engineering tool overcame many of these challenges and allowed synthetic gene drives to be engineered in a standard molecular laboratory.
- CRISPR-based gene drives have been engineered in yeast, D. melanogaster, three species of mosquitoes and most recently in mice. The gene drives developed in mice have had lower transmission rates than previous invertebrate gene drives. Studies of synthetic mouse gene drives have revealed the challenges of engineering gene drives in mammals.
- Future research efforts aim to optimise vertebrate gene drive designs to improve transmission rates. For homing gene drives, this includes identifying or engineering promoters that provide robust and specific SpCas9 expression during meiosis I. In the immediate future, non-homing gene drive designs, such as tCRISPR or X-shredder may be more feasible in vertebrate species.
- Gene drives for real-world application should include built-in molecular containment strategies to help contain the spread and stop unforeseen impacts. There is a range of molecular-safeguarding options for vertebrates, including targeting private alleles and daisy drives. These molecular containment approaches should be used in parallel to efforts that physically isolate the target population.
- Public acceptance and governance will determine the speed and extent of application. Public acceptance will hinge on the trustworthiness of research, the problem the technology addresses and the outcomes of initial trials. Before gene drives can be applied as a vertebrate biocontrol tool, updating governance and regulations to include the environmental release of gene-drive organisms is required.
- The application of gene drive for vertebrate biocontrol will have profound implications for society, our economy and the environment. With technological development progressing swiftly, public and political engagement must move in parallel to ensure applications align with society’s values.
Author Contributions
Writing—original draft preparation, G.R.M.; writing—review and editing, G.R.M., C.B.A.W. and S.G.L. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support for this work was provided by NSW Department of Primary Industries, the Commonwealth Scholarship Commission UK, the European Squirrel Initiative, and the Bio-technology and Biological Sciences Research Council (BBSRC) Institute Strategic Programme Grant BB/P013759/1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Krull, C.R.; Galbraith, J.A.; Glen, A.S.; Nathan, H.W. Invasive vertebrates in Australia and New Zealand. In Austral Ark: The State of Wildlife in Australia and New Zealand; Stow, A., Holwell, G.I., Maclean, N., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 197–226. [Google Scholar]
- Robertson, P.A.; Adriaens, T.; Lambin, X.; Mill, A.; Roy, S.; Shuttleworth, C.M.; Sutton-Croft, M. The large-scale removal of mammalian invasive alien species in Northern Europe. Pest Manag. Sci. 2017, 73, 273–279. [Google Scholar] [CrossRef]
- Witmer, G.W.; Fuller, P.L. Vertebrate species introductions in the United States and its territories. Curr. Zool. 2011, 57, 559–567. [Google Scholar] [CrossRef]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Tatem, A.J. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef] [PubMed]
- Singleton, G. Integrated management of rodents: A Southeast Asian and Australian perspective. Belg. J. Zool. 1997, 127, 157–169. [Google Scholar]
- Singleton, G. Impacts of Rodents on Rice Production in Asia; IRRI Discussion Paper Series; IRRI: Los Baños, Philippines, 2003. [Google Scholar]
- Howal, G.; Donald, J.; Galvan, J.P.; Russel, J.; Parkes, J.; Samaniego, A.; Tershy, B. Invasive Rodent Eradication on Islands. Conserv. Biol. 2007, 21, 1258–1268. [Google Scholar] [CrossRef]
- Towns, D.R.; Atkinson, I.A.E.; Daugherty, C.H. Have the Harmful Effects of Introduced Rats on Islands been Exaggerated? Biol. Invasions 2006, 8, 863–891. [Google Scholar] [CrossRef]
- Meerburg, B.G.; Brom, F.W.; Kijlstra, A. The ethics of rodent control. Pest Manag. Sci. 2008, 64, 1205–1211. [Google Scholar] [CrossRef]
