
Nutrigenomics in Animal Feeding: Digital Gene Expression Analysis in Poultry Fed Tenebrio molitor Larvae Meal
 , ,
, ,  , , ,
, , ,  ,
,  , , ,
, , ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Sample Preparation, and RNA Extraction
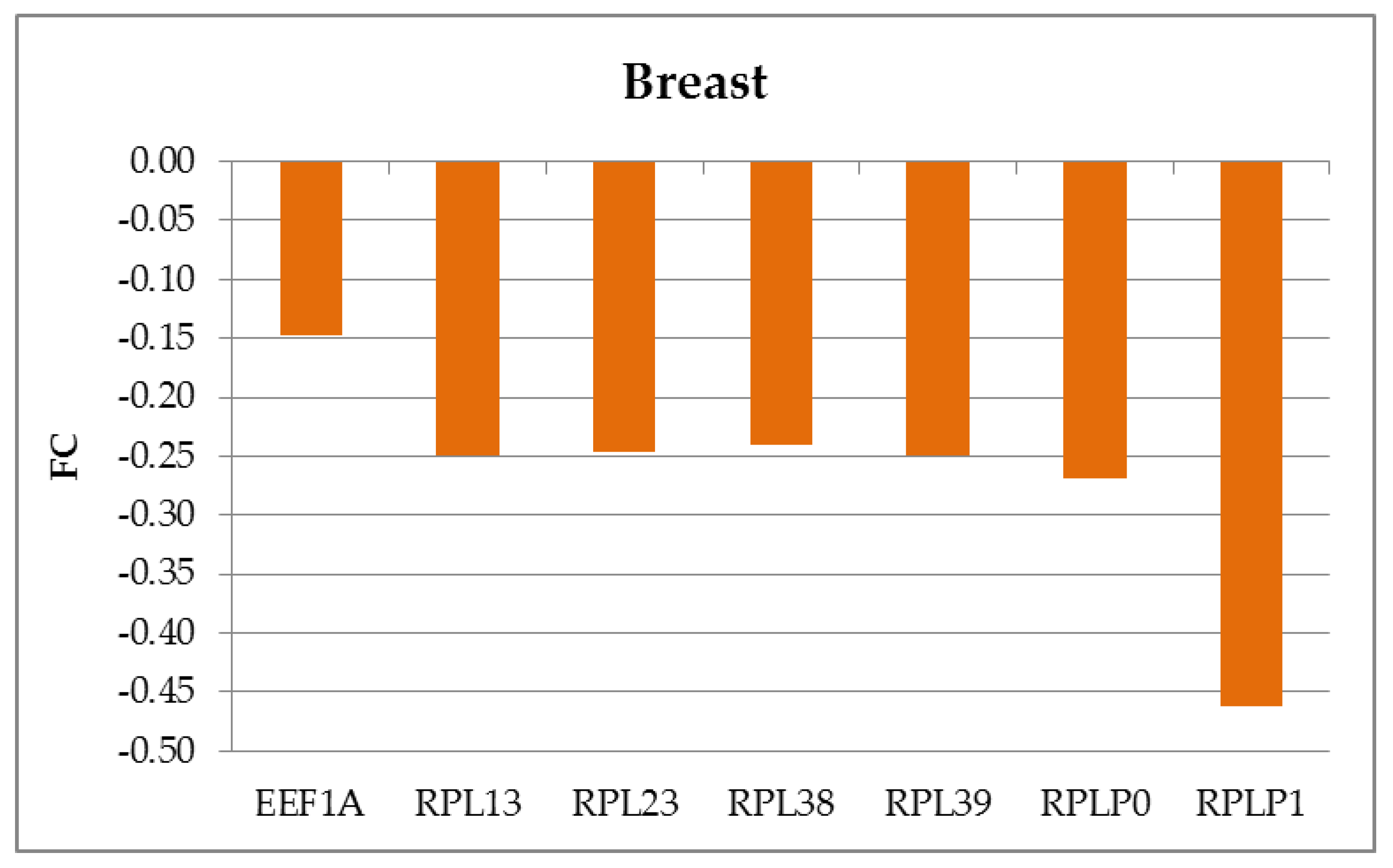
2.3. RNA Sequencing and Data Analysis
2.4. Data Analysis
2.5. Functional Analysis
3. Results
3.1. RNA-Seq
3.2. DEGs Analysis
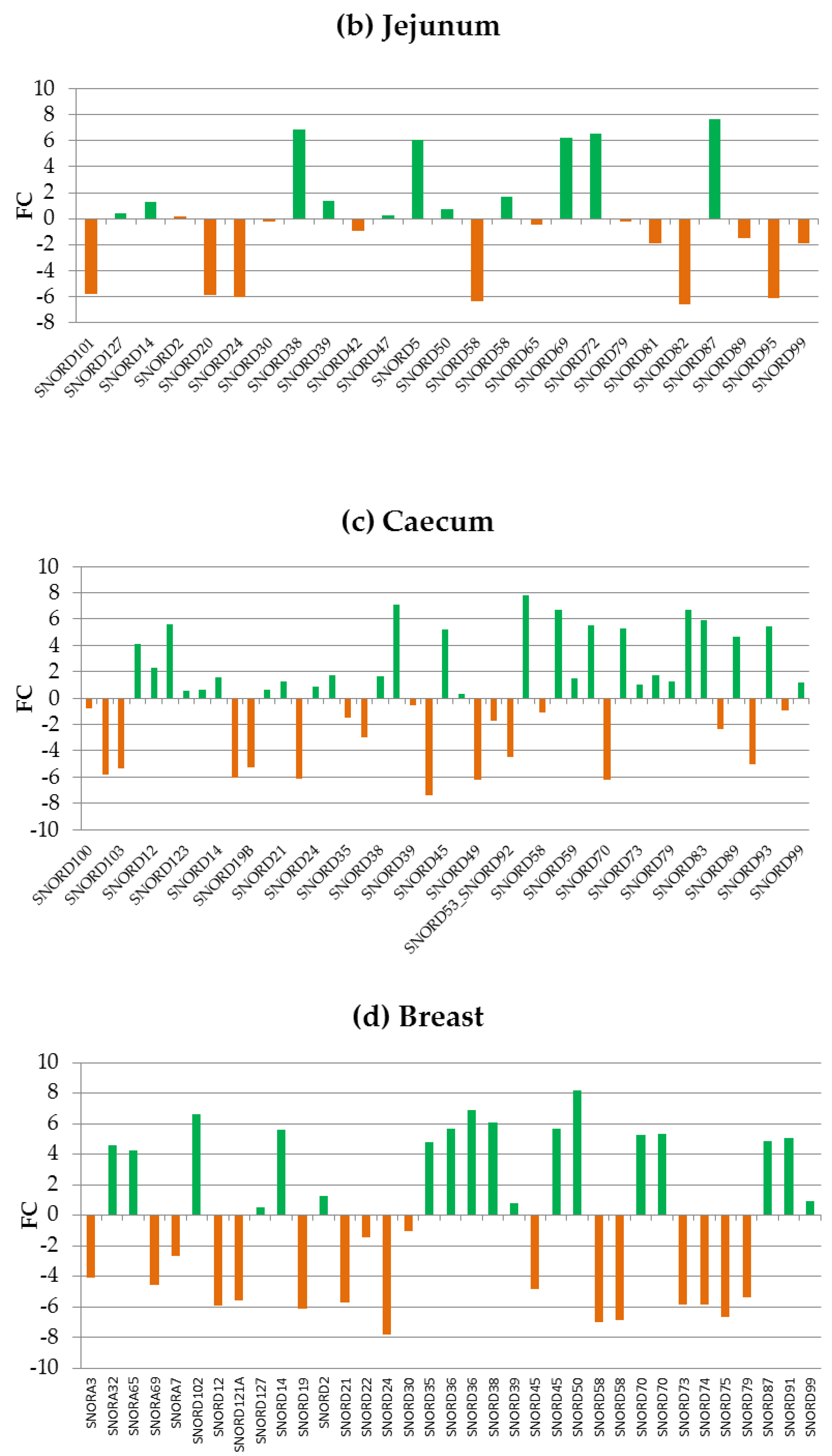
3.2.1. Caecum DEGs
3.2.2. Jejunum DEGs
3.2.3. Breast DEGs
3.2.4. Liver DEGs
3.3. Functional Analysis of DEGs
3.3.1. Expression Pathway Analysis in Breast
3.3.2. Expression Pathway Analysis in Gut and Liver
3.3.3. Protein Metabolism Expression Pathway Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Errico, S.; Spagnoletta, A.; Verardi, A.; Moliterni, S.; Dimatteo, S.; Sangiorgio, P. Tenebrio molitor as a source of interesting natural compounds, their recovery processes, biological effects, and safety aspects. Compr. Rev. Food Sci. Food Saf. 2022, 21, 148–197. [Google Scholar] [CrossRef] [PubMed]
- van Huis, A. Prospects of insects as food and feed. Org. Agric. 2021, 11, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Sogari, G.; Amato, M.; Biasato, I.; Chiesa, S.; Gasco, L. The potential role of insects as feed: A multi-perspective review. Animals 2019, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Biasato, I.; Ferrocino, I.; Biasibetti, E.; Grego, E.; Dabbou, S.; Sereno, A.; Gai, F.; Gasco, L.; Schiavone, A.; Cocolin, L.; et al. Modulation of intestinal microbiota, morphology and mucin composition by dietary insect meal inclusion in free-range chickens. BMC Vet. Res. 2018, 14, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marco, M.; Martínez, S.; Hernandez, F.; Madrid, J.; Gai, F.; Rotolo, L.; Belforti, M.; Bergero, D.; Katz, H.; Dabbou, S.; et al. Nutritional value of two insect larval meals (Tenebrio molitor and Hermetia illucens) for broiler chickens: Apparent nutrient digestibility, apparent ileal amino acid digestibility and apparent metabolizable energy. Anim. Feed. Sci. Technol. 2015, 209, 211–218. [Google Scholar] [CrossRef]
