
Nutritional Balance Matters: Assessing the Ramifications of Vitamin A Deficiency on Poultry Health and Productivity

Abstract
:1. Introduction
- Describe the critical contributions of vitamin A to the metabolism, growth, immunity, antioxidant capacity, and reproduction of avian species.
- Identify the key factors that contribute to vitamin A deficiency in domestic fowl.
- Identify the main clinical manifestations of vitamin A deficiency in poultry.
- Discuss effective intervention strategies for preventing and managing vitamin A deficiency in poultry populations.
- Address considerations for meeting the vitamin A requirements of poultry.
2. The Crucial Contributions of Vitamin A: Metabolism, Growth, Immunity, Antioxidant Capacity, and Reproduction
- (1)
- Digestion and Metabolism:
- (2)
- Growth and Development:
- (3)
- Immune Function:
- (4)
- Antioxidant Capacity:
- (5)
- Reproduction:
- (6)
- Vision:
3. Causes of Vitamin A Deficiency: Identifying Key Factors
- (1)
- Inadequate feed formulation: One prominent factor contributing to vitamin A deficiency in poultry diets is inadequate feed formulation [80]. When the formulation of poultry feed does not include sufficient quality sources of preformed vitamin A, it fails to provide the necessary levels of this essential micronutrient. To address this issue, careful consideration should be given to the feed formulation process, including the use of least-cost feed formulation methods [81,82]. It is crucial to ensure the inclusion of stable and highly bioavailable commercial sources of vitamin A to prevent deficiencies.
- (2)
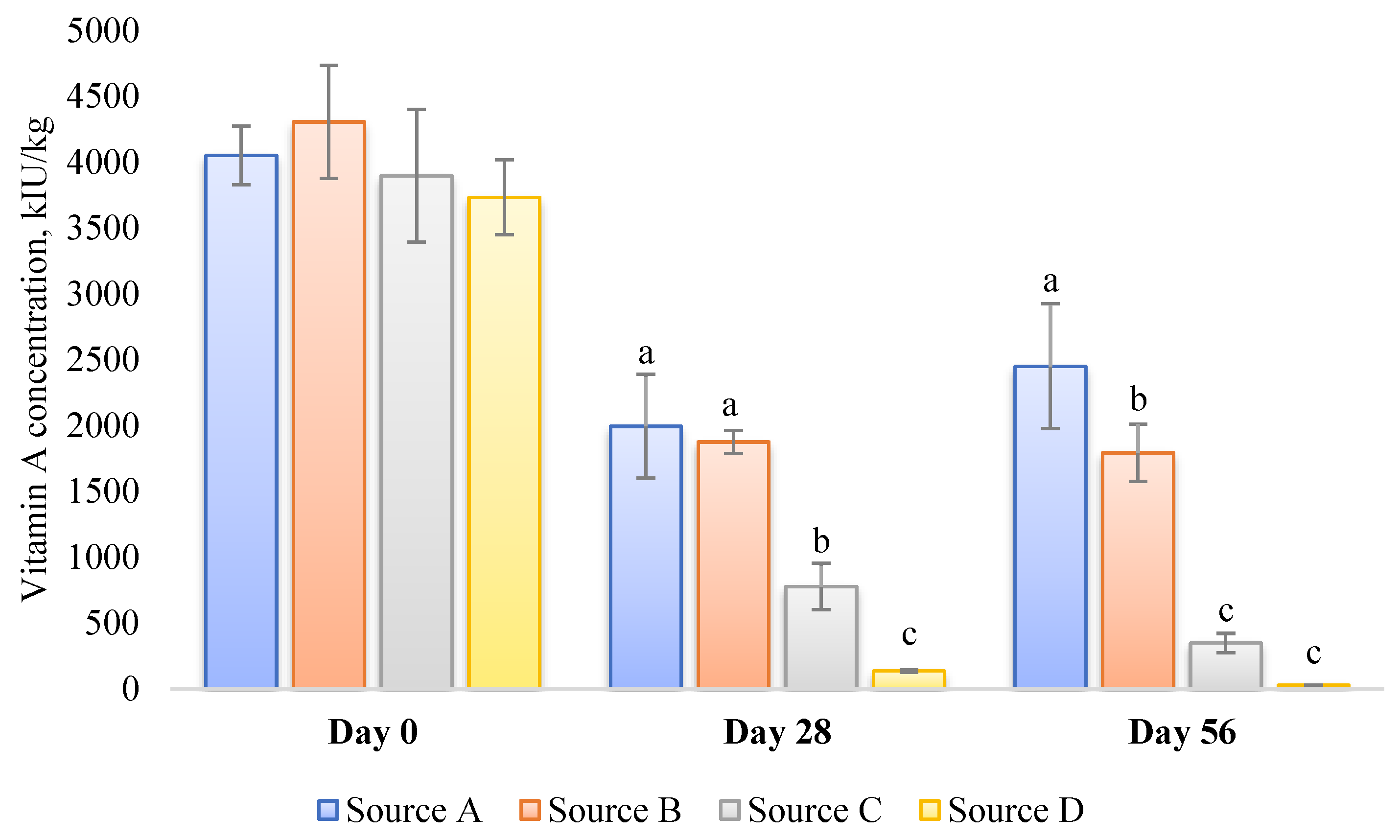
- Mishandling and improper storage of poultry premixes and feed can contribute to the deficiency of essential vitamins, including vitamin A. Over time, exposure to light, heat, and oxygen can lead to the degradation of vitamins in the premixes and feed [83]. For instance, some of the vitamin A sources available on the market, particularly those that are less stable, may lose their vitamin A activity completely if stored at elevated temperatures within a short period (Figure 1) [84]. Therefore, it is crucial to store premixes and feed in cool, dry, and dark conditions for the shortest possible time to maintain optimal levels of vitamin A. Regular quality checks and timely replenishment of premixes and feed stocks are also vital to preserve the potency of vitamin A and prevent its degradation. Furthermore, paying meticulous attention to the thermal treatment of feed is of the utmost importance [85]. Considering the specific example of pelleting, it is crucial to avoid subjecting the feed to excessively high temperatures for prolonged periods of time, as this can result in significant losses in vitamin A activity [86]. Under such conditions, implementing an additional safety margin for vitamin activity becomes sensible.
- (3)
- Factors affecting absorption and utilization: Factors that affect the absorption and utilization of fat-soluble vitamins should not be overlooked [3]. Gut health disorders, parasitic infections, imbalances, or deficiencies of other nutrients (dietary fat, other fat-soluble vitamins etc.), stress, mycotoxins in feed, and certain diseases can impair the absorption of retinol in the digestive tract [87]. To mitigate these factors, implementing measures to promote good gut health, such as using intestinal health promoters or ensuring proper sanitation practices, is important [88]. These strategies optimize vitamin A absorption and utilization in poultry.