- Lorvelec, O.; Pascal, M. French attempts to eradicate non-indigenous mammals and their consequences for native biota. Biol. Invasions 2005, 7, 135–140. [Google Scholar] [CrossRef]
- Burt, A.; Trivers, R. Genes in Conflict: The Biology of Selfish Genetic Elements; Harvard University Press: Cambrideg, MA, USA, 2016. [Google Scholar]
- Esvelt, K.M.; Smidler, A.L.; Catteruccia, F.; Church, G.M. Concerning RNA-guided gene drives for the alteration of wild populations. Elife 2014, 3, e03401. [Google Scholar] [CrossRef]
- Fraser, M.J., Jr. Insect transgenesis: Current applications and future prospects. Annu. Rev. Entomol. 2012, 57, 267–289. [Google Scholar] [CrossRef]
- Jasin, M. Genetic manipulation of genomes with rare-cutting endonucleases. Trends Genet. 1996, 12, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Belfort, M.; Bonocora, R.P. Homing endonucleases: From genetic anomalies to programmable genomic clippers. Methods Mol. Biol. 2014, 1123, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Windbichler, N.; Papathanos, P.A.; Catteruccia, F.; Ranson, H.; Burt, A.; Crisanti, A. Homing endonuclease mediated gene targeting in Anopheles gambiae cells and embryos. Nucleic Acids Res. 2007, 35, 5922–5933. [Google Scholar] [CrossRef] [PubMed]
- McDermott, S.R.; Noor, M.A.F. The role of meiotic drive in hybrid male sterility. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 1265–1272. [Google Scholar] [CrossRef]
- Ardlie, K.G. Putting the brake on drive: Meiotic drive of t haplotypes in natural populations of mice. Trends Genet. 1998, 14, 189–193. [Google Scholar] [CrossRef]
- Silver, L.M. The peculiar journey of a selfish chromosome: Mouse t haplotypes and meiotic drive. Trends Genet. 1993, 9, 250–254. [Google Scholar] [CrossRef]
- Bauer, H.; Schindler, S.; Charron, Y.; Willert, J.; Kusecek, B.; Herrmann, B.G. The nucleoside diphosphate kinase gene Nme3 acts as quantitative trait locus promoting non-Mendelian inheritance. PLoS Genet. 2012, 8, e1002567. [Google Scholar] [CrossRef]
- Bauer, H.; Véron, N.; Willert, J.; Herrmann, B.G. The t-complex-encoded guanine nucleotide exchange factor Fgd2 reveals that two opposing signaling pathways promote transmission ratio distortion in the mouse. Genes Dev. 2007, 21, 143–147. [Google Scholar] [CrossRef]
- Herrmann, B.G.; Bauer, H. The mouse t—Haplotype: A selfish chromosome—Genetics, molecular mechanism, and evolution. Evol. House Mouse 2012, 3, 297. [Google Scholar]
- Hickey, W.A.; Craig, G.B. Genetic Distortion of Sex Ratio in a Mosquito Aedes aegypti. Genetics 1966, 53, 1177. [Google Scholar] [CrossRef]
- Craig, G.B.; Hickey, W.A.; Vandehey, R.C. Inherited Male-Producing Factor in Aedes aegypti. Science 1960, 132, 1887–1889. [Google Scholar] [CrossRef] [PubMed]
- Burt, A. Site-specific selfish genes as tools for the control and genetic engineering of natural populations. Proc. R. Soc. B Biol. Sci. 2003, 270, 921–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Academies of Sciences, Engineering, and Medicine. Gene Drives on the Horizon: Advancing Science, Navigating Uncertainty, and Aligning Research with Public Values; The National Academies Press: Washington, DC, USA, 2016. [Google Scholar]
- McFarlane, G.R.; Whitelaw, C.B.A.; Lillico, S.G. CRISPR-Based Gene Drives for Pest Control. Trends Biotechnol. 2018, 36, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Adelman, Z.N.; Tu, Z. Control of Mosquito-Borne Infectious Diseases: Sex and Gene Drive. Trends Parasitol. 2016, 32, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Galizi, R.; Doyle, L.; Menichelli, M.; Bernardini, F.; Deredec, A.; Burt, A.; Crisanti, A. A synthetic sex ratio distortion system for the control of the human malaria mosquito. Nat. Commun. 2014, 5, 3977. [Google Scholar] [CrossRef]
- Hammond, A.; Galizi, R.; Kyrou, K.; Simoni, A.; Siniscalchi, C.; Katsanos, D.; Nolan, T. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nat. Biotechnol. 2016, 34, 78–83. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. World Malaria Report 2019; World Health Organization: Geneva, Switzerland, 2019.