- Biasato, I.; De Marco, M.; Rotolo, L.; Renna, M.; Lussiana, C.; Dabbou, S.; Capucchio, M.; Biasibetti, E.; Costa, P.; Gai, F.; et al. Effects of dietary Tenebrio molitor meal inclusion in free-range chickens. J. Anim. Physiol. Anim. Nutr. 2016, 100, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Oddon, S.B.; Biasato, I.; Imarisio, A.; Pipan, M.; Dekleva, D.; Colombino, E.; Capucchio, M.T.; Meneguz, M.; Bergagna, S.; Barbero, R.; et al. Black Soldier Fly and Yellow Mealworm live larvae for broiler chickens: Effects on bird performance and health status. J. Anim. Physiol. Anim. Nutr. 2021, 105, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Colombino, E.; Biasato, I.; Ferrocino, I.; Bellezza Oddon, S.; Caimi, C.; Gariglio, M.; Dabbou, S.; Caramori, M.; Battisti, E.; Zanet, S.; et al. Effect of insect live larvae as environmental enrichment on poultry gut health: Gut mucin composition, microbiota and local immune response evaluation. Animals 2021, 11, 2819. [Google Scholar] [CrossRef]
- Gariglio, M.; Dabbou, S.; Crispo, M.; Biasato, I.; Gai, F.; Gasco, L.; Piacente, F.; Odetti, P.; Bergagna, S.; Plachà, I.; et al. Effects of the dietary inclusion of partially defatted black soldier fly (Hermetia illucens) meal on the blood chemistry and tissue (Spleen, Liver, Thymus, and Bursa of Fabricius) histology of muscovy ducks (Cairina moschata domestica). Animals 2019, 9, 307. [Google Scholar] [CrossRef] [Green Version]
- Dabbou, S.; Ferrocino, I.; Gasco, L.; Schiavone, A.; Trocino, A.; Xiccato, G.; Barroeta, A.C.; Maione, S.; Soglia, D.; Biasato, I.; et al. Antimicrobial effects of black soldier fly and yellow mealworm fats and their impact on gut microbiota of growing rabbits. Animals 2020, 10, 1292. [Google Scholar] [CrossRef] [PubMed]
- Biasato, I.; Ferrocino, I.; Grego, E.; Dabbou, S.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Gut microbiota and mucin composition in female broiler chickens fed diets including yellow mealworm (Tenebrio molitor, L.). Animals 2019, 9, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biasato, I.; Gasco, L.; De Marco, M.; Renna, M.; Rotolo, L.; Dabbou, S.; Capucchio, M.T.; Biasibetti, E.; Tarantola, M.; Sterpone, L.; et al. Yellow mealworm larvae (Tenebrio molitor) inclusion in diets for male broiler chickens: Effects on growth performance, gut morphology, and histological findings. Poult. Sci. 2018, 97, 540–548. [Google Scholar] [CrossRef]
- Donohue, M.; Cunningham, D. Effects of grain and oilseed prices on the costs of US poultry production. J. Appl. Poult. Res. 2009, 18, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Biasato, I.; Ferrocino, I.; Grego, E.; Dabbou, S.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Yellow mealworm inclusion in diets for heavy-size broiler chickens: Implications for intestinal microbiota and mucin dynamics. Animals 2020, 10, 1909. [Google Scholar] [CrossRef] [PubMed]