- (4)
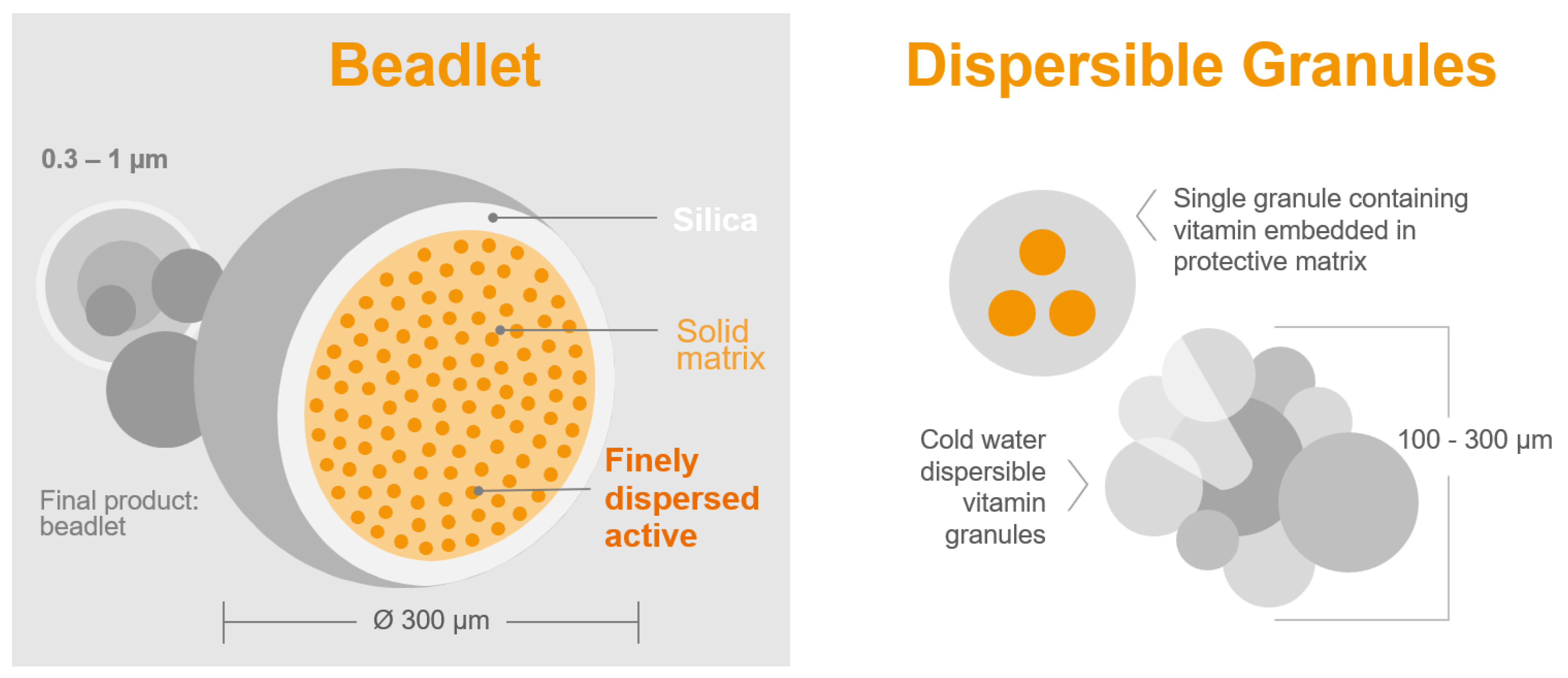
- Bioavailability: The bioavailability of vitamin A sources is often overlooked, but it plays a crucial role [89]. Unlike other vitamins, vitamin A in the form of retinyl acetate is commonly formulated in small solid beadlet particles by different suppliers (Figure 2). This can result in variations in stability and bioavailability among the vitamin A products available on the market [84]. Experienced formulators can easily create highly stable vitamin A products that can withstand challenging conditions with the help of special formulation aids. However, the main challenge lies in the digestive tract of poultry, where these stable formulations must release retinyl acetate in the intestinal lumen. If the formulations are too stable, they may have reduced or no biological value at all [90]. Therefore, it is essential to strike a delicate balance between the overall stability and bioavailability of a vitamin A commercial product. This balance ensures the product’s ability to withstand harsh storage conditions in premixes and high pelleting temperatures in feed, while also facilitating easy release in the digestive tract.
4. Identifying Clinical Manifestations of Vitamin A Deficiency
- (1)
- Reduced growth, high mortality, and poor feed conversion:
- (2)
- Decreased immune response:
- (3)
- Ocular abnormalities:
- (4)
- Reproductive disorders:
- (5)
- Neurological disorders
5. Addressing Effective Intervention Strategies
- (1)
- Dietary Modifications:
- (2)
- Supplementation:
- (3)
- Injection:
- (4)
- Oral application:
6. Balancing Vitamin A Intake and Hypervitaminosis Risks
7. Fulfilling Vitamin A Requirements
- (1)
- The NASEM and GfE are reputable scientific organizations that offer evidence-based guidance and requirement estimates for various nutrients, including vitamins. They conduct thorough reviews of scientific literature and consult experts to develop their specifications. Breeding companies, on the other hand, specialize in specific aspects of poultry production, focusing on developing and breeding poultry strains that excel in certain traits such as growth rate, feed conversion, and disease resistance. Their recommendations are often based on their own research, experience, and published scientific data.
- (2)
- The primary goal of NASEM and GfE is to provide scientifically rigorous and unbiased recommendations to meet the basic nutritional requirements of poultry. They may consider factors such as physiological needs, growth rates, reproductive performance, and environmental conditions. Breeding companies aim to develop and provide specific genetic lines or breeds of poultry that perform well under practical conditions. Their recommendations for vitamins and other nutrients are tailored to support the genetic potential of their poultry strains and achieve maximal performance in terms of growth, feed conversion, and overall production efficiency.
- (3)
- NASEM and GfE base their requirement estimates and allowances on extensive scientific research and systematic reviews of the available literature. They evaluate a wide range of sources, including published studies, experimental data, and meta-analyses, to establish nutrient requirement estimates for different poultry species and production stages. Breeding companies conduct their own research, often using proprietary data and genetic selection strategies. Their recommendations are specific to the genetic lines they develop and are typically derived from their internal studies, field trials, and the performance data of their poultry strains.
- (4)
- The requirement estimates and allowances of NASEM and GfE are widely accessible to the public and scientific community. They publish reports and guidelines that provide detailed information on the nutrient requirements for poultry and other animal species, including vitamins and other essential nutrients. Recommendations from breeding companies may be proprietary and shared only with their customers or collaborators.
8. Conclusions
- (1)
- Adequate nutrition, including sufficient vitamin A, is essential for maintaining optimal health, growth, and productivity in poultry flocks.
- (2)
- Vitamin A plays a crucial role in supporting various physiological functions in domestic fowl, including growth and development, immune function, antioxidant capacity, reproduction, and vision.
- (3)
- Vitamin A deficiency in poultry diets can result from inadequate feed formulation and the mishandling and improper storage of premixes and feed, factors affecting absorption and utilization.
- (4)
- Clinical manifestations of vitamin A deficiency in domestic fowl include reduced growth, high mortality, poor feed conversion, decreased immune response, ocular abnormalities, reproductive disorders, and neurological pathologies.
- (5)
- Effective intervention strategies for preventing and treating vitamin A deficiency in poultry encompass a range of approaches, including dietary modifications, supplementation, injection, and oral application. It is essential to evaluate the efficacy, practicality, and potential challenges associated with these strategies based on the specific circumstances.
- (6)
- Meeting the vitamin A requirements of poultry is crucial for their well-being and productivity. Differentiating between the requirement estimates and allowances provided by scientific committees and recommendations from breeding companies is important when addressing vitamin A nutrition.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; Tiwari, R.; Yatoo, M.I.; Karthik, K.; Michalak, I.; Dhama, K. Nutritional significance of amino acids, vitamins and minerals as nutraceuticals in poultry production and health—A comprehensive review. Vet. Q. 2020, 41, 1–29. [Google Scholar] [CrossRef]
- Khan, R.U.; Khan, A.; Naz, S.; Ullah, Q.; Puvača, N.; Laudadio, V.; Mazzei, D.; Seidavi, A.; Ayasan, T.; Tufarelli, V. Pros and Cons of Dietary Vitamin A and Its Precursors in Poultry Health and Production: A Comprehensive Review. Antioxidants 2023, 12, 1131. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. Delving into Vitamin A Supplementation in Poultry Nutrition: Current Knowledge, Functional Effects, and Practical Implications. World's Poult. Sci. J. 2023, 79, 1–23. [Google Scholar] [CrossRef]
- Shojadoost, B.; Yitbarek, A.; Alizadeh, M.; Kulkarni, R.R.; Astill, J.; Boodhoo, N.; Sharif, S. Centennial Review: Effects of vitamins A, D, E, and C on the chicken immune system. Poult. Sci. 2021, 100, 100930. [Google Scholar] [CrossRef] [PubMed]
- McDowell, L.R. Vitamins in Animal and Human Nutrition; McDowell, L.R., Ed.; Iowa State University Press: Ames, IA, USA, 2000; pp. 15–90. [Google Scholar]
- Çalişlar, S. The Important of Beta Carotene on Poultry Nutrition. Selcuk. J. Agric. Food Sci. 2019, 33, 252–259. [Google Scholar] [CrossRef]
- Moreno, J.A.; Díaz-Gómez, J.; Nogareda, C.; Angulo, E.; Sandmann, G.; Portero-Otin, M.; Serrano, J.C.E.; Twyman, R.M.; Capell, T.; Zhu, C.; et al. The distribution of carotenoids in hens fed on biofortified maize is influenced by feed composition, absorption, resource allocation and storage. Sci. Rep. 2016, 6, 35346. [Google Scholar] [CrossRef] [PubMed]
