1. Introduction
Edible mushrooms are considered functional foods and an excellent source of nutraceuticals with important roles in promoting human health and preventing diseases, such as cancer. However, our knowledge of the biocompounds responsible for these effects is still limited [
1,
2]. Recently, an ethanol-insoluble and water-soluble small RNA (sRNA) fraction, purified by anion-exchange chromatography from some edible mushrooms species such as
Cantharellus cibarius (CCI) and
Boletus edulis (BED), emerged as potent biomolecules, exhibiting the ability to induce apoptosis and inhibit cell proliferation in cancer cells without adverse effects on normal cells [
3,
4,
5].
To unravel the molecular nature of this sRNA, the sRNA fractions from Boletus edulis, Agaricus bisporus Portobello (ABI), and Cantharellus cibarius as well as microRNAs from ABI and BED were analyzed for their anti-cancer properties.
2. Methods
Mushrooms. In this work, the following two species of mushrooms were used:
B. edulis and
A. bishoprus (harvested in different regions of Trás-os-Montes and Alto Douro in late 2022). To ensure the preservation of the material, the frozen mushrooms were subjected to a freeze-drying process. The
C. cibarius sRNA fraction (CCI3), previously isolated and characterized [
5], was used as a control throughout this work.
Cell lines. Caco-2 (tumor cell line, HTB-37, ATCC) and HDFn (normal cell line, Human Dermal Fibroblasts, C-004-5C, Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) cells were cultured in an Advanced-DMEM medium supplemented with 13% AmnioMAX basal medium, 10% fetal bovine serum (FBS), 2% AmnioMAX supplement, 1% L-glutamine (200 mM), and 1% antibiotic (penicillin 5 mg/mL, streptomycin 5 g/mL, and neomycin 10 mg/mL) (Gibco, Thermo Fisher Scientific). The cells were maintained in a humidified atmosphere of 97% with 5% CO2 at 37 °C. Cell handling was performed using standard in vitro animal cell culture methodologies in a class II vertical flow chamber.
The extraction and purification of mushroom small RNAs by anion-exchange chromatography. Ethanol insoluble and water-soluble small RNA fractions were obtained as described by Lemieszek and colleagues [
3,
4,
5].
miRNA commercial kit. B. edulis and A. bisporus powder and HMWM (High Molecular Weight Material) were used for miRNA isolation. A MirVana miRNA Isolation Kit was used following manufacturer instruction.
MTT assay. Caco-2 and HDFn cells were plated on 96-well microplates at a density of 3 × 104 cells/mL. The next day, the culture medium was replaced by a fresh medium with serial dilutions of the tested fractions in triplicate, and control (untreated) cells were used for each assay. After 96 h, the cells were incubated for 4 h with MTT solution (5 mg/mL). Formazan crystals were solubilized with DMSO, and the product was quantified spectrophotometrically by measuring the absorbance at a 510 nm wavelength using a Tristar 3 Microplate Reader (Berthold Technologies, Bad Wildbad, Germany).
RNA Fluorescent in situ hybridization (RNA-FISH). Slides were prepared with 1 × 10
5 of Caco-2 cells. After 24 h, 250 µg/mL of CCI3 and BEDB were added to the cells, with untreated cells being used as a control. The RNA-FISH procedure was performed 96 h after exposure, following the method described by [
6]. The probe used was a cloned sequence isolated from a CCI3 sRNA fraction that was labelled with digoxigenin-11-dUTP (Roche Applied Science, Penzberg, Germany). Microscopy images were acquired in a Zeiss ImagerZ2 microscope coupled to an ORCA-Flash 4.0 digital camera (Hamamatsu Photonics, Shizuoka, Japan) and captured with SmartCapture 4 software (Digital Scientifc UK, Cambridge, UK). All the images were prepared for color and contrast optimization in the Adobe Photoshop software (version 7.0) as a whole.
Statistical analysis. The data are presented as mean ± standard deviation. Statistical significance was determined by one way ANOVA, multiple comparison Šidák test. ns—non-significant; p > 0.05; * p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; **** p ≤ 0.0001.