- Biasato, I.; Gasco, L.; De Marco, M.; Renna, M.; Rotolo, L.; Dabbou, S.; Capucchio, M.; Biasibetti, E.; Tarantola, M.; Bianchi, C.; et al. Effects of yellow mealworm larvae (Tenebrio molitor) inclusion in diets for female broiler chickens: Implications for animal health and gut histology. Anim. Feed. Sci. Technol. 2017, 234, 253–263. [Google Scholar] [CrossRef]
- Gasco, L.; Biasato, I.; Dabbou, S.; Schiavone, A.; Gai, F. Animals fed insect-based diets: State-of-the-art on digestibility, performance and product quality. Animals 2019, 9, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alaux, C.; Dantec, C.; Parrinello, H.; Le Conte, Y. Nutrigenomics in honey bees: Digital gene expression analysis of pollen’s nutritive effects on healthy and varroa-parasitized bees. BMC Genom. 2011, 12, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Sun, J.; Chen, J.; Wang, L.; Li, A.; Helm, M.; Dubovsky, S.L.; Bacanu, S.-A.; Zhao, Z.; Chen, X. RNA-Seq analysis implicates dysregulation of the immune system in schizophrenia. BMC Genom. 2012, 13, S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W. Potential roles of noncoding RNAs in environmental epigenetic transgenerational inheritance. Mol. Cell. Endocrinol. 2014, 398, 24–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frías-Lasserre, D.; Villagra, C.A. The importance of ncRNAs as epigenetic mechanisms in phenotypic variation and organic evolution. Front. Microbiol. 2017, 8, 2483. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.O.; Jang, H.-J.; Rengaraj, D.; Yang, S.-Y.; Han, J.Y.; Lamont, S.J.; Womack, J.E. Tissue expression and antibacterial activity of host defense peptides in chicken. BMC Vet. Res. 2016, 12, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milona, P.; Townes, C.L.; Bevan, R.M.; Hall, J. The chicken host peptides, gallinacins 4, 7, and 9 have antimicrobial activity against Salmonella serovars. Biochem. Biophys. Res. Commun. 2007, 356, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Cuperus, T.; Coorens, M.; van Dijk, A.; Haagsman, H.P. Avian host defense peptides. Dev. Comp. Immunol. 2013, 41, 352–369. [Google Scholar] [CrossRef] [PubMed]
- Trakaki, A.; Marsche, G. High-Density Lipoprotein (HDL) in allergy and skin diseases: Focus on immunomodulating functions. Biomedicines 2020, 8, 558. [Google Scholar] [CrossRef] [PubMed]
