

Acidogenesis of Pentose Liquor to Produce Biohydrogen and Organic Acids Integrated with 1G–2G Ethanol Production in Sugarcane Biorefineries
 , , , , and
, , , , and
Abstract
:
1. Introduction
2. Materials and Methods
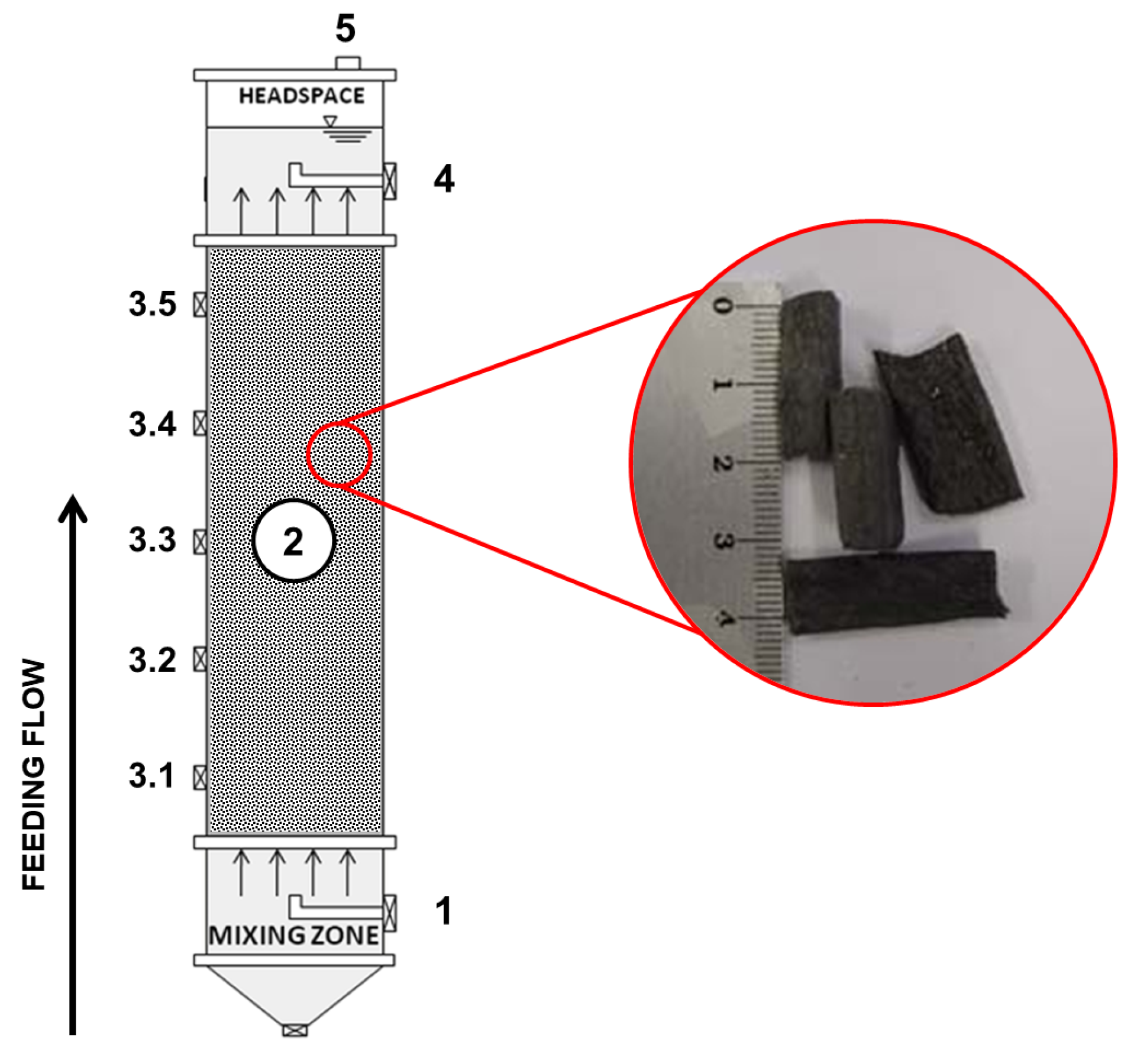
2.1. Bioreactor and Support Material
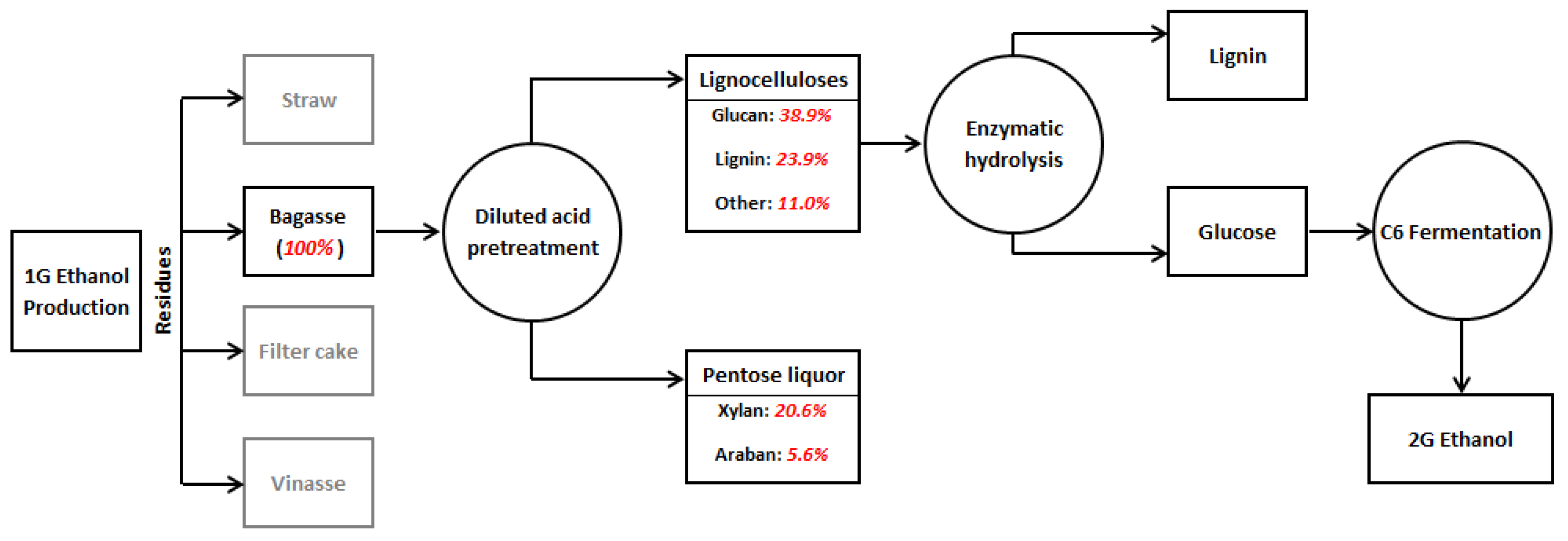
2.2. Substrates
2.3. Experimental Procedure
2.4. Monitoring Procedure and Analytical Methods
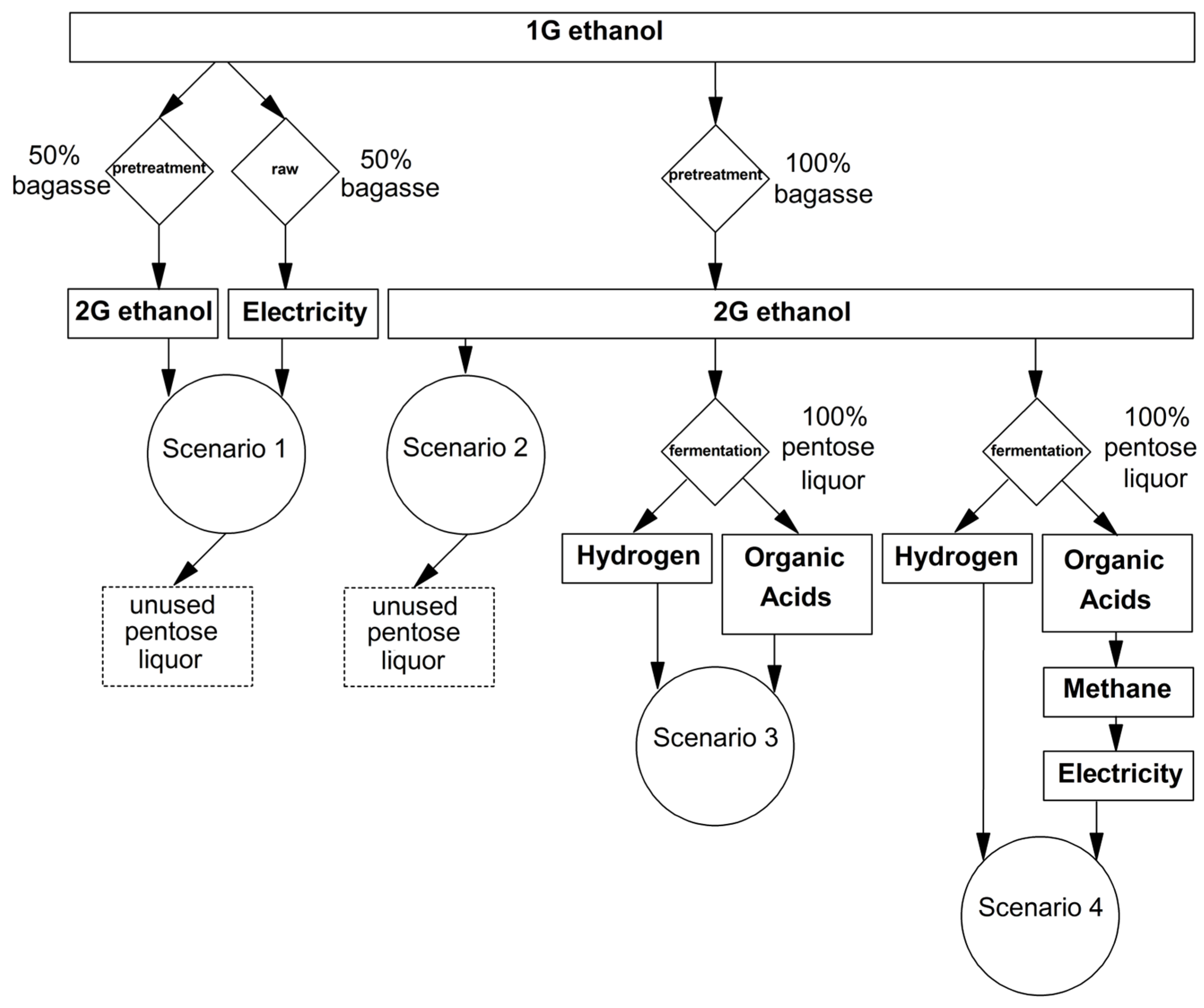
2.5. Scenario Assessment
3. Results and Discussion
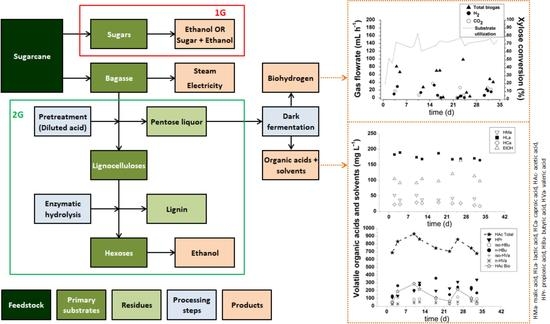
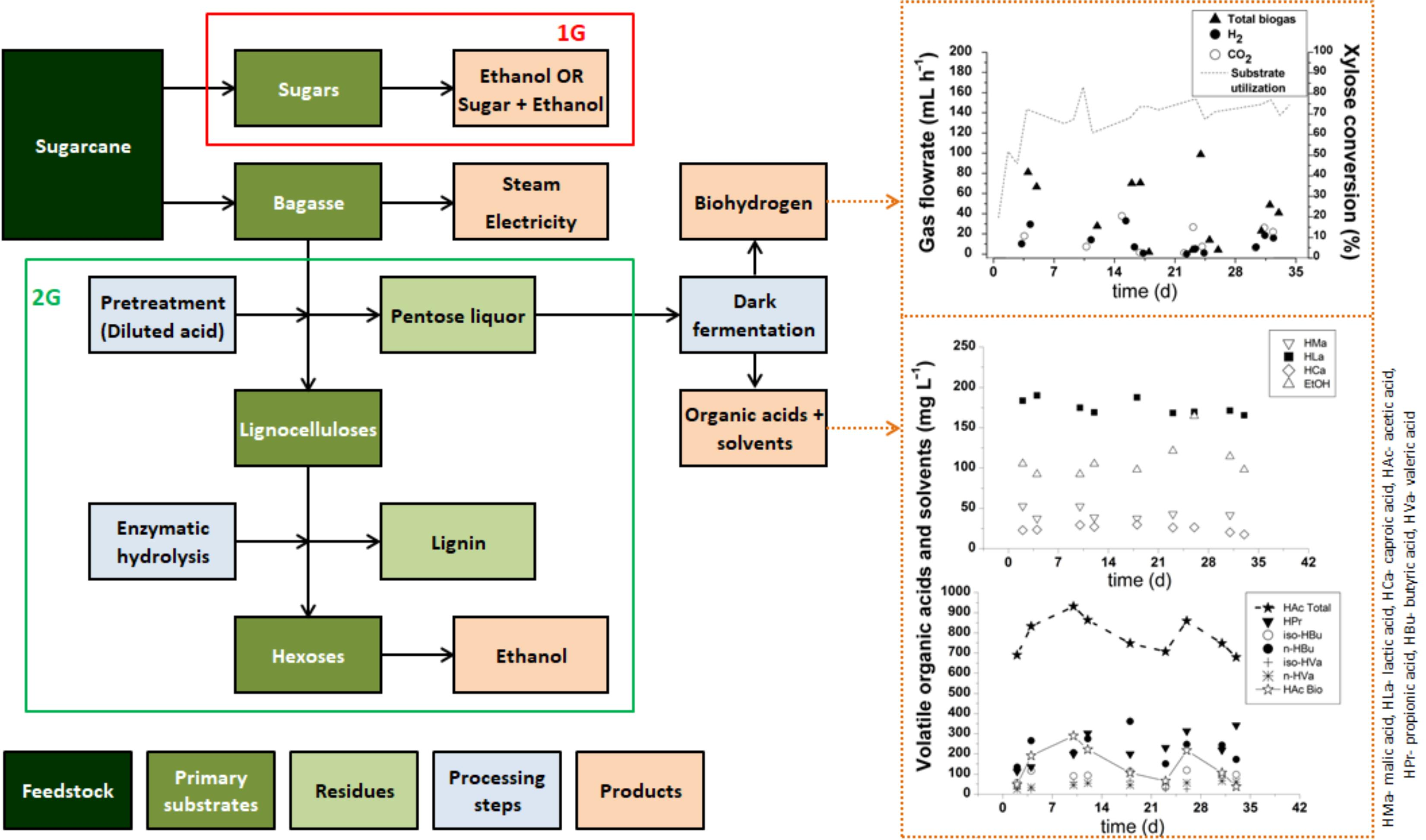
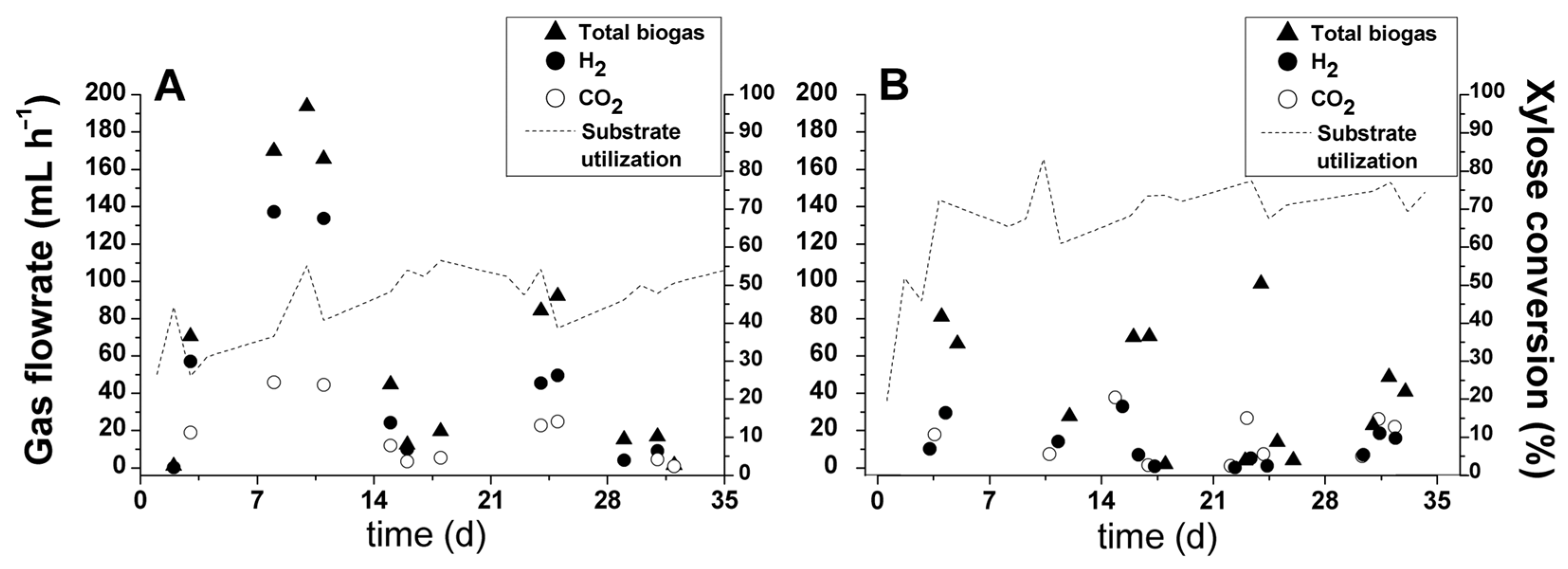
3.1. Biohydrogen Production
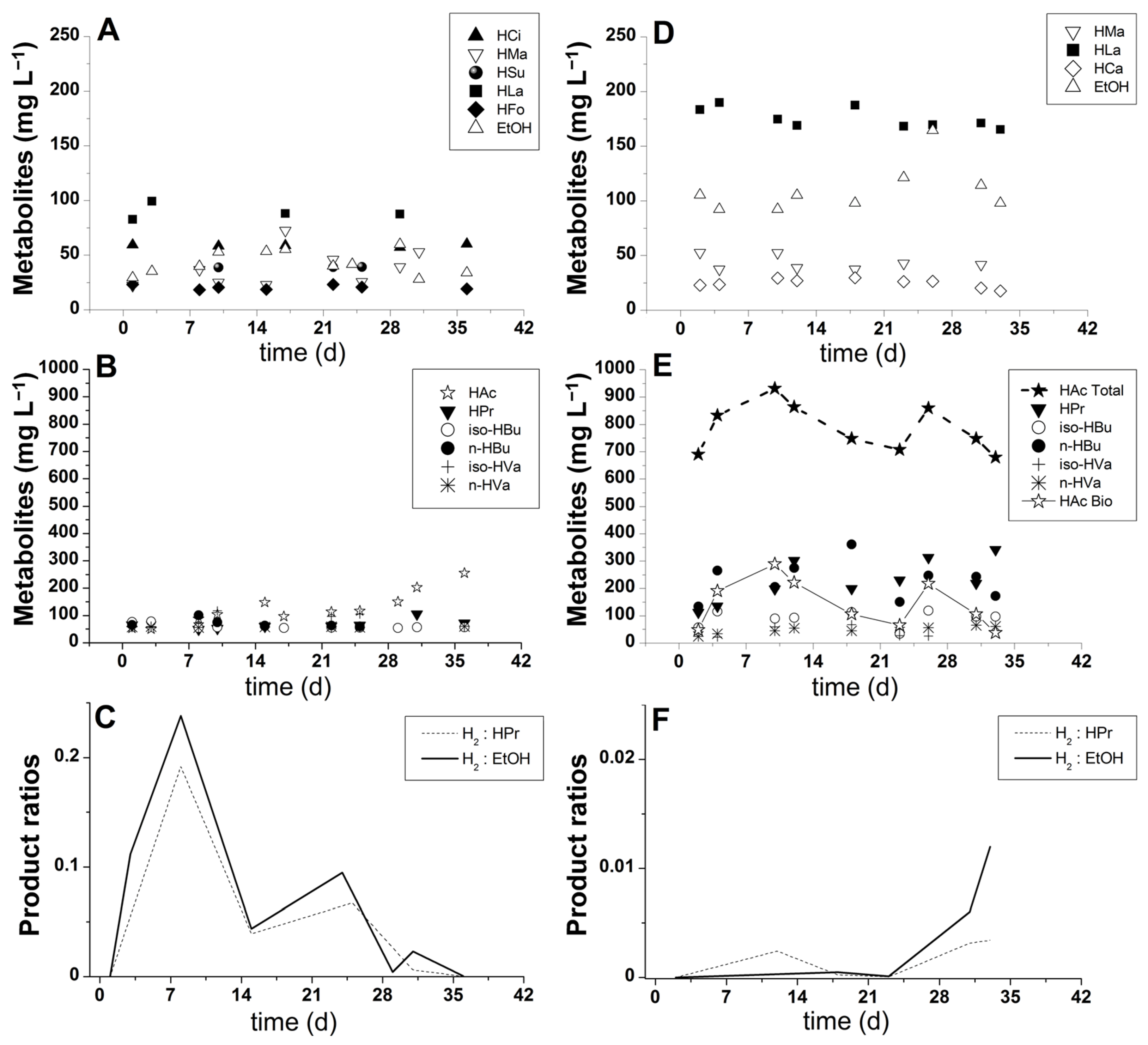
3.2. Organic Acid Production
3.3. Preliminary Analysis of Biohydrogen, VOA and Cogeneration Potential in a Pentose Liquor-Based Biorefinery
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lago, A.C.; Bonomi, A.; Cavalett, O.; Cunha, M.P.; Lima, M.A. Sugarcane as a carbon source: The Brazilian case. Biomass Bioenergy 2012, 46, 5–12. [Google Scholar] [CrossRef]
- Poggi-Varaldo, H.M.; Munoz-Paez, K.M.; Escamilla-Alvarado, C.; Robledo-Narváez, P.N.; Ponce-Noyola, M.T.; Calva-Calva, G.; Ríos-Leal, E.; Galíndez-Mayer, J.; Estrada-Vázquez, C.; Ortega-Clemente, A.; et al. Biohydrogen, biomethane and bioelectricity as crucial components of biorefinery of organic wastes: A review. Waste Manag. Res. 2014, 32, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Morais, E.R.; Junqueira, T.L.; Sampaio, I.L.M.; Dias, M.O.S.; Rezende, M.C.A.F.; Jesus, C.D.F.; Klein, B.C.; Gómez, E.O.; Mantelatto, P.E.; Maciel Filho, R.; et al. Biorefinery alternatives. In Virtual biorefinery: An Optimization Strategy for Renewable Carbon Valorization; Bonomi, A., Cavalett, O., Cunha, M.P., Lima, M.A.P., Eds.; Springer: London, UK, 2016; pp. 53–132. [Google Scholar]
- Dias, M.O.S.; Junqueira, T.L.; Sampaio, I.L.M.; Chagas, M.F.; Watanabe, M.D.B.; Morais, E.R.; Gouveia, V.L.R.; Klein, B.C.; Rezende, M.C.A.F.; Cardoso, T.F.; et al. Use of the VSB to assess biorefinery strategies. In Virtual biorefinery: An Optimization Strategy for Renewable Carbon Valorization; Bonomi, A., Cavalett, O., Cunha, M.P., Lima, M.A.P., Eds.; Springer: London, UK, 2016; pp. 189–256. [Google Scholar]
- Rabelo, S.C.; Carrere, H.; Maciel Filho, R.; Costa, A.C. Production of bioethanol, methane and heat from sugarcane bagasse in a biorefinery concept. Bioresour. Technol. 2011, 102, 7887–7895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CONAB. Acompanhamento da Safra Brasileira de Cana-de-Açúcar; Safra 2023/24—Primeiro Levantamento; CONAB: Brasília, Brasil, 2023. Available online: https://www.conab.gov.br/info-agro/safras/cana/boletim-da-safra-de-cana-de-acucar/item/download/47193_22dfa86f63aa61cd8e405419b30d9262 (accessed on 27 June 2023). (In Portuguese)
- Dias, M.O.S.; Junqueira, T.L.; Cavalett, O.; Pavanello, L.G.; Cunha, M.P.; Jesus, C.D.F.; Maciel Filho, R.; Bonomi, A. Biorefineries for the production of first and second generation ethanol and electricity from sugarcane. Appl. Energy 2013, 109, 72–78. [Google Scholar] [CrossRef]
- Dias, M.O.S.; Junqueira, T.L.; Rossel, C.E.; Maciel Filho, R.; Bonomi, A. Evaluation of process configurations for second generation integrated with first generation bioethanol production from sugarcane. Fuel Process. Technol. 2013, 109, 84–89. [Google Scholar] [CrossRef]