- Green, A.S.; Fascetti, A.J. Meeting the Vitamin A Requirement: The Efficacy and Importance of β-Carotene in Animal Species. Sci. World J. 2016, 2016, 7393620. [Google Scholar] [CrossRef] [PubMed]
- Surai, P. The antioxidant properties of canthaxanthin and its potential effects in the poultry eggs and on embryonic development of the chick. Part 1. World's Poult. Sci. J. 2012, 68, 465–476. [Google Scholar] [CrossRef]
- Guo, S.; He, L.; Zhang, Y.; Niu, J.; Li, C.; Zhang, Z.; Li, P.; Ding, B. Effects of Vitamin A on Immune Responses and Vitamin A Metabolism in Broiler Chickens Challenged with Necrotic Enteritis. Life 2023, 13, 1122. [Google Scholar] [CrossRef] [PubMed]
- Cortes, P.L.; Tiwary, A.K.; Puschner, B.; Crespo, R.M.; Chin, R.P.; Bland, M.; Shivaprasad, H.L. Vitamin A deficiency in turkey poults. J. Vet. Diagn. Investig. 2006, 18, 489–494. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). Scientific opinion of the panel on additives and products or substances used in animal feed (FEEDAP) on a request from the European Commission on the consequences for the consumer of the use of vitamin A in animal nutrition. EFSA J. 2008, 873, 1–81. [Google Scholar]
- Rautenschlein, S.; Ryll, M. Nicht-infektiöse Erkrankungen und Veränderungen. In Erkrankungen des Nutzgeflügels; Eugen Ulmer Verlag: Stuttgart, Germany, 2014; pp. 210–220. [Google Scholar]
- Hedman, H.D.; Vasco, K.A.; Zhang, L. A Review of Antimicrobial Resistance in Poultry Farming within Low-Resource Settings. Animals 2020, 10, 1264. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.S.R.; Çetingül, I.S. Nutritional advances in production performance and product quality of poultry husbandry under heat stress. Online J. Anim. Feed. Res. 2022, 12, 53–65. [Google Scholar]
- Abd El-Hack, M.E.; Alagawany, M.; Mahrose, K.M.; Arif, M.; Saeed, M.; Arain, M.A.; Soomro, R.N.; Siyal, F.A.; Fazlani, F.A.; Fowler, J. Productive performance, egg quality, hematological parameters and serum chemistry of laying hens fed diets supplemented with certain fat-soluble vitamins, individually or combined, during summer season. Anim. Nutr. 2019, 5, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Carazo, A.; Macákova, K.; Matoušová, K.; Krčmová, L.K.; Protti, M.; Mladěnka, P. Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity. Nutrients 2021, 13, 1703. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). Scientific Opinion on the Safety and Efficacy of Vitamin A (Retinyl Acetate, Retinyl Palmitate and Retinyl Propionate) as a Feed Additive for All Animal Species and Categories. EFSA J. 2013, 11, 3037. [Google Scholar] [CrossRef]
- Harrison, E.H.; Hussain, M.M. Mechanisms Involved in the Intestinal Digestion and Absorption of Dietary Vitamin A. J. Nutr. 2001, 131, 1405–1408. [Google Scholar] [CrossRef]
- Sideeg, R.M. Effect of Dietary Vitamin A and Nigella Sativa on the Performance of Broiler Chicks. Master’s Thesis, University of Khartoum, Khartoum, Sudan, 1996. [Google Scholar]
- Yin, H.D.; Tian, K.; Li, D.Y.; Gilbert, E.R.; Xiao, L.H.; Chen, S.Y.; Wang, Y.; Liu, Y.P.; Zhao, X.L.; Zhu, Q. Expression Profiles of Cellular Retinol-binding Protein, Type II (CRBP II) in Erlang Mountainous Chickens. Asian-Australas. J. Anim. Sci. 2014, 27, 310–315. [Google Scholar] [CrossRef]
- Senoo, H.; Imai, K.; Mezaki, Y.; Miura, M.; Morii, M.; Fujiwara, M.; Blomhoff, R. Accumulation of vitamin A in the hepatic stellate cell of arctic top predators. Anat. Rec. 2012, 295, 1660–1668. [Google Scholar] [CrossRef]
- Kim, D.H.; Lee, J.; Suh, Y.; Cressman, M.; Lee, K. Research Note: All-trans retinoic acids induce adipogenic differentiation of chicken embryonic fibroblasts and preadipocytes. Poult. Sci. 2020, 99, 7142–7146. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Y.; Li, R.; Chen, G. Transcriptional Factors Mediating Retinoic Acid Signals in the Control of Energy Metabolism. Int. J. Mol. Sci. 2015, 16, 14210–14244. [Google Scholar] [CrossRef]
- Sporn, M.B.; Roberts, A.B.; Goodman, D.S. The Retinoids; Academic Press: Orlando, FL, USA, 1984. [Google Scholar]
- Barua, A.B.; Olson, J.A. Retinoyl beta-glucuronide: An endogenous compound of human blood. Am. J. Clin. Nutr. 1986, 43, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Huan, J. Effect of Dietary Pyrrolizidine Alkaloids on Copper and Vitamin A Metabolism in the Chicken and Japanese Quail. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 1991. [Google Scholar]
- Kam, R.K.; Deng, Y.; Chen, Y.; Zhao, H. Retinoic acid synthesis and functions in early embryonic development. Cell. Biosci. 2012, 2, 11. [Google Scholar] [CrossRef]
- Yu, M.; Guan, K.; Zhang, C. The promoting effect of retinoic acid on proliferation of chicken primordial germ cells by increased expression of cadherin and catenins. Amino Acids 2010, 40, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Underhill, T.M.; Weston, A.D. Retinoids and their receptors in skeletal development. Microsc. Res. Tech. 1998, 43, 137–155. [Google Scholar] [CrossRef]
- Wang, J.L.; Swartz-Basile, D.A.; Rubin, D.C.; Levin, M.S. Retinoic acid stimulates early cellular proliferation in the adapting remnant rat small intestine after partial resection. J. Nutr. 1997, 127, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Seiler, K.M.; Waye, S.E.; Kong, W.; Kamimoto, K.; Bajinting, A.; Goo, W.H.; Onufer, E.J.; Courtney, C.; Guo, J.; Warner, B.W.; et al. Single-Cell Analysis Reveals Regional Reprogramming During Adaptation to Massive Small Bowel Resection in Mice. Cell. Mol. Gastroenterol. Hepatol. 2019, 8, 407–426. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Ghaffari, M.; Whitsett, J.A.; Zeng, X.; Sever, Z.; Lin, S. Retinoic acid-receptor activation of SP-B gene transcription in respiratory epithelial cells. Am. J. Physiol. Cell. Mol. Physiol. 1998, 275, L239–L246. [Google Scholar] [CrossRef]
- Fernandes-Silva, H.; Araújo-Silva, H.; Correia-Pinto, J.; Moura, R.S. Retinoic Acid: A Key Regulator of Lung Development. Biomolecules 2020, 10, 152. [Google Scholar] [CrossRef]
- Danielpour, D.; Song, K. Cross-talk between IGF-I and TGF-beta signaling pathways. Cytokine Growth Factor Rev. 2006, 17, 59–74. [Google Scholar] [CrossRef]
- Oliveira, L.M.; Teixeira, F.M.E.; Sato, M.N. Impact of Retinoic Acid on Immune Cells and Inflammatory Diseases. Mediat. Inflamm. 2018, 2018, 3067126. [Google Scholar] [CrossRef]
- Halevy, O.; Arazi, Y.; Melamed, D.; Friedman, A.; Sklan, D. Retinoic acid receptor-alpha gene expression is modulated by dietary vitamin A and by retinoic acid in chicken T lymphocytes. J. Nutr. 1994, 124, 2139–2146. [Google Scholar] [CrossRef]
- Liu, Z.-M.; Wang, K.-P.; Ma, J.; Zheng, S.G. The role of all-trans retinoic acid in the biology of Foxp3+ regulatory T cells. Cell Mol. Immunol. 2015, 12, 553–557. [Google Scholar] [CrossRef]
- Jiang, H.; Promchan, K.; Lin, B.R.; Lockett, S.; Chen, D.; Marshall, H.; Badralmaa, Y.; Natarajan, V. LZTFL1 Upregulated by All-Trans Retinoic Acid during CD4+ T Cell Activation Enhances IL-5 Production. J. Immunol. 2016, 196, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Jäger, A.; Kuchroo, V.K. Effector and regulatory T-cell subsets in autoimmunity and tissue inflammation. Scand. J. Immunol. 2010, 72, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.C.; Esterhazy, D.; Sarde, A.; London, M.; Pullabhatla, V.; Osma-Garcia, I.; Al-Bader, R.; Ortiz, C.; Elgueta, R.; Arno, M.; et al. Retinoic acid is essential for Th1 cell lineage stability and prevents transition to a Th17 cell program. Immunity 2015, 42, 499–511. [Google Scholar] [CrossRef]
