
Environmental Regulation of Weed Seed Dormancy and Germination †


1
Department of Biology, Mount Saint Vincent University, 166 Bedford Highway, Halifax, NS B3M 2J6, Canada
2
Department of Biology, Saint Mary’s University, 923 Robie Street, Halifax, NS B3H 3C3, Canada
3
Department of Biology, Life Science Centre, Dalhousie University, 1355 Oxford Street, Halifax, NS B3H 4R2, Canada
†
This paper is dedicated to the memory of the late Dr. Paul Brethen Cavers (18 January 1938–18 April 2023), in recognition of his valuable contributions to the Ecology of Weeds and Seeds.
Seeds 2023, 2(3), 259-277; https://doi.org/10.3390/seeds2030020
Submission received: 15 January 2023
/
Revised: 27 June 2023
/
Accepted: 27 June 2023
/
Published: 30 June 2023
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Many weeds produce dormant seeds that are unable to complete germination under favourable conditions. There are two types of seed dormancy: primary dormancy (innate dormancy), in which seeds are in a dormant state upon release from the parent plant, and secondary dormancy (induced dormancy), in which dormancy develops in seeds through some experience after release from the parent plant. Mechanisms of seed dormancy are categorized as embryo dormancy and coat-imposed dormancy. In embryo dormancy, the control of dormancy resides within the embryo itself, and in coat-imposed dormancy, it is maintained by the structures enclosing the embryo. Many factors can influence seed dormancy during development and after dispersal; they can be abiotic, biotic, or a combination of both. Most weeds deposit a large number of seeds in the seed bank, which can be one of two types—transient or persistent. In the transient type, all viable seeds in the soil germinate or die within one year, and there is no carry-over until a new crop is deposited. In the persistent type, at least some seeds survive in the soil for more than one year and there is always some carry-over until a new crop is deposited. Some dormant seeds require after-ripening—changes in dry seeds that cause or improve germination. Nondormant, viable seeds can germinate if they encounter appropriate conditions. In the face of climate change, including global warming, some weeds produce a large proportion of nondormant seeds, which germinate shortly after dispersal, and a smaller, more transient seed bank. Further studies are required to explore this phenomenon.
1. Introduction
Environmental factors influence the parent plant during growth and its progeny during development, after dispersal, and subsequent germination [1,2,3,4]. Pre-dispersal factors, including temperature, moisture, light, nutrients, and gases, affect the metabolic activities of the parent plant and, in turn, influence seeds [5,6,7]. During maturation, morphological, physiological, biochemical, and molecular aspects of seeds are affected by environmental factors [4], and changes are portrayed by the seed size, shape [3,8], and germinability [1,3,9]. The position of seeds on the parent plant can also affect their colour, morphology, and germinability [2,10]. Environmental conditions affect plant reproduction and are critical factors in controlling seed dormancy and germination [11,12]. Among plants, many weeds produce seeds with different degrees of dormancy to ensure the perpetuation of future generations [13]. After seed dispersal, many weeds produce large seed banks and exhibit intermittent germination [14]. Such weeds are more successful in perpetuating their generation than those that have small seed banks [15]. Both climatic and edaphic factors can influence seeds in the seed bank by either promoting or inhibiting their germinability, which depends on the plant species and the interactive effects of environmental factors [15,16,17,18,19,20,21]. Many weeds produce seeds that require a period of after-ripening before they can germinate [7,15,22].
As the climate is changing, the response of plants, especially weeds, and their progeny will also change in response to climatic factors [6,23]. Seeds are at the most critical stage in the life of a plant and are important for the existence of new generations; thus, changes in their behaviour, especially germinability, determine the survival of a particular plant species [24]. By controlling the time of germination, seed dormancy strongly influences plant fitness and survival [25,26]. Weeds that produce seeds with various degrees of dormancy will be more successful than those that produce more germinable seeds because, in the latter case, an unexpected disaster can wipe out the whole population [14,27]. In terms of temporal and spatial distribution, seed germination is critical for the survival of seed plants. Seed morphological characteristics control the spatial distribution of germination, whereas seed physiological characteristics control the temporal distribution of germination [28]. In this review, I focus on seed dormancy and germination, seed banks, and the pre- and post-dispersal factors that affect weed seed dormancy with reference to Scotch thistle (Onopordum acanthium L.) as a model plant for such studies.
2. Seed Dormancy: A Survival Strategy
Dormancy regulates the life of most weed species by determining their seed germinability. The production of seeds with different degrees of dormancy is a survival strategy of many weeds, which is of great concern to agriculturists since these seeds can germinate intermittently over a long period. Therefore, without a comprehensive understanding of dormancy, problems created by weeds will exist forever [29]. “Seed dormancy is regarded as the failure of an intact viable seed to complete germination under favourable conditions” [30]. It is a population-based phenomenon since every seed independently senses its environment and responds physiologically to the perceived signal [31]. Seed dormancy is determined by both genetic and environmental factors [12,32], such as temperature, light, and soil nitrate, which are experienced by the mother plant during seed maturation [7]. Seed dormancy can be caused by developmental, physiological, chemical, and structural factors [33], and has been described in both physiological and ecological terms [15,22,34,35,36,37].
Different types of seed dormancy have been recognized [15]. For example, Marianna Nikolaeva (1982) classified seed dormancy into two general types: endogenous and exogenous. Endogenous dormancy includes physiological, morphological, and morpho-physiological, in which some characteristics of the embryo prevent germination. However, exogenous dormancy includes physical, chemical, and mechanical, in which some characteristics of the embryo covering prevent germination [35].
John Harper (1957) wrote, “some seeds are born dormant, some achieve dormancy, and some have dormancy thrust upon ’em”. He described these as “innate”, “induced”, and “enforced” dormancy, respectively. Innate dormancy occurs when seeds are in a dormant state upon release from the parent plant, whereas induced dormancy is used to describe a situation in which dormancy develops through some experience after release from the parent plant. Enforced dormancy is the inability of a seed to germinate due to an environmental restraint, such as a shortage of water, low temperatures, or poor aeration [38]. Even though this classification has been accepted by some authors [10,39], for others, it has been a topic of debate [15,37].
Vleeshouwers et al. (1995) opposed the concept of dormancy introduced by Harper (1957) and stated that “seed dormancy should not be identified with the absence of germination” and that the phenomenon, which was classified as “seeds’ dormancy”, should be called “seeds’ inability to germinate”. They proposed the following definition for seed dormancy: “dormancy is a seed characteristic, the degree of which defines what conditions should be met to make the seed germinate” [37].
Baskin and Baskin (2014) also discussed Harper’s terms, “induced dormancy” and “enforced dormancy”, and stated that they were unaware of any species whose seeds are nondormant at maturity but are induced into dormancy. They declared that it is inappropriate to use the term “induced dormancy” for the seeds that are dormant or conditionally dormant at maturity and later become nondormant and reenter secondary dormancy. According to them, the term “induced dormancy” overlaps with the term “secondary dormancy”. Moreover, they regarded “enforced dormancy” as equal to “quiescence” and insisted that if the term “enforced dormancy” is used, the lack of germination must be entirely due to an unfavourable environmental condition [15].
Even though Baskin and Baskin (2014) stated that there are no species that produce nondormant seeds that can be induced into dormancy [15], it appears that bull thistle (Cirsium vulgare) can exhibit this phenomenon. Fresh cypselae (indehiscent dry fruit) of C. vulgare have virtually no innate dormancy and germinate in either light/dark or constant-dark conditions. However, after a period of stratification in constant darkness, most of the cypselae require light for germination [40].
If freshly matured seeds are unable to germinate under favourable conditions, they have primary dormancy (innate dormancy). After dispersal, seeds with physiological dormancy experience the transitional period of conditional dormancy before they become fully nondormant. At first, seeds gain the ability to germinate under a narrow range of environmental conditions, which increases with the loss of dormancy until they become completely nondormant and germinate under a wide range of conditions. If nondormant seeds are unable to germinate, because of changes in the environment, they reenter conditional dormancy but can germinate under a full range of conditions. As time passes, the range of conditions within which germination is possible narrows to the extent that germination is not possible under any condition and the seed acquires secondary dormancy. Some seeds exhibit an annual cycle of dormancy/non-dormancy, while others exhibit a cycle of conditional dormancy/non-dormancy. Some others exhibit neither conditional nor primary dormancy and can germinate throughout the year [15].
Dormancy plays a critical role in weed population dynamics. Therefore, fundamental knowledge about the mechanisms that regulate seed dormancy is necessary to improve weed management practices [29]. Different mechanisms can be involved in various types of seed dormancy in different species. These mechanisms may be grouped into two broad categories: (1) embryo dormancy, where the control of dormancy resides within the embryo itself, and/or (2) coat-imposed dormancy, in which dormancy is maintained by the structures enclosing the embryo, such as the seed coat [22]. Embryo dormancy is a very complex mechanism, which has not been explained thoroughly. Two factors can be involved: (1) cotyledons and (2) germination inhibitors [22]. In embryo dormancy, the following factors are known as control mechanisms: (1) an undifferentiated and underdeveloped embryo, (2) blocks to the synthesis of nucleic acids and proteins, (3) failure of certain enzymes to mobilize food reserves for the embryo, (4) a deficiency of plant growth hormones, and (5) the presence of inhibitors in the embryo [41]. Coat-imposed dormancy is the exertion of a constraint by the tissues enclosing the embryo that the embryo cannot overcome. Structures responsible for imposing dormancy include endosperm, pericarp, or extrafloral organs. The tissues responsible for embryo restraint differ from species to species [22]. In coat-imposed dormancy, the following factors act as control mechanisms: (1) restriction in water uptake, (2) restriction in gas exchange, (3) prevention of the release of inhibitors from the embryo, (4) provision of inhibitors to the embryo, (5) filtration of incident light, and (6) physical prevention of radicle protrusion [17,41]. Either embryo or coat-imposed dormancy or a combination of the two can be involved in dormancy mechanisms in different species [22]. After the termination of dormancy by different environmental factors, germination can be initiated if seeds meet suitable conditions.