- Zhou, M.; Lü, X. Strategies on simultaneous fermentation of pentose and hexose to bioethanol. In Advances in 2nd Generation of Bioethanol Production; Lü, X., Ed.; Woodhead Publishing: Duxford, UK, 2021; pp. 161–211. [Google Scholar]
- Sriariyanun, M.; Gundupalli, M.P.; Phakeenuya, V.; Phusamtisampan, T.; Cheng, Y.S.; Venkatachalam, P. Biorefinery Approaches For Production Of Cellulosic Ethanol Fuel Using Recombinant Engineered Microorganisms. J. Appl. Sci. Eng. 2023, 27, 1985–2005. [Google Scholar]
- Ghimire, A.; Frunzo, L.; Pirozzi, F.; Trably, E.; Escudie, R.; Lens, P.N.L.; Esposito, G. A review on dark fermentative biohydrogen production from organic biomass: Process parameters and use of by-products. Appl. Energy 2015, 144, 73–95. [Google Scholar] [CrossRef]
- Elbeshbishy, E.; Dhar, B.R.; Nakhla, G.; Lee, H.S. A critical review on inhibition of dark biohydrogen fermentation. Renew. Sustain. Energy Rev. 2017, 79, 656–668. [Google Scholar] [CrossRef]
- Hawkes, F.R.; Dinsdale, R.; Hawkes, D.L.; Hussy, I. Sustainable fermentative hydrogen production: Challenges for process optimization. Int. J. Hydrogen Energy 2002, 27, 1339–1347. [Google Scholar] [CrossRef]
- Show, K.Y.; Lee, D.J.; Tay, J.H.; Lin, C.Y.; Chang, J.S. Biohydrogen production: Current perspectives and the way forward. Int. J. Hydrogen Energy 2012, 37, 15616–15631. [Google Scholar] [CrossRef]
- Das, D.; Veziroglu, T. Hydrogen production by biological processes: A survey of literature. Int. J. Hydrogen Energy 2001, 26, 13–28. [Google Scholar] [CrossRef]
- Leite, J.A.C.; Fenandes, B.S.; Pozzi, E.; Barboza, M.; Zaiat, M. Application of an anaerobic packed-bed biobioreactor for the production of hydrogen and organic acids. Int. J. Hydrogen Energy 2008, 33, 579–586. [Google Scholar] [CrossRef]
- Ramos, L.R.; Silva, E.L. Thermophilic hydrogen and methane production from sugarcane stillage in two-stage anaerobic fluidized bed reactors. Int. J. Hydrogen Energy 2020, 45, 5239–5251. [Google Scholar] [CrossRef]
- Ferreira, T.B.; Rego, G.C.; Ramos, L.R.; Menezes, C.A.; Silva, E.L. Improved dark fermentation of cane molasses in mesophilic and thermophilic anaerobic fluidized bed reactors by selecting operational conditions. Int. J. Energy Res. 2020, 44, 10442–10452. [Google Scholar] [CrossRef]
- Fuess, L.T.; Braga, A.F.M.; Eng, F.; Gregoracci, G.; Saia, F.T.; Zaiat, M.; Lens, P.N.L. Solving the bottlenecks of sugarcane vinasse biodigestion: Impacts of temperature and substrate exchange on sulfate removal during dark fermentation. Chem. Eng. J. 2023, 455, 140965. [Google Scholar] [CrossRef]
- Menezes, C.A.; Almeida, P.S.; Delforno, T.P.; Oliveira, V.M.; Sakamoto, I.K.; Varesche, M.B.A.; Silva, E.L. Relating biomass composition and the distribution of metabolic functions in the co-fermentation of sugarcane vinasse and glycerol. Int. J. Hydrogen Energy 2023, 48, 8837–8853. [Google Scholar] [CrossRef]
- Kim, H.; Jeon, B.S.; Sang, B.I. An efficient new process for the selective production of odd-chain carboxylic acids by simple carbon elongation using Megasphaera hexanoica. Sci. Rep. 2019, 9, 11999. [Google Scholar] [CrossRef] [Green Version]
- Bastidas-Oyanedel, J.R.; Schmidt, J.E. Increasing profits in food waste biorefinery—A techno-economic analysis. Energies 2018, 11, 1551. [Google Scholar] [CrossRef] [Green Version]
- Brady, G.S.; Clauser, H.R.; Vaccari, J.A. Materials Handbook, 15th ed.; McGraw-Hill Professional: New York, NY, USA, 2002. [Google Scholar]