- Bono, M.R.; Tejon, G.; Flores-Santibañez, F.; Fernandez, D.; Rosemblatt, M.; Sauma, D. Retinoic Acid as a Modulator of T Cell Immunity. Nutrients 2016, 8, 349. [Google Scholar] [CrossRef] [PubMed]
- Marks, E.; Ortiz, C.; Pantazi, E.; Bailey, C.S.; Lord, G.M.; Waldschmidt, T.J.; Noelle, R.J.; Elgueta, R. Retinoic Acid Signaling in B Cells Is Required for the Generation of an Effective T-Independent Immune Response. Front. Immunol. 2016, 7, 643. [Google Scholar] [CrossRef] [PubMed]
- Elias, P.M.; Friend, D.S. Vitamin-A-induced mucous metaplasia. An in vitro system for modulating tight and gap junction differentiation. J. Cell Biol. J. Cell Biol. 1976, 68, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Liu, S.; Liu, G.; Zhao, J.; Jiao, H.; Wang, X.; Song, Z.; Lin, H. Vitamin A Deficiency Impairs Mucin Expression and Suppresses the Mucosal Immune Function of the Respiratory Tract in Chicks. PLoS ONE. 2015, 10, e0139131. [Google Scholar] [CrossRef]
- Abdelhamid, L.; Luo, X.M. Retinoic Acid, Leaky Gut, and Autoimmune Diseases. Nutrients 2018, 10, 1016. [Google Scholar] [CrossRef]
- Lochbaum, R.; Schilpp, C.; Nonnenmacher, L.; Frick, M.; Dietl, P.; Wittekindt, O.H. Retinoic acid signalling adjusts tight junction permeability in response to air-liquid interface conditions. Cell. Signal 2020, 65, 109421. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Chang, S.H.; Martinez, G.J.; Yang, X.O.; Nurieva, R.; Kang, H.S.; Ma, L.; Watowich, S.S.; Jetten, A.M.; Tian, Q.; et al. Critical regulation of early Th17 cell differentiation by interleukin-1 signaling. Immunity 2009, 30, 576–587. [Google Scholar] [CrossRef]
- Reid, W.D.; Close, A.J.; Humphrey, S.; Chaloner, G.; Lacharme-Lora, L.; Rothwell, L.; Kaiser, P.; Williams, N.J.; Humphrey, T.J.; Wigley, P.; et al. Cytokine responses in birds challenged with the human food-borne pathogen Campylobacter jejuni implies a Th17 response. R. Soc. Open Sci. 2016, 3, 150541. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Allen, C.; Ballow, M. Retinoic acid enhances the production of IL-10 while reducing the synthesis of IL-12 and TNF-alpha from LPS-stimulated monocytes/macrophages. J. Clin. Immunol. 2007, 27, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhang, G.; Minton, J.E.; Ross, C.R.; Blecha, F. Regulation of cathelicidin gene expression: Induction by lipopolysaccharide, interleukin-6, retinoic acid, and Salmonella enterica serovar typhimurium infection. Infect. Immun. 2000, 68, 5552–5558. [Google Scholar] [CrossRef]
- Elloumi, H.Z.; Holland, S.M. Complex regulation of human cathelicidin gene expression: Novel splice variants and 5′UTR negative regulatory element. Mol. Immunol. 2008, 45, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Palace, V.P.; Khaper, N.; Qin, Q.; Singal, P.K. Antioxidant potentials of vitamin A and carotenoids and their relevance to heart disease. Free. Radic. Biol. Med. 1999, 26, 746–761. [Google Scholar] [CrossRef]
- Shastak, Y.; Gordillo, A.; Pelletier, W. The relationship between vitamin A status and oxidative stress in animal production. J. Appl. Anim. Res. 2023, 51, 546–553. [Google Scholar] [CrossRef]
- Landete, J.M. Dietary intake of natural antioxidants: Vitamins and polyphenols. Crit. Rev. Food Sci. Nutr. 2013, 53, 706–721. [Google Scholar] [CrossRef]
- Bohn, T.; Böhm, V.; Dulińska-Litewka, J.; Landrier, J.-F.; Bánáti, D.; Kucuk, O.; Borel, P.; Canas, J.A.; Rühl, R. Is vitamin A an antioxidant? Int. J. Vitam. Nutr. Res. 2022, a000752. [Google Scholar] [CrossRef]
- Tesoriere, L.; Ciaccio, M.; Bongiorno, A.; Riccio, A.; Pintaudi, A.; Livrea, M. Antioxidant activity of all-trans-retinol in homogeneous solution and in phosphatidylcholine liposomes. Arch. Biochem. Biophys. 1993, 307, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Ahlemeyer, B.; Bauerbach, E.; Plath, M.; Steuber, M.; Heers, C.; Tegtmeier, F.; Krieglstein, J. Retinoic acid reduces apoptosis and oxidative stress by preservation of SOD protein level. Free. Radic. Biol. Med. 2001, 30, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Szymański, L.; Skopek, R.; Palusińska, M.; Schenk, T.; Stengel, S.; Lewicki, S.; Kraj, L.; Kamiński, P.; Zelent, A. Retinoic Acid and Its Derivatives in Skin. Cells 2020, 9, 2660. [Google Scholar] [CrossRef]
- Gad, A.; Abu Hamed, S.; Khalifa, M.; Amin, A.; El-Sayed, A.; Swiefy, S.A.; El-Assal, S. Retinoic acid improves maturation rate and upregulates the expression of antioxidant-related genes in in vitro matured buffalo (Bubalus bubalis) oocytes. Int. J. Veter-Sci. Med. 2018, 6, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Chen, D.; Tian, G.; He, J.; Huang, Z.; Zheng, P.; Mao, X.; Yu, J.; Luo, J.; Luo, Y.; et al. All-Trans Retinoic Acid Attenuates Transmissible Gastroenteritis Virus-Induced Apoptosis in IPEC-J2 Cells via Inhibiting ROS-Mediated P38MAPK Signaling Pathway. Antioxidants 2022, 11, 345. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Andrés, C.M.C.; de la Lastra, J.M.P.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Chemistry of Hydrogen Peroxide Formation and Elimination in Mammalian Cells, and Its Role in Various Pathologies. Stresses 2022, 2, 256–274. [Google Scholar] [CrossRef]
- Mishra, B.; Jha, R. Oxidative Stress in the Poultry Gut: Potential Challenges and Interventions. Front. Veter. Sci. 2019, 6, 60. [Google Scholar] [CrossRef]
- Surai, P.F. Antioxidant Systems in Poultry Biology: Superoxide Dismutase. J. Anim. Res. Nutr. 2015, 1, 8. [Google Scholar] [CrossRef]
- Fu, Z.; Kato, H.; Sugahara, K.; Kubo, T. Retinoic acid accelerates the development of reproductive organs and egg production in Japanese quail (Coturnix coturnix japonica). Biol. Reprod. 2000, 63, 1795–1800. [Google Scholar] [CrossRef]
- Endo, T.; Mikedis, M.M.; Nicholls, P.K.; Page, D.C.; de Rooij, D.G. Retinoic Acid and Germ Cell Development in the Ovary and Testis. Biomolecules 2019, 9, 775. [Google Scholar] [CrossRef]
- Busada, J.T.; Geyer, C.B. The Role of Retinoic Acid (RA) in Spermatogonial Differentiation. Biol. Reprod. 2016, 94, 10. [Google Scholar] [CrossRef] [PubMed]
- Abdelnour, S.A.; Abd El-Hack, M.E.; Swelum, A.A.; Saadeldin, I.M.; Noreldin, A.E.; Khafaga, A.F.; Al-Mutary, M.G.; Arif, M.; Hussein, E.O.S. The Usefulness of Retinoic Acid Supplementation during In Vitro Oocyte Maturation for the In Vitro Embryo Production of Livestock: A Review. Animals 2019, 9, 561. [Google Scholar] [CrossRef]
- Wang, W.-D.; Hsu, H.-J.; Li, Y.-F.; Wu, C.-Y. Retinoic Acid Protects and Rescues the Development of Zebrafish Embryonic Retinal Photoreceptor Cells from Exposure to Paclobutrazol. Int. J. Mol. Sci. 2017, 18, 130. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Lee, J.; Kim, S.; Lillehoj, H.S.; Lee, K. Hypertrophy of Adipose Tissues in Quail Embryos by in ovo Injection of All-Trans Retinoic Acid. Front. Physiol. 2021, 12, 681562. [Google Scholar] [CrossRef] [PubMed]