- Rouillard, A.D.; Gundersen, G.W.; Fernandez, N.F.; Wang, Z.; Monteiro, C.D.; McDermott, M.G.; Ma’ayan, A. The harmonizome: A collection of processed datasets gathered to serve and mine knowledge about genes and proteins. Database 2016, 2016, baw100. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.K.; Bang, M.-L.; Witt, C.C.; Labeit, D.; Trombitas, C.; Watanabe, K.; Granzier, H.; McElhinny, A.S.; Gregorio, C.C.; Labeit, S. The muscle Ankyrin Repeat Proteins: CARP, ankrd2/Arpp and DARP as a family of Titin filament-based stress response molecules. J. Mol. Biol. 2003, 333, 951–964. [Google Scholar] [CrossRef]
- Stamenkovic, N.; Jasnic, J.; Novkovic, M.; Milosevic, E.; Boskovic, S.; Kojic, A.; Popic, K.; Stankovic, M.; Wang, Y.; Milenkovic, S.; et al. Cloning and expression profiling of muscle regulator ANKRD2 in domestic chicken Gallus gallus. Histochem. Cell Biol. 2020, 154, 383–396. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, B.; Li, W.; Oiso, N.; Novak, E.K.; Rusiniak, M.E.; Gautam, R.; Chintala, S.; O’Brien, E.P.; Zhang, Y.; et al. Ru2 and Ru encode mouse orthologs of the genes mutated in human Hermansky-Pudlak syndrome types 5 and 6. Nat. Genet. 2003, 33, 145–153. [Google Scholar] [CrossRef]
- Helip-Wooley, A.; Westbroek, W.; Dorward, H.M.; Koshoffer, A.; Huizing, M.; Boissy, R.E.; Gahl, W.A. Improper trafficking of melanocyte-specific proteins in Hermansky–Pudlak syndrome type-5. J. Investig. Dermatol. 2007, 127, 1471–1478. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Pan, Z.; Zhang, K.; Yu, M.; Yu, D.; Lu, Y.; Wang, J.; Zhang, J.; Zhang, K.; Du, W. Identification of the differentially expressed genes of muscle growth and intramuscular fat metabolism in the development stage of yellow broilers. Genes 2020, 11, 244. [Google Scholar] [CrossRef] [Green Version]
- Barnes, D.J.; Hookway, E.; Athanasou, N.; Kashima, T.; Oppermann, U.; Hughes, S.; Swan, D.; Lueerssen, D.; Anson, J.; Hassan, A.B. A germline mutation of CDKN2A and a novel RPLP1-C19MC fusion detected in a rare melanotic neuroectodermal tumor of infancy: A case report. BMC Cancer 2016, 16, 629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proud, C.G. Peptide-chain elongation in eukaryotes. Mol. Biol. Rep. 1994, 19, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.F.; Guo, Y.M.; Huang, B.Y.; Bun, S.; Zhang, L.B.; Li, J.H.; Liu, D.; Long, F.Y.; Yang, X.; Jiao, P. The effect of glucagon-like peptide 2 injection on performance, small intestinal morphology, and nutrient transporter expression of stressed broiler chickens. Poult. Sci. 2010, 89, 1967–1974. [Google Scholar] [CrossRef] [PubMed]
