
Human Nasal Epithelium Organoids for Assessing Neutralizing Antibodies to a Protective SARS-CoV-2 Virus-like Particle Vaccine
 , ,
, ,  , , , , , , , , , , and
, , , , , , , , , , and 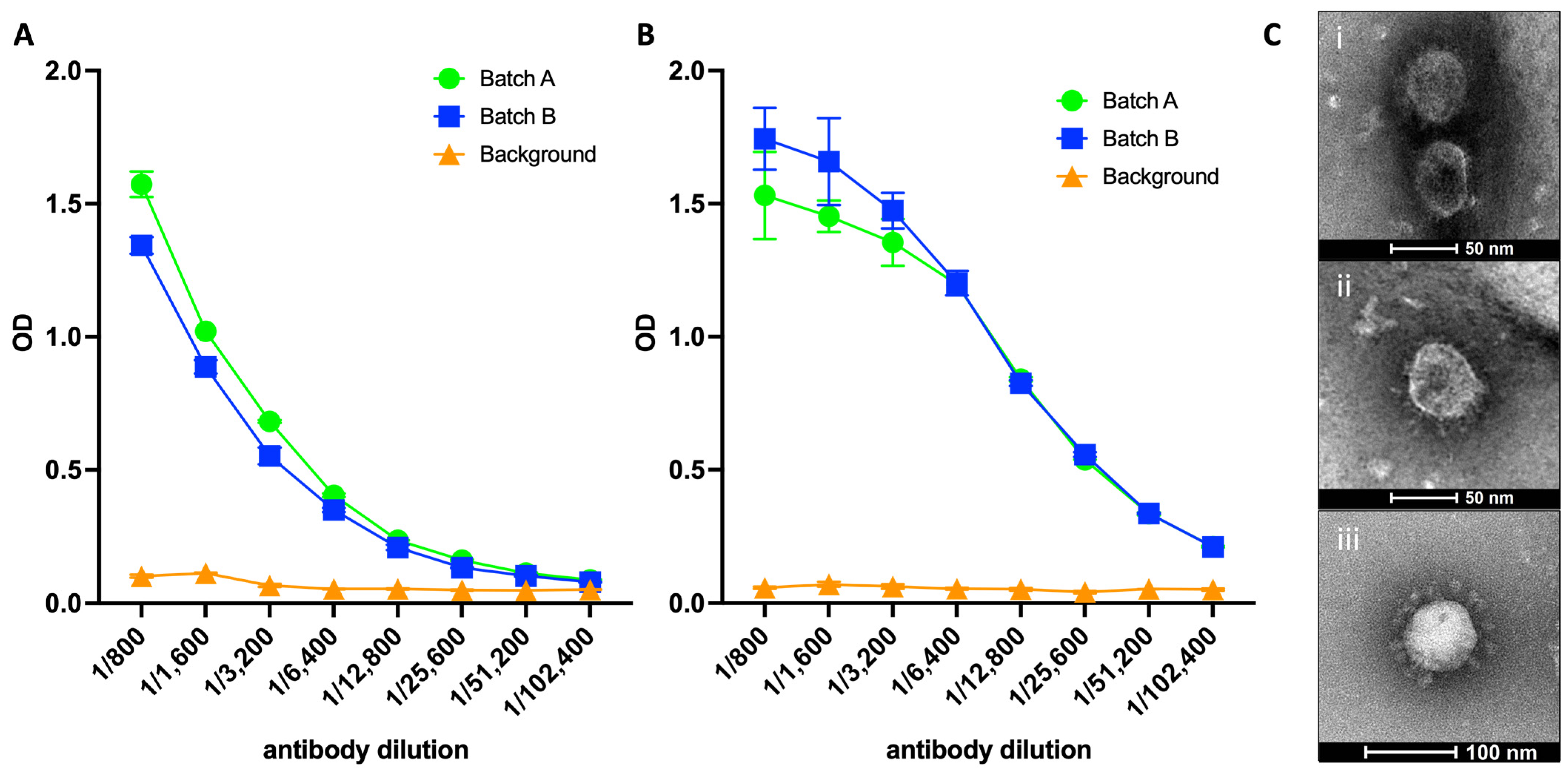{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. VLP Production and Purification
2.3. Direct-VLP Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Negative Staining by Transmission Electron Microscopy (TEM)
2.5. Mice and Vaccinations
2.6. Measurement of Viral Loads via 50% Tissue Culture Infectious Dose (TCID50)
2.7. SARS-CoV-2 Surrogate Virus Neutralization Test (sVNT) Assays
2.8. ALI-HNE Organoids
2.9. SARS-CoV-2 Propagation and ALI-HNE Infection
2.10. Immunofluorescence and Confocal Microscopy
2.11. ELISpot
2.12. Isolation of Primary Dendritic Cells
3. Results
3.1. SARS-CoV-2 VLPs Exhibit Reactivity for Spike Protein and RBD
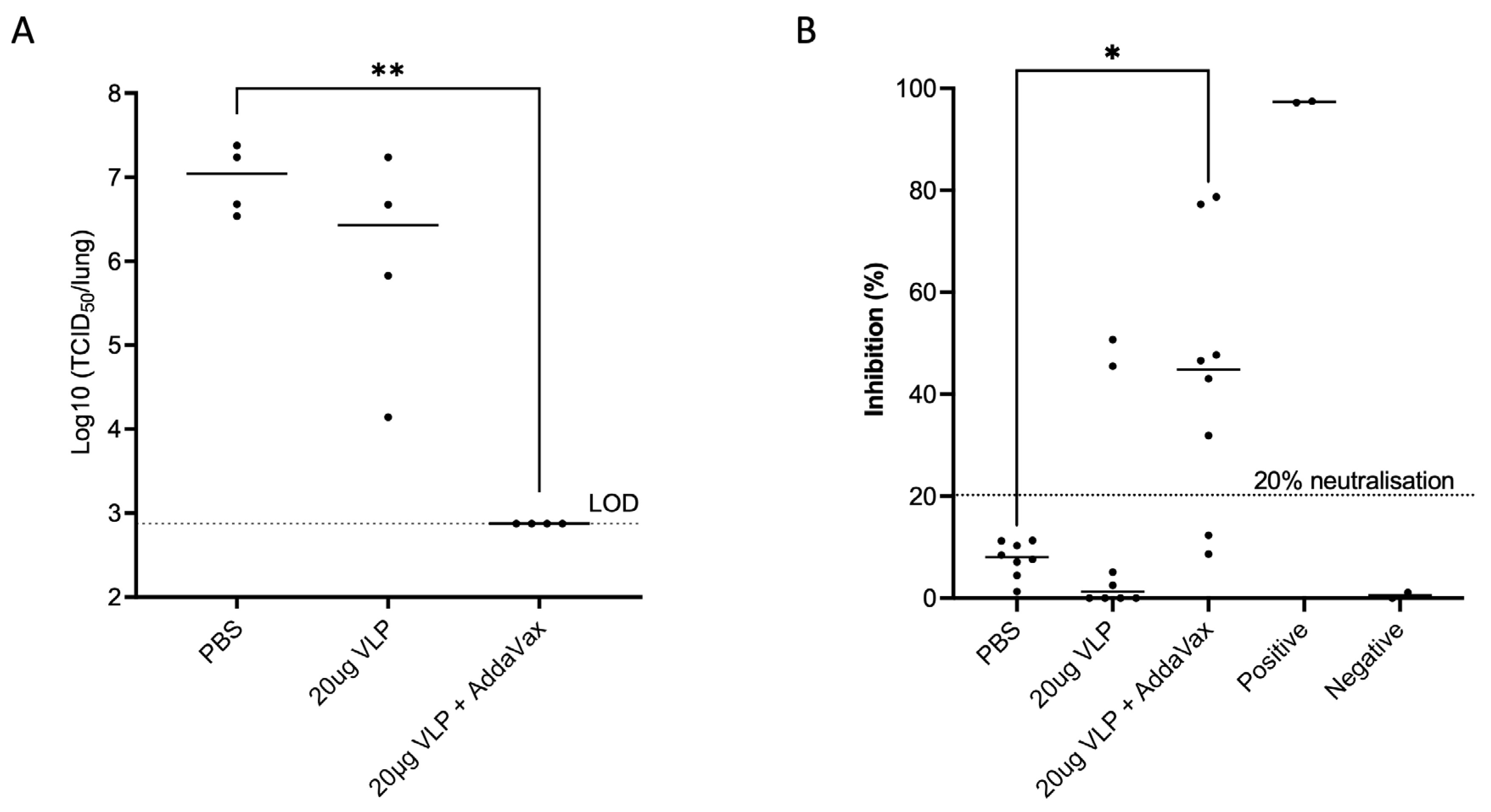
3.2. SARS-CoV-2 VLPs Induce Nab Responses and Completely Protect against Viral Challenge
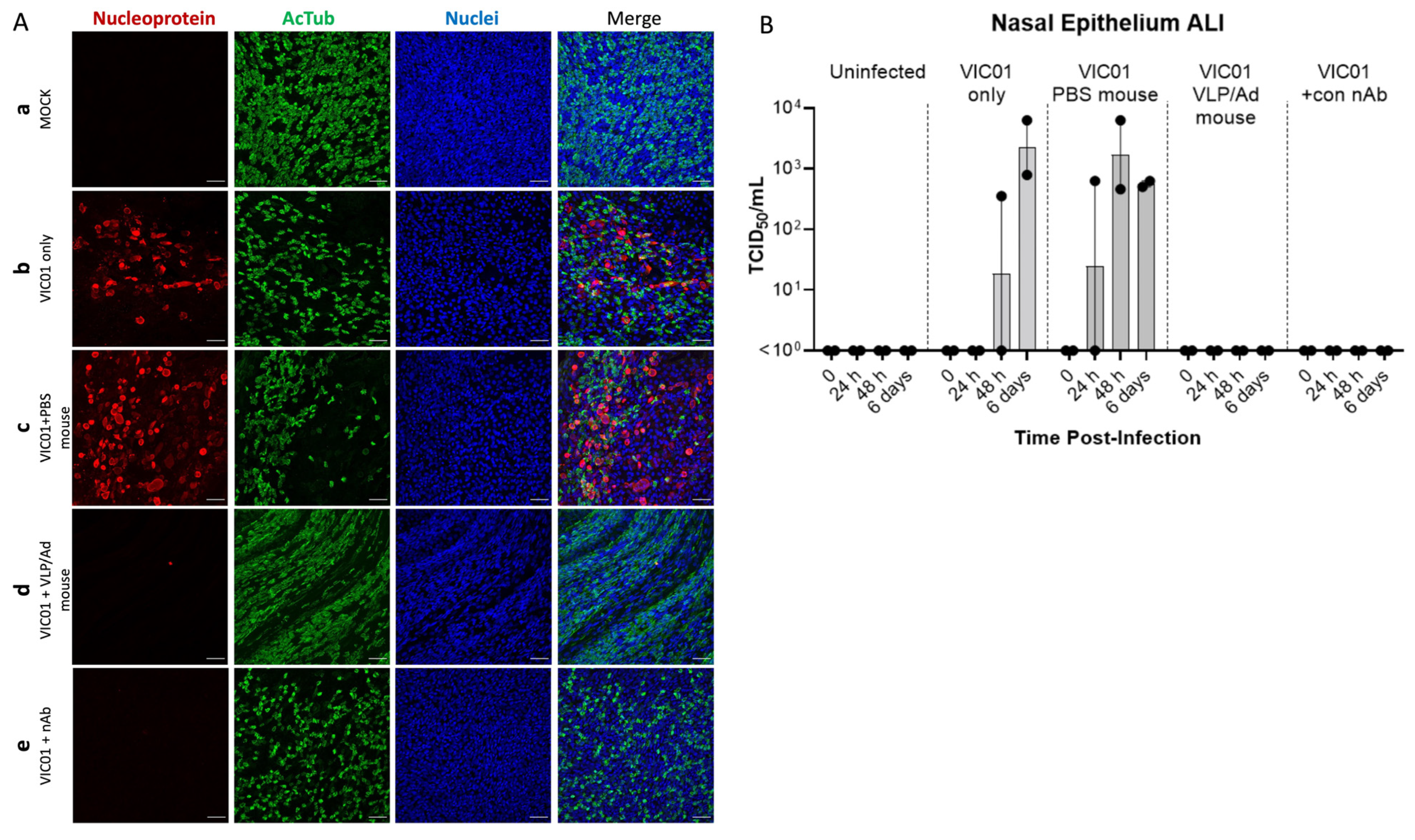
3.3. Mouse Sera from VLP Vaccinated Mice Completely Neutralizes SARS-CoV-2 Virus in an ALI-HNE Model
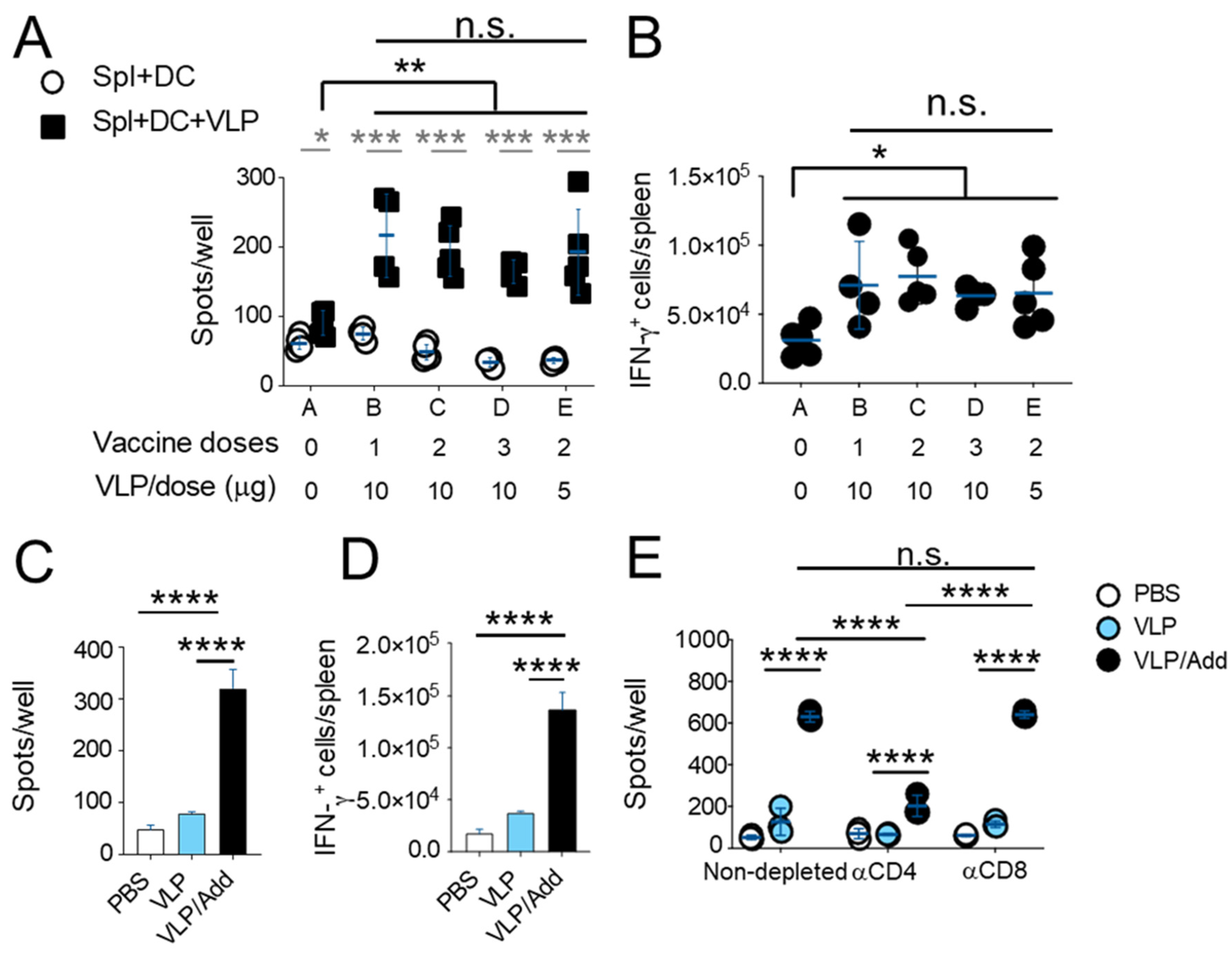
