
Phytochemicals from Cocoa Shell Protect Mitochondrial Function and Alleviate Oxidative Stress in Hepatocytes via Regulation of ERK and PI3K-AKT Pathways †

 , , and
, , and 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experiments
2.1. Materials
2.2. Aqueous Extraction of Phytochemicals from Cocoa Shell and UPLC-MS/MS Characterization
2.3. Cell Culture
2.4. Experimental Design
2.5. Cell Viability
2.6. Evaluation of the Effect of Phytochemicals from Cocoa Shell Intracellular ROS, Mitochondrial Superoxide, and Mitochondrial Membrane Potential (ΔΨm)
2.7. Evaluation of the Effect of Phytochemicals from Cocoa Shell on Hepatic Mitochondrial Content, Oxygen Consumption, and ATP Production
2.8. Evaluation of the Effect of Phytochemicals from Cocoa Shell on the Phosphorylation Pattern of Liver Cellular Key Signaling Pathways
2.9. Bioinformatic Analysis
2.10. Statistical Analysis
3. Results
3.1. Cocoa Shell Was Mainly Composed of Theobromine, Protocatechuic Acid, and Flavan-3-Ols
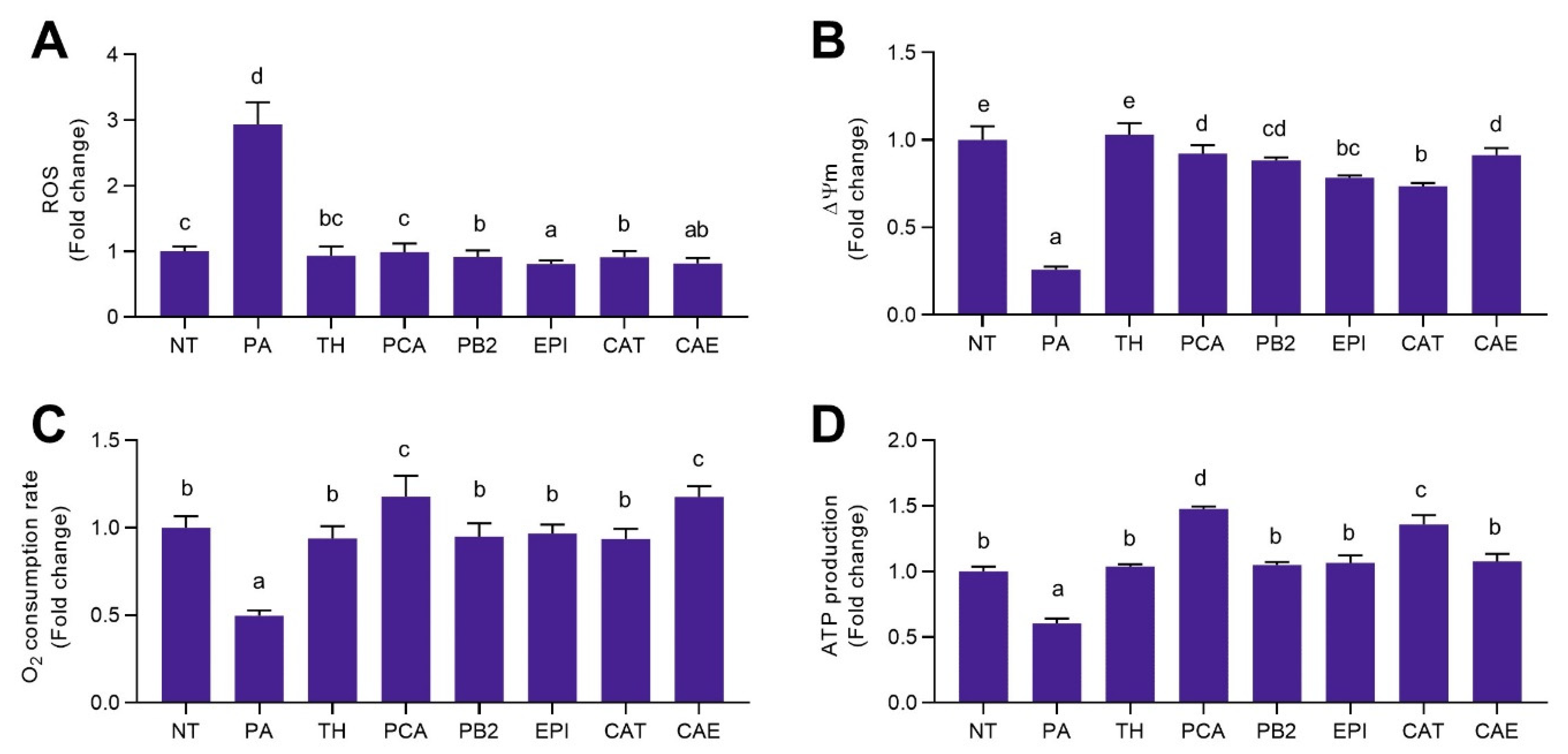
3.2. Phytochemicals Present in Cocoa Shell Attenuated Oxidative Stress in Palmitic Acid Treated Hepatocytes
3.3. Phytochemicals from Cocoa Shell Counteracted Palmitic Acid Effects on Mitochondrial Function in Hepatocytes
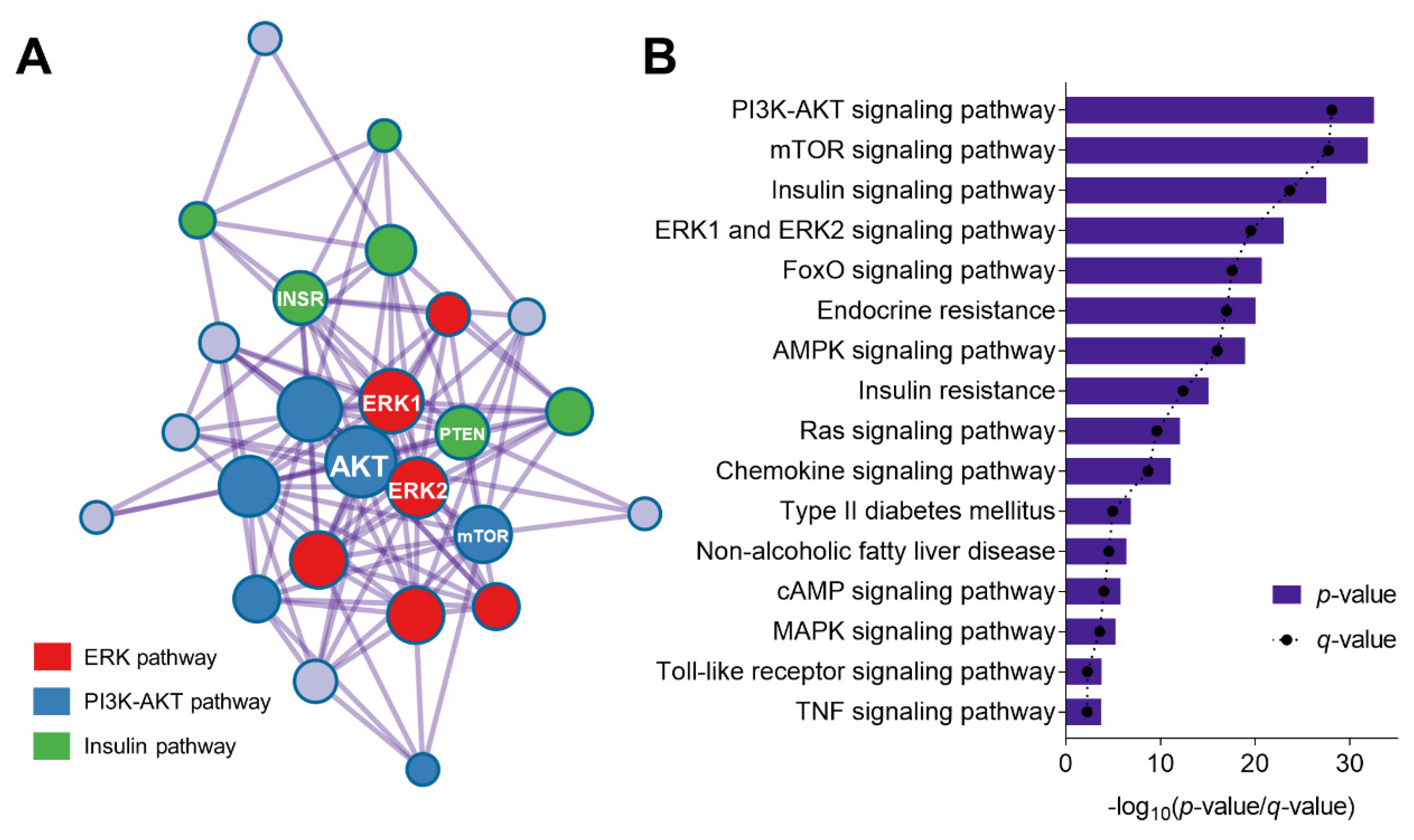
3.4. Phytochemicals from Cocoa Shell Protect Mitochondrial Function and Alleviate Oxidative Stress in Hepatocytes via Regulation of ERK and PI3K-AKT Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| NT | Nontreated cells |
| PA | Palmitic acid |
| PB2 | Procyanidin B2 |
| PBS | Phosphate-buffered saline |
| PCA | Protocatechuic acid |
| ROS | Reactive oxygen species |
| TH | Theobromine |
| CAE | Cocoa shell aqueous extract |
| CAT | Catechin |
| EDTA | Ethylenediaminetetraacetic acid |
| EPI | Epicatechin |
| FBS | Fetal bovine serum |
| MEM | Minimum essential medium |
| NAFLD | Nonalcoholic fatty liver disease |
References
- World Health Organization Noncommunicable Diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/noncommunicable-diseases (accessed on 20 May 2020).