A consensus view based on those presented above is that dormancy is an internal characteristic of a seed that prevents germination under favourable conditions of water, temperature, and gases. Therefore, the term “dormancy” includes “innate and induced dormancy”, whereas the term “quiescence” refers to so-called “enforced dormancy”. A comprehensive classification of seed dormancy has been provided by Baskin and Baskin [42,43].
3. Seed Germination: The Initiation of a New Generation
Nondormant, viable seeds, fresh or stored, can germinate if they encounter appropriate environmental conditions [5]. Germination includes all the events from the uptake of water by the quiescent dry seed to the elongation of the embryonic axis [22]. In the physiological sense, this starts with the initiation of embryo growth and terminates with the protrusion of the radicle through the covering tissue [31]. Penetration of the radicle through the covering structures is called visible germination [30]. According to Salisbury (1961), there are two main types of germination patterns: quasi-simultaneous and intermittent. If the vast majority of a batch of seeds germinate within a few days when there is sufficient moisture in the soil, that is a quasi-simultaneous pattern. However, if seeds from a single sample germinate/emerge at irregular intervals over weeks, months, or years, that is an intermittent pattern [27]. Most weeds produce seeds with different degrees of dormancy and have an intermittent pattern of germination. This pattern is important for short-lived monocarpic weeds because new seedling populations quickly replace those that have been eradicated. The time at which germination takes place is critical to the survival of seedlings since it determines the availability of resources for subsequent seedling growth [44,45]. The timing of germination can be controlled by seed response to several environmental factors, including moisture, temperature, light, gases, soil nutrients, and other soil chemicals [5,6,10,18,46,47].
From the above factors, only moisture, temperature, and, in most cases, oxygen, are universal prerequisites for the growth of an embryo [48]. However, light can promote or inhibit seed germination, which is commonly dependent upon temperature [49]. In positively photoblastic seeds, germination is promoted by white light, whereas in negatively phoblastic seeds, germination is inhibited [17]. It has been shown that chemicals from plant-generated smoke (e.g., karrikins) can affect the germination of photoblastically sensitive seeds [50]. Seeds of many weeds require exposure to light for germination. This is regulated by a bluish-green pigment-protein complex called phytochrome. Phytochrome occurs in two photo-interconvertible forms: the red-absorbing form (Pr) and the far-red-absorbing form (Pfr). The absorption maxima of Pr and Pfr are approximately 660 and 730 nm, respectively, but there can be considerable overlap between the two forms [48]. The Pfr form is biologically active, whereas Pr is inactive. Since Pr absorbs red light most efficiently, this wavelength will drive a high proportion of the molecule into the Pfr form, thereby inducing germination. Subsequent far-red light absorbed by the Pfr form will promote the formation of Pr, thereby cancelling the effect of the prior red light [51]. As shown, germination is more similar in dark than in light, when seeds are germinated at low temperatures. This is because the transformation of Pfr into Pr (in dark and in low temperatures) is very slow, and 100% transformation does not exist; the seeds show high sensitivity to low proportions (ca. 4%) of the remaining active form [52,53].
Seed germination is closely related to seed anatomy. In several species, the role of seed coats as dormancy regulators has been demonstrated [54,55,56]. Seeds with less primary dormancy will germinate under a wide range of environmental conditions, while those with stronger primary dormancy will only germinate within a very restricted range. Every seed has its own “germination window”, which can change with time. The germination window is defined as the range of conditions under which a seed will germinate [57]. Seeds are more germinable if the range of germination conditions is wider. Dormancy-relieving factors (e.g., temperature) widen the window (range), while dormancy-inducing factors narrow this range [58,59,60].
Three conditions are required for a seed to germinate: (1) viability, (2) non-dormancy, and (3) favourable conditions. The availability of these requirements may cause the initiation of germination, which will take place in five consecutive stages: (1) water uptake by the seed, (2) activation of hormones, enzymes, and cellular respiration, (3) breakdown and movement of stored reserves to the embryo, (4) use of the mobilized reserves by the embryo as a source of energy for cellular activity and growth, and (5) cell division and embryo elongation that enable the radicle to protrude from the seed coat [61]. These stages, which have been studied extensively in cereals, especially barley (Hordeum vulgare L.), are discussed briefly here. Caryopses of barley and other cereals have a special layer of endosperm cells, called the aleurone layer, that surrounds the centrally located starchy endosperm. This layer is cytologically and biochemically distinct from the starchy endosperm. Aleurone cells contain protein-storing organelles called protein bodies as well as lipid-storing oleosomes. Shortly after water uptake, when the seeds begin to germinate, the embryo releases gibberellins (GA) that activate certain genes, causing the synthesis of specific messenger RNA (mRNA) molecules, which, in turn, direct the synthesis of the hydrolytic enzymes in the aleurone layer. Abscisic acid (ABA) inhibits this GA-dependent enzyme synthesis by inhibiting the transcription of α-amylase mRNA. Both the scutellum and the aleurone layer secrete enzymes into the starchy endosperm. These enzymes catalyze the breakdown of starch and other macromolecules during germination. The two enzymes responsible for starch degradation are α- and β-amylase. α-amylase hydrolyzes starch chains to produce oligosaccharides consisting of α-1,4-linked glucose residues. β-amylase degrades these oligosaccharides to produce the disaccharide maltose. Maltase then converts maltose to glucose. The hydrolysis products are released in the form of solubilized sugars, amino acids, and other products, which are absorbed by the scutellum and then transported to the growing regions of the embryo [62].
Studies with tomato seeds have also shown that GA induced the enzymatic hydrolysis of the galactomannan-rich endosperm cell walls, and the activity of enzyme endo-β-mannanase was controlled completely by GA. Even though gibberellins are important in the promotion and maintenance of germination, they are not involved in the control of dormancy. There is much evidence that ABA is involved in regulating the onset and maintenance of dormancy [22,30,63]. The effects of ABA on seed dormancy and those of GA on seed germination have been known for decades. There is a “cross-talk” between these two hormones; one regulates the metabolism and signal transduction of the other. The inhibitory and promotive actions of ABA and GA, respectively, are the primary determinants of seed dormancy and germination. The relative activities of these hormones are related to their amounts as well as the sensitivity of the target cells/tissues to them; these depend on their respective perception and signalling pathways [22]. Environmental factors strongly influence the metabolism, sensitivity, and signalling of ABA and GA [64]. Ethylene and brassinosteroids can influence the signalling pathway of ABA by reducing its ability to inhibit germination by acting as its downstream synthesis in the signal transduction pathway [22]. It has also been shown that auxin plays a critical role in the induction and maintenance of seed dormancy, and its action requires the ABA signalling pathway, indicating the interdependent roles of these hormones in seed dormancy [65,66].
Some seeds may germinate shortly after dispersal, while others may remain dormant in the soil seed bank for several months to many years until suitable environmental conditions are available [10]. After dispersal, the primary seed dormancy can be broken by environmental signals, such as seasonal changes in temperature, soil nitrate levels, and smoke from forest fires [67].
4. The Seed Bank: A Rewarding Investment
All viable seeds present on the soil surface or buried in the soil make up the soil seed bank. As a bank, both deposits and withdrawals of seeds take place. Deposits are made through seed rain during seed dispersal, while withdrawals are made via germination, senescence, death, and predation [34]. Most weeds and invasive species deposit large numbers of seeds in the seed bank. Seeds are distributed vertically in the soil, with most of them buried near the surface [68]. Weed seedling emergence has been reported from a range of soil burial depths (0 to 9 cm) [69]. In general, there are two types of seed banks: (1) transient and (2) persistent. In the transient type, which is usually from grasses and forbs, all viable seeds in the soil germinate or die within one year, and there is no carry-over until a new crop is deposited. Species with a transient seed bank during the summer may germinate in the autumn, while species with a transient seed bank during the winter may germinate in the following spring. In the persistent type, which is represented by species from a wide range of habitats, at least some seeds survive in the soil for more than one year and there is always some carry-over until a new crop is deposited. There are some species with persistent seed banks in which most seeds germinate quickly, but some survive for at least one year. In contrast, other species with persistent seed banks have only a few seeds germinating soon after dispersal [70]. Dormancy regulates the persistence of weed soil seed banks and the timing of seedling emergence [71]. In the soil seed bank, seeds can sense a range of environmental signals and continually adjust their dormancy status [72]. In the soil, seeds may undergo long-term cycles of dormancy release and induction without completing their germination [57,73,74]. Plant hormones and dormancy proteins can control the release and induction of dormancy, which are influenced by environmental factors during seed maturation and storage [7]. In a study, seeds of wild mustard (Sinapis arvensis L.) were buried in a field. During exposure to high summer temperatures, the seeds became nondormant, but low water potential and constant supra-optimal temperatures induced them into secondary dormancy [75].