- Hutsko, S.; Meizlisch, K.; Wick, M.; Lilburn, M. Early intestinal development and mucin transcription in the young poult with probiotic and mannan oligosaccharide prebiotic supplementation. Poult. Sci. 2016, 95, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, E.; Li, H.; Emmerson, D.; Webb, K., Jr.; Wong, E. Dietary protein composition influences abundance of peptide and amino acid transporter messenger ribonucleic acid in the small intestine of 2 lines of broiler chicks. Poult. Sci. 2010, 89, 1663–1676. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, D.P.; Keene, D.R.; Corson, G.M.; Pöschl, E.; Bächinger, H.P.; Gambee, J.E.; Sakai, L.Y. Fibrillin-1: Organization in microfibrils and structural properties. J. Mol. Biol. 1996, 258, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Tang, X.; Meng, Q.; Zhang, H. Differential expression analysis of the broiler tracheal proteins responsible for the immune response and muscle contraction induced by high concentration of ammonia using iTRAQ-coupled 2D LC-MS/MS. Sci. China Life Sci. 2016, 59, 1166–1176. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Lin, Y.; Wu, H.; Zhang, M. Selenium accumulation in protein fractions of Tenebrio molitor larvae and the antioxidant and immunoregulatory activity of protein hydrolysates. Food Chem. 2021, 334, 127475. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.L.; Simonović, M. Synthesis and decoding of selenocysteine and human health. Croat. Med. J. 2012, 53, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| First Period (Day 1 to 12) | Second Period (Day 12 to 25) | Third Period (Day 25 to 53) | ||||
|---|---|---|---|---|---|---|
| Ingredients (g/kg as Fed) | CT | TM | CT | TM | CT | TM |
| Corn meal | 483.2 | 496.6 | 523.8 | 566.8 | 566.6 | 605.4 |
| Soybean meal | 345.0 | 262.0 | 317.0 | 203.9 | 275.5 | 164.0 |
| TM larvae meal | - | 150.0 | - | 150.0 | - | 150.0 |
| Corn gluten meal | 75.5 | 14.5 | 58.3 | - | 56.0 | - |
| Soybean oil | 54.0 | 34.8 | 64.9 | 41.6 | 68.9 | 45.8 |
| Dicalcium phosphate | 11.0 | 15.5 | 8.4 | 12.9 | 7.0 | 12.0 |
| Calcium carbonate | 17.5 | 15.0 | 15.0 | 13.0 | 14.5 | 12.2 |
| Sodium chloride | 2.3 | 2.3 | 2.3 | 2.3 | 2.3 | 2.3 |
| Sodium bicarbonae | 1.3 | 1.3 | 1.3 | 1.3 | 1.3 | 1.3 |
| DL-methionine | 0.8 | 0.8 | 0.8 | 0.8 | 0.4 | 0.5 |