- Mark, M.; Ghyselinck, N.B.; Chambon, P. Function of retinoic acid receptors during embryonic development. Nucl. Recept. Signal. 2009, 7, e002. [Google Scholar] [CrossRef]
- Duester, G. Retinoic acid synthesis and signaling during early organogenesis. Cell 2008, 134, 921–931. [Google Scholar] [CrossRef]
- Gur, M.; Bendelac-Kapon, L.; Shabtai, Y.; Pillemer, G.; Fainsod, A. Reduced Retinoic Acid Signaling During Gastrulation Induces Developmental Microcephaly. Front. Cell Dev. Biol. 2022, 10, 844619. [Google Scholar] [CrossRef]
- Aydelotte, M.B. Vitamin A deficiency in chickens. Br. J. Nutr. 1963, 17, 205–210. [Google Scholar] [CrossRef]
- Bridges, C.D.; Alvarez, R.A.; Fong, S.L.; Liou, G.I.; Ulshafer, R.J. Rhodopsin, vitamin A, and interstitial retinol-binding protein in the rd chicken. Invest. Ophthalmol. Vis. Sci. 1987, 28, 613–617. [Google Scholar] [PubMed]
- Kelley, M.W.; Turner, J.K.; Reh, T.A. Retinoic acid promotes differentiation of photoreceptors in vitro. Development 1994, 120, 2091–2102. [Google Scholar] [CrossRef]
- Lima, H.; Souza, L. Vitamin A in the diet of laying hens: Enrichment of table eggs to prevent nutritional deficiencies in humans. World's Poult. Sci. J. 2018, 74, 619–626. [Google Scholar] [CrossRef]
- Randhawa, S.R.; Deshmukh, S.; Banga, H.S.; Singh, N. Concomitant Vitamin A Deficiency Following Suspected Fowl pox virus Infection Leading to Esophageal Gland Metaplasia in a Layer Flock. J. Anim. Res. 2018, 8, 1087–1090. [Google Scholar] [CrossRef]
- Kleyn, R.; Chrystal, P. Vitamins. In Broiler Nutrition: Masterclass; Context Products Ltd.: Leicestershire, UK, 2020; pp. 129–142. [Google Scholar]
- Al-Deseit, B. Least-Cost Broiler Ration Formulation Using Linear Programming Technique. J. Anim. Vet. Advan. 2009, 8, 1274–1278. [Google Scholar]
- Mallick, P.; Muduli, K.; Biswal, J.N.; Pumwa, J. Broiler Poultry Feed Cost Optimization Using Linear Programming Technique. J. Oper. Strat. Plan. 2020, 3, 31–57. [Google Scholar] [CrossRef]
- Yang, P.; Wang, H.K.; Zhu, M.; Li, L.X.; Ma, Y.X. Degradation kinetics of vitamins in premixes for pig: Effects of choline, high concentrations of copper and zinc, and storage time. Anim. Biosci. 2021, 34, 701–713. [Google Scholar] [CrossRef]
- Hirai, R.A.; De Leon, D.; Randig-Biar, M.; Silva, A.; Sanchez, E.; McElroy, A.P.; Bailey, C.A.; Martinez, N.; Sokale, A.; Music, L. Evaluation of the stability of vitamin A acetate concentrates mixed in a vitamin-trace mineral premix over a 56-day high temperature and humidity storage stress. In Proceedings of the 2023 The International Poultry Scientific Forum, Atlanta, GA, USA, 23–24 January 2023; p. 112. [Google Scholar]
- Abdollahi, M.R.; Ravindran, V.; Svihus, B. Pelleting of broiler diets: An overview with emphasis on pellet quality and nutritional value. Anim. Feed. Sci. Technol. 2013, 179, 1–23. [Google Scholar] [CrossRef]
- Baker, D.H. Bioavailability of minerals and vitamins. In Swine Nutrition, 2nd ed.; Lewisand, A.J., Southern, L.L., Eds.; CRC Press LLC: Boca Raton, FL, USA, 2001; pp. 357–379. [Google Scholar]
- Leeson, S.; Summers, J.D. Commercial Poultry Nutrition. In Scott’s Nutrition of the Chicken, 3rd ed.; Leeson, S., Ed.; Nottingham University Press: Nottingham, UK, 2001; p. 398. [Google Scholar]
- Oviedo-Rondón, E.O. Holistic view of intestinal health in poultry. Anim. Feed. Sci. Technol. 2019, 250, 1–8. [Google Scholar] [CrossRef]
- Baker, D.H.; Stein, H.H. Bioavailability of Minerals and Vitamins in Feedstuffs. In Sustainable Swine Nutrition; Chiba, L.I., Ed.; Wiley-Blackwell Press: Ames, IA, USA, 2013; pp. 341–364. [Google Scholar]
- Teleki, A.; Hitzfeld, A.; Eggersdorfer, M. 100 Years of Vitamins: The Science of Formulation is the Key to Functionality. KONA Powder Part. J. 2013, 30, 144–163. [Google Scholar] [CrossRef]
- Haring, C.M.; Jaffa, M.E.; Beach, J.R. A nutritional disease of poultry. Calif. Sta. Rpt. 1920, 79. Available online: https://books.google.by/books?id=p-M4AAAAIAAJ&pg=PA684&lpg=PA684&dq=A+nutritional+disease+of+poultry++Calif.+Sta.+Rpt.&source=bl&ots=4Jt1Lw7piP&sig=ACfU3U1Qdh0-aZ_SdallrDmuhjWi-cMN9A&hl=en&sa=X&ved=2ahUKEwij8fG5hdqCAxXaPxAIHSugDTs4ChDoAXoECAIQAw#v=onepage&q=%20poultry&f=false (accessed on 3 August 2023).
- Hinshaw, W.R.; Lloyd, W.E. Vitamin-A defiency in turkeys. Hilgardia 1934, 8, 283–304. [Google Scholar] [CrossRef]
- Wolbach, S.B.; Hegsted, D.M. Vitamin A deficiency in the duck. Skeletal growth and the central nervous system. Arch. Pat Hot. 1952, 54, 548–563. [Google Scholar]
- Wilke, H.L. Vitamins In Poultry Nutrition; Iowa State University Veterinarian: Ames, IA, USA, 1940; Volume 2, p. 8. Available online: https://lib.dr.iastate.edu/iowastate_veterinarian/vol2/iss3/8 (accessed on 16 July 2023).
- Abercrombie, G.S. Vitamin Requirements of Growing Japanese Quail (Coturnix coturnix japonica). Master’s Thesis, Oregon State University, Oregon, OR, USA, 1966. [Google Scholar]
- Uni, Z.; Zaiger, G.; Gal-Garber, O.; Pines, M.; Rozenboim, I.; Reifen, R. Vitamin A deficiency interferes with proliferation and maturation of cells in the chicken small intestine. Br. Poult. Sci. 2000, 41, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, A.R.; Lauridsen, C.; Howlider, A.R.; Jakobsen, K. Importance of vitamin A supplementation for performance of Sonali chickens under smallholder farm conditions in a tropical climate. Livest. Res. Rural Dev. 2004, 16, 83. [Google Scholar]
- Feng, Y.L.; Xie, M.; Tang, J.; Huang, W.; Zhang, Q.; Hou, S.S. Effects of vitamin A on growth performance and tissue retinol of starter White Pekin ducks. Poult. Sci. 2019, 98, 2189–2192. [Google Scholar] [CrossRef]
- Xiao, X.; Liang, J.R.; Yang, H.M.; Wan, X.L.; Wang, Z.Y. Vitamin A deficiency or critical excess has negative effects on the growth performance, slaughter performance, and meat quality of goslings. Anim. Feed Sci. Technol. 2021, 280, 115064. [Google Scholar] [CrossRef]
- Swindell, E.C.; Eichele, G. Retinoid metabolizing enzymes in development. BioFactors 1999, 10, 85–89. [Google Scholar] [CrossRef]
- Fu, Z.W.; Kubo, T.; Sugahara, K.; Noguchi, T.; Kato, H. Retinoid nutritional status differently affects the expression of Japanese quail retinoic acid receptor-beta isoform transcripts in a tissue-specific manner. J. Endocrinol. 2001, 169, 281–290. [Google Scholar] [CrossRef]
- Fu, Z.; Noguchi, T.; Kato, H. Differential regulation of IGFBP-2 and IGFBP-5 gene expression by vitamin A status in Japanese quail. Am. J. Physiol. Metab. 2001, 281, E138–E146. [Google Scholar] [CrossRef]
- Sijtsma, S.; Rombout, J.; West, C.; van der Zijpp, A. Vitamin A deficiency impairs cytotoxic T lymphocyte activity in Newcastle disease virus-infected chickens. Vet. Immunol. Immunopathol. 1990, 26, 191–201. [Google Scholar] [CrossRef]
- Dalloul, R.; Lillehoj, H.; Shellem, T.; Doerr, J. Effect of vitamin A deficiency on host intestinal immune response to Eimeria acervulina in broiler chickens. Poult. Sci. 2002, 81, 1509–1515. [Google Scholar] [CrossRef]
- Titus, H.W.; Nutritional Diseases of Poultry. Yearbook of Agriculture 1075–1108. 1942. Available online: https://naldc.nal.usda.gov/download/IND43893805/PDF (accessed on 28 July 2023).