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Kader, S.M.; El-Den Ashmawy, E.M.S. Non-alcoholic fatty liver disease: The diagnosis and management. World J. Hepatol. 2015, 7, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef]
- Schulze, M.B.; Martínez-González, M.A.; Fung, T.T.; Lichtenstein, A.H.; Forouhi, N.G. Food based dietary patterns and chronic disease prevention. BMJ 2018, 361, k2396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panak Balentić, J.; Ačkar, Đ.; Jokić, S.; Jozinović, A.; Babić, J.; Miličević, B.; Šubarić, D.; Pavlović, N. Cocoa Shell: A By-Product with Great Potential for Wide Application. Molecules 2018, 23, 1404. [Google Scholar] [CrossRef] [Green Version]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martín-Cabrejas, M.A.; de Mejia, E.G. Cocoa Shell Aqueous Phenolic Extract Preserves Mitochondrial Function and Insulin Sensitivity by Attenuating Inflammation between Macrophages and Adipocytes In Vitro. Mol. Nutr. Food Res. 2019, 63, 1801413. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martín-Cabrejas, M.A.; de Mejia, E.G. Relationship of the phytochemicals from coffee and cocoa by-products with their potential to modulate biomarkers of metabolic syndrome in vitro. Antioxidants 2019, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Rebollo-Hernanz, M.; Cañas, S.; Taladrid, D.; Bartolomé, B.; Aguilera, Y.; Martin-Cabrejas, M.A. Extraction of phenolic compounds from cocoa shell: Modeling using response surface methodology and artificial neural networks. Sep. Purif. Technol. 2021, 270, 118779. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martín-Cabrejas, M.A.; Gonzalez de Mejia, E. Phenolic compounds from coffee by-products modulate adipogenesis-related inflammation, mitochondrial dysfunction, and insulin resistance in adipocytes, via insulin/PI3K/AKT signaling pathways. Food Chem. Toxicol. 2019, 132, 110672. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Reilly, S.M.; Saltiel, A.R. Adapting to obesity with adipose tissue inflammation. Nat. Rev. Endocrinol. 2017, 13, 633–643. [Google Scholar] [CrossRef]