| L-lysine | 3.1 | 0.9 | 2.0 | 0.8 | 1.3 | - |
| Threonine | 0.1 | 0.1 | - | 0.4 | - | 0.3 |
| Trace mineral-vitamin premix 2 | 5.0 | 5.0 | 5.0 | 5.0 | 5.0 | 5.0 |
| Choline chloride | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| 3-phytase | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Total (g/kg as fed) | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 |
| AMEn 3 (MJ/kg DM) | 12.89 | 12.89 | 13.28 | 13.28 | 13.54 | 13.54 |
| Nutrient composition (%) | ||||||
| DM | 86.6 | 86.6 | 86.7 | 86.8 | 86.8 | 86.8 |
| CP | 23.5 | 23.8 | 21.3 | 21.1 | 19.6 | 19.6 |
| EE | 7.9 | 9.6 | 9.0 | 10.3 | 9.5 | 10.8 |
| NDF | 9.4 | 10.1 | 9.4 | 10.1 | 9.4 | 10.1 |
| ADF | 3.8 | 4.4 | 3.7 | 4.1 | 3.5 | 4.0 |
| Transcripts | Breast | Liver | Jejunum | Caecum |
|---|---|---|---|---|
| with FPKM 1 > 5 | 6825 | 7294 | 9953 | 9389 |
| with ΔFPKM 1 > |5| | 1847 | 1989 | 2328 | 2292 |
| DEGs 2 | 237 | 234 | 288 | 308 |
| Upregulated DEGs 2 FC 3 > 2.5 | 117 55 | 118 33 | 119 25 | 182 39 |
| Downregulated DEGs 2 FC 3 < −2.5 | 120 40 | 116 41 | 168 39 | 126 52 |
| Annotated DEGs 2 | 181 | 184 | 223 | 252 |
| GO 4 gene | 91 | 85 | 116 | 117 |
| n° pathways | 4082 | 4072 | 4668 | 4602 |
| Category | Breast | Liver | Jejunum | Caecum |
|---|---|---|---|---|
| DEGs 1 | 237 | 234 | 289 | 308 |
| MiRNA 2 | 42 | 41 | 42 | 54 |
| SNOR 3 | 35 | 40 | 35 | 58 |
| MT 4 | 29 | 14 | 18 | 10 |
| No ID 5 | 56 | 50 | 65 | 56 |
| Coding Gene | 77 | 89 | 129 | 130 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soglia, D.; Viola, I.; Nery, J.; Maione, S.; Sartore, S.; Lasagna, E.; Perini, F.; Gariglio, M.; Bongiorno, V.; Moretti, R.; et al. Nutrigenomics in Animal Feeding: Digital Gene Expression Analysis in Poultry Fed Tenebrio molitor Larvae Meal. Poultry 2022, 1, 14-29. https://doi.org/10.3390/poultry1010003
Soglia D, Viola I, Nery J, Maione S, Sartore S, Lasagna E, Perini F, Gariglio M, Bongiorno V, Moretti R, et al. Nutrigenomics in Animal Feeding: Digital Gene Expression Analysis in Poultry Fed Tenebrio molitor Larvae Meal. Poultry. 2022; 1(1):14-29. https://doi.org/10.3390/poultry1010003
Chicago/Turabian StyleSoglia, Dominga, Irene Viola, Joana Nery, Sandra Maione, Stefano Sartore, Emiliano Lasagna, Francesco Perini, Marta Gariglio, Valentina Bongiorno, Riccardo Moretti, and et al. 2022. "Nutrigenomics in Animal Feeding: Digital Gene Expression Analysis in Poultry Fed Tenebrio molitor Larvae Meal" Poultry 1, no. 1: 14-29. https://doi.org/10.3390/poultry1010003