Seed longevity is an important aspect of the soil seed bank [10]. In a habitat, seed longevity depends on both environmental factors and seed dormancy/germination traits [76]. In a study with 28 herbaceous species, it was shown that seed longevity increased with increased burial depth, but there were significant differences among species [77]. Several burial studies have shown that seeds of many weed species can survive for long periods in the seed bank [15]. It is strange that many species with only “quiescent” seeds nevertheless survive for very long periods (many decades) in the seed bank. For example, round-leaved mallow (Malva rotundifolia L.), moth mullein (Verbascum blattaria L.), and mullein (Verbascum sp.) were the most long-lived species (120 years) in the longest-running experiment on the longevity of seeds under field conditions, which was initiated by Dr. W. J. Beal in East Lansing, Michigan, USA, in the autumn of 1879 [78].
The interactions of many factors, such as the intrinsic dormancy characteristics of seed populations; environmental conditions, such as water, temperature, light, and gases that affect the dormancy state; and biological interactions, such as predation and allelopathy, are involved in the longevity of seeds in the soil seed bank [19,79]. The condition and location of seeds in the soil profile can determine the intensity and manner of the interaction of these factors. In summary, genotypes; environmental factors during seed development, maturation, and after-ripening; and seed morphology, such as size, shape, and colour, affect a seed’s condition and dormancy state [68,80,81,82].
5. The Effects of Pre-Dispersal Factors on Seed Dormancy: Maturation Conditions
Many factors can influence seed dormancy during development (see Figure 1). Seeds develop from fertilized ovules. The seeds of flowering plants contain three genetically different tissues: (1) the typically diploid embryo, which develops from a zygote through equal contributions by both parents—the maternal contribution is the egg cell and the paternal contribution is the spermatic nucleus; (2) the typically triploid endosperm, formed by the fusion of the two polar nuclei with the second spermatic nucleus, in which maternal and paternal contributions are in the ratio of 2:1; and (3) the seed coat, formed from integuments (embryonic seed coat), in which there is only a maternal contribution [83]. Seed development can be a complex and prolonged process. When the ovule is large, it has the potential to gradually increase its volume, resulting in a large seed. However, when it is small, the developmental process is simple and short, and the volume of the ovule hardly increases, resulting in a small seed, often with a simple seed coat [84].
The process of embryo development, or embryogenesis, takes place after the fertilization of the egg cell with a sperm nucleus [85,86]. In a mature seed, the embryo consists of an embryonic axis with a plumule (shoot), a radicle (root) and one or two cotyledons or, in some cases, several (e.g., conifers), depending on the type of plant [87]. After the final stage of development (desiccation), the mature seed becomes independent of the mother plant and, under the appropriate environmental conditions, it resumes its growth into a new seedling and, ultimately, into a mature plant. In some species, seeds must retain relatively high moisture content (12–31%) to be viable. These seeds, which die if desiccated, are called “recalcitrant”. They are different from those that remain viable in a state of low moisture content and are called “orthodox” [33].
Almost always, the development of the endosperm precedes that of the embryo. The endosperm is a seed storage tissue, formed within the embryo sac from a second fertilization event between two polar nuclei and the second spermatic nucleus [88]. The endosperm can have an important role in the growth of the embryo or seedling. During seed development, the volume ratios of ovule, endosperm, and embryo change depending on the species, resulting in endospermous and exendospermous seeds. An endospermous seed is one in which endosperm is formed and retained as a storage tissue in the mature seed. In small seeds, the growth of the embryo starts much later than that of the endosperm, a small part of the endosperm is consumed during embryogenesis, and the embryo remains relatively small [84]. In this case, a complete breakdown of the endosperm takes place during seed germination. However, when the endosperm is crushed during development or the division of the endosperm terminates early, the resulting seed contains little or no endosperm and is termed exendospermous. There is no clear distinction between endospermous and exendospermous seeds since seeds with transitional endosperm are also known. Moreover, in several plant families, including the Asteraceae, the endosperm of a mature seed is reduced to a single layer, which is not visible macroscopically [84,88,89]. In these families, the endosperm may contribute to the expression of dormancy and germination [22].
The seed coat is derived from the ovule integuments and, therefore, has the genetic characteristics of the mother plant [90]. Different cell types, such as parenchyma, sclerenchyma, and cork, may be part of the seed coat. The sclerenchyma elements, sclereids, or fibres are thick-walled cells that may contain lignin. The epidermis, with its overlying cuticle, constitutes the outside of the seed and shows much variation in structure, cellular arrangement, and sculpture. It has many functions, including the secretion of epicuticular waxes [33,84].
During the last stages of maturation, a drastic reduction occurs in the metabolic activity of seeds, followed by rapid desiccation and compression of the seed coat layers. Seed coat shrinkage can cause cell compression, the closure of all openings in the coat, and the impregnation of the cell walls by water-repellent substances, such as suberin, cutin, lignin, and callose, which promote impermeability in seeds with hard tissues—the so-called “hard seeds” [22,91]. Seed coat impermeability that adversely affects water uptake and gas exchange and prevents germination is called “physical dormancy” [22,90]. This class of dormancy is found in many plant families, including Anacardiaceae, Fabaceae, Cannaceae, Chenopodiaceae, Convolvolaceae, Geraniaceae, Malvaceae, and Sapindaceae [22,92].
In mature seeds, seed coats mainly contain dead and hard tissues, which form an effective covering structure. The seed epidermis together with its overlying cuticle has a variety of functions, including mechanical and chemical protection against environmental factors [33]. Moreover, both embryo and seed coat can be rich in chemical compounds, such as phenolics, alkaloids, amino acids, tannins, and cyanogenic glycosides [84].
The effects of environmental conditions on seed development and subsequent germinability have been studied in many species [2,93,94,95]. Environmental factors, such as temperature, light, drought, nutrients [5], and carbon dioxide [6], can affect seed development and germinability. In particular, the effects of temperature on plant growth and seed development are profound [96]. The range of temperatures over which the seeds of a particular species can germinate is called the cardinal temperatures, and they are as follows: (1) the minimum or base temperature—the lowest temperature at which germination can occur, (2) the maximum or ceiling temperature—the highest temperature at which the seeds can germinate, and (3) the optimum temperature—the temperature at which germination is most rapid [22,97].
In general, high temperatures within the physiological range promote flowering [3,98], increase the rate of dry matter accumulation, and reduce the duration of seed development [3,99]. Plants that are grown in high temperatures produce smaller seeds than those grown in low temperatures [3,94]. A positive correlation between seed germinability and higher temperature during seed development has been found for many species [3,80,100,101,102,103,104]. In evening primrose (Oenothera biennis L.), for instance, maturation drying in higher temperatures resulted in more viable and faster-germinating seeds [4]. It has been postulated that the synthesis of germination inhibitors may be increased at low temperatures, while the synthesis of germination promoters may be increased at high temperatures [5]. This may be true for O. acanthium [3,13].
Carbon dioxide can also influence seed development and its subsequent germinability. A study has shown that elevated carbon dioxide increased single-sound seed mass and germination rate, but decreased germination percentage [6].
Photoperiod and light quality may affect seed development and germinability [14]. In some species, seeds that mature in short days become highly germinable. For example, seeds of lamb’s-quarters (Chenopodium album L.) that matured in 8 h days were more germinable than those that matured in 18 h days [105]. Additionally, light quality affects seed development and germinability. For example, in mouse-ear cress (Arabidopsis thaliana L.) plants that were grown in white light with either high or low red/far-red (R/FR) ratios, seeds that matured under a high R/FR ratio had a higher germination percentage in the dark than those that matured under a low R/FR ratio [106]. In several species, an inverse relationship was found between dark germinability and the chlorophyll content of the surrounding tissues of mature seeds [107].
During seed development, drought stress can also affect the subsequent germinability of seeds. However, the effects of drought stress vary with species. In species with mechanically imposed dormancy, drought usually increases seed coat thickness, reduces permeability, and results in decreased germinability. However, in species with chemically imposed dormancy, drought usually reduces dormancy by changing the balance between germination inhibitors and promoters [5]. Under water stress, seed dormancy decreased in Johnsongrass (Sorghum halepense (L.) Pers.) [108] and A. thaliana [109], while it increased in soybean (Glycine max (L.) Merr.) [110]. In seeds of pitcher’s stitchwort (Arenaria patula Michx. var. robusta) matured at the same site over three years, germinability was highest for seeds from the driest year [111].
Compared to other environmental factors discussed above, the effects of nutrients during seed development on subsequent seed germinability have not been studied extensively [5]. Large amounts of nitrogen in lamb’s-quarter (Chenopodium album L.) [112] and calcium in peanut (Arachis hypogaea L.) [113] resulted in enhanced germination in these species. In their book Seeds, Baskin and Baskin also listed several species in which the addition of mineral nutrients during seed development affected subsequent germinability [15].
6. The Effects of Post-Dispersal Factors on Seed Dormancy: After-Ripening Conditions
After-ripening can be defined as any changes that occur in seeds in the dry state (not imbibed) post-dispersal that cause or improve germination (see Figure 1). The term after-ripening, which has been applied to changes during dry storage [20,114,115], has also been used to describe changes during seed storage under either dry or moist conditions [116]. However, seed storage in the imbibed state, usually at low temperatures, is called stratification [114]. Here, the term “after-ripening” is used to describe physical or chemical changes in the seed coat, endosperm, and/or embryo that occur during the dry storage of seeds and that improve or cause germination. Since several to many factors regulate seed dormancy, after-ripening may involve more than one process [117].