- Whitelaw, B.; McFarlane, G.R. Accelerating evolution. Biologist 2019, 66, 18–21. [Google Scholar]
- Smithies, O.; Gregg, R.G.; Boggs, S.S.; Koralewski, M.A.; Kucherlapati, R.S. Insertion of DNA sequences into the human chromosomal β-globin locus by homologous recombination. Nature 1985, 317, 230–234. [Google Scholar] [CrossRef]
- Capecchi, M.R. Altering the genome by homologous recombination. Science 1989, 244, 1288–1292. [Google Scholar] [CrossRef]
- Rouet, P.; Smih, F.; Jasin, M. Introduction of double-strand breaks into the genome of mouse cells by expression of a rare-cutting endonuclease. Mol. Cell. Biol. 1994, 14, 8096–8106. [Google Scholar] [CrossRef]
- Smih, F.; Rouet, P.; Romanienko, P.J.; Jasin, M. Double-strand breaks at the target locus stimulate gene targeting in embryonic stem cells. Nucleic Acids Res. 1995, 23, 5012–5019. [Google Scholar] [CrossRef]
- Liu, M.; Rehman, S.; Tang, X.; Gu, K.; Fan, Q.; Chen, D.; Ma, W. Methodologies for Improving HDR Efficiency. Front. Genet. 2019, 9, 691. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, T.; Nakade, S.; Sakane, Y.; Suzuki, K.-I.T.; Yamamoto, T. MMEJ-assisted gene knock-in using TALENs and CRISPR-Cas9 with the PITCh systems. Nat. Protoc. 2016, 11, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Yanik, M.; Ponnam, S.P.G.; Wimmer, T.; Trimborn, L.; Müller, C.; Gambert, I.; Stieger, K. Development of a Reporter System to Explore MMEJ in the Context of Replacing Large Genomic Fragments. Mol. Ther. Nucleic Acid 2018, 11, 407–415. [Google Scholar] [CrossRef]
- Windbichler, N.; Menichelli, M.; Papathanos, P.A.; Thyme, S.B.; Li, H.; Ulge, U.Y.; Crisanti, A. A synthetic homing endonuclease-based gene drive system in the human malaria mosquito. Nature 2011, 473, 212–215. [Google Scholar] [CrossRef]
- Burt, A.; Koufopanou, V. Homing endonuclease genes: The rise and fall and rise again of a selfish element. Curr. Opin. Genet. Dev. 2004, 14, 609–615. [Google Scholar] [CrossRef]
- Rocha-Martins, M.; Cavalheiro, G.R.; Matos-Rodrigues, G.E.; Martins, R.A. From Gene Targeting to Genome Editing: Transgenic animals applications and beyond. Acad. Bras. Cienc. 2015, 87 (Suppl. S2), 1323–1348. [Google Scholar] [CrossRef] [PubMed]
- Sander, J.D.; Joung, J.K. CRISPR-Cas systems for editing, regulating and targeting genomes. Nat. Biotechnol. 2014, 32, 347–355. [Google Scholar] [CrossRef]
- Pratt, J.; Venkatraman, N.; Brinker, A.; Xiao, Y.; Blasberg, J.; Thompson, D.C.; Bourner, M. Use of zinc finger nuclease technology to knock out efflux transporters in C2BBe1 cells. Curr. Protoc. Toxicol. 2012, 52, 23. [Google Scholar] [CrossRef]
- Boch, J.; Bonas, U. Xanthomonas AvrBs3 Family-Type III Effectors: Discovery and Function. Annu. Rev. Phytopathol. 2010, 48, 419–436. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Zhang, H.; Xu, N.; Zhang, B.; Gou, F.; Zhu, J.-K. Application of the CRISPR–Cas system for efficient genome engineering in plants. Mol. Plant 2013, 6, 2008–2011. [Google Scholar] [CrossRef]
- Simoni, A.; Siniscalchi, C.; Chan, Y.-S.; Huen, D.S.; Russell, S.; Windbichler, N.; Crisanti, A. Development of synthetic selfish elements based on modular nucleases in Drosophila melanogaster. Nucleic Acids Res. 2014, 42, 7461–7472. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Zhang, F. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Lander, E.S. The heroes of CRISPR. Cell 2016, 164, 18–28. [Google Scholar] [CrossRef]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the immune system of bacteria and archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Montoliu, L. On the Origin of CRISPR-Cas Technology: From Prokaryotes to Mammals. Trends Microbiol. 2016, 24, 811–820. [Google Scholar] [CrossRef]