- Godoy-Matos, A.F.; Silva Júnior, W.S.; Valerio, C.M. NAFLD as a continuum: From obesity to metabolic syndrome and diabetes. Diabetol. Metab. Syndr. 2020, 12, 60. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Cañas, S.; Aguilera, Y.; Benitez, V.; Gila-Díaz, A.; Rodriguez-Rodriguez, P.; Cobeta, I.M.; de Pablo, A.L.L.; Gonzalez, M.C.; Arribas, S.M. Validation of Cocoa Shell as a Novel Antioxidant Dietary Fiber Food Ingredient: Nutritional Value, Functional Properties, and Safety. Curr. Dev. Nutr. 2020, 4, 773. [Google Scholar] [CrossRef]
- Nassir, F.; Ibdah, J.A. Role of mitochondria in nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2015, 15, 8713–8742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begriche, K.; Igoudjil, A.; Pessayre, D.; Fromenty, B. Mitochondrial dysfunction in NASH: Causes, consequences and possible means to prevent it. Mitochondrion 2006, 6, 1–28. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Willis, L.; Aguilera, Y.; Martin-Cabrejas, M.A.; Gonzalez de Mejia, E. Fibroblast Growth Factor 21 Signaling Activation by Selected Bioactive Compounds from Cocoa Shell Modulated Metabolism and Mitochondrial Function in Hepatocytes. Curr. Dev. Nutr. 2020, 4, 459. [Google Scholar] [CrossRef]
- Lv, H.; Ren, H.; Wang, L.; Chen, W.; Ci, X. Lico A Enhances Nrf2-Mediated Defense Mechanisms against t-BHP-Induced Oxidative Stress and Cell Death via Akt and ERK Activation in RAW 264.7 Cells. Oxid. Med. Cell. Longev. 2015, 2015, 709845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasek-Gajda, E.; Jurkowska, H.; Jasińska, M.; Lis, G.J. Targeting the MAPK/ERK and PI3K/AKT Signaling Pathways Affects NRF2, Trx and GSH Antioxidant Systems in Leukemia Cells. Antioxidants 2020, 9, 633. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ding, H.; Liu, L.; Song, Y.; Du, X.; Feng, S.; Wang, X.; Li, X.; Wang, Z.; Li, X.; et al. Non-esterified Fatty Acid Induce Dairy Cow Hepatocytes Apoptosis via the Mitochondria-Mediated ROS-JNK/ERK Signaling Pathway. Front. Cell Dev. Biol. 2020, 8, 245. [Google Scholar] [CrossRef]