- Napoothiri, K.M.; Nair, N.R.; John, R.P. An overview of the recent developments in polylactide (PLA) research. Bioresour Technol. 2010, 101, 6493–8501. [Google Scholar]
- Oliveira, G.H.D.; Niz, M.Y.K.; Zaiat, M.; Rodrigues, J.A.D. Effects of organic loading rate on polyhydroxyalkanoate production from sugarcane stillage by mixed microbial cultures. Appl. Biochem. Biotechnol. 2019, 189, 1039–1055. [Google Scholar] [CrossRef]
- Pattra, S.; Sangyoka, S.; Boonmee, M.; Reungsang, A. Bio-hydrogen Production from the Fermentation of Sugarcane Bagasse Hydrolysate by Clostridium butyricum. Int. J. Hydrogen Energy 2008, 33, 5256–5265. [Google Scholar] [CrossRef]
- Kongjan, P.; Min, B.; Angelidaki, I. Biohydrogen production from xylose at extreme thermophilic temperatures (70 ºC) by mixed culture fermentation. Water Res. 2009, 43, 1414–1424. [Google Scholar] [CrossRef] [PubMed]
- Baêta, B.E.L.; Lima, D.R.S.; Balena Filho, J.G.; Adarme, O.F.H.; Gurgel, L.V.A.; Aquino, S.F. Evaluation of hydrogen and methane production from sugarcane bagasse hemicellulose hydrolysates by two-stage anaerobic digestion process. Bioresour Technol. 2016, 218, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Thungklin, P.; Sittijunda, S.; Reungsang, A. Sequential fermentation of hydrogen and methane from steam-exploded sugarcane bagasse hydrolysate. Int. J. Hydrogen Energy 2018, 43, 9924–9934. [Google Scholar] [CrossRef]
- Chatterjee, S.; Mohan, S.V. Simultaneous production of green hydrogen and bioethanol from segregated sugarcane bagasse hydrolysate streams with circular biorefinery design. Chem. Eng. J. 2021, 425, 130386. [Google Scholar] [CrossRef]
- Fangkum, A.; Reungsang, A. Biohydrogen production from mixed xylose/arabinose at thermophilic temperature by anaerobic mixed cultures in elephant dung. Int. J. Hydrogen Energy 2011, 36, 13928–13938. [Google Scholar] [CrossRef]
- Maintinguer, S.I.; Fernandes, B.S.; Duarte, I.C.S.; Saavedra, N.K.; Adorno, M.A.T.; Varesche, M.B.A. Fermentative hydrogen production with xylose by Clostridium and Klebsiella species in anaerobic batch bioreactors. Int. J. Hydrogen Energy 2011, 36, 13508–13517. [Google Scholar] [CrossRef]
- Lin, C.; Cheng, C. Fermentative hydrogen production from xylose using anaerobic mixed microflora. Int. J. Hydrogen Energy 2006, 31, 832–840. [Google Scholar] [CrossRef]
- Lin, C.; Wu, C.; Hung, C. Temperature effects on fermentative hydrogen production from xylose using mixed anaerobic cultures. Int. J. Hydrogen Energy 2008, 33, 43–50. [Google Scholar] [CrossRef]
- Zhao, C.; Karakashev, D.; Lu, W.; Wang, H.; Angelidaki, I. Xylose fermentation to biofuels (hydrogen and ethanol) by extreme thermophilic (70 °C) mixed culture. Int. J. Hydrogen Energy 2010, 5, 3415–3422. [Google Scholar] [CrossRef]
- Wu, S.Y.; Lin, C.Y.; Lee, K.S.; Hung, C.H.; Chang, J.S.; Lin, P.J.; Chang, F.Y. Dark Fermentative Hydrogen Production from Xylose in Different Bioreactors Using Sewage Sludge Microflora. Energy Fuels 2008, 22, 113–119. [Google Scholar] [CrossRef]
- Dessi, P.; Lakaniemi, A.M.; Lens, P.N.L. Biohydrogen production from xylose by fresh and digested activated sludge at 37, 55 and 70 °C. Water Res. 2017, 115, 120–129. [Google Scholar] [CrossRef]
- Baik, J.H.; Jung, J.H.; Sim, Y.B.; Park, J.H.; Kim, S.M.; Yang, J.; Kim, S.H. High-rate biohydrogen production from xylose using a dynamic membrane bioreactor. Bioresour. Technol. 2022, 344, 126205. [Google Scholar] [CrossRef]