- Yao, R.; Liu, D.; Jia, X.; Zheng, Y.; Liu, W.; Xiao, Y. CRISPR-Cas9/Cas12a biotechnology and application in bacteria. Synth. Syst. Biotechnol. 2018, 3, 135–149. [Google Scholar] [CrossRef]
- Deltcheva, E.; Chylinski, K.; Sharma, C.M.; Gonzales, K.; Chao, Y.; Pirzada, Z.A.; Charpentier, E. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 2011, 471, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 2020, 38, 824–844. [Google Scholar] [CrossRef]
- Garneau, J.E.; Dupuis, M.-È.; Villion, M.; Romero, D.A.; Barrangou, R.; Boyaval, P.; Moineau, S. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 2010, 468, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.M.; Musunuru, K. Expanding the genetic editing tool kit: ZFNs, TALENs, and CRISPR-Cas9. J. Clin. Investig. 2014, 124, 4154–4161. [Google Scholar] [CrossRef] [PubMed]
- DiCarlo, J.E.; Chavez, A.; Dietz, S.L.; Esvelt, K.M.; Church, G.M. Safeguarding CRISPR-Cas9 gene drives in yeast. Nat. Biotechnol. 2015, 33, 1250–1255. [Google Scholar] [CrossRef]
- Gantz, V.M.; Bier, E. The mutagenic chain reaction: A method for converting heterozygous to homozygous mutations. Science 2015, 348, 442. [Google Scholar] [CrossRef]
- Gantz, V.M.; Jasinskiene, N.; Tatarenkova, O.; Fazekas, A.; Macias, V.M.; Bier, E.; James, A.A. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proc. Natl. Acad. Sci. USA 2015, 112, E6736–E6743. [Google Scholar] [CrossRef]
- Kyrou, K.; Hammond, A.M.; Galizi, R.; Kranjc, N.; Burt, A.; Beaghton, A.K.; Crisanti, A. A CRISPR–Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nat. Biotechnol. 2018, 36, 1062–1066. [Google Scholar] [CrossRef]
- Gierus, L.; Birand, A.; Bunting, M.D.; Godahewa, G.I.; Piltz, S.G.; Oh, K.P.; Thomas, P.Q. Leveraging a natural murine meiotic drive to suppress invasive populations. Proc. Natl. Acad. Sci. USA 2022, 119, e2213308119. [Google Scholar] [CrossRef]
- Grunwald, H.A.; Gantz, V.M.; Poplawski, G.; Xu, X.-R.S.; Bier, E.; Cooper, K.L. Super-Mendelian inheritance mediated by CRISPR–Cas9 in the female mouse germline. Nature 2019, 566, 105–109. [Google Scholar] [CrossRef]
- Weitzel, A.J.; Grunwald, H.A.; Weber, C.; Levina, R.; Gantz, V.M.; Hedrick, S.M.; Cooper, K.L. Meiotic Cas9 expression mediates gene conversion in the male and female mouse germline. PLoS Biol. 2021, 19, e3001478. [Google Scholar] [CrossRef]
- Galizi, R.; Hammond, A.; Kyrou, K.; Taxiarchi, C.; Bernardini, F.; O’Loughlin, S.M.; Crisanti, A. A CRISPR-Cas9 sex-ratio distortion system for genetic control. Sci. Rep. 2016, 6, 31139. [Google Scholar] [CrossRef] [Green Version]
- Oberhofer, G.; Ivy, T.; Hay, B.A. Behavior of homing endonuclease gene drives targeting genes required for viability or female fertility with multiplexed guide RNAs. Proc. Natl. Acad. Sci. USA 2018, 115, E9343–E9352. [Google Scholar] [CrossRef]
- Yang, E.; Metzloff, M.; Langmüller, A.M.; Xu, X.; Clark, A.G.; Messer, P.W.; Champer, J. A homing suppression gene drive with multiplexed gRNAs maintains high drive conversion efficiency and avoids functional resistance alleles. G3 Genes Genomes Genet. 2022, 12, jkac081. [Google Scholar] [CrossRef]
- Berry, R.; Scriven, P. The house mouse: A model and motor for evolutionary understanding. Biol. J. Linn. Soc. 2005, 84, 335–347. [Google Scholar] [CrossRef]
- Pocock, M.J.; Hauffe, H.C.; Searle, J.B. Dispersal in house mice. Biol. J. Linn. Soc. 2005, 84, 565–583. [Google Scholar] [CrossRef]
- Champer, J.; Buchman, A.; Akbari, O.S. Cheating evolution: Engineering gene drives to manipulate the fate of wild populations. Nat. Rev. Genet. 2016, 17, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, W.D. Extraordinary sex ratios. A sex-ratio theory for sex linkage and inbreeding has new implications in cytogenetics and entomology. Science 1967, 156, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Backus, G.A.; Gross, K. Genetic engineering to eradicate invasive mice on islands: Modeling the efficiency and ecological impacts. Ecosphere 2016, 7, e01589. [Google Scholar] [CrossRef]