3. Results and Discussion
In this work, the previously characterized sRNA fraction isolated from
C. cibarius [
5] (named as CCI3) was used to compare the anti-cancer activity of RNA-enriched fractions that were ethanol-insoluble- and water-soluble-purified via anion-exchange chromatography from other mushroom species such as
Boletus edulis (BED) and
Agaricus bisporus (ABI). Using this purification method, it was possible to isolate two fractions from BED, named BEDA and BEDB. The BEDB fraction was enriched with nucleic acids, similar to what was previously reported [
4,
5]. On the other hand, it was not possible to isolate a similar fraction from
A. bisporus, suggesting that not all the mushrooms presented the same characteristics for the isolation of sRNA fractions, nor did this sRNA. However, the fraction ABIA was also tested for its anti-cancer properties. In order to understand whether these sRNA-enriched fractions were a mixture of sRNAs or a specific sequence, we extracted small RNAs (miRNAs) from the powder and HMWM of
B. edulis and
A. bisporus, named BEDS, BEDH and ABIS, ABIH, respectively.
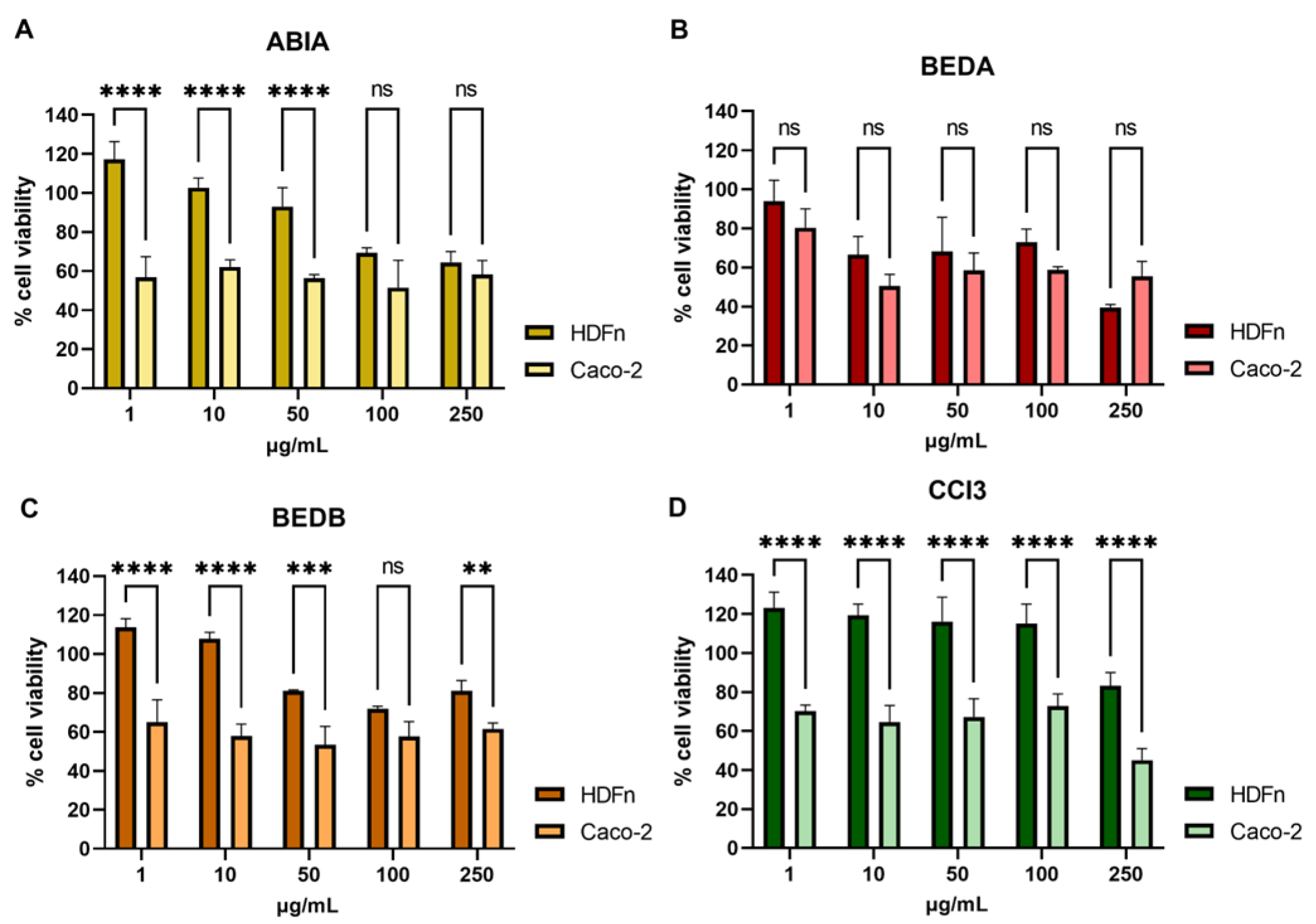
3.1. The Anti-Tumor Effect of Fractions Obtained from Caco-2 and HDFn Cells
With the aim of demonstrating the anti-proliferative activity of the different samples, the cell lines Caco-2 (tumor cell line) and HDFn (normal cell line) were treated with different concentrations of these samples for 96 h, followed by an assay MTT in order to assess cell viability. We were able to verify that the ABIA fraction (
Figure 1A) only causes a significant decrease in cell viability at a 50 µg/mL concentration in tumor cells, being cytotoxic to normal cells above this concentration. The BEDA fraction (
Figure 1B) did not show significant differences between normal and tumor cells. Regarding the BEDB (
Figure 1C) and CCI3 (
Figure 1D) fractions (sRNA-enriched), both showed significant decreases in the cell viability of the tumor cell line, showing cytotoxicity to normal cells only at high concentrations (250 µg/mL for CCI3 and from 50 µg/mL for BEDB). These data agree with the previously reported data [
5], and they highlight the potential of CCI3 for application in cancer therapy.