3.4. T Cells Play a Role in Response against SARS-CoV-2 after Vaccination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Kurhade, C.; Zou, J.; Xia, H.; Liu, M.; Chang, H.C.; Ren, P.; Xie, X.; Shi, P.Y. Low neutralization of SARS-CoV-2 Omicron BA.2.75.2, BQ.1.1 and XBB.1 by parental mRNA vaccine or a BA.5 bivalent booster. Nat. Med. 2023, 29, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Qu, P.; Evans, J.P.; Faraone, J.N.; Zheng, Y.M.; Carlin, C.; Anghelina, M.; Stevens, P.; Fernandez, S.; Jones, D.; Lozanski, G.; et al. Enhanced neutralization resistance of SARS-CoV-2 Omicron subvariants BQ.1, BQ.1.1, BA.4.6, BF.7, and BA.2.75.2. Cell Host Microbe 2023, 31, 9–17.e13. [Google Scholar] [CrossRef] [PubMed]
- Takashita, E.; Kinoshita, N.; Yamayoshi, S.; Sakai-Tagawa, Y.; Fujisaki, S.; Ito, M.; Iwatsuki-Horimoto, K.; Halfmann, P.; Watanabe, S.; Maeda, K.; et al. Efficacy of Antiviral Agents against the SARS-CoV-2 Omicron Subvariant BA.2. N. Engl. J. Med. 2022, 386, 1475–1477. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Kaki, M.; Potluri, V.S.; Kahar, P.; Khanna, D. A comprehensive review of SARS-CoV-2 vaccines: Pfizer, Moderna & Johnson & Johnson. Hum. Vaccines Immunother. 2022, 18, 2002083. [Google Scholar] [CrossRef]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Stralin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168.e114. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e1415. [Google Scholar] [CrossRef]