- Aye, P.P.; Morishita, T.Y.; Saif, Y.M.; Latshaw, J.D.; Harr, B.S.; Cihla, F.B. Induction of vitamin A deficiency in turkeys. Avian Dis. 2000, 44, 809–817. [Google Scholar] [CrossRef]
- Leutskaya, Z.; Fais, D. Antibody synthesis stimulation by vitamin A in chickens. Biochim. Biophys. Acta 1977, 475, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Bang, B.G.; Bang, F.B.; Foard, M.A. Lymphocyte depression induced in chickens on diets deficient in vitamin A and other components. Am. J. Pathol. 1972, 68, 147–162. [Google Scholar] [PubMed]
- Idi, A.; Permin, A.; Jensen, S.K.; Murrell, K.D. Effect of a minor vitamin A deficiency on the course of infection with Ascaridia galli (Schrank, 1788) and the resistance of chickens. Helminthologia 2007, 44, 3–9. [Google Scholar] [CrossRef]
- Asmundson, V.S.; Kratzer, F.H. Observation on vitamin A deficiency in turkey breeding stock. Poult. Sci. 1952, 31, 71–73. [Google Scholar] [CrossRef]
- Sommer, A. Vitamin A deficiency and clinical disease: An historical overview. J. Nutr. 2008, 138, 1835–1839. [Google Scholar] [CrossRef] [PubMed]
- Waters, J.W. Effect of Vitamin A-Deficiency on the Dark Adaptation of the Pigeon. Nature 1952, 169, 484. [Google Scholar] [CrossRef]
- Fu, Z.; Kato, H.; Sugahara, K.; Kubo, T. Vitamin A deficiency reduces the responsiveness of pineal gland to light in Japanese quail (Coturnix japonica). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1998, 119, 593–598. [Google Scholar] [CrossRef]
- Chen, F.; Jiang, Z.; Jiang, S.; Li, L.; Lin, X.; Gou, Z.; Fan, Q. Dietary Vitamin A Supplementation Improved Reproductive Performance by Regulating Ovarian Expression of Hormone Receptors, Caspase-3 and Fas in Broiler Breeders. Poult. Sci. 2015, 95, 30–40. [Google Scholar] [CrossRef]
- Thompson, J.N.; Howell, J.M.; Pitt, G.A.J.; McLaughlin, C.I. The biological activities of retinoic acid in the domestic fowl and effects of vitamin A deficiency on the chick embryo. Br. J. Nutr. 1969, 23, 471–490. [Google Scholar] [CrossRef]
- Bermudez, A.J.; Swayne, D.E.; Squires, M.W.; Radin, M.J. Effects of vitamin A deficiency on the reproductive system of mature White Leghorn hens. Avian Dis. 1993, 37, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Weber, G. Improvement of flock productivity through supply of vitamins for higher laying performance and better egg quality. World's Poult. Sci. J. 2009, 65, 443–458. [Google Scholar] [CrossRef]
- Maden, M.; Graham, A.; Gale, E.; Rollinson, C.; Zile, M. Positional apoptosis during vertebrate CNS development in the absence of endogenous retinoids. Development 1997, 124, 2799–2805. [Google Scholar] [CrossRef] [PubMed]
- Howell, J.M.; Thompson, J.N. Lesions in nervous tissue and bone in ataxic vitamin A deficient quail. Acta Neuropathol. 1970, 16, 285–292. [Google Scholar] [CrossRef]
- Friedman, A.; Sklan, D. Impaired T lymphocyte immune response in vitamin A depleted rats and chicks. Br. J. Nutr. 1989, 62, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Sklan, D.; Melamed, D.; Friedman, A. The effect of varying levels of dietary vitamin A on immune response in the chick. Poult. Sci. 1994, 73, 843–847. [Google Scholar] [CrossRef]
- Sklan, D.; Melamed, D.; Friedman, A. The effect of varying levels of vitamin A on immune response of the turkey. Br. Poult. Sci. 1995, 36, 385–392. [Google Scholar] [CrossRef]
- Davis, C.Y.; Sell, J.L. Effect of all-trans retinol and retinoic acid nutriture on the immune system of chicks. J. Nutr. 1983, 113, 1914–1919. [Google Scholar] [CrossRef]
- Sanda, M.E.; Oyewole, B.O. Effects of vitamins A and C on performance, immune response and haematology of broilers vaccinated against Newcastle disease. Int. J. Res. Stud. Biosci. 2015, 3, 37–42. [Google Scholar]
- Rizvi, F.; Asghar, K.; Hafeez, M.A. Effect of vitamin A deficient diet on immune response in Newcastle disease infected broilers. Pak. J. Life Soc. Sci. 2003, 1, 13–16. [Google Scholar]
- Zhang, L.; Hou, Y.; Ma, Z.; Xie, J.; Fan, J.; Jiao, Y.; Wang, F.; Han, Z.; Liu, S.; Ma, D. Effect of oral vitamin A supplementation on host immune response to infectious bronchitis virus infection in SPF chicken. Poult. Sci. 2023, 102, 102701. [Google Scholar] [CrossRef] [PubMed]
- Sepehri Moghaddam, H.; Emadi, M. The effect of threonine and vitamin a on immune system in broiler chickens. Int. J. Adv. Biol. Biomed. Res. 2014, 2, 756–763. [Google Scholar]
- Guo, X.; Han, J.; Yan, S.; Li, Y.; Shi, B.; Zhao, Y. Effects of vitamin A on growth performance, immunity and antioxidant unction of Broilers. Chin. J. Anim. Nutr. 2019, 31, 3582–3589. [Google Scholar]
- Lessard, M.; Hutchings, D.; Cave, N.A. Cell-mediated and humoral immune responses in broiler chickens maintained on diets containing different levels of vitamin A. Poult. Sci. 1997, 76, 1368–1378. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Roshdy, A.R.; Guo, Y.; Wang, Y.; Guo, S. Effect of dietary vitamin A on reproductive performance and immune response of broiler breeders. PLoS ONE 2014, 9, e105677. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.L.; Zheng, X.C.; Liang, J.R.; Xiao, X.; Yang, H.M.; Wang, Z.Y. Dietary vitamin A supplementation improves intestinal morphology and immune performance of goslings. J. Anim. Feed. Sci. 2022, 31, 217–223. [Google Scholar] [CrossRef]
- Yang, H.; Liang, J.; Dai, H.; Wan, X.; Wang, Z. Effects of vitamin A supplementation in the diet of breeding geese on offspring intestinal tissue morphology and immune performance. Asian-Australas. J. Anim. Sci. 2020, 33, 1463–1469. [Google Scholar] [CrossRef]
- Li, L.; Liu, Z.; Fang, B.; Xu, J.; Dong, X.; Yang, L.; Zhang, Z.; Guo, S.; Ding, B. Effects of vitamin A and K3 on immune function and intestinal antioxidant capacity of aged laying hens. Braz. J. Poult. Sci. 2022, 24, eRBCA-2021-1572. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; Mahrose, K.; Askar, A.A.; Alagawany, M.; Arif, M.; Saeed, M.; Abbasi, F.; Soomro, R.N.; Siyal, F.A.; Chaudhry, M.T. Single and Combined Impacts of Vitamin A and Selenium in Diet on Productive Performance, Egg Quality, and Some Blood Parameters of Laying Hens During Hot Season. Biol. Trace Elem. Res. 2017, 177, 169–179. [Google Scholar] [CrossRef]
- Jukes, T.H. Experiments on the storage of vitamin A by growing turkeys. Poult. Sci. 1942, 21, 357–360. [Google Scholar] [CrossRef]
- Abd El-Wahab, A.; Visscher, C.; Ratert, C.; Kölln, M.; Diephaus, D.; Beineke, A.; Kamphues, J. Outcome of an Experimental Study in Growing Turkeys Suspected of Having a Diet Related, Uncommon and Uncoordinated Gait. Vet. Sci. 2017, 4, 49. [Google Scholar] [CrossRef] [PubMed]
- Seifried, O. Studies of A-avitaminosis in chickens. l. Lesions of the respiratory tract and their relations to some infectious disease. J. Exp. Med. 1930, 52, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Seifried, O. Studies on A Avitaminosis in chickens. II. Lesions of the upper alimentary tract and their relation to some infectious diseases. J. Exp. Med. 1930, 52, 533–538. [Google Scholar] [CrossRef]
- Howell, J.M.; Thompson, J.N. Observations on the lesions in vitamin A deficient adult fowls with particular reference to changes in bone and central nervous system. Br. J. Exp. Pathol. 1967, 48, 450–454. [Google Scholar] [PubMed]
- Howell, J.M.; Thompson, J.N. Vitamin A deficiency in chickens and CNS Changes. Nutr. Rev. 1965, 23, 313–314. [Google Scholar] [CrossRef]
- Adamstone, F.B. Histologic comparison of the brains of vitamin A-deficient and vitamin E-deficient chicks. Arch. Pathol. 1947, 43, 301–312. [Google Scholar]
- Coggeshall, R.E.; Bieri, J.G. Pathology of the brain in single and mixed deficiencies of vitamins A and E in the chick. J. Nutr. 1960, 70, 272–277. [Google Scholar] [CrossRef]