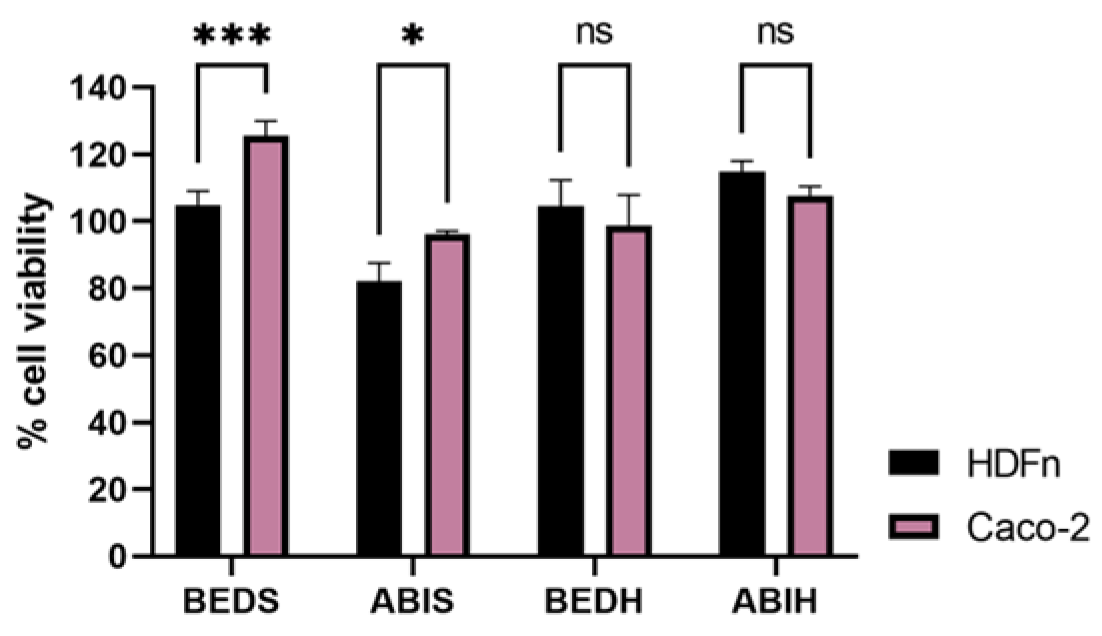
Regarding the results obtained in the MTT assay of the ABIS, ABIH, BEDS, and BEDH samples (
Figure 2), it was possible to verify that none of the samples caused a greater decrease in cell viability in the tumor cells. Despite the similar purity and molecular weight when compared with the CCI3 and BEDB fractions, the BEDS and BEDH samples did not show the same biological effect. These data suggest a different primary structure of sRNA and a sequence-dependent effect.