- Koopman, P.; Gubbay, J.; Vivian, N.; Goodfellow, P.; Lovell-Badge, R. Male development of chromosomally female mice transgenic for Sry. Nature 1991, 351, 117–121. [Google Scholar] [CrossRef]
- Manser, A.; Cornell, S.J.; Sutter, A.; Blondel, D.V.; Serr, M.; Godwin, J.; Price, T.A.R. Controlling invasive rodents via synthetic gene drive and the role of polyandry. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190852. [Google Scholar] [CrossRef] [PubMed]
- Manser, A.; Lindholm, A.K.; Simmons, L.W.; Firman, R.C. Sperm competition suppresses gene drive among experimentally evolving populations of house mice. Mol. Ecol. 2017, 26, 5784–5792. [Google Scholar] [CrossRef] [Green Version]
- Sutter, A.; Lindholm, A.K. Detrimental effects of an autosomal selfish genetic element on sperm competitiveness in house mice. Proc. R. Soc. B Biol. Sci. USA 2015, 282, 20150974. [Google Scholar] [CrossRef] [PubMed]
- Manser, A.; Lindholm, A.K.; König, B.; Bagheri, H.C. Polyandry and the decrease of a selfish genetic element in a wild house mouse population. Evol. Int. J. Org. Evol. 2011, 65, 2435–2447. [Google Scholar] [CrossRef] [PubMed]
- Hay, B.A.; Guo, M. Gene drive-mediated population elimination for biodiversity conservation. When you come to a fork in the road, take it. Proc. Natl. Acad. Sci. USA 2022, 119, e2218020119. [Google Scholar] [CrossRef] [PubMed]
- Prowse, T.A.A.; Cassey, P.; Ross, J.V.; Pfitzner, C.; Wittmann, T.A.; Thomas, P. Dodging silver bullets: Good CRISPR gene-drive design is critical for eradicating exotic vertebrates. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170799. [Google Scholar] [CrossRef] [PubMed]
- Pfitzner, C.; White, M.A.; Piltz, S.G.; Scherer, M.; Adikusuma, F.; Hughes, J.N.; Thomas, P.Q. Progress Toward Zygotic and Germline Gene Drives in Mice. CRISPR J. 2020, 3, 388–397. [Google Scholar] [CrossRef]
- Prowse, T.A.A.; Adikusuma, F.; Cassey, P.; Thomas, P.; Ross, J.V. A Y-chromosome shredding gene drive for controlling pest vertebrate populations. Elife 2019, 8, e41873. [Google Scholar] [CrossRef]
- Bull, J. Evolutionary decay and the prospects for long-term disease intervention using engineered insect vectors. Evol. Med. Public Health 2015, 2015, 152–166. [Google Scholar] [CrossRef]
- Champer, J.; Kim, I.K.; Champer, S.E.; Clark, A.G.; Messer, P.W. Performance analysis of novel toxin-antidote CRISPR gene drive systems. BMC Biol. 2020, 18, 27. [Google Scholar] [CrossRef] [PubMed]
- Hammond, A.; Kyrou, K.; Bruttini, M.; North, A.; Galizi, R.; Karlsson, X.; Crisanti, A. The creation and selection of mutations resistant to a gene drive over multiple generations in the malaria mosquito. PLoS Genet. 2017, 13, e1007039. [Google Scholar] [CrossRef] [PubMed]
- Sudweeks, J.; Hollingsworth, B.; Blondel, D.V.; Campbell, K.J.; Dhole, S.; Eisemann, J.D.; Lloyd, A.L. Locally Fixed Alleles: A method to localise gene drive to island populations. Sci. Rep. 2019, 9, 15821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, K.; Burt, A. Double drives and private alleles for localised population genetic control. PLoS Genet. 2021, 17, e1009333. [Google Scholar] [CrossRef]
- Champer, J.; Liu, J.; Oh, S.Y.; Reeves, R.; Luthra, A.; Oakes, N.; Messer, P. Reducing resistance allele formation in CRISPR gene drive. Proc. Natl. Acad. Sci. USA 2018, 115, 5522–5527. [Google Scholar] [CrossRef]
- Champer, S.; Oh, S.Y.; Liu, C.; Wen, Z.; Clark, A.; Messer, P.; Champer, J. Computational and experimental performance of CRISPR homing gene drive strategies with multiplexed gRNAs. Sci. Adv. 2020, 6, eaaz0525. [Google Scholar] [CrossRef]