Dry storage increases seed germinability in many species, such as wild oats (Avena fatua L.) [118], hairyfruit chervil (Chaerophyllum tainturieri Hook.) [119], downy brome (Bromus tectorum L.) [120], capeweed (Arctotheca calendula (L.) K. Lewin) [121], night-flowering catchfly (Silene noctiflora L.) [6], and milk thistle (Silybum marianum (L.) Gaertn.) [122]. In some cases, however, dry storage decreases germination percentage, as was shown for sea oats (Uniola paniculate L.) [123], European hazelnut (Corylus avellana L.) [124], and snow gum (Eucalyptus pauciflora Sieber ex Spreng.) [125], and germination rate, as was found in giant ragweed (Ambrosia trifida L.) [126].
After-ripening is influenced strongly by storage conditions and duration [20,22,127]. Cyclic changes in seed dormancy can occur in dry storage at constant temperatures, as was shown for rough bluegrass (Poa trivialis L.). In this species, the pattern can be regulated by an endogenous rhythm present within the seeds and/or by seasonal changes in temperature [128]. In many species, after-ripening is dependent on temperature, which determines the rate of dormancy loss [15,19]. Studies have revealed that dry storage can improve seed germinability through alterations to the seed coat, resulting in increased imbibition and, consequently, high germination percentages [41]. After-ripening, which widens the range of germination temperatures, generally takes place in dry seeds or seeds with low water content. However, if seeds become too dry (lower than 5% moisture), the process will be delayed. Seed moisture content, temperature, and oxygen availability are important factors in after-ripening [22].
Several studies have revealed that during after-ripening, the loss of dormancy can be caused by a gradual shift in the threshold level of temperature or water potential to more negative values. As a result, germination percentages and rates increase, and seeds can germinate synchronously at low water potentials [129,130,131]. It was speculated that progressive dormancy loss in a seed population could be related to a progressive decrease in the mean base water potential [132].
Attempts have been made to correlate after-ripening with changes in seed chemical properties [7,133]. Narasimhareddy and Swamy (1979) reported that in peanuts (Arachis hypogaea L.), the level of ABA–like inhibitors decreased during after-ripening, while that of cytokinins increased [133]. In another study with after-ripened A. hypogaea seeds, it was shown that ABA and phenolic compounds antagonized IAA (indole-3-acetic acid)- and GA3 (gibberellic acid)-induced metabolic processes. In this case, during after-ripening, the amounts of ABA and phenolics decreased, while those of IAA and GA3 increased, concomitant with an increase in germination [134]. Additionally, an inverse relationship was found between the concentration of phenolic compounds and the length of the storage period [135].
Analytical methods have also been used to obtain further information on the chemical aspects of after-ripening. For instance, it was reported that volatile compounds might be involved in the after-ripening of common cocklebur (Xanthium pensylvanicum Wallr.) seeds [136]. Furthermore, in barley (Hordeum vulgare L.), qualitative and quantitative changes in phenolic acids were found during after-ripening and were suggested as indications of a positive correlation between phenolic acids and dormancy levels [137]. In O. acanthium, after-ripening decreased moisture content and wax load and increased phenolic compounds, but had no effect on lignin content [115].
7. Scotch Thistle: A Model Plant Species to Study Seed Dormancy
O. acanthium is a noxious weed with a wide range of distribution, from cold areas such as Scandinavia and Western and Eastern Siberia to warm areas such as North Africa, Italy, Greece, Spain, Portugal, Australia, California, and Texas [138]. It occurs most commonly in wastelands, pastures, fence lines, gravelly riverbanks, and well-drained sandy or gravelly soils [139]. Additionally, it has been found in agricultural fields and causes problems in infested areas [3,140,141,142]. The density and vigour of this species vary from year to year, probably because of climatic conditions, but this is not well understood [143].
O. acanthium is a monocarpic winter annual, biennial, or short-lived perennial reproducing almost entirely by cypselae (see Figure 2). Depending on size, a single plant can produce from as few as 100 to as many as 50,000 cypselae [3]. After maturation, cypselae are released from the parent plant; however, some are retained in the capitulum for weeks or months. The only significant method of dispersal is by cypselae, each of which is equipped with a stout pappus (see Figure 2A) [144]. Some cypselae germinate as soon as they reach the soil in late summer or early autumn if adequate germination conditions are met. Before the onset of winter, the resulting seedlings form sizable rosettes. Spring-germinating plants are often larger than autumn-germinating plants, ensuring that they flower in the next year. The following year, bolting rosettes flower, produce cypselae, and then die. If cypselae do not germinate soon after dispersal, they are incorporated into seed banks and overwinter in a dormant state. The next year, they may germinate in spring or early summer, remain in the rosette stage until the summer of the following year, then bolt, flower, produce cypselae, and die. These two kinds of plants are winter annuals and biennials, respectively. If flowering plants are damaged by ineffective cutting, cultivation, or herbicides, they may produce some cypselae, but then become short-lived perennials by producing regrowth that will bolt in the following year, produce cypselae, and then die [138].
Cypselae of O. acanthium have a wide range of germination responses. Some cypselae may germinate in the autumn shortly after dispersal [138], while others may remain dormant for at least 40 years in the soil [15]. It was found that cypselae from a single plant could differ greatly in seed dormancy [145]. The germination responses of O. acanthium cypselae to high temperatures, dry heat, gibberellic acid, soil nitrate, scarification, stratification, leaching, red light, and other stimulating factors have been reported [141,146,147,148,149]. In a recent study with two local populations of O. acanthium, the application of potassium nitrate, gibberellic acid, sulfuric acid, and wet and dry prechilling resulted in increased cypsela germination [150]. These studies have been performed in different parts of the world and contain contradictory results.
O. acanthium shows an increase in cypsela germinability after maturation under high temperatures or dry storage and exhibits intermittent germination after maturation under cool field conditions [20,138]. In a study using four collections from each of two populations of O. acanthium, seedling emergence was monitored over three years. During this time, only 26% of the cypselae emerged, and the non-germinated cypselae had significantly more surface wax than the germinated ones [81]. Cypselae that ripened during the warmest temperatures of the season had the highest emergence percentage in the year of collection, while more of those that ripened under cooler temperatures emerged in later years. Under warmer conditions, smaller cypselae with thinner coats and smaller amounts of phenolics and waxes were produced. As a result, these cypselae were highly germinable [3,46].
After-ripening significantly increased germination percentages and rates for both warm- and cool-matured cypselae and eliminated differences between them [3,46]. Under a field environment in soil, cypselae gradually lose their dormancy. When there is precipitation, changes in wax content and the resulting increase in the solubility of phenolic compounds due to the alteration in the permeability of the cypsela coat may allow the removal of the water-soluble inhibitors from cypselae and enhance their germinability [13].
8. Concluding Remarks
In this review, I highlighted aspects of weed seed dormancy and germination that are useful for elucidating the population dynamics of weeds with reference to O. acanthium. As shown, O. acanthium produces cypselae that are either dormant or nondormant (see Figure 3). After separation from the parent plant, cypselae with primary dormancy can become nondormant through after-ripening or stratification under favourable environmental conditions. Adverse environmental conditions can cause death in both dormant and nondormant cypselae. If the germination conditions are suitable, nondormant cypselae will germinate. However, if there is a period of wetting and drying or the germination conditions are not suitable because of some other environmental factor(s), such as low temperature or low oxygen, cypselae may enter conditional dormancy and, finally, into secondary dormancy [15]. Few of these cypselae may die, but the majority will remain dormant until suitable conditions for relieving secondary dormancy are met.
The population dynamics of O. acanthium with regards to pre- and post-dispersal factors, particularly temperature, are presented in a “tentative” model of germination (see Figure 4). In this model, the total number of cypselae produced by the mother plant in high temperatures was set to 100 and the proportions of cypselae from low temperatures were calculated accordingly. The model reflects the fact that plants grown in higher temperatures produce fewer cypselae than those grown in lower temperatures. During maturation, higher temperatures cause more cypselae to abort. This could be related to the smaller size of the plants and the shorter maturation time, in which insufficient resources are available for allocation to developing cypselae. Additionally, cypselae that mature under warmer conditions have thinner and rougher coats, lower phenolic and wax contents, and correspondingly higher germinability [3,13]. A large proportion of the viable cypselae from plants grown in warmer temperatures are able to germinate in late summer or autumn shortly after dispersal. If the cypselae remain non-germinated, they are vulnerable to unpredictable environmental disasters because of their thinner and rougher coats.
After germination in late summer, these cypselae can produce sizeable rosettes. After overwintering, these rosettes bolt, flower, and produce cypselae in the following year. However, cypselae that mature under cooler conditions have thicker and smoother coats and are resistant to adverse environmental conditions. Very few of them germinate in the autumn of the year of their maturation. Dormant cypselae are incorporated into the persistent soil seed bank and germinate intermittently over a long period. Since every cypsela has its own microenvironment, each experiences a unique set of conditions. Thus, some cypselae germinate, while others remain non-germinated in the soil because of unsuitable germination conditions, even though they may be quiescent. From any dormancy state, there are few cypselae that cannot survive adverse environmental conditions, either before or after entering the seed bank. Global warming is currently a “hot topic” because of its relationship to adverse effects on many aspects of the lives of humans and other creatures. Increases in air temperature will affect the population dynamics of O. acanthium. As a result of global warming, this species will produce a large proportion of nondormant cypselae that will germinate shortly after dispersal. Thus, a smaller, more transient seed bank will be produced. Since a large seed bank is key to the overall success of a weed, this species should become less important as a weed in these warmer areas. However, the opposite is true for plants that produce dormant seeds under higher temperatures.