- Nguyen, T.H.O.; Rowntree, L.C.; Petersen, J.; Chua, B.Y.; Hensen, L.; Kedzierski, L.; van de Sandt, C.E.; Chaurasia, P.; Tan, H.X.; Habel, J.R.; et al. CD8(+) T cells specific for an immunodominant SARS-CoV-2 nucleocapsid epitope display high naive precursor frequency and TCR promiscuity. Immunity 2021, 54, 1066–1082.e1065. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Quadeer, A.A.; McKay, M.R. Preliminary Identification of Potential Vaccine Targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies. Viruses 2020, 12, 254. [Google Scholar] [CrossRef]
- Keating, G.M.; Noble, S. Recombinant hepatitis B vaccine (Engerix-B): A review of its immunogenicity and protective efficacy against hepatitis B. Drugs 2003, 63, 1021–1051. [Google Scholar] [CrossRef]
- Venters, C.; Graham, W.; Cassidy, W. Recombivax-HB: Perspectives past, present and future. Expert Rev. Vaccines 2004, 3, 119–129. [Google Scholar] [CrossRef]
- Roy, V.; Jung, W.; Linde, C.; Coates, E.; Ledgerwood, J.; Costner, P.; Yamshchikov, G.; Streeck, H.; Juelg, B.; Lauffenburger, D.A.; et al. Differences in HPV-specific antibody Fc-effector functions following Gardasil(R) and Cervarix(R) vaccination. NPJ Vaccines 2023, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Zaman, K.; Dudman, S.; Stene-Johansen, K.; Qadri, F.; Yunus, M.; Sandbu, S.; Gurley, E.S.; Overbo, J.; Julin, C.H.; Dembinski, J.L.; et