- Champer, J.; Reeves, R.; Oh, S.Y.; Liu, C.; Liu, J.; Clark, A.G.; Messer, P.W. Novel CRISPR/Cas9 gene drive constructs reveal insights into mechanisms of resistance allele formation and drive efficiency in genetically diverse populations. PLoS Genet. 2017, 13, e1006796. [Google Scholar] [CrossRef]
- Metzloff, M.; Yang, E.; Dhole, S.; Clark, A.G.; Messer, P.W.; Champer, J. Experimental demonstration of tethered gene drive systems for confined population modification or suppression. BMC Biol. 2022, 20, 119. [Google Scholar] [CrossRef]
- Min, J.; Noble, C.; Najjar, D.; Esvelt, K.M. Daisy quorum drives for the genetic restoration of wild populations. BioRxiv 2017, 115618. [Google Scholar] [CrossRef]
- Min, J.; Noble, C.; Najjar, D.; Esvelt, K.M. Daisyfield gene drive systems harness repeated genomic elements as a generational clock to limit spread. BioRxiv 2017, 104877. [Google Scholar] [CrossRef]
- Noble, C.; Min, J.; Olejarz, J.; Buchthal, J.; Chavez, A.; Smidler, A.L.; Esvelt, K.M. Daisy-chain gene drives for the alteration of local populations. Proc. Natl. Acad. Sci. USA 2019, 116, 8275–8282. [Google Scholar] [CrossRef]
- Oberhofer, G.; Ivy, T.; Hay, B.A. Cleave and Rescue, a novel selfish genetic element and general strategy for gene drive. Proc. Natl. Acad. Sci. USA 2019, 116, 6250–6259. [Google Scholar] [CrossRef]
- Esvelt, K.M.; Gemmell, N. Conservation demands safe gene drive. PLoS Biol. 2017, 15, e2003850. [Google Scholar] [CrossRef] [PubMed]
- Esvelt, K.M. Daisy Drive: Safe Genes Research Plan. Responsive Science. Available online: https://www.responsivescience.org/pub/daisy-drive (accessed on 20 November 2020).
- Esvelt, K.M. Daisy Drive Systems. MIT Media Lab. Available online: http://www.sculptingevolution.org/daisydrives (accessed on 20 November 2020).
- Emerson, C.; James, S.; Littler, K.; Randazzo, F. Principles for gene drive research. Science 2017, 358, 1135–1136. [Google Scholar] [CrossRef]
- Long Kanya, C.; Alphey, L.; Annas George, J.; Bloss Cinnamon, S.; Campbell Karl, J.; Champer, J.; Akbari Omar, S. Core commitments for field trials of gene drive organisms. Science 2020, 370, 1417–1419. [Google Scholar] [CrossRef]
- Stelmach, A.; Nerlich, B.; Hartley, S. Gene Drives in the UK, US, and Australian Press (2015–2019): How a New Focus on Responsibility Is Shaping Science Communication. Sci. Commun. 2022, 44, 143–168. [Google Scholar] [CrossRef] [PubMed]
- Buchthal, J.; Evans, S.W.; Lunshof, J.; Telford, S.R.; Esvelt, K.M. Mice Against Ticks: An experimental community-guided effort to prevent tick-borne disease by altering the shared environment. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180105. [Google Scholar] [CrossRef] [PubMed]
- Funk, C.; Hefferon, M. Most Americans Accept Genetic Engineering of Animals That Benefits Human Health, but Many Oppose Other Uses. Available online: https://www.pewinternet.org/wp-content/uploads/sites/9/2018/08/PS_2018.08.16_biotech-animals_FINAL.pdf (accessed on 18 January 2022).
- Teem, J.L.; Alphey, L.; Descamps, S.; Edgington, M.P.; Edwards, O.; Gemmell, N.; Roberts, A. Genetic Biocontrol for Invasive Species. Front. Bioeng. Biotechnol. 2020, 8, 452. [Google Scholar] [CrossRef]
- Brossard, D.; Belluck, P.; Gould, F.; Wirz, C.D. Promises and perils of gene drives: Navigating the communication of complex, post-normal science. Proc. Natl. Acad. Sci. USA 2019, 116, 7692. [Google Scholar] [CrossRef]
- Kelsey, A.; Stillinger, D.; Pham, T.B.; Murphy, J.; Firth, S.; Carballar-Lejarazú, R. Global Governing Bodies: A Pathway for Gene Drive Governance for Vector Mosquito Control. Am. J. Trop. Med. Hyg. 2020, 103, 976–985. [Google Scholar] [CrossRef]
- UNCBD Ad Hoc Technical Expert Group (AHTEG). Report of the Ad Hoc Technical Expert Group on Risk Assessment. Available online: https://www.cbd.int/doc/c/a763/e248/4fa326e03e3c126b9615e95d/cp-ra-ahteg-2020-01-05-en.pdf (accessed on 12 May 2021).