- Silva, V.; Rabelo, C.A.B.S.; Camargo, F.P.; Sakamoto, I.K.; Silva, E.L.; Varesche, M.B.A. Optimization of Key Factors Affecting Hydrogen and Ethanol Production from Xylose by Thermoanaerobacterium calidifontis VCS1 Isolated from Vinasse Treatment Sludge. Waste Biomass Valorization 2022, 13, 1897–1912. [Google Scholar] [CrossRef]
- Lima, D.M.F.; Zaiat, M. The influence of the degree of back-mixing on hydrogen production in an anaerobic fixed-bed reactor. Int. J. Hydrogen Energy 2012, 37, 9630–9635. [Google Scholar] [CrossRef]
- Peixoto, G.; Saavedra, N.K.; Varesche, M.B.A.; Zaiat, M. Hydrogen production from soft-drink wastewater in an upflow anaerobic packed-bed bioreactor. Int. J. Hydrogen Energy 2011, 36, 8953–8966. [Google Scholar] [CrossRef]
- Penteado, E.D.; Lazaro, C.Z.; Sakamoto, I.K.; Zaiat, M. Influence of seed sludge and pretreatment method on hydrogen production in packed-bed anaerobic reactors. Int. J.Hydrogen Energy 2013, 38, 6137–6145. [Google Scholar] [CrossRef]
- Ferraz Jr, A.D.N.; Etchebehere, C.; Zaiat, M. High organic loading rate on thermophilic hydrogen production and metagenomic study at an anaerobic packed-bed reactor treating a residual liquid stream of a Brazilian biorefinery. Bioresour Technol. 2015, 186, 81–88. [Google Scholar] [CrossRef]
- Fuess, L.T.; Kiyuna, L.S.M.; Garcia, M.L.; Zaiat, M. Operational strategies for long-term biohydrogen production from sugarcane stillage in continuous acidogenic packed-bed reactor. Int. J. Hydrogen Energy 2016, 41, 8132–8145. [Google Scholar] [CrossRef] [Green Version]
- Corbari, S.D.M.L.; Andreani, C.L.; Torres, D.G.B.; Eng, F.; Gomes, S.D. Strategies to improve the biohydrogen production from cassava wastewater in fixed-bed reactors. Int. J. Hydrogen Energy 2019, 44, 17214–17223. [Google Scholar] [CrossRef]
- Aguilar, R.; Ramirez, J.A.; Garrote, G.; Vazquez, M. Kinetic study of the acid hydrolysis of sugarcane bagasse. J. Food Eng. 2002, 55, 309–318. [Google Scholar] [CrossRef]
- APHA; AWWA; WEF. Standard Methods for the Examination of Water and Wastewater, 22nd ed.; APHA: Washington, DC, USA, 2012. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric methods for determination of sugar and related substance. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Mockaitis, G.; Rodrigues, J.A.D.; Foresi, E.; Zaiat, M. Toxic effects of cadmium (Cd2+) on anaerobic biomass: Kinetic and metabolic implications. J. Environ. Manag. 2012, 106, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Perna, V.; Castelló, E.; Wenzel, J.; Zampol, C.; Lima, D.M.F.; Borzacconi, L.; Varesche, M.B.; Zaiat, M.; Etchebehere, C. Hydrogen production in an upflow anaerobic packed bed reactor used to treat cheese whey. Int. J. Hydrogen Energy 2013, 38, 54–62. [Google Scholar] [CrossRef]
- Ferraz, A.D.N., Jr.; Etchebehere, C.; Zaiat, M. Mesophilic hydrogen production in acidogenic packed-bed reactors (APBR) using raw sugarcane vinasse as substrate: Influence of support materials. Anaerobe 2015, 34, 94–105. [Google Scholar]
- Lima, D.M.F.; Moreira, W.K.; Zaiat, M. Comparison of the use of sucrose and glucose as a substrate for hydrogen production in an upflow anaerobic fixed-bed reactor. Int. J. Hydrogen Energy 2013, 38, 15074–15083. [Google Scholar] [CrossRef]