In conclusion, understanding maturation conditions, especially temperature; the substrate on which seeds are dispersed; and the germination performance of seeds produced at that temperature on the particular substrate help us (1) design a model that can predict seed dormancy loss, germination patterns in the field, the size and persistence of a resulting seed bank, and possibly the density of emerging seedling cohorts in a subsequent population; (2) develop effective control methods; and (3) increase agricultural production in the face of climate change.
Funding
This work was financially supported by a Discovery grant from the Natural Sciences and Engineering Research Council (NSERC) of Canada to M.M.Q.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
I thank NSERC for financial support, Mount Saint Vincent University for logistic support, and Paul Cavers for useful comments on the previous version of this manuscript. I appreciate constructive comments on the manuscript from the Academic Editor and three anonymous referees.
Conflicts of Interest
The author claims no conflict of interest.
References
- Gutterman, Y. Environmental factors and survival strategies of annual plant species in the Negev Desert, Israel. Plant Species Biol. 2000, 15, 113–125. [Google Scholar] [CrossRef]
- Gutterman, Y. Maternal effects on seeds during development. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CAB International: Wallingford, UK, 2000; pp. 59–84. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B.; Bernards, M.A. Pre- and post-dispersal factors regulate germination patterns and structural characteristics of Scotch thistle (Onopordum acanthium) cypselas. New Phytol. 2003, 159, 263–278. [Google Scholar] [CrossRef] [PubMed]
- LeFait, B.M.; Qaderi, M.M. Maternal environmental effects of temperature and exogenous gibberellic acid on seed and seedling traits of four populations of evening primrose (Oenothera biennis). Seeds 2022, 1, 110–125. [Google Scholar] [CrossRef]
- Fenner, M. The effects of the parent environment on seed germinability. Seed Sci. Res. 1991, 1, 75–84. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Reid, D.M. Combined effects of temperature and carbon dioxide on plant growth and subsequent seed germinability of Silene noctiflora. Int. J. Plant Sci. 2008, 169, 1200–1209. [Google Scholar] [CrossRef]
- Née, G.; Xiang, Y.; Soppe, W.J.J. The release of dormancy, a wake-up call for seeds to germinate. Curr. Opin. Plant Biol. 2017, 35, 8–14. [Google Scholar] [CrossRef]
- Gray, D.; Steckel, J.R.A.; Dearman, J.; Brocklehurst, P.A. Some effects of temperature during seed development on carrot (Daucus carota) seed growth and quality. Ann. Appl. Biol. 1988, 112, 367–376. [Google Scholar] [CrossRef]
- Wulff, R.D. Environmental maternal effects on seed quality and germination. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 491–505. [Google Scholar]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Klupczyńska, E.A.; Pawłowski, T.A. Regulation of seed dormancy and germination mechanisms in changing environment. Int. J. Mol. Sci. 2021, 22, 1357. [Google Scholar] [CrossRef]
- Nautiyal, P.C.; Sivasubramaniam, K.; Dadlani, M. Seed dormancy and regulation of germination. In Seed Science and Technology: Biology, Production, Quality; Dadlani, M., Yadava, D.K., Eds.; Springer: Gateway East, Singapore, 2023; pp. 39–66. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B.; Bernards, M.A. Isolation and structural characterization of a water-soluble germination inhibitor from Scotch thistle (Onopordum acanthium) cypselas. J. Chem. Ecol. 2003, 29, 2425–2438. [Google Scholar] [CrossRef]
- Cavers, P.B.; Qaderi, M.M.; Manku, R.; Downs, M.P. Intermittent germination: Causes and ecological implications. In Seed Biology: Advances and Applications; Black, M., Bradford, K.J., Vázquez-Ramos, J., Eds.; CAB International: Wallingford, UK, 2000; pp. 363–374. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Smith, R.G.; Birthisel, S.K.; Bosworth, S.C.; Brown, B.; Davis, T.M.; Gallandt, E.R.; Hazelrigg, A.; Venturini, E.; Warren, N.D. Environmental correlates with germinable weed seedbanks on organic farms across northern New England. Weed Sci. 2018, 66, 78–93. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Physiology and Biochemistry of Seeds. 2. Viability, Dormancy, and Environmental Control; Springer: Berlin/Heidelberg, Germany, 1982. [Google Scholar]
- Egley, G.H.; Duke, S.O. Physiology of weed seed dormancy and germination. In Weed Physiology. Vol. I. Reproduction and Ecophysiology; Duke, S.O., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 27–64. [Google Scholar]
- Simpson, G.M. Seed Dormancy in Grasses; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Qaderi, M.M.; Presti, A.; Cavers, P.B. Dry storage effects on germinability of Scotch thistle (Onopordum acanthium) cypselas. Acta Oecol. 2005, 27, 67–74. [Google Scholar] [CrossRef]
- Zhang, R.; Baskin, J.M.; Baskin, C.C.; Mo, Q.; Chen, L.; Hu, X.; Wang, Y. Effect of population, collection year, after-ripening and incubation condition on seed germination of Stipa bungeana. Sci. Rep. 2017, 7, 13893. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Bentsink, L.; Koornneef, M. Seed dormancy and germination. Arab. Book 2008, 6, e0119. [Google Scholar] [CrossRef]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; de Casas, R.R.; NESCent Germination Working Group. The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Sun, L.; Baskin, C.C.; Baskin, J.M.; Cao, M.; Yang, J. Seed dormancy in space and time: Global distribution, paleoclimatic and present climatic drivers, and evolutionary adaptations. New Phytol. 2022, 234, 1770–1781. [Google Scholar] [CrossRef]
- Salisbury, E.J. Weeds and Aliens; Collins: London, UK, 1961. [Google Scholar]
- Nonogaki, H. Seed dormancy and germination—emerging mechanisms and new hypotheses. Front. Plant Sci. 2014, 5, 233. [Google Scholar] [CrossRef]
- Foley, M.E. Weeds, seeds, and buds–opportunities and systems for dormancy investigations. Weed Sci. 2002, 50, 267–272. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef]
- Bradford, K.J. Applications of hydrothermal time to quantifying and modeling seed germination and dormancy. Weed Sci. 2002, 50, 248–260. [Google Scholar] [CrossRef]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J.J. Molecular mechanisms of seed dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef]
- Werker, E. Seed Anatomy; Gebrüder Borntraeger: Berlin/Stuttgart, Germany, 1997. [Google Scholar]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977. [Google Scholar]
- Nikolaeva, M.G. Factors affecting the seed dormancy pattern. In The Physiology and Biochemistry of Seed Development, Dormancy and Germination; Khan, A.A., Ed.; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1982; pp. 51–74. [Google Scholar]
- Hilhorst, H.W.M. A critical update on seed dormancy. I. Primary dormancy. Seed Sci. Res. 1995, 5, 61–73. [Google Scholar] [CrossRef]
- Vleeshouwers, L.M.; Boumeester, H.J.; Karssen, C.M. Redefining seed dormancy: An attempt to integrate physiology and ecology. J. Ecol. 1995, 83, 1031–1037. [Google Scholar] [CrossRef]
- Harper, J.L. The ecological significance of dormancy and its importance in weed control. Proc. IVth Int. Cong. Crop Prot. 1957, 4, 415–420. [Google Scholar]
- Rees, M. Seed dormancy. In Plant Ecology, 2nd ed.; Crawley, M.J., Ed.; Blackwell Science Ltd.: Oxford, UK, 1997; pp. 214–238. [Google Scholar]
- Doucet, C.; Cavers, P.B. Induced dormancy and colour polymorphism in seeds of the bull thistle Cirsium vulgare (Savi) Ten. Seed Sci. Res. 1997, 7, 399–407. [Google Scholar] [CrossRef]
- Bradbeer, J.W. Seed Dormancy and Germination; Chapman & Hall: New York, NY, USA, 1988. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. The great diversity in kinds of seed dormancy: A revision of the Nikolaeva-Baskin classification system for primary seed dormancy. Seed Sci. Res. 2021, 31, 249–277. [Google Scholar] [CrossRef]
- Kitajima, K.; Fenner, M. Ecology of seedling regeneration. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CAB International: Wallingford, UK, 2000; pp. 331–359. [Google Scholar]
- de Casas, R.R.; Willis, C.G.; Pearse, W.D.; Baskin, C.C.; Baskin, J.M.; Cavender-Bares, J. Global biogeography of seed dormancy is determined by seasonal and seed size: A case study in the legumes. New Phytol. 2017, 214, 1527–1536. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Cavers, P.B.; Hamill, A.S.; Downs, M.P.; Bernards, M.A. Maturation temperature regulates germinability and chemical constituents of Scotch thistle (Onopordum acanthium) cypselas. Can. J. Bot. 2006, 84, 28–38. [Google Scholar] [CrossRef]