Figure 1.
Examples of the three known primary mechanisms by which selfish genetic elements achieve ‘drive’. (a). Interference. A killer gene (K) can kill non-K-bearing sperm. This would benefit the K gene’s chance of fertilising an egg. (b). Over-replication. A transposable element (T) makes a second copy of itself, and one daughter cell inherits one copy while the other inherits two copies. (c). Gonotaxis. A B-chromosome replicates itself, and both copies move to the germline and avoid the somatic cells. Image adapted from Burt and Trivers [11].
Figure 1.
Examples of the three known primary mechanisms by which selfish genetic elements achieve ‘drive’. (a). Interference. A killer gene (K) can kill non-K-bearing sperm. This would benefit the K gene’s chance of fertilising an egg. (b). Over-replication. A transposable element (T) makes a second copy of itself, and one daughter cell inherits one copy while the other inherits two copies. (c). Gonotaxis. A B-chromosome replicates itself, and both copies move to the germline and avoid the somatic cells. Image adapted from Burt and Trivers [11].

Figure 2.
Mendelian and gene drive inheritance. Left: Mendelian inheritance of an altered gene. Right: Inheritance of a gene drive. Image adapted from McFarlane, Whitelaw, and Lillico [27].
Figure 2.
Mendelian and gene drive inheritance. Left: Mendelian inheritance of an altered gene. Right: Inheritance of a gene drive. Image adapted from McFarlane, Whitelaw, and Lillico [27].

Figure 3.
Nuclease-assisted genetic engineering. A double-strand DNA break (DSB) from a site-specific nuclease could lead to precise modification through homology-directed repair (HDR) in the presence of a DNA repair template, in the form of double-stranded DNA or a single-strand DNA oligonucleotide, both of which must contain homology arms. DSBs can also be repaired through non-homologous end joining (NHEJ), which frequently leads to small insertions and deletions (indels), or microhomology-mediated end-joining (MMEJ), which often results in deletion.
Figure 3.
Nuclease-assisted genetic engineering. A double-strand DNA break (DSB) from a site-specific nuclease could lead to precise modification through homology-directed repair (HDR) in the presence of a DNA repair template, in the form of double-stranded DNA or a single-strand DNA oligonucleotide, both of which must contain homology arms. DSBs can also be repaired through non-homologous end joining (NHEJ), which frequently leads to small insertions and deletions (indels), or microhomology-mediated end-joining (MMEJ), which often results in deletion.

Figure 4.
A schematic representation of Streptococcus pyogenes Cas9 (SpCas9). SpCas9 is programmed here with a single guide RNA (sgRNA). The sgRNA comprises a site-specific CRISPR RNA (crRNA; black) and an auxiliary transactivating crRNA (tracrRNA; pink). Two nuclease domains (RuvC and HNH) both cut one DNA strand 3 bp upstream from the protospacer adjacent motif (PAM; 5’-NGG; red) to generate a double-strand DNA break. Image created with Biorender.com.
Figure 4.
A schematic representation of Streptococcus pyogenes Cas9 (SpCas9). SpCas9 is programmed here with a single guide RNA (sgRNA). The sgRNA comprises a site-specific CRISPR RNA (crRNA; black) and an auxiliary transactivating crRNA (tracrRNA; pink). Two nuclease domains (RuvC and HNH) both cut one DNA strand 3 bp upstream from the protospacer adjacent motif (PAM; 5’-NGG; red) to generate a double-strand DNA break. Image created with Biorender.com.

Figure 5.
X-Shredder (XS). During sperm production, Cas nuclease and guide RNA(s) are expressed from the XS cassette (orange) located on the Y-chromosome (Y) shred the X-chromosome (X) beyond repair. This results in a bias towards the production of Y-bearing sperm and most offspring being XS males.
Figure 5.
X-Shredder (XS). During sperm production, Cas nuclease and guide RNA(s) are expressed from the XS cassette (orange) located on the Y-chromosome (Y) shred the X-chromosome (X) beyond repair. This results in a bias towards the production of Y-bearing sperm and most offspring being XS males.