- Anzola-Rojas, M.P.; Fonseca, S.G.; Silva, C.C.; Oliveira, V.M.; Zaiat, M. The use of the carbon/nitrogen ratio and specific organic loading rate as tools for improving biohydrogen production in fixed-bed reactors. Biotechnol. Rep. 2015, 5, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Saady, N.M.C. Homoacetogenesis during hydrogen production by mixed cultures dark fermentation. Int. J. Hydrogen Energy 2013, 38, 13172–13191. [Google Scholar] [CrossRef]
- Ferreira, T.B.; Rego, G.C.; Ramos, L.R.; Menezes, C.A.; Soares, L.A.; Sakamoto, I.K.; Varesche, M.B.A.; Silva, E.L. HRT control as a strategy to enhance continuous hydrogen production from sugarcane juice under mesophilic and thermophilic conditions in AFBRs. Int. J. Hydrogen Energy 2019, 44, 19719–19729. [Google Scholar] [CrossRef]
- Menezes, C.A.; Silva, E.L. Hydrogen production from sugarcane juice in expanded granular sludge bed reactors under mesophilic conditions: The role of homoacetogenesis and lactic acid production. Ind. Crop. Prod. 2019, 138, 111586. [Google Scholar] [CrossRef]
- Freitas, I.B.F.; Menezes, C.A.; Silva, E.L. An alternative for value aggregation to the sugarcane chain: Biohydrogen and volatile fatty acids production from sugarcane molasses in mesophilic expanded granular sludge bed reactors. Fuel 2020, 260, 116419. [Google Scholar] [CrossRef]
- Bernal, A.P.; Menezes, C.A.; Silva, E.L. A new side-looking at the dark fermentation of sugarcane vinasse: Improving the carboxylates production in mesophilic EGSB by selection of the hydraulic retention time and substrate concentration. Int. J. Hydrogen Energy 2021, 46, 12758–12770. [Google Scholar] [CrossRef]
- Niz, M.Y.K.; Etchelet, I.; Fuentes, L.; Etchebehere, C.; Zaiat, M. Extreme thermophilic condition: An alternative for long-term biohydrogen production from sugarcane vinasse. Int. J. Hydrogen Energy 2019, 44, 22876–22887. [Google Scholar] [CrossRef]
- Piffer, M.A.; Oliveira, C.A.; Bovio-Winkler, P.; Eng, F.; Etchebehere, C.; Zaiat, M.; Nascimento, C.A.O.; Fuess, L.T. Sulfate- and pH-driven metabolic flexibility in sugarcane vinasse dark fermentation stimulates biohydrogen evolution, sulfidogenesis or homoacetogenesis. Int. J. Hydrogen Energy 2022, 47, 31202–31222. [Google Scholar] [CrossRef]
- Singh, A.; Müller, B.; Fuxelius, H.H.; Schnürer, A. AcetoBase: A functional gene repository and database for formyltetrahydrofolate synthetase sequences. Database 2019, 2019, baz142. [Google Scholar] [CrossRef]
- Klemps, R.; Cypionka, H.; Widdel, F.; Pfennig, N. Growth with hydrogen, and further physiological characteristics of Desulfotomaculum species. Arch. Microbiol. 1985, 143, 203–208. [Google Scholar] [CrossRef]
- Okamoto, M.; Miyahara, T.; Mizuno, O.; Noike, T. Biological hydrogen potential of materials characteristic of the organic fraction of municipal solid wastes. Water Sci. Technol. 2000, 41, 25–32. [Google Scholar] [CrossRef]
- Garay-Arroyo, A.; Covarrubias, A.A.; Clark, I.; Niño, I.; Gosset, G.; Martínez, A. Response to different environmental stress conditions of industrial an laboratory Saccharomyces cerevisiae strains. Appl. Microbiol. Biotechnol. 2004, 63, 734–741. [Google Scholar] [CrossRef]
- Ferraz, A.D.N., Jr.; Wenzel, J.; Etchebehere, C.; Zaiat, M. Effect of organic loading rate on hydrogen production from sugarcane vinasse in thermophilic acidogenic packed bed reactors. Int. J. Hydrogen Energy 2014, 39, 16852–16862. [Google Scholar]
- Fuess, L.T.; Zaiat, M.; Nascimento, C.A.O. Novel insights on the versatility of biohydrogen production from sugarcane vinasse via thermophilic dark fermentation: Impacts of pH-driven operating strategies on acidogenesis metabolite profiles. Bioresour. Technol. 2019, 286, 121379. [Google Scholar] [CrossRef]