al. HEV study protocol: Design of a cluster-randomised, blinded trial to assess the safety, immunogenicity and effectiveness of the hepatitis E vaccine HEV 239 (Hecolin) in women of childbearing age in rural Bangladesh. BMJ Open 2020, 10, e033702. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wei, M.; Sun, G.; Wang, X.; Li, M.; Lin, Z.; Li, Z.; Li, Y.; Fang, M.; Zhang, J.; et al. Real-time stability of a hepatitis E vaccine (Hecolin®) demonstrated with potency assays and multifaceted physicochemical methods. Vaccine 2016, 34, 5871–5877. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, A.Y.; Shehzad, A.; Islam, S.U.; Al-Suhaimi, E.A.; Lee, Y.S. Mosquirix RTS, S/AS01 Vaccine Development, Immunogenicity, and Efficacy. Vaccines 2022, 10, 713. [Google Scholar] [CrossRef] [PubMed]
- Collett, S.; Earnest, L.; Carrera Montoya, J.; Edeling, M.A.; Yap, A.; Wong, C.Y.; Christiansen, D.; Roberts, J.; Mumford, J.; Lecouturier, V.; et al. Development of virus-like particles with inbuilt immunostimulatory properties as vaccine candidates. Front. Microbiol. 2023, 14, 1065609. [Google Scholar] [CrossRef] [PubMed]