- Gardarin, A.; Dürr, C.; Colbach, N. Prediction of germination rates of weed species: Relationships between germination speed parameters and species traits. Ecol. Modell. 2011, 222, 626–636. [Google Scholar] [CrossRef]
- Casal, J.J.; Sánchez, R.A. Phytochromes and seed germination. Seed Sci. Res. 1998, 8, 317–329. [Google Scholar] [CrossRef]
- Malavert, C.; Batlla, D.; Benech-Arnold, R.L. Light sensitivity changes during dormancy induction in Polygonum aviculare L. seeds: Development of a predictive model of annual changes in seed-bank light sensitivity in relation to soil temperature. Weed Res. 2021, 61, 115–125. [Google Scholar] [CrossRef]
- Bączek-Kwinta, R. An interplay of light and smoke compounds in photoblastic seeds. Plants 2022, 11, 1773. [Google Scholar] [CrossRef] [PubMed]
- Pons, T.L. Seed responses to light. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI International: Wallingford, UK, 2000; pp. 237–260. [Google Scholar]
- Hilhorst, H.W.M.; Toorop, P.E. Review on dormancy, germinability and germination in crop and weed seeds. Adv. Agron. 1997, 61, 111–165. [Google Scholar]
- Taylorson, R.B. Environmental and chemical manipulation of weed seed dormancy. Rev. Weed Sci. 1987, 3, 135–154. [Google Scholar]
- Werker, E. Seed dormancy as explained by the anatomy of embryo envelopes. Isr. J. Bot. 1980, 29, 22–44. [Google Scholar]
- Kelly, K.M.; Van Staden, J.; Bell, W.E. Seed coat structure and dormancy. Plant Growth Regul. 1992, 11, 201–209. [Google Scholar] [CrossRef]
- Pereira, M.P.; Corrêa, F.F.; Polo, M.; de Castro, E.M.; Cardoso, A.A.; Pereira, F.J. Seed germination of Schinus molle L. (Anacardiaceae) as related to its anatomy and dormancy alleviation. Seed Sci. Res. 2016, 26, 351–361. [Google Scholar] [CrossRef]
- Karssen, C.M. Seasonal patterns of dormancy in weed seeds. In The Physiology and Biochemistry of Seed Development, Dormancy and Germination; Khan, A.A., Ed.; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1982; pp. 243–270. [Google Scholar]
- Benech-Arnold, R.L.; Sánchez, R.A.; Forcella, F.; Kruk, B.C.; Ghersa, C.M. Environmental control of dormancy in weed seed banks in soil. Field Crops Res. 2000, 67, 105–122. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Just, M.; Cross, A.T.; Lewandrowski, W.; Turner, S.R.; Merritt, D.J.; Dixon, K. Seed dormancy alleviation by warm stratification progressively widens the germination window in Mediterranean climate Rutaceae. Aust. J. Bot. 2023, 71, 55–66. [Google Scholar] [CrossRef]
- Pearson, C.F.; Ison, R.L. Agronomy of Grassland Systems, 2nd ed.; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Taiz, L.; Møller, I.M.; Murphy, A.; Zeiger, E. Plant Physiology and Development, 7th ed.; Sinauer: Sunderland, MA, USA, 2023. [Google Scholar]
- Hilhorst, H.W.M.; Karssen, C.M. Seed dormancy and germination: The role of abscisic acid and gibberellins and the importance of hormone mutants. Plant Growth Regul. 1992, 11, 225–238. [Google Scholar] [CrossRef]
- Footitt, S.; Walley, P.G.; Lynn, J.R.; Hambidge, A.J.; Penfield, S.; Finch-Savage, W.E. Trait analysis reveals DOG1 determines initial depth of seed dormancy, but not changes during dormancy cycling that result in seedling emergence timing. New Phytol. 2020, 225, 2035–2047. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.-Q.; Luan, S.; Li, J.; He, Z.-H. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.-D.; Xie, Q.; He, Z.-H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef]
- MacGregor, D.R.; Zhang, N.; Iwasaki, M.; Chen, M.; Dave, A.; Lopez-Molina, L.; Penfield, S. ICE1 and ZOU determine the depth of primary seed dormancy in Arabidopsis independently of their role in endosperm development. Plant J. 2019, 98, 277–290. [Google Scholar] [CrossRef]
- Radosevich, S.; Holt, J.; Ghersa, C. Weed Ecology: Implications for Management, 2nd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Balyan, R.S.; Bhan, V.M. Germination of horse purslane (Trianthema portulacastrum) in relation to temperature, storage conditions, and seedling depths. Weed Sci. 1986, 34, 513–515. [Google Scholar] [CrossRef]
- Thompson, K.; Grime, J.P. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. J. Ecol. 1979, 67, 893–921. [Google Scholar] [CrossRef]
- Nakabayashi, K.; Leubner-Metzger, G. Seed dormancy and weed emergence: From simulating environmental change to understanding trait plasticity, adaptive evolution, and population fitness. J. Exp. Bot. 2021, 72, 4181–4185. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Footitt, S. Seed dormancy cycling and the regulation of dormancy mechanisms to time germination in variable field environments. J. Exp. Bot. 2017, 68, 843–856. [Google Scholar] [CrossRef]
- Wang, G.; Baskin, C.C.; Baskin, J.M.; Yang, X.; Liu, G.; Zhang, X.; Ye, X.; Huang, Z. Timing of seed germination in two alpine herbs on the southeastern Tibetan plateau: The role of seed dormancy and annual dormancy cycling in soil. Plant Soil 2017, 421, 465–476. [Google Scholar] [CrossRef]
- Zhang, K.; Ji, Y.; Fu, G.; Yao, L.; Liu, H.; Tao, J.; Walck, J.L. Dormancy cycles in Aquilegia oxysepala Trautv. et Mey. (Ranunculaceae), a species with non-deep simple morphophysiological dormancy. Plant Soil 2021, 464, 223–235. [Google Scholar] [CrossRef]
- Soltani, E.; Baskin, C.C.; Baskin, J.M.; Soltani, A.; Galeshi, S.; Ghaderi-far, F.; Zeinali, E. A quantitative analysis of seed dormancy and germination in the winter annual weed Sinapis arvensis (Brassicaceae). Botany 2016, 94, 289–300. [Google Scholar] [CrossRef]
- Garcia, Q.S.; Barreto, L.C.; Bicalho, E.M. Environmental factors driving seed dormancy and germination in tropical ecosystems: A perspective from campo rupestre species. Environ. Exp. Bot. 2020, 178, 104164. [Google Scholar] [CrossRef]
- Mašková, T.; Poschlod, P. Soil seed bank persistence across time and burial depth in calcareous grassland habitats. Front. Plant Sci. 2022, 12, 790867. [Google Scholar] [CrossRef]
- Telewski, F.W.; Zeevaart, J.A.D. The 120-yr period for Dr. Beal’s seed viability experiment. Am. J. Bot. 2002, 89, 1285–1288. [Google Scholar] [CrossRef] [PubMed]
- Fenner, M. Ecology of seed banks. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 507–528. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B. Interpopulation and interyear variation in germination in Scotch thistle, Onopordum acanthium L., grown in a common garden: Genetics vs environment. Plant Ecol. 2002, 162, 1–8. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Cavers, P.B.; Bernards, M.A. Seed bank dynamics of Onopordum acanthium: Emergence patterns and chemical attributes. J. Ecol. 2002, 90, 672–683. [Google Scholar] [CrossRef]
- Loades, E.; Pérez, M.; Turečková, V.; Tarkowská, D.; Strnad, M.; Seville, A.; Nakabayashi, K.; Leubner- Metzger, G. Distinct hormonal and morphological control of dormancy and germination in Chenopodium album dimorphic seeds. Front. Plant Sci. 2023, 14, 1156794. [Google Scholar] [CrossRef]
- Evert, R.F.; Eichhorn, S.E. Raven Biology of Plants, 8th ed.; W.H. Freeman & Company: New York, NY, USA, 2013. [Google Scholar]
- Boesewinkel, F.D.; Bouman, F. The seed: Structure and function. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 1–24. [Google Scholar]
- Yadegari, R.; Goldberg, R.B. Embryogenesis in dicotyledonous plants. In Cellular and Molecular Biology of Plant Seed Development; Larkins, B.A., Vasil, I.K., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997; pp. 3–52. [Google Scholar]
- Perez-Grau, L. Plant embryogenesis–the cellular design of a plant. In Plant Reproduction; O’Neill, S.D., Roberts, J.A., Eds.; Sheffield Academic Press: Sheffield, UK, 2002; pp. 154–192. [Google Scholar]
- Bryant, J.A. Seed Physiology; Edward Arnold (Publishers) Ltd.: London, UK, 1985. [Google Scholar]
- Demason, D.A. Endosperm structure and development. In Cellular and Molecular Biology of Plant Seed Development; Larkins, B.A., Vasil, I.K., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997; pp. 73–115. [Google Scholar]
- Brown, R.C.; Lemmon, B.E.; Nguyen, H. Endosperm development. In Plant Reproduction; O’Neill, S.D., Roberts, J.A., Eds.; Sheffield Academic Press: Sheffield, UK, 2002; pp. 193–220. [Google Scholar]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 27, R874–R878. [Google Scholar] [CrossRef]
- Tran, V.N.; Cavanagh, A.K. Structural aspects of dormancy. In Seed Physiology. 2. Germination and Reserve Mobilization; Murray, D.R., Ed.; Academic Press Australia: North Ryde, NSW, Australia, 1984; pp. 1–44. [Google Scholar]
- Zhang, Y.; Zhang, K.; Ji, Y.; Tao, J. Physical dormancy and soil seed bank dynamics in seeds of Melilotus albus (Fabaceae). Flora 2020, 266, 151600. [Google Scholar] [CrossRef]
- Roach, D.A.; Wulff, R.D. Maternal effects in plants. Ann. Rev. Ecol. Syst. 1987, 18, 209–235. [Google Scholar] [CrossRef]