- Rogeri, R.C.; Fuess, L.T.; Eng, F.; Borges, A.V.; Araujo, M.A.; Damianovic, M.H.R.Z.; Silva, A.J. Strategies to control pH in the dark fermentation of sugarcane vinasse: Impacts on sulfate reduction, biohydrogen production and metabolite distribution. J. Environ. Manag. 2023, 325, 116495. [Google Scholar] [CrossRef] [PubMed]
- Fuess, L.T.; Zaiat, M.; Nascimento, C.A.O. Molasses vs. juice: Maximizing biohydrogen production in sugarcane biorefineries to diversify renewable energy generation. J. Water Process Eng. 2020, 37, 101534. [Google Scholar] [CrossRef]
- Fuess, L.T.; Fuentes, L.; Bovio-Winkler, P.; Eng, F.; Etchebehere, C.; Zaiat, M.; Nascimento, C.A.O. Full details on continuous biohydrogen production from sugarcane molasses are unraveled: Performance optimization, self-regulation, metabolic correlations and quanti-qualitative biomass characterization. Chem. Eng. J. 2021, 414, 128934. [Google Scholar] [CrossRef]
- Jo, J.H.; Lee, D.S.; Park, D.; Park, J.M. Biological hydrogen production by immobilized cells of Clostridium tyrobutyricum JM1 isolated from a food waste treatment process. Bioresour. Technol. 2008, 99, 6666–6672. [Google Scholar] [CrossRef]
- Piffer, M.A.; Zaiat, M.; Nascimento, C.A.O.; Fuess, L.T. Dynamics of sulfate reduction in the thermophilic dark fermentation of sugarcane vinasse: A biohydrogen-independent approach targeting enhanced bioenergy production. J. Environ. Chem. Eng. 2021, 9, 105956. [Google Scholar] [CrossRef]
- Oliveira, C.A.; Fuess, L.T.; Soares, L.A.; Damianovic, M.H.R.Z. Thermophilic biohydrogen production from sugarcane molasses under low pH: Metabolic and microbial aspects. Int. J. Hydrogen Energy 2020, 45, 4182–4192. [Google Scholar] [CrossRef]
- Ramos, L.R.; Silva, E.L. Continuous hydrogen production from agricultural wastewaters at thermophilic and hyperthermophilic temperatures. Appl. Biochem. Biotechnol. 2017, 182, 846–869. [Google Scholar] [CrossRef]
- Fuess, L.T.; Garcia, M.L.; Zaiat, M. Seasonal characterization of sugarcane vinasse: Assessing environmental impacts from fertirrigation and the bioenergy recovery potential through biodigestion. Sci. Total Environ. 2018, 634, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Lepe, J.A.S.; Leal, B.I. Microbiología Enológica: Fundamentos de Vinificacion, 3rd ed.; Ediciones Mundi-Prensa: Madrid, Spain, 2004. (In Spanish) [Google Scholar]
- Kolb, S.; Otte, H.; Nagel, B.; Schink, B. Energy conservation in malolactic fermentation by Lactobacillus plantarum and Lactobacillus sake. Arch. Microbiol. 1992, 157, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Fuess, L.T.; Santos, G.M.; Delforno, T.P.; Moraes, B.S.; Silva, A.J. Biochemical butyrate production via dark fermentation as an energetically efficient alternative management approach for vinasse in sugarcane biorefineries. Renew. Energy 2020, 158, 3–12. [Google Scholar] [CrossRef]
- Ding, H.B.; Amy, T.G.; Wang, J.Y. Caproate formation in mixed-culture fermentative hydrogen production. Bioresour Technol. 2010, 101, 9550–9559. [Google Scholar] [CrossRef]
- Cavalcante, W.A.; Leitão, R.C.; Gehring, T.A.; Angenent, L.T.; Santaella, S.T. Anaerobic fermentation for n-caproic acid production: A review. Process Biochem. 2017, 54, 106–119. [Google Scholar] [CrossRef]