- Tran, B.M.; Grimley, S.L.; McAuley, J.L.; Hachani, A.; Earnest, L.; Wong, S.L.; Caly, L.; Druce, J.; Purcell, D.F.J.; Jackson, D.C.; et al. Air-Liquid-Interface Differentiated Human Nose Epithelium: A Robust Primary Tissue Culture Model of SARS-CoV-2 Infection. Int. J. Mol. Sci. 2022, 23, 835. [Google Scholar] [CrossRef] [PubMed]
- Hierholzer, J.C.; Killington, R.A. Virus isolation and quantitation. In Virology Methods Manual; Academic Press: Cambridge, MA, USA, 1996; pp. 25–46. [Google Scholar]
- Awatade, N.T.; Wong, S.L.; Capraro, A.; Pandzic, E.; Slapetova, I.; Zhong, L.; Turgutoglu, N.; Fawcett, L.K.; Whan, R.M.; Jaffe, A.; et al. Significant functional differences in differentiated Conditionally Reprogrammed (CRC)- and Feeder-free Dual SMAD inhibited-expanded human nasal epithelial cells. J. Cyst. Fibros 2021, 20, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Deliyannis, G.; Gherardin, N.A.; Wong, C.Y.; Grimley, S.L.; Cooney, J.P.; Redmond, S.J.; Ellenberg, P.; Davidson, K.C.; Mordant, F.L.; Smith, T.; et al. Broad immunity to SARS-CoV-2 variants of concern mediated by a SARS-CoV-2 receptor-binding domain protein vaccine. EBioMedicine 2023, 92, 104574. [Google Scholar] [CrossRef]
- Mach, N.; Gillessen, S.; Wilson, S.B.; Sheehan, C.; Mihm, M.; Dranoff, G. Differences in dendritic cells stimulated in vivo by tumors engineered to secrete granulocyte-macrophage colony-stimulating factor or Flt3-ligand. Cancer Res. 2000, 60, 3239–3246. [Google Scholar] [PubMed]