- Fenner, M. Environmental influences on seed size and composition. Hortic. Rev. 1992, 13, 183–213. [Google Scholar]
- Murdoch, A.J.; Ellis, R.H. Dormancy, viability and longevity. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CAB International: Wallingford, UK, 2000; pp. 183–214. [Google Scholar]
- Huang, Z.; Footitt, S.; Tang, A.; Finch-Savage, W.E. Predicted global warming scenarios impact on the mother plant to alter seed dormancy and germination behaviour in Arabidopsis. Plant Cell Environ. 2018, 41, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Fitter, A.H.; Hay, R.K.M. Environmental Physiology of Plants, 3rd ed.; Academic Press: New York, NY, USA, 2002. [Google Scholar]
- Lyndon, R.F. The environmental control of reproductive development. In Fruit and Seed Production: Aspects of Development, Environmental Physiology and Ecology; Marshall, C., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1992; pp. 9–32. [Google Scholar]
- Duffus, C.M. Control of grain growth and development. In Fruit and Seed Production: Aspects of Development, Environmental Physiology and Ecology; Marshall, C., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1992; pp. 125–149. [Google Scholar]
- Boyce, K.G. Effect of temperature and dormancy on germination of tall fescue. Crop Sci. 1976, 16, 15–18. [Google Scholar] [CrossRef]
- Van der Vegte, F.W. Population differentiation and germination ecology in Stellaria media (L.) Vill. Oecologia 1978, 37, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Sawhney, R.; Naylor, J.M. Dormancy studies in seed of Avena fatua. 12. Influence of temperature on germination behavior of nondormant families. Can. J. Bot. 1980, 58, 578–581. [Google Scholar] [CrossRef]
- Somody, C.N.; Nalewaja, J.D.; Miller, S.E. The response of wild oat (Avena fatua) and Avena sterilis accessions to photoperiod and temperature. Weed Sci. 1984, 32, 206–213. [Google Scholar] [CrossRef]
- Probert, R.J.; Smith, R.D.; Birch, P. Germination responses to light and alternating temperatures in European populations of Dactylis glomerata L. II. The genetic and environmental components of germination. New Phytol. 1985, 99, 317–322. [Google Scholar] [CrossRef]
- Karssen, C.M. The light promoted germination of the seeds of Chenopodium album L. III. Effect of the photoperiod during growth and development of the plants on the dormancy of the produced seeds. Acta Bot. Neerl. 1970, 19, 81–94. [Google Scholar] [CrossRef]
- McCullough, J.M.; Shropshire, W. Physiological predetermination of responses in Arabidopsis thaliana (L.) Heynh. Plant Cell Physiol. 1970, 11, 139–148. [Google Scholar] [CrossRef]
- Cresswell, E.G.; Grime, J.P. Induction of a light requirement during seed development and its ecological consequences. Nature 1981, 291, 583–585. [Google Scholar] [CrossRef]
- Benech-Arnold, R.L.; Fenner, M.; Edwards, P.J. Changes in dormancy level in Sorghum halepense seeds induced by water stress during seed development. Funct. Ecol. 1992, 6, 596–605. [Google Scholar] [CrossRef]
- Abo Gamar, M.I.; Qaderi, M.M. Interactive effects of temperature, carbon dioxide and watering regime on seed germinability oftwo genotypes of Arabidopsis thaliana. Seed Sci. Res. 2019, 29, 12–20. [Google Scholar] [CrossRef]
- Hill, H.J.; West, S.H.; Hinson, K. Effect of water stress during seed fill on impermeable seed expression in soybean. Crop Sci. 1986, 26, 807–812. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Year-to-year variation in the germination of freshly-harvested seeds of Arenaria patula var. robusta from the same site. J. Tenn. Acad. Sci. 1975, 50, 106–108. [Google Scholar]
- Fawcett, R.S.; Slife, F.W. Effects of field applications of nitrate on weed seed germination and dormancy. Weed Sci. 1978, 26, 594–596. [Google Scholar] [CrossRef]
- Coffelt, T.A.; Hallock, D.L. Soil fertility responses of Virginia-type peanut cultivars. Agron. J. 1986, 78, 131–137. [Google Scholar] [CrossRef]
- Mayer, A.M.; Poljakoff-Mayber, A. The Germination of Seeds, 4th ed.; Pergamon Press: Oxford, UK, 1989. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B.; Hamill, A.S.; Bernards, M.A. Effects of collection time and after-ripening on chemical constituents and germinability of Scotch thistle (Onopordum acanthium) cypselas. Botany 2012, 90, 755–762. [Google Scholar] [CrossRef]
- Lewak, S.; Rudnicki, R.M. After-ripening in cold-requiring seeds. In The Physiology and Biochemistry of Seed Development, Dormancy and Germination; Khan, A.A., Ed.; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1982; pp. 193–217. [Google Scholar]
- Thomson, J.R. An Introduction to Seed Technology; Leonard Hill: Glasgow, UK, 1979. [Google Scholar]
- Adkins, S.W.; Simpson, G.M. The physiological basis of seed dormancy in Avena fatua. IX. Characterization of two dormancy states. Physiol. Plant. 1988, 73, 15–20. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Germination ecophysiology of seeds of the winter annual Chaerophyllum tainturiei: A new type of morphophysiological dormancy. J. Ecol. 1990, 78, 993–1004. [Google Scholar] [CrossRef]
- Allen, P.S.; Meyer, S.E.; Beckstead, J. Patterns of seed after-ripening in Bromus tectorum L. J. Exp. Bot. 1995, 46, 1737–1744. [Google Scholar] [CrossRef]
- Chaharsoghi, A.T.; Jacobs, B. Manipulating dormancy of Capeweed (Arctotheca calendula L.) seed. Seed Sci. Res. 1998, 8, 139–146. [Google Scholar] [CrossRef]
- Liava, V.; Ntatsi, G.; Karkanis, A. Seed germination of three milk thistle (Silybum marianum (L.) Gaertn.) populations of Greek origin: Temperature, duration, and storage conditions effects. Plants 2023, 12, 1025. [Google Scholar] [CrossRef]
- Westra, R.N.; Loomis, N.E. Seed dormancy in Uniola paniculata. Am. J. Bot. 1966, 53, 407–411. [Google Scholar] [CrossRef]
- Bradbeer, J.W. Studies in seed dormancy. IV. The role of endogenous inhibitors and gibberellin in the dormancy and germination of Corylus avellana L. seeds. Planta 1968, 78, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Beardsell, D.; Mullett, J. Seed germination of Eucalyptus pauciflora Sieb. ex Spreng. from low and high altitude populations in Victoria. Aust. J. Bot. 1984, 32, 475–480. [Google Scholar] [CrossRef]
- Dong, H.; Liu, T.; Liu, Z.; Song, Z. Fate of the soil seed bank of giant ragweed and its significance in preventing and controlling its invasion in grasslands. Ecol. Evol. 2020, 20, 4854–4866. [Google Scholar] [CrossRef]
- Rojas-Aréchiga, M.; García-Morales, E. Dormancy and viability of Ferocactus peninsulae (Cactaceae) seeds. Plant Species Biol. 2022, 37, 173–181. [Google Scholar] [CrossRef]
- Froud-Williams, R.J.; Hilton, J.R.; Dixon, J. Evidence for an endogenous cycle of dormancy in dry stored seeds of Poa trivialis. New Phytol. 1986, 102, 123–131. [Google Scholar] [CrossRef]
- Christensen, M.; Meyer, S.E.; Allen, P.S. A hydrothermal time model of seed after-ripening in Bromus tectorum L. Seed Sci. Res. 1996, 6, 1–9. [Google Scholar] [CrossRef]
- Bauer, M.C.; Meyer, S.E.; Allen, P.S. A simulation model to predict seed dormancy loss in the field for Bromus tectorum L. J. Exp. Bot. 1998, 49, 1235–1244. [Google Scholar]
- Meyer, S.E.; Debaene-Gill, S.B.; Allen, P.S. Using hydrothermal time concepts to model seed germination response to temperature, dormancy loss, and priming effects in Elymus elymoides. Seed Sci. Res. 2000, 10, 213–223. [Google Scholar] [CrossRef]
- Bradford, K.J. Water relations in seed germination. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 351–396. [Google Scholar]
- Narasimhareddy, S.B.; Swamy, P.M. Abscisic acid-like inhibitors and cytokinins during after-ripening of dormant peanut seeds (Arachis hypogaea). Physiol. Plant. 1979, 46, 191–193. [Google Scholar] [CrossRef]
- Rao, M.R.K.; Rao, I.M. Dormancy regulation in groundnut seeds (Arachis hypogaea L.) C. Var. TMV-3. Curr. Sci. 1979, 48, 1000–1001. [Google Scholar]
- Kigel, J. Seed germination in arid and semiarid regions. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 645–699. [Google Scholar]
- Esashi, Y.; Zhang, M.; Segawa, K.; Furihata, T.; Nakaya, M.; Maeda, Y. Possible involvement of volatile compounds in the after-ripening of cocklebur seeds. Physiol. Plant. 1994, 90, 577–583. [Google Scholar] [CrossRef]
- Weidner, S.; Paprocka, J.; Lukaszewicz, D. Changes in free, esterified and glycosidic phenolic acids in cereal grains during the after-ripening. Seed Sci. Technol. 1996, 24, 107–114. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B. Variation in germination response within Scotch thistle, Onopordum acanthium L., populations matured under greenhouse and field conditions. Écoscience 2000, 7, 57–65. [Google Scholar] [CrossRef]
- Moore, R.J.; Frankton, C. The Thistles of Canada; Monograph No. 10; Canada Department of Agriculture: Ottawa, ON, Canada, 1974.