- Dou, X.; Ding, Q.; Lai, S.; Jiang, F.; Song, Q.; Zhao, X.; Fu, A.; Moustaid-Moussa, N.; Su, D.; Li, S. Salidroside alleviates lipotoxicity-induced cell death through inhibition of TLR4/MAPKs pathway, and independently of AMPK and autophagy in AML-12 mouse hepatocytes. J. Funct. Foods 2020, 65, 103691. [Google Scholar] [CrossRef]
- Kim, J.; Kim, J.; Shim, J.; Lee, C.Y.; Lee, K.W.; Lee, H.J. Cocoa Phytochemicals: Recent Advances in Molecular Mechanisms on Health. Crit. Rev. Food Sci. Nutr. 2014, 54, 1458–1472. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T.; Mcdougall, G.J.; Alegría, A.; Alminger, M.; Arrigoni, E.; Aura, A.M.; Brito, C.; Cilla, A.; El, S.N.; Karakaya, S.; et al. Mind the gap-deficits in our knowledge of aspects impacting the bioavailability of phytochemicals and their metabolites-a position paper focusing on carotenoids and polyphenols. Mol. Nutr. Food Res. 2015, 59, 1307–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebollo-Hernanz, M.; Aguilera, Y.; Martin-Cabrejas, M.A.; de Mejia, E.G. Phytochemicals from Cocoa Shell Protect Mitochondrial Function and Alleviate Oxidative Stress in Hepatocytes via Regulation of ERK and PI3K-AKT Pathways. Med. Sci. Forum 2021, 2, 25. https://doi.org/10.3390/msf2021002025
Rebollo-Hernanz M, Aguilera Y, Martin-Cabrejas MA, de Mejia EG. Phytochemicals from Cocoa Shell Protect Mitochondrial Function and Alleviate Oxidative Stress in Hepatocytes via Regulation of ERK and PI3K-AKT Pathways. Medical Sciences Forum. 2021; 2(1):25. https://doi.org/10.3390/msf2021002025
Chicago/Turabian StyleRebollo-Hernanz, Miguel, Yolanda Aguilera, Maria A. Martin-Cabrejas, and Elvira Gonzalez de Mejia. 2021. "Phytochemicals from Cocoa Shell Protect Mitochondrial Function and Alleviate Oxidative Stress in Hepatocytes via Regulation of ERK and PI3K-AKT Pathways" Medical Sciences Forum 2, no. 1: 25. https://doi.org/10.3390/msf2021002025