- Vremec, D.; Pooley, J.; Hochrein, H.; Wu, L.; Shortman, K. CD4 and CD8 expression by dendritic cell subtypes in mouse thymus and spleen. J. Immunol. 2000, 164, 2978–2986. [Google Scholar] [CrossRef] [PubMed]
- Collett, S.; Torresi, J.; Silveira, L.E.; Truong, V.K.; Christiansen, D.; Tran, B.M.; Vincan, E.; Ramsland, P.A.; Elbourne, A. Investigating virus-host cell interactions: Comparative binding forces between hepatitis C virus-like particles and host cell receptors in 2D and 3D cell culture models. J. Colloid Interface Sci. 2021, 592, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Bahnan, W.; Wrighton, S.; Sundwall, M.; Blackberg, A.; Larsson, O.; Hoglund, U.; Khakzad, H.; Godzwon, M.; Walle, M.; Elder, E.; et al. Spike-Dependent Opsonization Indicates Both Dose-Dependent Inhibition of Phagocytosis and That Non-Neutralizing Antibodies Can Confer Protection to SARS-CoV-2. Front. Immunol. 2021, 12, 808932. [Google Scholar] [CrossRef]
- Zhang, A.; Stacey, H.D.; D’Agostino, M.R.; Tugg, Y.; Marzok, A.; Miller, M.S. Beyond neutralization: Fc-dependent antibody effector functions in SARS-CoV-2 infection. Nat. Rev. Immunol. 2023, 23, 381–396. [Google Scholar] [CrossRef]