- Smith, H.A.; Johnson, W.S.; Scott Shonkwiler, J.; Swanson, S.R. The implications of variable or constant expansion rates in invasive weed infestations. Weed Sci. 1999, 47, 62–66. [Google Scholar] [CrossRef]
- Cavers, P.B.; Qaderi, M.M.; Threadgill, P.F.; Steel, M.G. The biology of Canadian weeds. 147. Onopordum acanthium L. Can. J. Plant Sci. 2011, 91, 739–758. [Google Scholar] [CrossRef]
- Wei, C.; Zhou, S.; Shi, K.; Zhang, C.; Shao, H. Chemical profile and phytotoxic action of Onopordum acanthium essential oil. Sci. Rep. 2020, 10, 13568. [Google Scholar] [CrossRef] [PubMed]
- Michael, P.W. Control of the biennial thistle, Onopordum, by amitrole and five perennial grasses. Aust. J. Exp. Agric. Anim. Husb. 1968, 8, 331–339. [Google Scholar] [CrossRef]
- Hyde-Wyatt, B.H.; Morris, D.I. The Noxious and Secondary Weeds of Tasmania; Department of Agriculture: Tasmania, Australia, 1980.
- Meier, L.R. Variation in Seeds of Onopordum acanthium. Master’s Thesis, Department of Plant Sciences, University of Western Ontario, London, ON, Canada, 1995. [Google Scholar]
- Scifres, C.J.; McCarty, M.K. Some factors affecting germination and seedling growth of Scotch thistle. Res. Bull. Nebr. Agric. Exp. Stn. 1969, 228, 1–29. [Google Scholar]
- Young, J.A.; Evans, R.A. Germination and persistence of achenes of Scotch thistle. Weed Sci. 1972, 20, 98–101. [Google Scholar] [CrossRef]
- Pérez-García, F. Effect of the origin of cypsela on germination of Onopordum acanthium L. (Asteraceae). Seed Sci. Technol. 1993, 21, 187–195. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B. Effects of dry heat on the germinability and viability of Scotch thistle (Onopordum acanthium) cypselas: Interpopulation and interposition variation. Can. J. Bot. 2003, 81, 684–697. [Google Scholar] [CrossRef]
- Sarabi, V.; Bagheri, A.; Chitband, A.A. Evaluation of different methods for breaking cypsela dormancy in two populations of Scotch thistle (Onopordum acanthium L.) collected from the west and northwest of Iran. Can J. Plant Sci. 2022, 102, 902–910. [Google Scholar] [CrossRef]
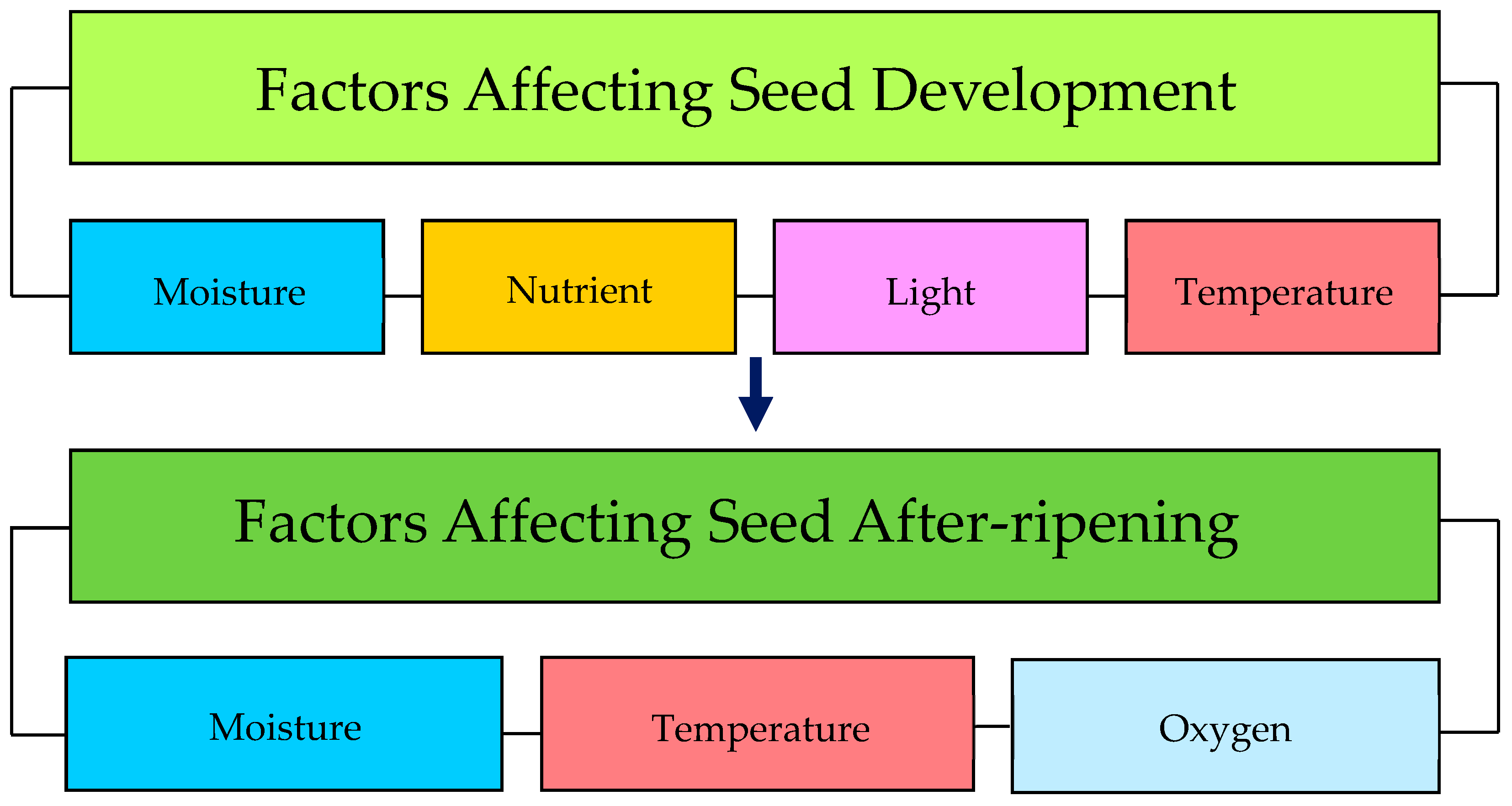
Figure 1.
Pre- and post-dispersal factors that regulate weed seed dormancy.

Figure 2.
Stereo zoom microscopy of Scotch thistle (Onopordum acanthium) cypselae. (A) Cypselae with pappi, (B) a typical cypsela matured under low (left) and high (right) temperatures, (C) a dissected cypsela showing coat layers and embryo, and (D) a germinating cypsela. Scale bars: 5 mm.
Figure 2.
Stereo zoom microscopy of Scotch thistle (Onopordum acanthium) cypselae. (A) Cypselae with pappi, (B) a typical cypsela matured under low (left) and high (right) temperatures, (C) a dissected cypsela showing coat layers and embryo, and (D) a germinating cypsela. Scale bars: 5 mm.

Figure 3.
A general model of changes that can take place in ripe seeds, including cypselae of Scotch thistle (Onopordum acanthium), from the time of dispersal.
Figure 3.
A general model of changes that can take place in ripe seeds, including cypselae of Scotch thistle (Onopordum acanthium), from the time of dispersal.

Figure 4.
A tentative model for cypsela population dynamics in Scotch thistle (Onopordum acanthium). Cypselae were collected from plants grown in higher and lower temperatures in a greenhouse. The number of cypselae per head that matured in higher temperatures was set to 100 and the relative proportions for other cypsela categories were calculated (values in parentheses) for those matured in both conditions. The thickness of each arrow indicates the relative number of cypselae. This figure is based on data from the cited literature [3,13,46,80,81,115].
Figure 4.
A tentative model for cypsela population dynamics in Scotch thistle (Onopordum acanthium). Cypselae were collected from plants grown in higher and lower temperatures in a greenhouse. The number of cypselae per head that matured in higher temperatures was set to 100 and the relative proportions for other cypsela categories were calculated (values in parentheses) for those matured in both conditions. The thickness of each arrow indicates the relative number of cypselae. This figure is based on data from the cited literature [3,13,46,80,81,115].

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Qaderi, M.M. Environmental Regulation of Weed Seed Dormancy and Germination. Seeds 2023, 2, 259-277. https://doi.org/10.3390/seeds2030020
AMA Style
Qaderi MM. Environmental Regulation of Weed Seed Dormancy and Germination. Seeds. 2023; 2(3):259-277. https://doi.org/10.3390/seeds2030020
Chicago/Turabian StyleQaderi, Mirwais M. 2023. "Environmental Regulation of Weed Seed Dormancy and Germination" Seeds 2, no. 3: 259-277. https://doi.org/10.3390/seeds2030020