3.2. RNA Fluorescent In Situ Hybridization (RNA-FISH)
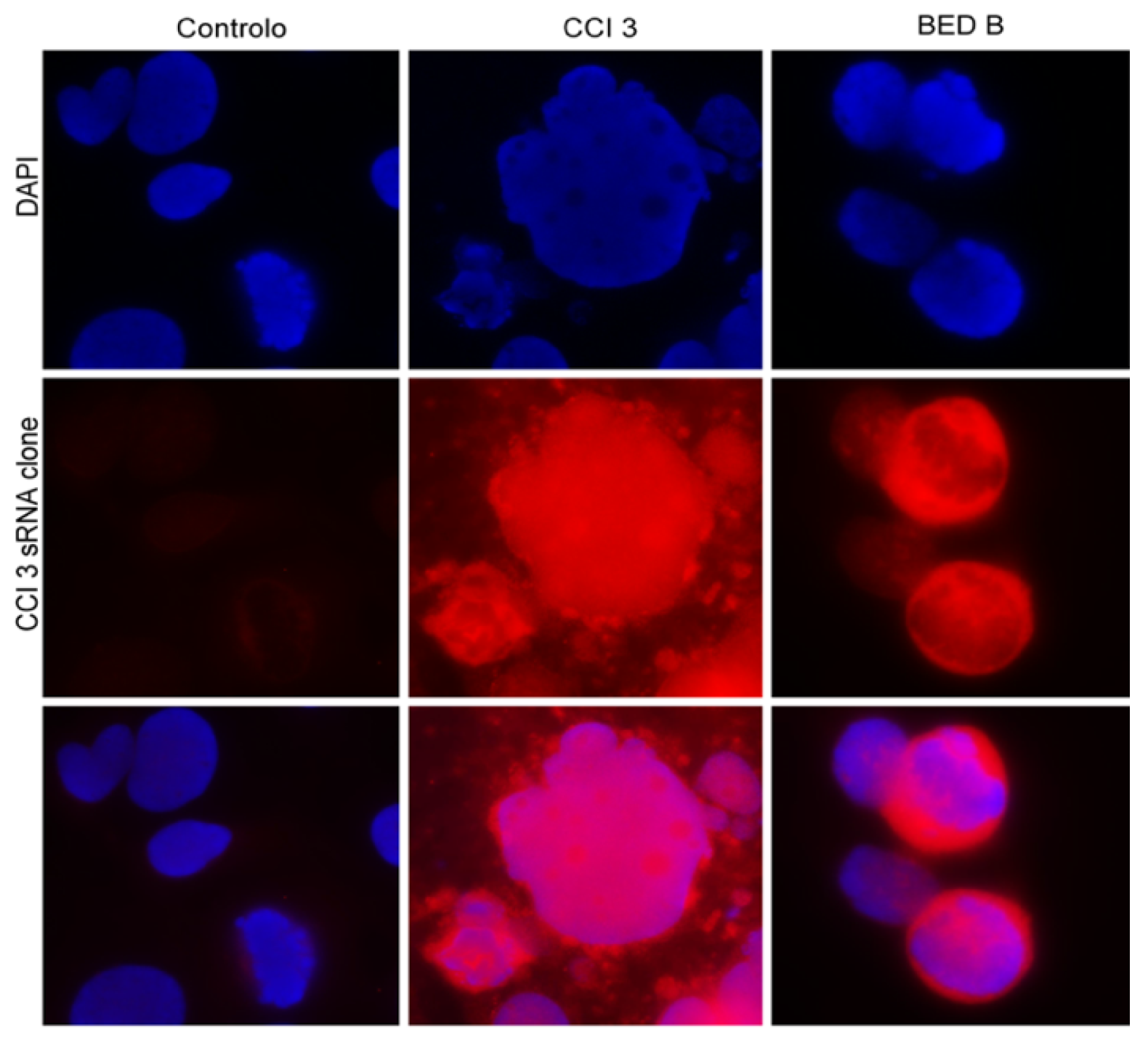
In order to understand whether the effect of CCI3 and BEDB was sequence-dependent and if the sequence isolated in these fractions was similar, we carried out an RNA-FISH in Caco-2 cells that were untreated and treated with CCI3 and BEDB for 96 h, using a clone isolated from CCI3 as a probe. In
Figure 3, it is possible to see the absence of the hybridization signal in the untreated cells. On the other hand, in Caco-2 cells treated with the CCI3 and the BEDB fractions, the hybridization of the CCI3 sRNA probe (in red) is clearly evident, both in the nucleus and in the cytoplasm of the cells. These data indicate that the sRNAs present in the CCI3 and BEDB fractions are similar in their sequence (80% stringency applied in the post-hybridization baths), suggesting that these fractions are enriched in the same sRNA sequence. However, further functional studies will be needed in order to understand their target in tumor cells and the mechanism behind their anti-tumor capacity.



 ,
, {kind=link}
{kind=link}
{kind=link}