- Thompson, J.N.; Howell, J.M.; Pitt, G.A.J.; Houghton, C.I. Biological activity of retinoic acid ester in the domestic fowl: Production of vitamin a deficiency in the early chick embryo. Nature 1965, 205, 1006–1007. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Gou, Z.; Chen, F.; Fan, Q.; Lin, X.; Ye, J.; Zhang, C.; Jiang, S. Effects of maternal and dietary vitamin A on growth performance, meat quality, antioxidant status, and immune function of offspring broilers. Poult. Sci. 2020, 99, 3930–3940. [Google Scholar] [CrossRef]
- Donalson, L.M.; Kim, W.K.; Woodward, C.; Herrera, P.; Kubena, L.F.; Nisbet, D.J.; Ricke, S.C. Utilizing different ratios of alfalfa and layer ration for molt induction and performance in commercial laying hens. Poult. Sci. 2005, 84, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Grashorn, M. Feed additives for influencing chicken meat and egg yolk color. In Handbook on Natural Pigments in Food and Beverages; Carle, R., Schweigert, R.M., Eds.; Woodhead Publishing: Sawston, UK, 2016; pp. 283–302. [Google Scholar]
- Schiedt, K.; Leuenberger, F.J.; Vecchi, M.; Glinz, E. Absorption, retention and metabolic transformations of carotenoids in rainbow trout, salmon and chicken. Pure Appl. Chem. 1985, 57, 685–692. [Google Scholar] [CrossRef]
- Brush, A.H. Metabolism of carotenoid pigments in birds. FASEB J. 1990, 4, 2969–2977. [Google Scholar] [CrossRef] [PubMed]
- Wilder, O.; Bethke, R. The Loss of Carotene in Machine-Dried Alfalfa Meal Under Variable Conditions of Storage. Poult. Sci. 1941, 20, 304–312. [Google Scholar] [CrossRef]
- Park, Y.; Anderson, M.; Walters, J.; Mahoney, A. Effects of processing methods and agronomic variables on carotene contents in forages and predicting carotene in alfalfa hay with near-infrared-reflectance spectroscopy. J. Dairy Sci. 1983, 66, 235–245. [Google Scholar] [CrossRef]
- Zurak, D.; Grbeša, D.; Duvnjak, M.; Kiš, G.; Međimurec, T.; Kljak, K. Carotenoid Content and Bioaccessibility in Commercial Maize Hybrids. Agriculture 2021, 11, 586. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. Vitamin A supply in swine production: Current science and practical considerations. Appl. Anim. Sci. 2023, 39, 289–305. [Google Scholar] [CrossRef]
- Akinyemi, F.; Adewole, D. Environmental Stress in Chickens and the Potential Effectiveness of Dietary Vitamin Supplementation. Front. Anim. Sci. 2021, 2, 775311. [Google Scholar] [CrossRef]
- EMA (European Medicines Agency). Injectable Veterinary Medicinal Products Containing Vitamin A for Use in Food Producing Species. An Official Website of the European Union. 2021. Available online: https://www.ema.europa.eu/en/medicines/veterinary/referrals/injectable-veterinary-medicinal-products-containing-vitamin-use-food-producing-species (accessed on 27 September 2023).
- Goudah, A.; El Sooud, K.A.; El-Aty, A.M.A. Pharmacokinetics and tissue residue profiles of erythromycin in broiler chickens after different routes of administration. Dtsch. Tierarztl. Wochenschr. 2004, 111, 162–165. [Google Scholar]
- Hezarjaribi, A.; Rezaeipour, V.; Abdollahpour, R. Effects of intramuscular injections of vitamin E-selenium and a gonadotropin releasing hormone analogue (GnRHa) on reproductive performance and blood metabolites of post-molt male broiler breeders. Asian Pac. J. Reprod. 2016, 5, 156–160. [Google Scholar] [CrossRef]
- Wolbach, S.B.; Hegsted, D.M. Hypervitaminosis A and the skeleton of growing chicks. Arch. Pathol. 1952, 54, 30–38. [Google Scholar]
- NASEM (National Academies of Sciences, Engineering, and Medicine). Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Veltmann, J.R., Jr.; Jensen, L. Vitamin A toxicosis in the chick and turkey poults. Poult. Sci. 1986, 65, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Coskun, B.; Inal, F.; Celik, I.; Erganis, O.; Tiftik, A.M.; Kurtoglu, F.; Kuyucuoglu, Y.; Ok, U. Effects of dietary levels of vitamin A on the egg yield and immune responses of laying hens. Poult. Sci. 1998, 77, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Bozhkov, A.; Ionov, I.; Kurhuzova, N.; Novikova, A.; Katerynych, O.; Akzhyhitov, R. Vitamin A intake forms resistance to hypervitaminosis A and affects the functional activity of the liver. Clin. Nutr. Open Sci. 2021, 41, 82–97. [Google Scholar] [CrossRef]
- GfE (Gesellschaft für Ernährungsphysiologie). Empfehlungen zur Energie- und Nährstoffversorgung von Legehennen und Masthühnern (Broiler); DLG-Verlag: Frankfurt, Germany, 1999. [Google Scholar]
- Cobb 500 Broiler Performance & Nutrition Supplement. 2022. Available online: https://www.cobb-vantress.com/assets/Cobb-Files/product-guides/5502e86566/2022-Cobb500-Broiler-Performance-Nutrition-Supplement.pdf (accessed on 28 July 2023).
- ROSS Broiler: Nutrition Specifications. 2022. Available online: https://aviagen.com/eu/brands/ross/products/ross-308 (accessed on 28 July 2023).
- Nicholas and, B.U.T. Heavy Lines Feeding Guidelines. 2015. Available online: https://www.aviagenturkeys.com/uploads/2015/11/20/NU06%20Feeding%20Guidelines%20for%20Nicholas%20&%20BUT%20Heavy%20Lines%20EN.pdf (accessed on 28 July 2023).
- Lohmann LSL-Lite Management Guide. 2014. Available online: https://lohmann-breeders.com/media/strains/cage/management/LOHMANN-LSL-Lite-Cage-1.pdf (accessed on 25 August 2016).
- Taghinejad-Roudbaneh, M.; Babaee, M.J.; Afrooziyeh, M.; Alizadeh, B. Estimation of Dietary Threonine Requirement for Growth and Immune Responses of Broilers. J. Appl. Anim. Res. 2013, 41, 474–483. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. The role of vitamin A in non-ruminant immunology. Front. Anim. Sci. 2023, 4, 1197802. [Google Scholar] [CrossRef]
- Rostangno, H.S.; Albino, L.F.T.; Donzele, J.L.; Gomez, P.C.; de Oliviera, R.F.; Lopez, D.C.; Ferreira, A.S.; de Toledo Barreto, S.L.; Euclides, R.F. Brazilian Tables for Poultry and Swine. In Composition of Feedstuffs and Nutritional Requirements, 3rd ed.; Rostagno, H.S., Ed.; Universidade Federal de Viçosa-Departamento de Zootecnia: Viçosa, Brazil, 2011. [Google Scholar]
- De Blas, C.; Mateos, G.G.; Rebollar, P.G.; FEDNA (Fundacion Espanola Deserrollo Nutricion Animal). Necesidades nutricionales para avicultura. Normas FEDNA. In Fundacion Espanola para el Desarrollo Nutricion Animal, 2nd ed.; Santoma, G., Mateos, G.G., Eds.; FEDNA: Madrid, Spain, 2018. [Google Scholar]
- Hubbard Management Guide. 2006. Available online: https://www.hubbardbreeders.com/media/manual_broiler_management_en___013796700_1441_27062016.pdf (accessed on 28 July 2023).
- Hybrid Turkeys Vitamin and Trace Mineral Supplementation. 2016. Available online: https://www.hybridturkeys.com/en/resources/commercial-management/feed-and-water/vitamin-and-trace-mineral-supplementation/ (accessed on 28 July 2023).
- Hy-Line W-36 Commercial Layers. 2020. Available online: https://www.hyline.com/filesimages/Hy-Line-Products/Hy-Line-Product-PDFs/W-36/36%20COM%20ENG.pdf (accessed on 25 July 2023).
- H&N Management Guide. 2016. Available online: https://www.hngb.co.uk/uploadedresources/1652881742-use_this_one_-_brown_management_guide_feb_22__version_6.pdf (accessed on 23 February 2018).
- ISA Nutrition Management Guide. 2018. Available online: https://www.isa-poultry.com/en/product/isa-brown/ (accessed on 22 February 2018).
- Dekalb Nutrition Management Guide. 2017. Available online: https://www.dekalbpoultry (accessed on 22 February 2018).
- Babcock Nutrition Management Guide. 2017. Available online: https://www.babcock-poultry.com/en/news/laying-hens-management-guide-alternative-systems/ (accessed on 22 February 2018).