- Chen, W.M.; Tseng, Z.J.; Lee, K.S.; Chang, J.S. Fermentative hydrogen production with Clostridium butyricum CGS5 isolated from anaerobic sewage sludge. Int. J. Hydrogen Energy 2005, 30, 1063–1070. [Google Scholar] [CrossRef]
- Infantes, D.; González del Campo, A.; Villaseñor, J.; Fernández, F.J. Influence of pH, temperature and volatile fatty acids on hydrogen production by acidogenic fermentation. Int. J. Hydrogen Energy 2011, 36, 15595–15601. [Google Scholar] [CrossRef]
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 1977, 41, 100–180. [Google Scholar] [CrossRef]
- Fernandes, B.S.; Peixoto, G.; Albrecht, F.R.; Del Aguila, N.K.S.; Zaiat, M. Potential to produce biohydrogen from various wastewaters. Energy Sustain. Dev. 2010, 14, 143–148. [Google Scholar] [CrossRef]
- Wu, K.J.; Saratale, G.D.; Lo, Y.C.; Chen, W.M.; Tseng, Z.J.; Chang, M.C.; Tsai, B.C.; Sud, A.; Chang, J.S. Simultaneous production of 2,3-butanediol, ethanol and hydrogen with a Klebsiella sp. strain isolated from sewage sludge. Bioresour. Technol. 2008, 99, 7966–7970. [Google Scholar] [CrossRef]
- CEPEA. CEPEA/ESALQ Hydrous Ethanol Index (Fuel)—São Paulo State. Available online: https://cepea.esalq.usp.br/en/indicator/ethanol.aspx (accessed on 27 June 2023).
- EPE. Leilões. Available online: https://www.epe.gov.br/pt/leiloes-de-energia/leiloes (accessed on 28 June 2023). (In Portuguese)
- Harahap, B.M.; Ahring, B.K. Acetate Production from Syngas Produced from Lignocellulosic Biomass Materials along with Gaseous Fermentation of the Syngas: A Review. Microorganisms 2023, 11, 995. [Google Scholar] [CrossRef]
- Sauer, M.; Porro, D.; Mattanovich, D.; Branduardi, P. Microbial production of organic acids: Expanding the markets. Trends Biotechnol. 2008, 26, 100–108. [Google Scholar] [CrossRef]
- Angelidaki, I.; Sanders, W. Assessment of the anaerobic biodegradability of macropollutants. Rev. Environ. Sci. Biotechnol. 2004, 3, 117–129. [Google Scholar] [CrossRef]
- EIA. Open Data: Natural Gas. Available online: https://www.eia.gov/opendata/browser/natural-gas/pri (accessed on 28 June 2023).
- Honda. Honda FCEV Concept Makes World Debut at Los Angeles International Auto Show. November 2013. Available online: https://hondanews.com/en-US/releases/release-47803fbebd894dddb275faf0fb0c0a28-honda-fcev-concept-makes-world-debut-at-los-angeles-international-auto-show (accessed on 27 June 2023).
- Peixoto, G.; Pantoja-Filho, J.L.R.; Agnelli, J.A.B.; Barboza, M.; Zaiat, M. Hydrogen and methane production, energy recovery, and organic matter removal from effluents in a two-stage fermentative process. Appl. Biochem. Biotechnol. 2012, 168, 651–671. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.O.S.; Junqueira, T.L.; Cavalett, O.; Cunha, M.P.; Jesus, C.D.F.; Mantelatto, P.E.; Rosell, C.E.V.; Maciel-Filho, R.; Bonomi, A. Cogeneration in integrated first and second generation ethanol from sugarcane. Chem. Eng. Res. Des. 2013, 91, 1411–1417. [Google Scholar] [CrossRef]
- Dantas, G.A.; Legey, L.F.L.; Mazzone, A. Energy from sugarcane bagasse in Brazil: An assessment of the productivity and cost of different technological routes. Renew. Sustain. Energy Rev. 2013, 21, 356–364. [Google Scholar] [CrossRef]
- Clean Cities. Alternative Fuel Price Report; U.S. Department of Energy: Washington, DC, USA, 2016. Available online: http://www.afdc.energy.gov/uploads/publication/alternative_fuel_price_report_april_2016.pdf (accessed on 27 June 2023).
- MOLBASE. Available online: https://www.molbase.com/ (accessed on 28 June 2023).