- Low, J.S.; Vaqueirinho, D.; Mele, F.; Foglierini, M.; Jerak, J.; Perotti, M.; Jarrossay, D.; Jovic, S.; Perez, L.; Cacciatore, R.; et al. Clonal analysis of immunodominance and cross-reactivity of the CD4 T cell response to SARS-CoV-2. Science 2021, 372, 1336–1341. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrera Montoya, J.; Collett, S.; Fernandez Ruiz, D.; Earnest, L.; Edeling, M.A.; Yap, A.H.Y.; Wong, C.Y.; Cooney, J.P.; Davidson, K.C.; Roberts, J.; et al. Human Nasal Epithelium Organoids for Assessing Neutralizing Antibodies to a Protective SARS-CoV-2 Virus-like Particle Vaccine. Organoids 2024, 3, 18-31. https://doi.org/10.3390/organoids3010002
Carrera Montoya J, Collett S, Fernandez Ruiz D, Earnest L, Edeling MA, Yap AHY, Wong CY, Cooney JP, Davidson KC, Roberts J, et al. Human Nasal Epithelium Organoids for Assessing Neutralizing Antibodies to a Protective SARS-CoV-2 Virus-like Particle Vaccine. Organoids. 2024; 3(1):18-31. https://doi.org/10.3390/organoids3010002
Chicago/Turabian StyleCarrera Montoya, Julio, Simon Collett, Daniel Fernandez Ruiz, Linda Earnest, Melissa A. Edeling, Ashley Huey Yiing Yap, Chinn Yi Wong, James P. Cooney, Kathryn C. Davidson, Jason Roberts, and et al. 2024. "Human Nasal Epithelium Organoids for Assessing Neutralizing Antibodies to a Protective SARS-CoV-2 Virus-like Particle Vaccine" Organoids 3, no. 1: 18-31. https://doi.org/10.3390/organoids3010002