- Hisex Nutrition Management Guide. 2017. Available online: https://www.hisex.com/en/news/laying-hens-management-guide-alternative-systems/ (accessed on 22 February 2018).
- Koutsos, E.A.; Klasing, K.C. Vitamin A nutrition of growing cockatiel chicks (Nymphicus hollandicus). J. Anim. Physiol. Anim. Nutr. 2005, 89, 379–387. [Google Scholar] [CrossRef]
- Sibbald, I.; Slinger, S. The effects of breed, sex, an arsenical and nutrient density on the utilization of dietary energy. Poult. Sci. 1963, 42, 1325–1332. [Google Scholar] [CrossRef]
- Leenstra, F.; Sambeek, F. Breeding of Laying Hens. LowInputBreeds Technical Note. Available online: www.lowinputbreeds.org (accessed on 27 September 2023).
- McDowell, L.R.; Ward, N.E. Optimum vitamin nutrition for poultry. Int. Poult. Prod. 2008, 16, 27–34. [Google Scholar]



{kind=link}
{kind=link}
| Disorder | Poultry Species | Manifestation | Reference |
|---|---|---|---|
| Immune deficiency | Chick | Reduced T lymphocyte activity after a challenge | [120] |
| Chick, turkey poult | Reduced antibody production and T cell proliferative response after a challenge | [121,122] | |
| Chick | Decreased cytotoxic T lymphocyte activity after a vaccination | [103] | |
| Chick | Reduced growth of the bursa of Fabricius and thymus | [123] | |
| Turkey | Squamous metaplasia and hyperkeratinization of glandular epithelium in bursa of Fabricius | [11] | |
| Broiler chicken | Lowered haemagglutination inhibition titers after a vaccination | [124] | |
| Broiler chicken | Fewer intraepithelial lymphocytes expressing surface markers CD3, CD4, CD8, αβTCR, and γβTCR after a challenge; higher shedding of Eimeria oocysts and lower levels of interferon-γ | [104] | |
| Broiler chicken | Elevated excretion of A. galli eggs | [109] | |
| Broiler chicken | Lower geometric antibody mean titer | [125] | |
| Broiler chicken | Increased viral replication and reduced serum immunoglobulin G levels | [126] | |
| Broiler chicken | Reduced cutaneous basophil hypersensitivity and heterophil and lymphocyte counts | [127] | |
| Broiler chicken | Reduced cytokine production | [128] | |
| Broiler breeder | Reduction in antibody titers after a vaccination | [129,130] | |
| Goose | Lower serum immunoglobulin A and G levels | [131] | |
| Goose | Lower immune organ weight, immune organ index, and immunoglobulin content | [132] | |
| Laying hen | Lower plasma immunoglobulin G content and mRNA expression of interleukin-10 in the spleen | [133] | |
| Reproductive dysfunction | Laying hen | Decrease in egg production and lower hatchability rates | [94] |
| Turkey breeder | Decrease in egg production, lower fertility and hatchability rates, and an increase in embryonic mortality | [110] | |
| Broiler breeder | Reduced ovarian stromal and white follicle weights along with oviduct weight and length; reduction in insulin-like growth factor 1 receptor transcripts in the ovarian stroma and the walls of yellow follicles, follicle stimulating hormone receptor expression in the walls of white and yellow follicles, and luteinizing hormone receptor and growth hormone receptor transcripts in the walls of yellow follicles; expression of apoptosis gene transcripts in the ovary; reduced egg weight | [114] | |
| Cock | Reduction in testicular size, loss of spermatids, and degeneration of the seminiferous epithelium | [115] | |
| Laying hen | Increase in the number of atretic follicles, including moderate to severe hemorrhages | [116] | |
| Laying hen | Reduction in egg number, egg mass, and albumen quality | [134] | |
| Abnormalities in digestive, respiratory, cardiovascular and urinary system | Turkey breeder | Pustules in the mouth and esophagus, urates in the ureters and kidneys | [110] |
| Turkey | Squamous metaplasia and hyperkeratinization of glandular epithelium in the esophagus, proventriculus, and nasal glands | [11] | |
| Turkey | Pustules in the mouth and pharynx, urates in the intestinal mesentery and pericardium | [135] | |
| Turkey | Squamous metaplasia in the oesophagus, hyperuricemia | [136] | |
| Chicken | Gross lesions characterized by the presence of white to yellowish caseous, coalescing nodules in the mucous glands and ducts of the pharynx and esophagus | [137,138] | |
| Goose | Reduced villus height and width, crypt depth, and muscular layer thickness in the duodenum, jejunum, and ileum | [131] | |
| Chick | Metaplasia of the nasal respiratory epithelium to a stratified squamous form | [119] | |
| Turkey | Squamous metaplasia affecting the epithelia of the digestive, respiratory, and urinary tracts | [35,106] | |
| Neurological maladies | Chick | Constriction of the central nervous system and the occurrence of Wallerian degeneration specifically in the ventral and lateral columns of the cervical spinal cord | [119] |
| Laying hen | Herniation of cerebral tissue, thinning of the cerebellar flocculi, ridging of the lumbosacral spinal cord, and early Wallerian degeneration in the cervical cord | [139] | |
| Chicken | Constriction of the brain and spinal cord | [140] | |
| Chick | Pin-pointed areas of degeneration in the brain stem, the base of the cerebellum, the optic chiasma, and the cerebrum | [141] | |
| Chick | The brains exhibited dispersed pyknotic neurons, predominantly concentrated in the optic tectum and the Purkinje cell layer of the cerebellum | [142] | |
| Japanese quail | Abnormalities in neural tissue | [119] | |
| Ophthalmic anomalies | Turkey | Increased lacrimation, inflammation of the nictitating membrane | [92] |
| Pigeon | Decrease in retinal sensitivity | [112] | |
| Chicken | Conjunctivitis, corneal ulcers, and excessive lacrimation | [78] | |
| Japanese quail | Diminished sensitivity of the pineal gland to light stimuli | [113] | |
| Duck | Keratoconjunctivitis sicca | [98] | |
| Turkey | Development of watery eyes followed by the accumulation of caseated matter in the eyes and blindness | [110] | |
| Skeletal disorders | Chick | Pathological changes in the epiphyseal cartilage zone of the femur and in the periosteum of the vertebrae; the vertebral periosteum exhibited heightened osteoblast activity and excessive cartilage formation | [119] |
| Japanese quail | Overgrowth of periosteum | [143] | |
| Duck | Altered endochondral bone growth leading to disproportional axial skeleton development | [93] | |
| Laying hen | Lesions in the bone tissue | [139] | |
| Hindered growth and development | Chick | Hindered potential for growth | [94] |
| Broiler chick | Reduced growth performance | [121] | |
| Turkey poult | Stunted growth | [122] | |
| Broiler chicken | Reduced growth performance | [128] | |
| Duck | Decreased growth | [98] | |
| Broiler chicken | Diminished growth | [144] |
| Source | Vitamin A Requirement, IU/kg Feed | |||
|---|---|---|---|---|
| Broilers | Laying Hens | Broiler Breeders | Turkeys | |
| NASEM [158] | 1500 | 3750 4 | n/a | 5000 |
| GfE [162] | 2500 | 4500 | 4500 | n/a |
| Brazilian tables [169] | 7053–13,538 | 9000–12,216 | 11,000 | n/a |
| FEDNA [170] | 7000–10,000 | 9000–10,000 | 9000–10,000 | 8000–13,000 |
| Cobb [163] | 10,000–13,000 | n/a | 10,000–13,000 | n/a |
| Aviagen 1 [164,165] | 10,000–13,000 | n/a | 10,000–12,000 | 5000–12,000 |
| Hubbard [171] | 10,000–15,000 | n/a | 12,000–14,000 | n/a |
| Hendrix (Hybrid turkeys) [172] | n/a | n/a | n/a | 9500–12,500 |
| EW Group (laying hens) 2 [166,173,174] | n/a | 8000–13,000 | n/a | n/a |
| Hendrix (laying hens) 3 [175,176,177,178] | n/a | 10,000–13,000 | n/a | n/a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shastak, Y.; Pelletier, W. Nutritional Balance Matters: Assessing the Ramifications of Vitamin A Deficiency on Poultry Health and Productivity. Poultry 2023, 2, 493-515. https://doi.org/10.3390/poultry2040037
Shastak Y, Pelletier W. Nutritional Balance Matters: Assessing the Ramifications of Vitamin A Deficiency on Poultry Health and Productivity. Poultry. 2023; 2(4):493-515. https://doi.org/10.3390/poultry2040037
Chicago/Turabian StyleShastak, Yauheni, and Wolf Pelletier. 2023. "Nutritional Balance Matters: Assessing the Ramifications of Vitamin A Deficiency on Poultry Health and Productivity" Poultry 2, no. 4: 493-515. https://doi.org/10.3390/poultry2040037