- Chiesa, P.; Macchi, E. A Thermodynamic Analysis of Different Options to Break 60% Electric Efficiency in Combined Cycle Power Plants. J. Eng. Gas Turbines Power 2004, 126, 770–785. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Xylose-Based Wastewater | Pentose Liquor |
|---|---|---|
| Total carbohydrates (mg·L−1) | 1495 ± 733 | 1459 ± 440 |
| Xylose (%) | 99.9 | 82.4 |
| Arabinose (%) | – | 7.12 |
| Rhamnose (%) | – | 6.14 |
| Glucose (%) | – | 1.58 |
| pH | 6.7 ± 0.1 | 5.9 ± 0.1 |
| Mass Flow Rate (mg·h−1) | Condition I (Xylose-Based Wastewater) | Condition II (Pentose Liquor) | ||
|---|---|---|---|---|
| Maximum | Mean | Maximum | Mean | |
| Influent xylose | 4171.12 (10) | 2096.68 ± 811.4 | 2256.85 (33) | 1886.92 ± 141.3 |
| Effluent xylose | 2591.34 (4) | 1148.13 ± 491.2 | 1312.13 (2) | 610.28 ± 173.7 |
| Citric acid | 72.28 (36) | 70.71 ± 1.07 | ND | ND |
| Malic acid | 87.19 (17) | 45.89 ± 16.8 | 69.31 (2) | 56.82 ± 6.6 |
| Succinic acid | 47.22 (25) | 46.88 ± 0.3 | ND | ND |
| Lactic acid | 119.36 (3) | 107.37 ± 7.0 | 248.73 (4) | 229.87 ± 9.1 |
| Formic acid | 28.11 (22) | 24.86 ± 2.0 | ND | ND |
| Acetic acid | 305.76 (36) | 150.66 ± 61.2 | 379.20 1 (10) | 186.72 1 ± 89.8 |
| Propionic acid | 125.92 (31) | 79.63 ± 17.0 | 447.80 (33) | 298.52 ± 78.6 |
| Isobutyric acid | 94.07 (3) | 72.33 ± 8.7 | 156.48 (26) | 119.34 ± 28.0 |
| n-Butyric acid | 120.80 (8) | 85.53 ± 15.2 | 473.39 (18) | 299.23 ± 71.1 |
| Isovaleric acid | 140.55 (10) | 109.02 ± 22.4 | 88.20 (18) | 53.24 ± 18.8 |
| n-Valeric acid | 72.65 (36) | 69.61 ± 1.6 | 85.62 (31) | 64.43 ± 13.3 |
| Caproic acid | ND | ND | 32.40 (18) | 17.47 ± 4.0 |
| Ethanol | 72.05 (29) | 51.46 ± 11.0 | 215.66 (26) | 144.47 ± 22.5 |
| Hydrogen | 11.24 (8) | 1.84 ± 0.003 | 2.70 (16) | 0.55 ± 0.0008 |
| Carbon dioxide | 82.43 (8) | 15.70 ± 0.02 | 68.04 (16) | 13.22 ± 0.02 |
| Sum (soluble + gas metabolites) | NC | 2079.62 | NC | 2094.16 |
| Correspondence (%) 2 | NC | 99.2 | NC | 111.0 |
| Reactor | Xylose Concentration (gCOD·L−1) | Temp. | pH | HY | VHPR | VOA Production (mg·h−1) | Reference | ||
|---|---|---|---|---|---|---|---|---|---|
| HBu | HPr | HAc | |||||||
| CSTR | 20.0 | 50 | 6.5 | 0.4 | 6600 | 264 | 1767 | 1601 | [36] |
| AGSB | 20.0 | 40 | 6.5 | 0.6 | 19,680 | 835 | 173 | 820 | [36] |
| Batch 2 | 1.5 | 37 | 5.5 | 1.5 | - | 853 1 | 4.53 1 | 149 1 | [26] |
| UAPBR | 1.7 | 25 | 6.7 | 0.08 | 398.4 | 120.8 | 125.9 | 305.76 | This study |
| UAPBR 2 | 1.7 | 25 | 5.9 | 0.04 | 120 | 473.4 | 447.8 | 379.20 | This study |
| Scenarios/Products | 1 | 2 | 3 | 4 | |
|---|---|---|---|---|---|
| 1G Ethanol 50%—2G Ethanol 50%—Cogeneration | 1G Ethanol 2G Ethanol | 1G Ethanol 2G Ethanol Hydrogen Organic Acids | 1G Ethanol 2G Ethanol Hydrogen Methane | ||
| 1G Ethanol | Production (m3·d−1) 1 | 987.60 | 987.60 | 987.60 | 987.60 |
| Econ. Pot. (USD·d−1) 2 | 521,924.96 | 521,924.96 | 521,924.96 | 521,924.96 | |
| 2G Ethanol | Production (m3·d−1) 3 | 149.69 | 299.37 | 299.37 | 299.37 |
| Econ. Pot. (USD·d−1) 2 | 79,106.83 | 158,213.66 | 158,213.66 | 158,213.66 | |
| Hydrogen | Production (m3·d−1) 4 | NA | NA | 19.06 | 19.06 |
| Econ. Pot. (USD·d−1) 5 | NA | NA | 2897.44 | 2897.44 | |
| Organic Acids (HLa, HAc, HPr, HBu) | Production (kg·d−1) 6 | NA | NA | 201,144.05 | NA |
| Econ. Pot. (USD·d−1) 7 | NA | NA | 162,693.67 | NA | |
| Energy (cogeneration 8 /methane 11,12) | Production (m3·d−1) Energ. Pot. (MWh) | NA | NA | NA | 118,931.60 11 |
| 334.32 8 | NA | NA | 806.73 12 | ||
| Econ. Pot. (USD·d−1) | 23,422.82 9 | NA | NA | 52,192.02 9 | |
| Total Energetic potential (MWh) | 334.32 | NA | NA | 806.73 | |
| Total Economic potential (USD·d−1) | 622,660.68 | 680,138.61 | 845,729.72 | 735,228.08 | |
| Total Environmental potential (COD removal percentage) | NA | NA | 62.7 10 | 74.7 11 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peixoto, G.; Mockaitis, G.; Moreira, W.K.; Lima, D.M.F.; de Lima, M.A.; Ferreira, F.V.; Fuess, L.T.; Polikarpov, I.; Zaiat, M. Acidogenesis of Pentose Liquor to Produce Biohydrogen and Organic Acids Integrated with 1G–2G Ethanol Production in Sugarcane Biorefineries. Waste 2023, 1, 672-688. https://doi.org/10.3390/waste1030040
Peixoto G, Mockaitis G, Moreira WK, Lima DMF, de Lima MA, Ferreira FV, Fuess LT, Polikarpov I, Zaiat M. Acidogenesis of Pentose Liquor to Produce Biohydrogen and Organic Acids Integrated with 1G–2G Ethanol Production in Sugarcane Biorefineries. Waste. 2023; 1(3):672-688. https://doi.org/10.3390/waste1030040
Chicago/Turabian StylePeixoto, Guilherme, Gustavo Mockaitis, Wojtyla Kmiecik Moreira, Daniel Moureira Fontes Lima, Marisa Aparecida de Lima, Filipe Vasconcelos Ferreira, Lucas Tadeu Fuess, Igor Polikarpov, and Marcelo Zaiat. 2023. "Acidogenesis of Pentose Liquor to Produce Biohydrogen and Organic Acids Integrated with 1G–2G Ethanol Production in Sugarcane Biorefineries" Waste 1, no. 3: 672-688. https://doi.org/10.3390/waste1030040