Figure 6.
CRISPR-based homing gene drive (GD). The GD cassette (purple) expresses Cas and one or more guide RNAs that cut the sister allele on the homologous wild-type (WT) chromosome. The cell then repairs the cut by homology-direct repair (HDR), using the GD chromosome as the repair template. This process copies the GD cassette and any genetic payload (trait) onto the WT chromosome and ensures that most of the mature gametes in the animal carry the GD cassette, passing on to most or all the offspring. With this strategy, the guide RNAs must not cut the GD cassette.
Figure 6.
CRISPR-based homing gene drive (GD). The GD cassette (purple) expresses Cas and one or more guide RNAs that cut the sister allele on the homologous wild-type (WT) chromosome. The cell then repairs the cut by homology-direct repair (HDR), using the GD chromosome as the repair template. This process copies the GD cassette and any genetic payload (trait) onto the WT chromosome and ensures that most of the mature gametes in the animal carry the GD cassette, passing on to most or all the offspring. With this strategy, the guide RNAs must not cut the GD cassette.

Figure 7.
Daisy-chains drive system. (a) A daisy chain separates the CRISPR components throughout the genome, but these components functionally operate so that each daisy element drives the next element in the chain. The element at the end of the chain, in this case, C is not copied and is lost in half of offspring. In those animals that have lost C, B is no longer copied and is lost; this process continues until the drive system is inactivated. (b) The loss of nondriving elements to mendelian inheritance is analogous to a rocket running out of fuel. Adding more elements (genetic fuel) to the daisy chain allows the system to spread further before it runs out of genetic fuel and no longer spreads in the population. Image from Esvelt & Gemmell [98] (CC BY 4.0).
Figure 7.
Daisy-chains drive system. (a) A daisy chain separates the CRISPR components throughout the genome, but these components functionally operate so that each daisy element drives the next element in the chain. The element at the end of the chain, in this case, C is not copied and is lost in half of offspring. In those animals that have lost C, B is no longer copied and is lost; this process continues until the drive system is inactivated. (b) The loss of nondriving elements to mendelian inheritance is analogous to a rocket running out of fuel. Adding more elements (genetic fuel) to the daisy chain allows the system to spread further before it runs out of genetic fuel and no longer spreads in the population. Image from Esvelt & Gemmell [98] (CC BY 4.0).

Figure 8.
Daisyfield system. Daisyfield drive systems scatter multiple daisy elements encoding the same guide RNAs. (a) The simple Daisyfield design depicted has four elements that target the wild-type allele where the Cas nuclease and genetic payload are to be inserted. Cutting and subsequent homology-directed repair (HDR) copies the payload and Cas nuclease into the cut site in the wild type allele. (b) Daisyfield drives are equivalent to using multiple parallel rocket boosters, half of which run out of genetic fuel and are lost to Mendelian inheritance with each generation of mating to wild-type animals. Image from Esvelt [100] (CC BY 4.0).
Figure 8.
Daisyfield system. Daisyfield drive systems scatter multiple daisy elements encoding the same guide RNAs. (a) The simple Daisyfield design depicted has four elements that target the wild-type allele where the Cas nuclease and genetic payload are to be inserted. Cutting and subsequent homology-directed repair (HDR) copies the payload and Cas nuclease into the cut site in the wild type allele. (b) Daisyfield drives are equivalent to using multiple parallel rocket boosters, half of which run out of genetic fuel and are lost to Mendelian inheritance with each generation of mating to wild-type animals. Image from Esvelt [100] (CC BY 4.0).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
McFarlane, G.R.; Whitelaw, C.B.A.; Lillico, S.G. Gene Drive: Past, Present and Future Roads to Vertebrate Biocontrol. Appl. Biosci. 2023, 2, 52-70. https://doi.org/10.3390/applbiosci2010006
AMA Style
McFarlane GR, Whitelaw CBA, Lillico SG. Gene Drive: Past, Present and Future Roads to Vertebrate Biocontrol. Applied Biosciences. 2023; 2(1):52-70. https://doi.org/10.3390/applbiosci2010006
Chicago/Turabian StyleMcFarlane, Gus R., C. Bruce A. Whitelaw, and Simon G. Lillico. 2023. "Gene Drive: Past, Present and Future Roads to Vertebrate Biocontrol" Applied Biosciences 2, no. 1: 52-70. https://doi.org/10.3390/applbiosci2010006